Embed Size (px)
Citation preview

Research in Veterinary Science 93 (2012) 975–982
Contents lists available at SciVerse ScienceDirect
Research in Veterinary Science
journal homepage: www.elsevier .com/locate / rvsc
Defects in small intestinal epithelial barrier function and morphology associatedwith peri-weaning failure to thrive syndrome (PFTS) in swine
Adam J. Moeser a,⇑, Luke B. Borst a, Beth L. Overman a, Jeremy S. Pittman b
a Department of Population Health and Pathobiology, North Carolina State University, College of Veterinary Medicine, Raleigh, NC 27606, United Statesb Murphy-Brown, LLC. Waverly, VA 23890, United States
a r t i c l e i n f o
Article history:Received 26 January 2011Accepted 3 January 2012
Keywords:PigsWeaningIntestineBarrier functionFailure-to-thrive
0034-5288/$ - see front matter � 2012 Elsevier Ltd. Adoi:10.1016/j.rvsc.2012.01.003
⇑ Corresponding author. Tel.: +1 919 513 0711; faxE-mail address: [email protected] (A.J. Moe
a b s t r a c t
The objective of this study was to investigate intestinal function and morphology associated with peri-weaning failure to thrive syndrome (PFTS) in swine. Jejunum and distal ileum from control and pigsexhibiting PFTS was harvested at weaning, 4 and 11 days post-weaning (PW) for intestinal barrier func-tion studies and histological analyses (n = 6 pigs per group). Marked disturbances in intestinal barrierfunction was observed in PFTS pigs, compared with controls, indicated by lower (p < 0.05) TER andincreased (p < 0.01) permeability to FITC dextran (4 kDa). Intestines from weaned pigs, subjected to a4-day fast, exhibited minor disturbances in intestinal barrier function. Villus atrophy and crypt hyperpla-sia were observed in the PFTS intestine compared with control and fasted pigs. These data demonstratethat PFTS is associated with profound disturbances in intestinal epithelial barrier function and alterationsin mucosal and epithelial morphology in which anorexia is not the sole factor.
� 2012 Elsevier Ltd. All rights reserved.
1. Introduction
The weaning transition is one of the most stressful events ayoung pig encounters in swine production; therefore significantreductions in feed intake and performance (Boudry et al., 2002,2004), compromised intestinal health (Moeser et al., 2007b), andincreased susceptibility to disease (Madec et al., 2000) are com-monly observed. Several studies have shown that weaning has adeleterious impact on intestinal barrier function in the pig result-ing in increased intestinal permeability and inflammation, contrib-uting to impaired performance and disease resistance (Moeseret al., 2007a,b; Smith et al., 2010; Spreeuwenberg et al., 2001).Despite this decline in intestinal health and performance, mostweaned pigs will gradually recover over the first week of post-weaning (PW) as they adapt to environmental and nutritional-re-lated weaning stressors; however, there is a subset of pigs that failto make the weaning transition. These healthy and robust pigs atweaning exhibit depressed to no feed consumption PW and theirbody condition rapidly declines until many are euthanized for hu-mane reasons. This condition has become increasingly recognizedover the past years and has been initially characterized by severalgroups in Canada and the United States (Dufresne et al., 2008;Harding, 2009, 2010) and was previously referred to as post-wean-ing wasting/catabolic syndrome (PWCS); however, its name hasbeen recently changed to peri-weaning failure to thrive syndrome
ll rights reserved.
: +1 919 515 3044.ser).
(PFTS). To date, there are no peer-reviewed scientific publicationson PFTS; however, several field reports indicate that diagnosticinvestigations with PFTS pigs failed to provide any conclusive asso-ciations with major post-weaning swine pathogens (e.g., PRRS,PCV2, Escherichia coli, etc.). Overall, this suggests the possibilitythat PFTS may be a result of non-infectious causes; (Harding,2010; Harding and Gauvreau, 2010; Pittman and Rovira, 2011a,b)however, this does not rule out potential unknown infectiousagents at this time that may be contributing to PFTS.
Field reports indicate that multiple strategies (management,therapeutic, and nutritional) to control PFTS have been unreward-ing and frustrating; however there have been no well-controlled,peer-reviewed experiments investigating the role of these strate-gies on PFTS. Best management practices appear to help, but donot fully mitigate the effects. Field reports indicate that environ-mental management (i.e., ventilation, temperature, water quality,stray voltage) of some cases has not been beneficial. Some farmsare more affected than others, but as a system and as an estimateof the industry, the overall prevalence may be around 3% of totalpigs affected with half of that percentage resulting in depressedgrowth and the other half resulting in eventual death. In specificproduction flows the impact may be as high as 20%.
PFTS appears to affect pigs independent of age or size at wean-ing, as increasing wean age fails to correct the problem and the factthat many of the affected pigs have body frame/skeletal structuresimilar to the largest pigs in the group. PFTS is thought to be moreprevalent in white breeds compared with dark breeds such asDurocs but this has not been conclusively determined. In addition

976 A.J. Moeser et al. / Research in Veterinary Science 93 (2012) 975–982
to the mortality levels seen with PFTS, it is evident that nurserieswith moderate to severe PFTS show a decreased growth rate in sur-viving group-mates, both clinically affected and apparently healthyanimals. It is likely that whatever instigating ‘‘event’’ is occurring,even in mild cases, is having an impact of growth rate during thenursery phase and potentially into finishing. Despite extensiveefforts to treat and manage PFTS, results have been unrewardingindicating that there are unknown pathophysiologic processesplaying a major role. The objective of the present study was gaina more fundamental understanding of PFTS pathophysiology byevaluating intestinal function and morphology in pigs exhibitingPFTS.
