Embed Size (px)
Citation preview

1
DETECTION OF AMINOPEPTIDASE – P HOMOLOGOUS GENE
AND ANALYSIS OF ITS EXPRESSION IN MUSCA DOMESTICA
(HOUSEFLY)
Submitted in partial fulfillment of the M.Sc. degree course
By Aditi A. Bapat
Department of Zoology
University of Pune
Under the guidance of Dr. D. N. Deobagkar

2
Certificate This is to certify that the project entitled “Detection of
Aminopeptidase – P Homologous Gene and Analysis of its Expression in
Musca domestica (Housefly) ” submitted by Miss Aditi A. Bapat
(Department of Zoology) was carried out under my guidance, for the partial
fulfillment of the M.Sc. degree course.
Project guide: Head of Department:
Dr. D. N. Deobagkar Dr. D. N. Deobagkar
Department of Zoology Department of Zoology
University of Pune University of Pune

3
Acknowledgement I would like to extend my deepest thanks to Dr. D. N. Deobagkar for giving me this
opportunity to work as a project student. I thank him for giving me valuable guidance
during the course of my project. I would also like to thank him for allowing me the use of
the laboratory and other facilities in the department.
I would also like to thank Dr. d. D. Deobagkar for the valuable inputs given by her.
My thanks to Gauri and Prafful, whose help has been invaluable to me.
A big thanks to Dr. Shobha Bhargava for her help in setting up the Housefly cultures.
Last but not the least, I thank my classmates for being such wonderful people and making
my two years in the department truly memorable!
Aditi A. Bapat

4
Contents
1. ACKNOWLEDGEMENT
2. INTRODUCTION
3. MATERIALS AND METHODS
4. RESULTS
5. DISCUSSION
6. REFERENCES

5
INTRODUCTION About the housefly (Musca domestica) Houseflies are a major pest of man and domestic animals. They are also vectors of many diseases. They are known to carry disease-causing organisms of typhoid, cholera, bacillary dysentery, diarrhea and many types of worms. There have been a variety of approaches used to control Houseflies. Use of pesticides is a very common means. However, due to their similarity with Drosophila melanogster, these organisms also form a model system for molecular, genetic and developmental studies. The Housefly is much larger in size than the fruit fly and consequently all its structures are also larger in size and thus could be easier to study. This can be used to compare the already existing systems in Drosophila with those of the Housefly. The major morphological feature, which distinguishes flies from other insects, is their reduced hind wings, termed halteres. The halteres are small structures that function as balancing organs during flight. Thus adult flies have only one pair of functional wings, hence their scientific name - Diptera (di - two, pteron - wing). Because of the reliance on the forewings for flight, the mesothorax has become enlarged to contain the enormous flight muscles, and the pro- and metathorax are correspondingly reduced. The mouthparts of flies are also characteristically suctorial. Musca domestica is one of the most widely distributed organisms on earth and occupies a basic position in the food chain. It is in every stage of its life cycle food for some other animal or plant. The Housefly – Musca domestica The Housefly has four stages of development - the Egg, Larva, Pupa, and Adult. The eggs are deposited in considerable numbers on decaying organic matter. Manure and wet feed around barns are excellent sources of organic matter for egg laying. A single female housefly may lay as many as 180 eggs at a time. The female will lay approximately five to six times during her lifetime (about 30 days) and may produce 1,000 eggs. The young larvae emerge from the eggs within a day during normal summer temperatures. They feed for about 8 to 10 days before they are ready to pupate. The adults emerge from the pupa case in about 5 days. Thus, the cycle from eggs to adults takes about 10 to 15 days.

Life Cycle and Description
The housefly has a comand Adult. Warm summerHousefly. Egg The white eggs, about Each female fly can lay upover a three to four day per Larva The mature larva is 3 ttapering toward the headspiracles are slightly raisemaggots emerge from theimmediately feed on and grown maggots have a grea The larvae go through to a dried, cool place near Pupa The pupae are dark broformed from the last larvaas the pupa ages. The emalternately swelling and shit uses like a pneumatic ha
EGGS
MAGGOT
6
plete metamorphosis with distinct Egg, Larva or Maggot, Pupa conditions are generally optimum for the development of the
1.2 mm in length, are laid singly but pile up in small masses. to 500 eggs in several batches of about 75 to 150 eggs, each iod. Eggs hatch within 24 hours.
o 9 mm long, typical creamy whitish in color, cylindrical but . The head contains one pair of dark hooks. The posterior d and the spiracular openings are sinuous slits. The legless eggs in warm weather within eight to 20 hours, and they develop in the material where the eggs were laid. The full-sy, cream-colored appearance and are 8 to 12 mm long.
three instars. When the maggots are full-grown, they crawl up breeding material and transform to the pupal stage.
wn and 8 mm long. The pupal stage is passed in a pupal case l skin, which varies in color from yellow, red, brown, to black erging fly escapes from the pupal case through the use of an rinking sac, called the ptilinum, on the front of its head, which mmer.
PUPA
ADULT