2. Materials and methods
2.1. Animals
This study was conducted with pigs from a farm experiencingPFTS for at least 4 years. The farm was a 1000 all gilt one-site far-row-to-finish parity segregation farm with internal replacement ofbreeding stock. Creep feed was not provided to piglets on this farm.Nursery mortality averaged 6–15% during the period of this study.Previous attempts to control and treat pigs exhibiting PFTS in-cluded management changes, improved biosecurity, sanitationchanges, supportive nutrition, antibiotics, and vaccines; however,this did not impact morbidity and mortality. Historical routinediagnostics on this farm have not revealed any consistent patho-gens among PFTS-affected pigs. Previous diagnostic tests includedculture (aerobic, anaerobic, Brachyspira spp., Salmonella enrich-ment), Immunohistochemisty (Leptospira, rotavirus), PCR (PRRS,PCV2, SIV, Rotavirus Type A/B/C, TGE, CSF, HEV, PCMV, Haemophi-lus parasuis (Hps), Mycoplasma hyopneumoniae (Mhp), Mycoplasmahyorhinis (Mhr), Mycoplasma hyosynoviae (Mhs), Lawsonia intracell-ularis, Leptospira spp.), ELISA ( Clostridium difficile toxin, etc.), HI(HEV, etc.), virus isolation (4 cell lines), toxic mineral analysis, boneash, electron microscopy and fecal floatation.
2.2. Experimental groups
A total of 36 weaned pigs (Large White-Landrace cross) derivedfrom the same weaning group were used in this study and theywere allocated to the following experimental groups: (1) Un-weaned pigs (day 0 weaning), (2) control pigs at 4 days PW, (3) PFTSpigs at 4 days PW, (4) Fasted pigs at 4 days PW, (5) control pigs at11 days PW, and (6) PFTS pigs at 11 days PW. The Fasted groupwas not allowed to consume feed for 4 days PW but given ad libitumaccess to water and served as a control for potential confoundingeffects of anorexia observed with PFTS. All fasted pigs were moni-tored twice daily and besides body weight loss, there were no othersigns of debilitating health. Selection of ‘‘PFTS pigs’’ was based onphysical appearance relative to the same-age pigs weaned on thesame day. Clinical selection criteria was based on gauntness in flankregion (due to loss of gut fill), visible or palpation of spine, visibleshoulder blades and hip prominences and looseness of skin (dehy-dration). Clinically normal (control) pigs were identified as lackingthe above clinical signs and void of any signs of other illness, includ-ing diarrhea. The average piglet weaning ages for each experimen-tal group are presented in Table 1. Average weaning age was similarbetween the control and PFTS experimental groups (weaningage = 20.8 days (range = 17–24) for control pigs and 21.3 days(range 18–24) for PFTS pigs, respectively). Given that the selectionof pigs was based on clinical criteria of pigs within large pens at4 days PW, feed intake for each group could not be accuratelymeasured; however, weaning weight and final body weights wererecorded and growth rate was calculated.
2.2.1. Ussing chamber studiesOn Days 0, 4, and 11 PW, selected pigs were euthanized by
administering an overdose of sodium pentobarbital via intrave-nous injection. Mid-jejunum and distal ileum was harvested fromeach pig immediately after euthanasia, and the intestinal mucosawas stripped from the seromuscular layer in oxygenated (95%O2–5% CO2) Ringer solution (in mmol/l: 154 Na+, 6.3 K+, 137 Cl�,0.3 H2PO4, 1.2 Ca2+, 0.7 Mg2+, 24 HCO�3 ; pH 7.4). To avoid themounting of Peyer’s patches on the Ussing chambers, ileal loopswere opened along the anti-mesenteric border and thus measure-ments were obtained from ileal mucosa associated with the mes-enteric side of the intestine. Jejunal and ileal mucosal tissueswere mounted in 1.13 cm2 aperture Ussing chambers, as describedpreviously (Argenzio and Liacos, 1990; Moeser et al., 2007b). Tis-sues were bathed on the serosal and mucosal sides with 10 mlRinger solution. The serosal bathing solution contained 10 mM glu-cose, which was osmotically balanced on the mucosal side with10 mM mannitol. Bathing solutions were oxygenated (95% O2–5%CO2) and circulated in water-jacketed reservoirs maintained at37 �C. The spontaneous potential difference (PD) was measuredusing Ringer-agar bridges connected to calomel electrodes, andthe PD was short-circuited through Ag–AgCl electrodes using avoltage clamp that corrected for fluid resistance. Tissues weremaintained in the short-circuited state, except for brief intervalsto record the open-circuit PD. Transepithelial electrical resistance(X cm2) was calculated from the spontaneous PD and short-circuitcurrent (Isc), as previously described (Argenzio and Liacos, 1990).After a 30-min equilibration period on Ussing chambers, TER andIsc was recorded at 15-min intervals over a 1-h period and thenaveraged to derive the basal TER and Isc values for a given animal.TER and Isc values are presented as the average ± SE of n = 6 pigs/treatment group.
2.2.2. Mucosal-to-serosal fluxes of FITC dextranAfter a 30-min equilibration period on Ussing chambers, 4 kDa
FITC-dextran (FD4) was added to the serosal bathing reservoir ofUssing chambers at a concentration of 100 mg/ml. After a 15-minequilibration period, standards were taken from the serosal sideof each chamber. Samples were taken from the serosal chambersafter a 60-min period. The quantity of FD4 was established by mea-suring the fluorescence in serosal reservoir fluid samples in a fluo-rescence plate reader at 540 nm. Data was presented as the rate ofFD4 flux (lmol cm2 h) over a 60-min flux period.