7
Adult The Housefly is 6 to 7 mm long, with the female usually larger than the male. The eyes are reddish and the mouthparts are sponging. The thorax bears four narrow black stripes and there is a sharp upward bend in the fourth longitudinal wing vein. The abdomen is gray or yellowish with dark midline and irregular dark markings on the sides. The underside of the male is yellowish. Noting the space between the eyes, which in females is almost twice as broad as in males, the sexes can be readily separated. Adults usually live 15 to 25 days. The potential reproductive capacity of flies is tremendous. Adults suck liquids containing sweet or decaying substances. Larvae feed on moist food rich in organic matter. Although they are attracted to a variety of food materials, houseflies have mouthparts, which allow them to ingest only liquid materials. Solid materials are liquefied by means of regurgitated saliva. The flies are inactive at night. Houseflies are not agricultural pests but are vectors of human and animal diseases. Indeed they are the greatest carriers of dirt and disease known to man. They breed in a wide variety of substrates, which provide moist, fermenting or putrefying conditions. The adults have a keen sense of smell. As mentioned earlier, extending already know information from Drosophila melanogaster for investigations in Musca domestica would be relatively easier, and also useful. There are a group of small peptides known in Drosophila melanogaster to be functioning in neurotransmission. These neuropeptides are important molecules in larval as well as adult stages of the animal, since sensory and motor systems would at one time or the other depend on them. A first report of these enzymes involved in specific processing of neuropeptides has been recently published, wherein an enzyme Aminopeptidase – P has been characterized in Drosophila melanogaster. Yet another novel gene from Drosophila melanogaster has been reported from this department. This gene encodes a 122 amino acid polypeptide involved in regulation of expression of homeobox genes.the protein has a Helix – Turn – Helix structure and binds specifically to the homeobox binding DNA motif. Presence of this sequence was also checked. A novel 369 base pair ORF A 1.75 kilo base region, immediately upstream of the gene encoding the larval serum protein 1β at 21DE in Drosophila melanogaster has been found to encode homologous transcripts in Drosophila melanogaster, Anopheles stephensi, mouse and man (Deshpande, 1998). This region harbors a novel open reading frame (ORF) of 369 base pairs (EMBL nucleotide sequence database Acc. N.o. Z29571) (Samant, 1992). This 369 base pair ORF has an upstream putative TATA box, a translation start site, two tandem termination codons and a polyadenylation signal. This ORF appears to be conserved through out evolution. This 369 base pair ORF codes for a 122 amino acid protein.

8
Computer modeling predicted the protein with a helix-turn-helix (HTH) motif towards its carboxy terminal (Deobagkar & Kolaskar, unpublished data). Homeodomain proteins are a family of transcriptional factors (Travers, 1993) characterized by a 60-amino acid domain (homeodomain) that binds to certain regions of DNA. Such proteins activate genes that specify the unique properties of that segment. They fold into three α-helices, the latter two forming the HTH motif, which recognizes an AT-rich region and fits into the major groove of the soluble helix. The homeobox is highly conserved with respect to its size and sequence thorough evolution. The putative 122 amino acid protein was modeled along a 20 base pair; AT-rich target sequence of the Drosphila melanogaster engrailed protein and predicted to contact the double helix via six hydrogen bonds. The mechanism by which homeodomain proteins can regulate transcription is known to an extent and some of their target genes have been identified. Aminopeptidase-P from Drosophila melanogaster A cytosolic form of Aminopeptidase – P from Drosophila melanogaster has been identified and characterized for the first time in this department. The complete sequence of a 12.5 kilo-base pair genomic clone from Drosophila melanogaster showed the presence of a 1,839 base pair ORF, encoding a protein of 613 amino acids with a calculated molecular mass of 68.5 kDa. The deduced amino acid sequence was 48% identical and 66% similar to rat and human cytosolic aminopeptidase – P. The recombinant enzyme expressed in Escherichia coli hydrolyzed the amino terminal Xaa-Pro bond of substance P and bradykinin, revealing its functional identity. Further enzyme characterization showed the enzyme to be a manganese-dependent metallopeptidase. DAP-P is located exclusively in the cytosol and is temporally regulated during Drosophila development. In the adult fly, the protein could be detected in gut, testis and ovary, with a high level of expression in brain. Aminopeptidase – P (AP-P; X-Pro Aminopeptidase; EC 3.4.11.9) has the unique ability to cleave the N-terminal amino acid residue from peptides having proline as the penultimate amino acid residue. Biologically active peptides comprise an important and diverse class of extracellular chemical messengers that mediate a wide range of intercellular interactions. Several bioactive peptides including hormones, neuropeptides, neurotransmitters escape non-specific protease degradation by having an Xaa-Pro motif at their amino termini. Due to its cyclic nature, proline confers resistance to such peptide bonds so that aminopeptidases with broad specificity cannot act upon such peptides. There are a limited number of peptidases that act on peptide bonds involving a proline residue, such as dipeptidyl peptidase II (DPPII) and dipeptidyl peptidase IV (DPPIV) (which remove Xaa-Pro dipeptides from the N-terminus of peptides), and prolidase (which cleaves the Xaa-Pro bond only in dipeptides), or endopeptidases such as prolyl endopeptidase (which cleaves on the carbonyl side of proline residues within a protein or