2.2.3. HistologyJejunum and ileum were fixed in 10% neutral buffered formalin
and processed for paraffin-embedding and staining with hematox-ylin and eosin (H&E) for histological analysis. Histological analysiswas performed by a board-certified veterinary pathologist (L.B.)who was blinded to experimental groups. Photomicrographs wereacquired with 20� and 60� magnifications at a resolution of1360 � 1024 using imaging software (Olympus DP2-BSW version2.2) running a high resolution digital camera (Olympus DP72)equipped to a clinical light microscope (Olympus BX45). Prior toimaging, the system was calibrated at each magnification using astage micrometer. Measurements were taken using the arbitraryline tool and exported into a spreadsheet program (Excel2007,Microsoft). Villi were measured using the 20� objective. Villi cho-sen for measurement were based on the criteria that (1) the entirecrypt and villi were captured in cross section and (2) the centrallacteal was present. Villi overlying gut associated lymphoid tissuewas excluded from measurement. A minimum of three villi fromeach pig were measured. Epithelial height from the villus tip andcrypt was measured at 60� magnification without using the con-denser. Measurements were taken through the nuclear axis from

Table 1Growth performance.
Days post-weaning
0 4 11
Unweaned Control PFTS Fasted Control PFTS
Weaning weight, kg 5.5 ± 0.3 6.4 ± 0.2 5.6 ± 0.5 6.1 ± 0.4 7.4 ± 0.3 6.0 ± 0.5#
Final weight, kg 6.9 ± 0.3 5.8 ± 0.4# 5.8 ± 0.4� 8.7 ± 0.4 6.2 ± 0.5#
ADG, kg/day 0.063 ± 0.01 0.035 ± 0.03# �0.07 ± 0.01� 0.341 ± 0.6 0.050 ± 0.05#
Values represent means ± SE from n = 6 pigs/experimental group.Data were analyzed using a 2-Way ANOVA with PW time point (4 and 11 days PW) and treatment as the main factors.#,� p < 0.05 compared with controls within each PW time point.
4 days PWUW
TER
, Ω.c
m2
0
10
20
30
40
50
60
70
Cont PFTS Fast PFTSCont
11 days PW
#
4 days PW UW
TER
, Ω.c
m2
0
10
20
30
40
50
60
70
Cont PFTS Fast PFTSCont
#
11 days PW
A
B
** *
*
*
**
*† #
Fig. 1. Transepithelial electrical resistance (TER) across porcine jejunum (A) andileum (B) from control and PFTS pigs. Values represent means ± SE from n = 6 pigs/experimental group. ⁄ p < 0.05 compared with unweaned (UW) pigs, #,� p < 0.05compared with controls within each PW time point. Data were analyzed using a 2-Way ANOVA with PW time point (4 and 11 days PW) and treatment as the mainfactors.
A.J. Moeser et al. / Research in Veterinary Science 93 (2012) 975–982 977
the basement membrane to the microvillus brush border from anaverage of 10 epithelial cells.
2.3. Statistics
All data were reported as means ± SE based on the experimentalnumber (n). All data were analyzed by using a standard two-wayANOVA (Sigmastat, Jandel Scientific, San Rafael, CA). A post hoc Tu-key’s test was used to determine differences between treatmentsfollowing ANOVA. Statistical significance was set at a level ofP < 0.05. P-values between P > 0.05 and P = 01.0 were consideredtrends.
3. Results
3.1. Growth performance and body temperature
From day 0 to day 4 PW, PFTS pigs exhibited lower (p < 0.05)ADG compared with controls (0.063 kg/day vs. 0.035 kg/day forcontrol and PFTS pigs, respectively) (Table 1). Fasted pigs exhibitedweight loss (�0.07 kg/day) over the 4 day fasting period. Impairedgrowth in PFTS was more pronounced compared with controls byday 11 PW (0.341 kg/day vs. 0.05 kg/day for control and PFTS pigs,respectively). Rectal temperatures were measured in all pigs anddid not differ between experimental groups at any time point inthis study (data not shown).
3.2. Intestinal barrier function in PFTS pigs
On day 4 PW, jejunal TER, was lower (p < 0.05) in control, PFTS,and fasted pigs compared with unweaned controls; however, nodifferences were observed between experimental groups at thisPW time point (Fig. 1A). On day 11 PW, a significant reduction injejunal TER was observed in PFTS pigs, compared with controls(p < 0.01). Mucosal-to-serosal permeability of FD4 in the jejunumwas similar between treatment groups on day 4 PW; however, asignificant elevation in FD4 flux was observed in jejunum fromPFTS on day 11 PW (Fig. 2A). Ileum from PFTS pigs exhibitedmarked reductions in TER (Fig. 1B) and elevations in FD4 flux(Fig. 2B) on day 4 and day 11 PW compared with control and fastedpigs (day 4 PW). Fasting did not have a significant effect on jejunalTER or FD4 flux compared with controls. Fasting caused reductionsin TER and elevations in FD4 flux in the ileum; however, these dis-turbances in barrier function were lesser in magnitude comparedwith age-matched PFTS pigs.
3.3. Isc across PFTS intestine
Transepithelial Isc, a measure of electrogenic ion transport acrossepithelium, was measured in the jejunum and distal ileum fromexperimental pigs in this study. Jejunum and ileum from controlpigs exhibited higher (p < 0.05) Isc values compared with intestinal
tissues from unweaned pigs (Fig. 3). Jejunum from PFTS pigs exhib-ited a marked, negatively-directed Isc compared with controls onday 4 and day 11 PW. Ileum from PFTS pigs exhibited elevated Isc
values on day 4 PW (p < 0.01); however, on day 11 PW, a distinct,negative Isc was observed. Fasting did not have a significant effecton Isc in the jejunum or ileum compared with controls on day 4 PW.
3.4. Intestinal morphology in PFTS intestine
On day 4 PW, reductions (p < 0.05) in villus height in the jeju-num and ileum were observed in all experimental groups com-pared with unweaned pigs (Table 2, Fig. 4). Jejunum and ileumfrom PFTS pigs had reduced (p < 0.05) villus heights compared with

FD4
flux,
μm
ol/c
m2 /h
0.00000
0.00005
0.00010
0.00015
0.00020
0.00025
#
*
*
*
4 days PW
UW Cont PFTS Fast PFTSCont
11 days PW
FD4
flux,
μm
ol/c
m2 /h
0.0000
0.0001
0.0002
0.0003
0.0004
0.0005
*
*
A
B
* * *
#
*
#
4 days PW
UW Cont PFTS Fast PFTSCont
11 days PW
Fig. 2. Mucosal-to-serosal fluxes of FITC-dextran (4 kDa) across porcine ilealmucosa from control and PFTS pigs. A single 60-min flux period was initiated aftera 30-min tissue equilibration period on Ussing chambers. Ileum from PFTS pigs wassignificantly more permeable to FITC-dextran compared with control tissues at 4and 11-days post-weaning. Values represent means ± SE from n = 6 pigs/experi-mental group. ⁄ p < 0.05 compared with unweaned (UW) pigs, #,� p < 0.05 comparedwith controls within each PW time point. Data were analyzed using a 2-WayANOVA with PW time point (4 and 11 days PW) and treatment as the main factors.
978 A.J. Moeser et al. / Research in Veterinary Science 93 (2012) 975–982
control pigs on days 4 and 11 PW. Reduced villi heights associatedwith PFTS were more pronounced in the ileum compared with thejejunum. Fasting caused significant reductions in villus height inthe jejunum (p < 0.05); however, no differences were observed inthe ileum. Crypt depth in the jejunum was similar between exper-imental groups on days 4 and 11 PW. In the ileum, crypt depth wasincreased in PFTS pigs compared with control and fasted pigs onday 4 PW (p < 0.05). Reductions in villus height in PFTS and fastedpigs resulted in reduced (p < 0.05) villus:crypt ratios in the jeju-num and ileum on days 4 and 11 PW.
3.5. Intestinal epithelial cell morphology in PFTS pigs
Intestinal epithelial cells from control and fasted pigs exhibiteda normal appearance of epithelial cells along the crypt–villus axiswith shortened, cuboidal epithelial cells present in the crypts(‘‘immature’’ epithelium) transitioning to tall, columnar epithelialcells (‘‘mature’’ epithelium) in the villus tip region (Fig. 5). In con-trast, epithelial cells lining the entire crypt-villus axis in ileum(Fig. 4) and jejunum (not shown) from PFTS exhibited an imma-ture, crypt-like phenotype.
4. Discussion
The failure-to-thrive nursery pig is a significant problem inswine production contributing to economic losses through morbid-ity, mortality, and the associated treatment and management
costs; however the pathogenesis remains poorly understood. Inthis study, we investigated the pathophysiology of PFTS by con-ducting functional and histopathologic analysis of the intestine ofpigs exhibiting clinical PFTS. These data demonstrate that PFTS ischaracterized by marked disturbances in intestinal barrier function(increased intestinal permeability), electrogenic ion transport, andintestinal villus and epithelial morphology. Moreover, intestinalbarrier dysfunction and intestinal morphological changes exhib-ited by PFTS pigs were more severe than age-matched, fasted pigsdemonstrating that these changes were not solely attributed toPFTS-associated anorexia.
In the present study, PFTS pigs failed to gain weight during thePW period. Although individual feed intakes for PFTS pigs could notbe measured in this study, it was apparent by daily observationand necropsy findings of PFTS-affected pigs that they were not con-suming feed. Interestingly, fasted pigs lost a significant amount ofbody weight during the 4-day PW period compared with PFTS pigs.It is unclear why PFTS pigs did not lose weight despite exhibitingsevere anorexia. This response either indicates that PFTS pigsmay have an impaired ability to catabolize body stores in the ab-sence of feed intake. In fact, clinical pathology tests were per-formed on PFTS and fasted pigs as part of another study(manuscript in preparation) and revealed that while fasted pigsexhibited markedly elevated urinary ketone levels, indicative ofcatabolic pathways, PFTS failed to exhibit elevated urinary ketones.The significance of this unique metabolic response observed inPFTS is unclear but may represent an underlying metabolic disor-der in PFTS pigs.