9
peptide). A protease from the human immunodeficiency (AIDS) virus also hydrolyses the Xaa-Pro bond of proteins and peptides. However, none of these enzymes have been reported to hydrolyze Xaa-Pro bonds located at the N-terminus of peptides and proteins. Therefore, role of AP-P is crucial in this respect. AP-P activity is ubiquitous and has been found in a wide range of organisms including bacteria, yeast and vertebrates. Mammalian AP-Ps exist in membrane-bound and cytosolic forms, which represent two distinct gene products. The cytosolic (soluble) form of Aminopeptidase – P is found in human leukocytes and rat brain. Both forms of AP-P can hydrolyze several peptides including bradykinin (a vasodilator) and neurotransmitters such as substance P, peptide YY, and neuropeptide Y. Immunochemical localization studies of some of these peptides show their cell-specific regulation in Drosophila melanogaster.
10
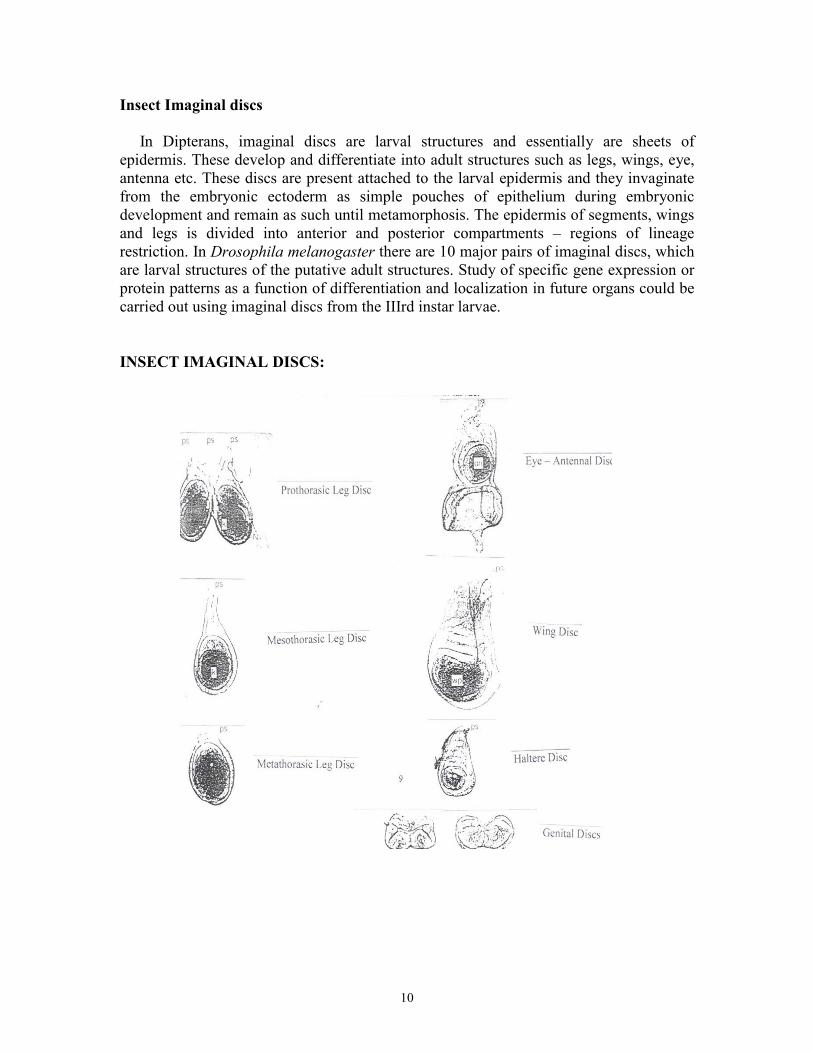
Insect Imaginal discs In Dipterans, imaginal discs are larval structures and essentially are sheets of epidermis. These develop and differentiate into adult structures such as legs, wings, eye, antenna etc. These discs are present attached to the larval epidermis and they invaginate from the embryonic ectoderm as simple pouches of epithelium during embryonic development and remain as such until metamorphosis. The epidermis of segments, wings and legs is divided into anterior and posterior compartments – regions of lineage restriction. In Drosophila melanogaster there are 10 major pairs of imaginal discs, which are larval structures of the putative adult structures. Study of specific gene expression or protein patterns as a function of differentiation and localization in future organs could be carried out using imaginal discs from the IIIrd instar larvae. INSECT IMAGINAL DISCS:

11
MATERIALS AND METHODS Breeding of Houseflies The houseflies are bread in jars with culture plates of milk and cotton from which they obtain their nutrition. The housefly has a complete metamorphosis with distinct egg, larva or maggot, pupa and adult stages. Warm summer conditions are generally optimum for the development of the housefly. Collection of Housefly embryos (Protocol as described by Pardue 1975 for collection of Drosophila embryos was modified and used)
Requirements: Bleach Dried Methanol Heptane Fixative : 4% formaldehyde n 1x PBS
Method:
The embryos were collected in baskets by washing in water. Dechorionate in baskets by adding bleach (Mediclor) for 5 min. Rinse well in water to remove all the bleach so that the smell of chlorine goes and remove all the excess water by blotting. Transfer the embryos with a brush to an eppendorf containing 500 µl of Fixative (4% formaldehyde in 1x PBS) + 500 µl heptane. Fix for 20 min by shaking vigorously. Remove the aqueous phase and add 500 µl of dried methanol and devitellinize for 5 min by shaking. Then remove the methanol and give three washes with methanol. Store at –20oC in methanol.
Genomic DNA Isolation (Ashburner 1989) Requirements:
Homogenization buffer: 60 mM KCl 15 mM NaCl 15 mM Tris-Cl (pH 7.4) (stock 1M) 2 mM EDTA (stock 0.5M)

12
T10E5: 10 mM Tris-Cl (pH 8.0) (stock 1M) 1mM EDTA (stock 0.5M)
5M NaCl Phenol is equilibrated with Tris-Cl (pH 8.0) buffer. Phenol : Chloroform (1:1) mix The alcohol is distilled and stored in clean bottles for use.
Method:
Wash the pupae in 1x PBS to remove the dirt and debris attached to it. Drain the 1x PBS and dry the pupae using tissue paper. Take 2 ml of Homogenization buffer (HB) and add 5 µl of NP40 to it. Transfer the pupae in the homogenizer with 1 ml of HB + NP40. Homogenize giving atleast 10-15 strokes. Then add the other 1 ml washing the sides of the homogenizer and transfer 0.5 ml into 4 eppendorf tubes. Centrifuge at low speed of 2000 rpm, 4oC, 2 min to get rid of the junk. Take the supernatant (with nuclei) into another tube and centrifuge at 8000 rpm, 4oC, 5 min to pellet the nuclei. Discard the supernatant and resuspend the pellet in 0.5 ml HB. Centrifuge at 8000 rpm, 4oC, 5 min. Remove the supernatant and resuspend the pellet in 200 µl of HB or T10E5. Add Proteinase K (stock 10 mg/ml) to a final concentration of 100 µg/ml followed by SDS (stock 10 %) to a final concentration of 0.1 %. Mix gently to lyse the nuclei. Keep this at 37oC for 2 hours. Then increase the volume to 400 µl with HB or T10E5. Then extract with equal volumes of Phenol. To the tubes add equal volumes of phenol. Mix and keep on ice for 5 min. then centrifuge at 10,000 rpm, 4oC, 5 min. Transfers the supernatant to another tube ant to it add equal volumes of phenol-chloroform:IAA (A mixture of phenol and chloroform:IAA is a good protein extractant). Mix and keep on ice for 5 min. and centrifuge at 10,000 rpm, 4oC, 5 min. Transfer the supernatant to another tube and add equal volumes of chloroform :IAA (Isoamyl alcohol prevents frothing of the chloroform). Mix and keep on ice for 5 min. and centrifuge at 10,000 rpm, 4oC, 5 min. Transfer the supernatant to a clean tube and add NaCl (stock 5M) to a final concentration of 150 mM. Mix and keep on ice for 5 min. Add 2 volumes of chilled distilled ethanol. A cottony white precipitate is seen. Keep at –20oC. Directly transfer the cottony white precipitate to a tube containing 70% ethanol to wash the precipitate. Centrifuge at 10,000 rpm, 4oC, 10 min. Completely remove all the 70% ethanol form the tube and dry the pellet at room temperature. Dissolve the pellet in small volume of T10E1.