The inciting mechanism of intestinal barrier dysfunction ob-served in PFTS is currently unclear. Given that clinical onset of PFTSoccurs after weaning, it appears that PW factors play a critical rolein the clinical onset of PFTS; however, it is possible that pre-wean-ing factors may play a contributing role. It is known that weaningin pigs triggers disturbances intestinal barrier dysfunction (Moeseret al., 2007a,b) and villus atrophy (Spreeuwenberg et al., 2001).These events are due to a combination of several factors includingstress (Moeser et al., 2007a,b), anorexia (Nabuurs et al., 1993;Spreeuwenberg et al., 2001) and diet changes (Miller et al., 1984;Nabuurs et al., 1993). In line with studies in these literature refer-ences, control pigs in the present study exhibited increased intes-tinal permeability (reduced TER and increased FD4 permeability)and villus atrophy at 4 days PW compared with unweaned pigs.Intestinal barrier function and architecture returned to unweanedcontrol levels by 11 days PW demonstrated by the decreased intes-tinal permeability and increase in villus heights in control pigs. InPFTS pigs, intestinal barrier function and villus architecture wasprofoundly disturbed compared with controls and exhibited littleevidence of recovery by 11 days PW. Furthermore, villus atrophyand intestinal barrier dysfunction observed with PFTS was shownto be more pronounced in the ileum compared with the jejunumfor unclear reasons at this time. The ileum is well known to be amajor site of antigen uptake and immune stimulation which couldhave been further provoked by in intestinal barrier disturbancesassociated with PFTS pigs. To determine if anorexia could inducesimilar defects in intestinal dysfunction compared with PFTS, wemeasured intestinal barrier function and villus morphology inage-matched pigs that were subjected to 4 day fasting. Fastingdid not significantly influence PW intestinal barrier function inthe jejunum and had relatively minor effects on TER in the ileum,compared with controls. Fasting did, however, have a negativeinfluence on villus height in the jejunum but not the ileum. Spreeu-wenberg et al. (2001) demonstrated that reduced feed intake atweaning was associated with reduced villus height and increasesin intestinal permeability (Spreeuwenberg et al., 2001); however,the true affect of reduced feed intake on intestinal measurementscould not be conclusively determined. Although PW anorexia did

4 days PWUW
I sc, μ
A .cm
2
-200
-100
0
100
200
Cont PFTS Fast PFTSCont
#
11 days PW
*
*
* #
**
* #
4 days PWUW
I sc, μ
A .cm
2
-600
-400
-200
0
200
Cont PFTS Fast PFTSCont11 days PW
*
** # *
* #
B
A
Fig. 3. Transepithelial short circuit current (Isc) across PFTS ileum. Values representmeans ± SE from n = 6 pigs/experimental group. Jejunal and ileal mucosa weremounted on Ussing chambers for measurement of Isc over a 60-min period.⁄ p < 0.05 compared with unweaned (UW) pigs, #,� p < 0.05 compared with othertreatments within each PW time point. Data were analyzed using a 2-Way ANOVAwith PW time point (4 and 11 days PW) and treatment as the main factors.
A.J. Moeser et al. / Research in Veterinary Science 93 (2012) 975–982 979
not appear to be associated with PFTS lesions, it remains to be fullyelucidated whether PW plays a role in disease pathogenesis. Forexample, it is possible that PW anorexia may serve as an importantPFTS trigger in ‘‘susceptible’’ pigs; however, this has not been dem-onstrated yet. Overall, findings from this study indicate that PWanorexia may have a more profound influence on villus structurecompared with intestinal barrier function. In addition, these datademonstrate that PW factors, in addition to anorexia, are playinga critical role in the intestinal barrier dysfunction and impairedvillus architecture observed with PFTS.
Table 2Morphologic measurements of jejunum and ileum.
Days post-weaning
0 4
Unweaned Control PFTS
JejunumVillus height, lm 314 ± 16 276 ± 9* 194 ± 1Crypt depth, lm 181 ± 8 200 ± 12 193 ± 1Villus:crypt ratio 2.0 ± 0.4 1.7 ± 0.3 1.2 ± 0
IlleumVillus height, lm 307 ± 14 230 ± 13* 105 ± 1Crypt depth, lm 128 ± 10 163 ± 10* 238 ± 1Villus:crypt ratio 2.1 ± 0.2 1.6 ± 0.2* 0.5 ± 0
Values represent means ± SE from n = 6 pigs/experimental group.Data were analyzed using a 2-Way ANOVA with PW time point (4 and 11 days PW) and* p < 0.05 compared with unweaned (UW) pigs.
# p < 0.05 compared with controls within each PW time point.
Impaired intestinal barrier function and damage to villus archi-tecture is a common sequela to enteric infections in weaned pigsincluding rotavirus and transmissible gastroenteritis (TGE) virus(Moeser and Blikslager, 2007; Tafazoli et al., 2001; Vellengaet al., 1988); however, a recent unpublished data from field diag-nostic investigations yield little support for an infectious cause ofPFTS (Harding, 2010; (Pittman and Rovira, 2011a,b). Rotavirustypes (A, B, and C) have been consistently found in PFTS pigs; how-ever similar findings have been found in unaffected cohort controls(Pittman and Rovira, 2011a,b). In addition to enteric infections,weaning stress, mediated via corticotropin releasing factor (CRF)receptor and mast cell activation, is an important cause of intesti-nal barrier dysfunction (Moeser et al., 2007a,b; Smith et al., 2010)in weaned pigs; however, in the present study, all pigs underwentthe same weaning stressors (removal from the dam, co-mingling,transport stress, dietary change) but only PFTS pigs exhibited se-vere barrier dysfunction. Although enteric pathogens and stresssignaling pathways in the gut have a deleterious influence on PWintestinal barrier function, it does not appear that these factorsare playing a major role in PFTS; however it is possible that a spe-cific combination of PW factors are needed in order to influence theclinical onset of PFTS.