13
Agarose gel electrophoresis Requirements:
50 x TAE 242g Tris base 57.1 ml Glacial Acetic Acid 100 ml 0.5M EDTA, pH 8.0 Distilled water to 1 lit
6x Loading buffer 0.25% Bromophenol Blue 40% (w/v) Sucrose in water Store at 4oC
Ethidium bromide 10 mg/ml Agarose 0.8% - 2.0%
Method:
Make 1% agarose in 1x TAE. Melt by heating (make sure to adjust the volume with water). Cast the gel. Let the gel harden. Remove the comb and place the gel in the tank with sufficient 1x TAE buffer. Mix DNA sample to be loaded with loading dye and load in the wells. Include a standard marker as well to estimate the size of DNA sample loaded. Run the gel at 80 V till the dye front is 75% across the gel. Remove the gel. Stain with 1 µg/ml Ethidium bromide for 20 min. Rinse with water to remove excess stain. Place on UV transilluminator to visualize DNA bands. Photograph the gel.
Isolation of RNA (Perbal 1988) Requirements:
Buffer I: 10 mM Tris-Cl (pH 8.5) (stock 1M) 140mM NaCl (stock 1M) 1.5mM MgCl2 (stock 2M)
Buffer II: 50mM Tris-Cl (pH 8.0) (stock 1M) 10mM NaCl (stock5M) 10mM EDTA (stock 0.5M) 0.5% SDS (stock 10%)
1x PBS: 130mM NaCl

14
7mM Na2HPO4 . 2 H2O 3mM Nah2PO4 . 2H2O Adjust pH to 7.0
5M NaCl Phenol equilibrated with DEPC treated water. Phenol : Chloroform (1:1) mix Chloroform : Isoamyl alcohol (24:1) Absolute ethanol. Autoclaved and DEPC treated tips and eppendorfs. Treat all solutions and glassware with 0.02% Diethyl pyrocarbonate (DEPC), a non-specific inhibitor of RNAse.
Method:
Wash the larvae in 1x PBS and freeze them. Take the larvae in a homogenizer and homogenize giving atleast 5-8 strokes in Buffer I on ice. Transfer the homogenate to a tube. To the tube add 10% NP40 to a final concentration of 0.1%. Vortex twice for 15 seconds each. Then centrifuge at 10,000 rpm, 4oC, 15 min to pellet the nuclei. Carefully remove the supernatant and add it to a tube containing 1/5th the volume of Buffer II. Mix gently and keep on ice for 5 min. Then add equal volumes of water saturated Phenol. Mix and keep on ice for 5 min. then centrifuge at 10,000 rpm, 4oC, 5 min. Remove supernatant and extract with equal volumes of chloroform:IAA. Mix and keep on ice for 5 min. centrifuge at 10,000 rpm, 4oC, 5 min. Remove the supernatant and add NaCl (stock 5M) to a final concentration of 200 mM. Keep on ice for 5 min. Then add 2.5 volumes of chilled distilled ethanol. A white flocculant precipitate of RNA will be seen.
Antibody staining of Imaginal discs (Pardue 1975) Requirements:
PBS-Tx-BSA: 1x PBS (stock 10x) 0.1% Trioton-X (stock 100%) 0.5% BSA
Blocking solution: 1x PBS (stock 10x) 0.1% Trioton-X (stock 100%) 3% BSA

15
Antibody dilution buffer: 1x PBS (stock 10x) 3% BSA
DAB: A stock of 10mg/ml DAB in 1x PBS is made. The working concentration used is 1mg/ml. Method:
The imaginal discs are isolated from IIIrd instar larvae. They are stored in fixative (4% formaldehyde in 1x PBS). Wash the imaginal discs in 1x PBS-Tx-BSA for 20 min x 3. Remove the last wash and add 1 ml of blocking solution. Keep on rocker for 2 hours. Then remove the blocking solution and wash the discs once with 1x PBS-Tx-BSA for 10 min. Remove this wash and add the primary antibody (appropriately diluted) to the discs and keep it at 4oC overnight. In the control tube add the equivalent amount of antibody dilution buffer. Simultaneously also keep the secondary antibody for preabsorbtion overnight at 4oC. Remove the primary antibody and store it. It can be reused. Then wash with 1x PBS-Tx-BSA for 30 min x 4. Remove the last wash and add the secondary antibody (appropriately diluted). Keep for 2 hours at room temperature. Remove the secondary antibody and wash with 1x PBS-Tx-BSA for 30 min x 4. Then add 500µl of DAB (Diaminobenzidine) + H2O2 (1µl in 1 ml). Develop for 5-10 min. Then wash with 1x PBS-Tx-BSA for 5 min x 3. Finally store the discs in 50% glycerol.
Antibody staining of embryos (Pardue 1975) Requirements:
PBS-Tx-BSA: 1x PBS (stock 10x) 0.1% Trioton-X (stock 100%) 0.5% BSA
Blocking solution: 1x PBS (stock 10x) 0.1% Trioton-X (stock 100%) 3% BSA
Antibody dilution buffer: 1x PBS (stock 10x) 3% BSA