Given the markedly impaired barrier function observed in PFTSintestine, it is plausible that bacteria, viruses, and antigens mayeasily traverse the intestinal epithelium inciting local and systemicinflammatory responses that may contribute to PFTS pathogenesis.Interestingly, there was no significant histopathological evidenceof inflammation in the PFTS intestine. It is well known that wean-ing triggers intestinal inflammation in pigs which may be a resultof PW anorexia (McCracken et al., 1999; Pie et al., 2004), increasedexposure to enteric pathogens (Shan et al., 2011), changes in dietcomposition (Hao et al., 2009), and stress (Smith et al., 2010).Although there were no histological evidence of inflammation inthe PFTS pig intestine, it is possible that specific pro-inflammatorycytokines such as TNF-a, IL-6, and prostaglandins or serum endo-toxin levels are elevated however these mediators were not mea-sured in this study. In a parallel experiment conducted with PFTSpigs, complete blood counts and blood chemistry values failed toreveal any evidence of a systemic inflammatory response associ-ated with PFTS (manuscript in preparation).
In the present study, small intestine from PFTS pigs exhibiteddramatically altered short Isc, an index of net electrogenic iontransport properties. In the jejunum, PFTS intestine displayedmarked, negative Isc on 4 and 11 days PW while in the ileum, alarge, negative Isc on day 11 PW was preceded by a positive Isc
on day 4 PW. The electrogenic nature of these unique Isc valuesin the PFTS intestine is presently unclear. Enhanced Isc in the smallintestine is commonly associated with increased electrogenic
11
Fasted Control PFTS
6*,# 198 ± 22*,# 337 ± 31 253 ± 16*,#
2 198 ± 10 247 ± 28* 235 ± 18*
.1*,# 1.2 ± 0.1*,# 1.6 ± 0.1 1.2 ± 0.2*,#
2*,# 211 ± 19* 355 ± 36 173 ± 11*,#
0*,# 164 ± 12* 216 ± 22* 226 ± 23*
.05*,# 1.3 ± 0.2* 1.4 ± 0.09* 0.6 ± 0.2*,#
treatment as the main factors.
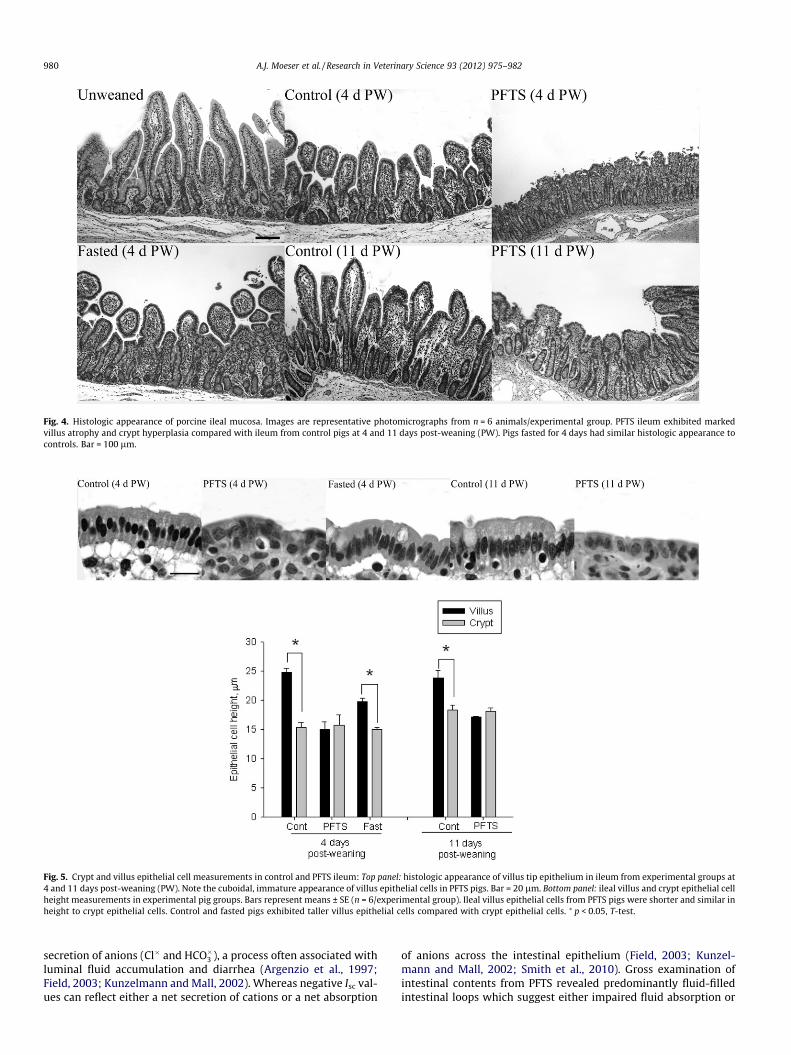
Fig. 4. Histologic appearance of porcine ileal mucosa. Images are representative photomicrographs from n = 6 animals/experimental group. PFTS ileum exhibited markedvillus atrophy and crypt hyperplasia compared with ileum from control pigs at 4 and 11 days post-weaning (PW). Pigs fasted for 4 days had similar histologic appearance tocontrols. Bar = 100 lm.
Fig. 5. Crypt and villus epithelial cell measurements in control and PFTS ileum: Top panel: histologic appearance of villus tip epithelium in ileum from experimental groups at4 and 11 days post-weaning (PW). Note the cuboidal, immature appearance of villus epithelial cells in PFTS pigs. Bar = 20 lm. Bottom panel: ileal villus and crypt epithelial cellheight measurements in experimental pig groups. Bars represent means ± SE (n = 6/experimental group). Ileal villus epithelial cells from PFTS pigs were shorter and similar inheight to crypt epithelial cells. Control and fasted pigs exhibited taller villus epithelial cells compared with crypt epithelial cells. ⁄ p < 0.05, T-test.