16
DAB: A stock of 10mg/ml DAB in 1x PBS is made. The working concentration used is 0.5mg/ml. Method:
Wash the embryos twice in methanol. Add 70% methanol + 1% H2O2 to the embryos and keep for 15 min (to inactivate the internal peroxidase). Then wash the embryos in 1x PBS-Tx-BSA for 5 min x 2. Wash the embryos in 1x PBS-Tx-BSA for 20 min x 3. Remove the last wash and add 1 ml of blocking solution. Keep on rocker for 2 hours. Then remove the blocking solution and wash the discs once with 1x PBS-Tx-BSA for 10 min. Remove this wash and add the primary antibody (appropriately diluted) to the discs and keep it at 4oC overnight. In the control tube add the equivalent amount of antibody dilution buffer. Simultaneously also keep the secondary antibody for pre-absorbtion overnight at 4oC for pre-absorbtion do not treat the embryos with 70% methanol + 1% H2O2. Remove the primary antibody and store it. It can be reused. Then wash with 1x PBS-Tx-BSA for 30 min x 4. Remove the last wash and add the secondary antibody (appropriately diluted). Keep for 2 hours at room temperature. Remove the secondary antibody and wash with 1x PBS-Tx-BSA for 30 min x 4. Then add 500µl of DAB (Diaminobenzidine) + H2O2 (1µl in 1 ml). Develop for 5-10 min. Then wash with 1x PBS-Tx-BSA for 5 min x 3. Finally store the discs in 50% glycerol.
Polymerase Chain Reaction (PCR) (Ausubel etal, 1987) Requirements:
Genomic DNA 10x Taq buffer 50 mM MgCl2 2mM dNTP mix Taq polymerase Distilled water Primers:
369 base pair ORF (primer I) (AT GGC ATC TAA TTT ACC TAC) 369 base pair ORF (primer II) (TAA ACG ATC ATA GTT TCG CG)

17
ATG_________________________________________TAA Aminopeptidase (DAP) I (GGA ATT CAT GAA GAG GAG CAC CAC CCA A) Aminopeptidase (DAP) II (GGA ATT CTT AAA TGG GCT GAA CTT CCT T) Aminopeptidase (DAP) III (GGA ATT CTC GGC GTT ACC TCC TGA ATT C) ATG_________________________________________TAA Dap I DapII Dap III 100 ng of Genomic DNA was used to amplify target sequences using specific primers. The components of the PCR reaction are as follows:
Components Volumes Final Concentrations Template 1 µl X
Primer (sense) 1.5 µl 0.5 µM Primer (anti-sense) 1.5 µl 0.5 µM
10x Taq buffer 3 µl 1 x 50mM MgCl2 1 µl 1.66 µM
2mM dNTP mix 1.5 µl 0.1 µM Taq polymerase 1 µl Distilled water 19.5 µl
Total 30 µl PCR Conditions:
1 cycle 35 cycles 1 cycle (final extension) Soak
Denature 94o C (2 min) 94o C (30 sec) 72o C (15 min) 4o C Anneal 50o C (1 min) 50o C (30 sec) Extend 72o C (2.5 min) 72o C (2.5 min)
Two different sets of primers were used. One was for the 369 bp ORF and the other was for DAP (I/II and I/III)

18
Gradient PCR: Along with the regular PCR a Gradient PCR was also set up. The conditions were the same as above, only the annealing temperatures were changed to 51oC, 52oC, 53oC for DAP primers I and III and to 52oC, 53oC, 54oC for the 369 base pair ORF primers. Three tubes were used, one at each temperature. The products were analyzed on a 1.5% agarose gel. Reverse-Transcriptase Polymerase Chain Reaction (RT-PCR) (Protocol as recommended by the manufacturer, One-Step RT-PCR system, Life Technologies, USA) Requirements:
Whole RNA from IIIrd Instar larvae of Housefly 2x Taq buffer RT and Taq polymerase mix Distilled water Primers
369 bp ORF (primer I) (AT GGC ATC TAA TTT ACC TAC) 369 bp ORF (primer II) (TAA ACG ATC ATA GTT TCG CG) Aminopeptidase (DAP) I (GGA ATT CAT GAA GAG GAG CAC CAC CCA A) Aminopeptidase (DAP) II (GGA ATT CTT AAA TGG GCT GAA CTT CCT T) Aminopeptidase (DAP) III (GGA ATT CTC GGC GTT ACC TCC TGA ATT C) 1.6 µg of total RNA from IIIrd instar larva of Housefly was used as template for amplifying DNA sequences corresponding to the Aminopeptidase and the 369 base pair ORF. The RT reaction was set up as given below:
Components Volumes Final Concentrations Template 1 µl X
Primer (sense) 1.5 µl 0.5 µM Primer (anti-sense) 1.5 µl 0.5 µM
2x Buffer 3 µl 1 x RT and Taq polymerase
mix 1 µl
Distilled water 19.5 µl Total 30 µl

19
PCR Conditions:
1 cycle 35 cycle 1 cycle (final extension) Soak
CDNA synthesis Denature 94o C (30 sec) 72o C (15 min) 4o C
46o C (30 min) Anneal 50o C (30 sec)
Extend 72o C (2.5 min)
Two different sets of primers were used. One was for the 369 base pair ORF and the other was for DAP (I/II and I/III) Gradient PCR: Along with the regular PCR a Gradient PCR was also set up. The conditions were the same as above, only the annealing temperatures were changed to 46oC 47oC 48oC. Three tubes were used, one at each temperature. The products were analyzed on a 1.5% agarose gel.

20
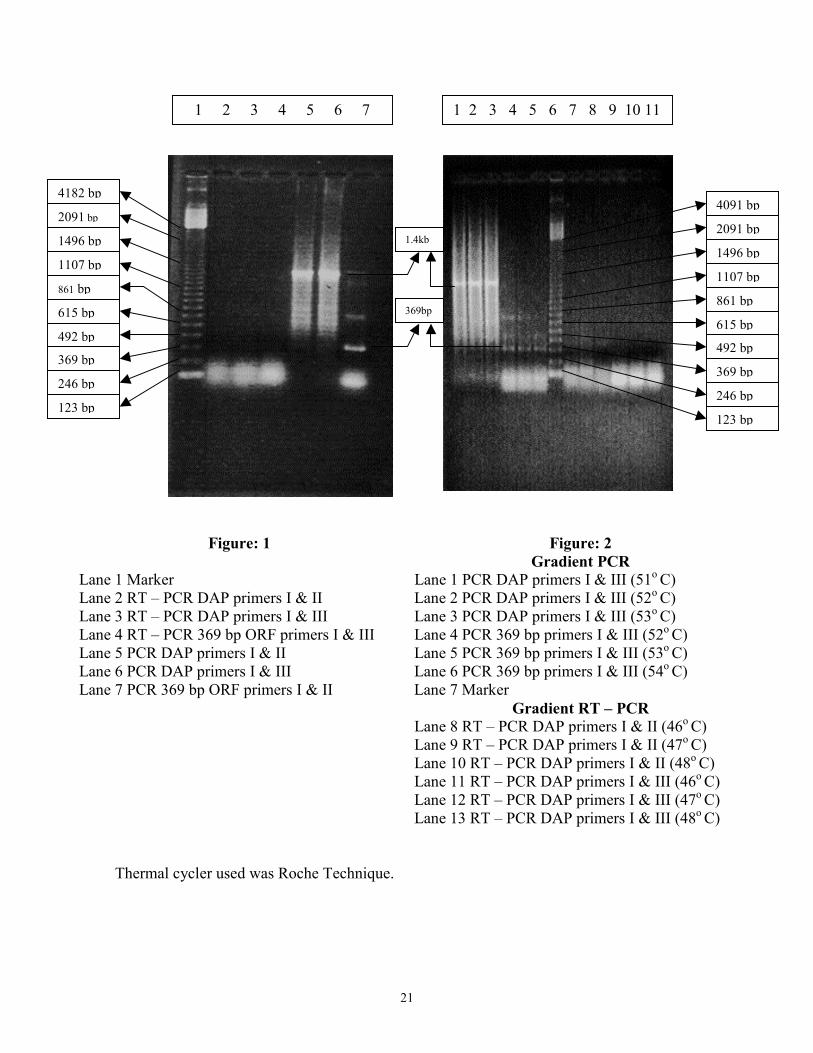
RESULTS Polymerase Chain Reaction Whole genomic DNA from housefly pupae was PCR amplified using primers for the 369 base pair ORF and homologous primer based amplification was also seen with Aminopeptidase – P. The 122 amino acid polypeptide encoded by the 369 base pair ORF has been shown to be present in a variety of organisms including insects such as Drosophila melanogaster, mosquitoes to mammals including mouse and human. Its conserved feature relates to its important function in differentiation of the Peripheral Nervous System. This gene has been localized on chromosome 1 in humans denoting that the amplified DNA is of similar size to that in other organisms, further confirming its presence in Musca domestica. This could therefore be extended in studies relating to the developmental and tissue specific expression in Musca domestica as well. Primers used in the PCR reaction showed selective amplification of a stretch of DNA 369 base pair long, confirming a similar coding region to be present. Apart form the 369 base pair fragment, two additional bands were also seen, suggesting that there could be a larger genomic sequence where the 122 amino acid protein may be a part of a larger polypeptide. Of course experimental artifact could not be ruled out at this stage. Using homologous primers for Aminopeptidase – P a single band of size 1.4 kb was observed to be amplified. But a smear was seen which might be due to non-specific amplification. Therefore a gradient PCR was also run where the annealing temperatures were changed such that the stringency was increased. Amplification was seen but the smear still remained and multiple bands were still seen with primers for the 369 base pair ORF also. It may therefore be necessary to standardize the conditions more specifically. Total RNA from IIIrd instar larvae was used in RT – PCR using the same primers for 369 base pair ORF and Aminopeptidase – P. But no amplification was seen with these primers, also in gradient RT – PCR. These observations however indicate the presence of related sequences in Musca domestica.
Lane 1 Marker Lane 2 RT – PCLane 3 RT – PCLane 4 RT – PCLane 5 PCR DALane 6 PCR DALane 7 PCR 36
Thermal c
p
p
p
p
p
p
p
p
1 2 3 4 5 6 7
21
Figure: 1
LaneR DAP primers I & II LaneR DAP primers I & III LaneR 369 bp ORF primers I & III LaneP primers I & II LaneP primers I & III Lane
9 bp ORF primers I & II Lane
LaneLaneLaneLaneLaneLane
ycler used was Roche Technique.
1.4kb
369bp
1 2 3 4 5 6 7 8 9 10 11
4182 b
p2091 bp
1496 b
1107 b
pp
615 bp
492 b861 bp
p
369 bp
246 bp
123 bFigure: 2 Gradient PCR
1 PCR DAP primers I & III (51o C) 2 PCR DAP primers I & III (52o C) 3 PCR DAP primers I & III (53o C) 4 PCR 369 bp primers I & III (52o C) 5 PCR 369 bp primers I & III (53o C) 6 PCR 369 bp primers I & III (54o C) 7 Marker
Gradient RT – PCR 8 RT – PCR DAP primers I & II (46o C) 9 RT – PCR DAP primers I & II (47o C) 10 RT – PCR DAP primers I & II (48o C 11 RT – PCR DAP primers I & III (46o C 12 RT – PCR DAP primers I & III (47o C 13 RT – PCR DAP primers I & III (48o C
4091 b
2091 bp
1496 bp
1107 b
861 b
615 b
492 b
369 b
246 b
p
123 b) ) ) )

22
In situ localization of DAP in embryos of Housefly The embryos were staged using Drosophila melanogaster embryos as a reference. Staining with the anti – DAP antibodies was seen in the early stages of development. But no definitive staining pattern was seen. Thus no conclusive interpretations could be drawn based on these results.
Fig 3: Control embryo. Fig 4: Experimental embryo.
Fig 5: Control embryo. Fig 6: Experimental embryo.