980 A.J. Moeser et al. / Research in Veterinary Science 93 (2012) 975–982
secretion of anions (Cl� and HCO�3 ), a process often associated withluminal fluid accumulation and diarrhea (Argenzio et al., 1997;Field, 2003; Kunzelmann and Mall, 2002). Whereas negative Isc val-ues can reflect either a net secretion of cations or a net absorption
of anions across the intestinal epithelium (Field, 2003; Kunzel-mann and Mall, 2002; Smith et al., 2010). Gross examination ofintestinal contents from PFTS revealed predominantly fluid-filledintestinal loops which suggest either impaired fluid absorption or

A.J. Moeser et al. / Research in Veterinary Science 93 (2012) 975–982 981
increased secretion; however these observations did not appear tocoincide with the direction (positive or negative) of the Isc values.Detailed unidirectional ion flux experiments would be needed tobetter understand these differences in ion transport activity inthe PFTS intestine.
Repair of injured intestinal mucosa involves three major eventsthat culminate in the restoration of epithelial continuity and nor-malization of permeability: (1) villus contraction, a mechanismto reduce surface area of injured mucosa, (2) epithelial restitution,where intestinal epithelial cells adjacent to the damaged orsloughed epithelium migrate across exposed basement membraneto restore epithelial continuity, and (3) closure of the paracellularspace and tight junctions where intestinal permeability propertiesare restored (Blikslager et al., 2007). These acute events are fol-lowed by crypt hyperplasia 18–24 h after the initial injury whichreplaces lost or damaged villus epithelial cells and restores normalvillus architecture and digestive and absorptive function (Blikslag-er and Roberts, 1997). In the present study, reductions in villusheight and increased crypt depth were observed in all weanedpig experimental groups compared with unweaned controls. Inweaned controls, reduced villus heights and crypt hyperplasiawas observed on day 4 PW. By day 11 PW, villus height in controlpigs was restored and was accompanied by continued increase incrypt depth thus representing the normal mechanisms of mucosalepithelial repair (crypt hyperplasia and restoration of barrier func-tion) (Blikslager and Roberts, 1997). PFTS pigs exhibited more pro-nounced reductions in villus height and increases and crypthyperplasia compared with controls indicating greater intestinalinjury in PFTS pigs. In contrast to controls, PFTS pigs only partiallyrecovered villus height by day 11 PW and had no evidence of con-tinued crypt hyperplasia between days 4 and 11 PW. Impairedcrypt proliferation may represent a mechanism by which the PFTSintestine fails to restore villus height during the PW period.
The crypt–villus axis divides the epithelium into secretory epi-thelium and absorptive epithelium. The epithelium that performsthese opposing functions is derived from stem cells located withinthe crypts. Newly formed epithelial cells migrate along the crypt–villus axis, maturing in appearance and function until they are ulti-mately sloughed from the tip of the villus approximately 5 daysafter their creation. Immature crypt epithelial cells are cuboidalin appearance and perform predominantly secretory functionswhereas mature villus tip epithelial cells are columnar in appear-ance and perform predominantly absorptive functions. In the pres-ent study we observed distinct differences in intestinal epithelialmorphology between PFTS pigs and controls. The PFTS intestinewas populated by predominantly immature, cuboidal-shaped epi-thelial cells demonstrated by reduction in epithelial height andphenotypic appearance whereas villus tip epithelial cells from con-trol and fasted pigs displayed normal columnar epithelial cells. Thereason for these differences in appearance of PFTS villus epitheliumis unclear. As discussed above, intestinal injury and crypt expan-sion as a result of intestinal injury can result in increased popula-tions of immature, crypt cells that are rapidly migrating up thevillus to restore the epithelial monolayer (Blikslager et al., 1999;Gookin et al., 2003). The functional consequence of this epithelialdefect is not entirely known at this point. However, the increasedproportion of crypt epithelium relative to mature villus epitheliumin the PFTS ileum may be a contributing factor to the barrierdysfunction observed as crypt epithelium is more permeable thanmature villus epithelium (Collett et al., 1997; Gookin et al., 2003;Marcial et al., 1984).
Data from this study revealed that PFTS is associated with pro-found alterations in intestinal barrier function, electrogenic iontransport, and mucosal and epithelial morphology. The uniquefunctional and structural differences observed in the PFTS intestinedo not fit well with known intestinal injury models including
infectious enteritis, stress, or local or systemic inflammatory dis-eases. Although the mechanisms of PFTS remain to be elucidated,findings presented from this study provide valuable clues intothe unique pathophysiology of PFTS that may potentially revealnovel management and therapeutic targets to mitigate thissyndrome.
Funding source
This work was funded by the North Carolina Pork Council, Pro-ject#10M75MF1. The funding agency had initial involvement withsuggestions for improved study design, there was no involvementin the collection, analysis and interpretation of data; in the writingof the manuscript; and in the decision to submit the manuscript forpublication.
Conflict of interest
None of the authors had any conflicts of interest regarding thiswork.
References
Argenzio, R.A., Liacos, J.A., 1990. Endogenous prostanoids control ion transportacross neonatal porcine ileum in vitro. American Journal of Veterinary Research51, 747.
Argenzio, R.A., Armstrong, M., Blikslager, A., Rhoads, J.M., 1997. Peptide YY inhibitsintestinal Cl-secretion in experimental porcine cryptosporidiosis through aprostaglandin-activated neural pathway. Journal of Pharmacology andExperimental Therapeutics 283, 692–697.