23
Fig 7: Control embryo Fig 8: Experimental embryo
24
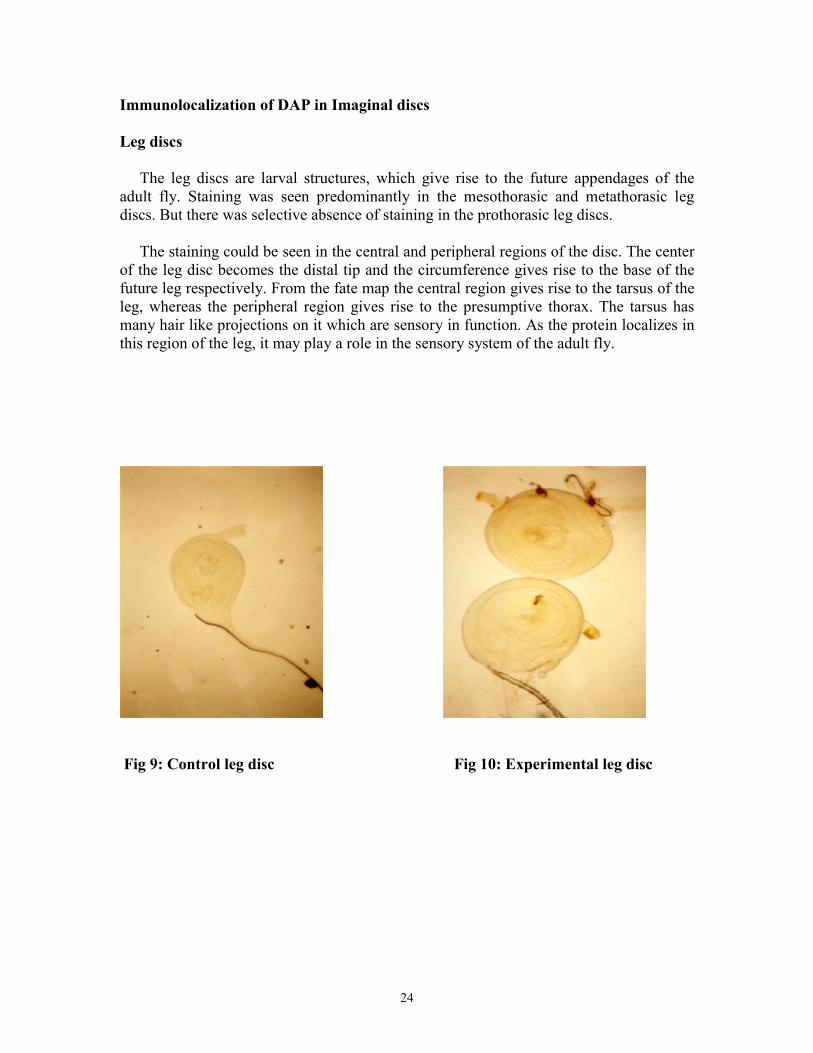
Immunolocalization of DAP in Imaginal discs Leg discs The leg discs are larval structures, which give rise to the future appendages of the adult fly. Staining was seen predominantly in the mesothorasic and metathorasic leg discs. But there was selective absence of staining in the prothorasic leg discs. The staining could be seen in the central and peripheral regions of the disc. The center of the leg disc becomes the distal tip and the circumference gives rise to the base of the future leg respectively. From the fate map the central region gives rise to the tarsus of the leg, whereas the peripheral region gives rise to the presumptive thorax. The tarsus has many hair like projections on it which are sensory in function. As the protein localizes in this region of the leg, it may play a role in the sensory system of the adult fly.
Fig 9: Control leg disc Fig 10: Experimental leg disc

25
Fig 11: Experimental leg disc Fig 12: Experimental leg disc
Fig 13: Experimental leg disc Fate Map of Leg Disc

26
Wing disc The wing disc is the larval structure, which gives rise to the future wings of the adult fly. From the fate map of the wing disc, the staining could be observed in the dorsal region of the presumptive thorax and in the region of the wing attachment i.e. the ventral hinge region. Thus the activity of this enzyme could indicate the presence of substrate molecules in this region, which need to be processed further. These proteins may have a role to play in the signal - transduction pathway during the differentiation of the imaginal discs into adult structures.
Fig 14: Control wing disc Fig 15: Experimental wing disc
Fig 16: Experimental wing disc Fate Map of Wing Disc
27
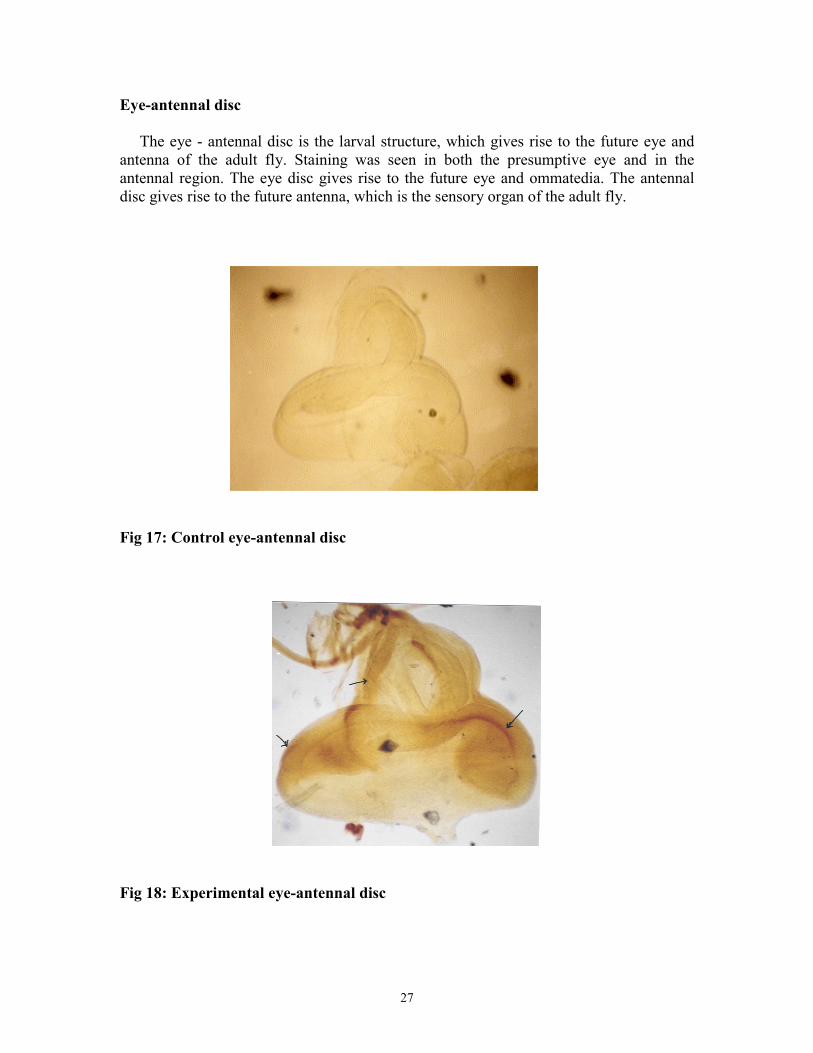
Eye-antennal disc The eye - antennal disc is the larval structure, which gives rise to the future eye and antenna of the adult fly. Staining was seen in both the presumptive eye and in the antennal region. The eye disc gives rise to the future eye and ommatedia. The antennal disc gives rise to the future antenna, which is the sensory organ of the adult fly.
Fig 17: Control eye-antennal disc
Fig 18: Experimental eye-antennal disc

28
Immunological staining was also seen in the larval brain. The staining was observed in the olfactory region of the brain. The stained regions could play a role in the sensory system of the larva wherein this enzyme could play a role in signal transmission. Presence of DAP like enzyme was detected in sub-oesophageal ganglion of the brain. This is the site where important neuropeptides have been previously reported. (Data not shown.)

29
DISCUSSION Houseflies are common pests of man. But they are also very similar to the Dipteran, Drosophila melanogster, which is a useful model organism of study. The Housefly is much larger in size than the fruit fly and consequently all its structures are also larger in size and is thus easier to study. This property of the Housefly can be used to compare the already existing systems in Drosophila melanogaster with those of the Housefly. To add to our knowledge of molecular biology certain genomic sequences and their functionalities can be compared in Musca domestica. Thus homologous sequences known in Drosophila melanogster could be easily identified and could be of interest. In this study the presence of the Aminopeptidase – P was checked. Using the Polymerase Chain Reaction, homologous primer based amplification with Aminopeptidase – P showed a single band of 1.4 kb present in the genome of the Housefly, suggesting that homologous sequences may be present in the genome. A similar amplification was done using primers of the 369 base pair ORF. This also showed a single prominent band, confirming a similar coding region to be present. Apart form this two additional bands were also seen, suggesting that there could be a larger genomic sequence where the 122 amino acid protein may be a part of a larger polypeptide. Of course experimental artifact can not be ruled out at this stage. A Revers Transcriptase – PCR was also run to detect any transcript in the total RNA of IIIrd instar larvae. But no amplification was seen and thus transcript could not be detected. Antibodies specific to Aminopeptidase – P were used to localize the protein in the embryos of the Housefly and imaginal discs from its IIIrd instar larvae. The staining was not conclusive in the embryos due to which solid conclusions could not be drawn as to its significance. Due to the time constraint, conditions specific for staining of the embryos could not be standardized. Of the imaginal discs stained, staining was observed in the leg discs, wing disc and eye – antennal disc of the IIIrd instar larva. Staining was seen in the distal region and the proximal region of the leg disc. The distal region gives rise to the tarsal segments of the leg. As they are distal most they are suggested to have a sensory function. Thus this protein may be localized in the sensory area of the nervous system. The circumference of the leg disc gives rise to the presumptive thorax in the adult fly. Staining in this region also indicates that it plays a role in the sensory relay system. Staining in the wing disc was observed in the dorsal region of the presumptive thorax and in the region of the wing attachment i.e. the ventral hinge. The presence of this protein the hinge region of the wing may suggest a possible role for this protein in the signal - transduction pathway. The protein is an enzyme and its activity could indicate that the peptides and polypeptides present in this region require further processing, leading to their activation or deactivation. These in turn could be responsible for the switching on or off of the signal - transduction pathway. This pathway may control the

30
development and differentiation of the imaginal discs into adult structures as the larva metamorphoses into the adult. In addition to staining in the imaginal discs and embryos, some staining was also seen in the larval brain. The regions of the brain stained were the olfactory area and the sub – oesophageal ganglion. These regions could play a role in the sensory system of the larva. Presence of DAP - like protein in adult Housefly brain has already been reported from this lab (unpublished results, S Bhargava etal). This enzyme therefore, seems to have an important function in the adult brain.

31
REFERENCES Amano K. (1985) Breeding of the House Fly, Musca domestica, (Diptera; Muscidae) in Fresh Dung of Cattle Fed on Pasture Grass. App. Entomol. Zool. 20:43-150. Ashburner M. (1989) Drosophila: A laboratory manual. Cold Spring Harbour Laboratory. Ausubel F. M., Brent R., Kingston R. E., Moore D. D., Seidman J. G., Smith J. A., Struhl K. (1987) Current protocols in Molecular Biology, Greene Publishing Associates and Wiley Interscience, New York. Campos – Ortega J. A., and Hertenstein V. (1985) The embryonic development of Drosophila melanogaster. Springer – Verlag, Berlin. Deobagkar D. N., Bhargava A., Kulkarni-Kale U. D., Kolaskar A. S., Samant S. A., (1995). Europ. Dros. Res. Conf. 14, p301. Deshpande M. (1998) Expression and characterization of a novel 369 bp open reading frame from Drosophila melanogaster. Ph. D Thesis. Hewitt C. G. (1914) The HouseFly, its Structure, Habits, Development, Relation to Disease Control. University Press, Cambridge England. Kulkarni G. V. and Deobagkar D. D. (2002) A cytosolic form of Aminopeptidase – P from Drosophila melanogaster : Molecular cloning and characterization. J Biochem (Tokyo) 131 (3), 445-52. Lynsk T. J. (1993) Adult Resting and Larval Development Sites of Stable Flies and House Flies (Diptera: Muscidae) on Dairies in Alberta. J. Econ. Entomol. 86: 1746-1753. Pardue M. L., Gall J. G. (1975) Nucleic acid hybridization to the DNA of cytological preparations : Methods in Cell Biology. (D.M. Prescott ed), Vol. X, 1-7.





