Blikslager, A.T., Roberts, M.C., 1997. Mechanisms of intestinal mucosal repair.Journal of the American Veterinary Medical Association 211, 1437–1441.
Blikslager, A.T., Roberts, M.C., Argenzio, R.A., 1999. Prostaglandin-induced recoveryof barrier function in porcine ileum is triggered by chloride secretion. AmericanJournal of Physiology 276, G28–36.
Blikslager, A.T., Moeser, A.J., Gookin, J.L., Jones, S.L., Odle, J., 2007. Restoration ofbarrier function in injured intestinal mucosa. Physiological Reviews 87, 545–564.
Boudry, G., Lalles, J.P., Malbert, C.H., Bobillier, E., Seve, B., 2002. Diet-relatedadaptation of the small intestine at weaning in pigs is functional rather thanstructural. Journal of Pediatric Gastroenterology and Nutrition 34, 180–187.
Boudry, G., Guerin, S., Henri Malbert, C., 2004. Effect of an abrupt switch from amilk-based to a fibre-based diet on gastric emptying rates in pigs: differencebetween origins of fibre. British Journal of Nutrition 92, 913–920.
Collett, A., Walker, D., Sims, E., He, Y.L., Speers, P., Ayrton, J., Rowland, M., Warhurst,G., 1997. Influence of morphometric factors on quantitation of paracellularpermeability of intestinal epithelia in vitro. Pharmaceutical Research 14, 767–773.
Dufrensne, L., Fangman, T.J., Henry, S., 2008. Post-weaning catabolic syndrome –complexities and perspectives. Proceedings from the Allen D. Leman SwineConference, University of Minnesota, St. Paul, MN, 2008.
Field, M., 2003. Intestinal ion transport and the pathophysiology of diarrhea. Journalof Clinical Investigation 111, 931–943.
Gookin, J.L., Galanko, J.A., Blikslager, A.T., Argenzio, R.A., 2003. PG-mediated closureof paracellular pathway and not restitution is the primary determinant ofbarrier recovery in acutely injured porcine ileum. American Journal ofPhysiology Gastrointestinal Liver Physiology 285, G967–979.
Hao, Y., Zhan, Z.F., Guo, P.F., Piao, X.S., Li, D.F., 2009. Soybean beta-conglycinin-induced gut hypersensitivity reaction in a piglet model. Archives of AnimalNutrition 63, 188.
Harding, 2009, Swine health challenges and emerging diseases in Western Canada.In: Proceedings from the Saskatchewan Pork Industry Symposium, Saskatoon,Saskatchewan, pp. 19–20.
Harding, 2010a, Porcine Postweaning Wasting/Catabolic Syndrome (PWCS): theinitial diagnostic investigation. 1. In: Proceedings of the 21st IPVS, 50.
Harding, H.Y., Gauvreau, H., 2010b, Porcine Postweaning Wasting/CatabolicSyndrome (PWCS): pathological features. In: Proceedings of the 21st IPVS, 49.
Kunzelmann, K., Mall, M., 2002. Electrolyte transport in the mammalian colon:mechanisms and implications for disease. Physiological Reviews 82, 245–289.
Madec, F., Bridoux, N., Bounaix, S., Cariolet, R., Duval-Iflah, Y., Hampson, D.J., Jestin,A., 2000. Experimental models of porcine post-weaning colibacillosis and theirrelationship to post-weaning diarrhoea and digestive disorders as encounteredin the field. Veterinary Microbiology 72, 295–310.
Marcial, M.A., Carlson, S.L., Madara, J.L., 1984. Partitioning of paracellularconductance along the ileal crypt-villus axis: a hypothesis based on structuralanalysis with detailed consideration of tight junction structure–functionrelationships. Journal of Membrane Biology 80, 59–70.
McCracken, B.A., Spurlock, M.E., Roos, M.A., Zuckermann, F.A., Gaskins, H.R., 1999.Weaning anorexia may contribute to local inflammation in the piglet smallintestine. Journal of Nutrition 129, 613–619.

982 A.J. Moeser et al. / Research in Veterinary Science 93 (2012) 975–982
Miller, B.G., Newby, T.J., Stokes, C.R., Bourne, F.J., 1984. Influence of diet onpostweaning malabsorption and diarrhea in the pig. Research in VeterinaryScience 36, 187.
Moeser, A.J., Blikslager, A.T., 2007. Mechanisms of porcine diarrheal disease. Journalof the American Veterinary Medical Association 231, 56–67.
Moeser, A.J., Klok, C.V., Ryan, K.A., Wooten, J.G., Little, D., Cook, V.L., Blikslager, A.T.,2007a. Stress signaling pathways activated by weaning mediate intestinaldysfunction in the pig. American Journal of Physiology GastrointestintestinalLiver Physiology 292, G173–G181.
Moeser, A.J., Ryan, K.A., Nighot, P.K., Blikslager, A.T., 2007b. Gastrointestinaldysfunction induced by early weaning is attenuated by delayed weaning andmast cell blockade in pigs. American Journal of Physiology GastrointestinalLiver Physiology 293, G413–421.
Nabuurs, M.J., Hoogendoorn, A., van der Molen, E.J., van Osta, A.L., 1993. Villusheight and crypt depth in weaned and unweaned pigs, reared under variouscircumstances in The Netherlands. Research in Veterinary Science 55, 78–84.
Pie, S., Lallès, J.P., Blazy, F., Laffitte, L., Seve, B., Oswald, I.P., 2004. Weaning isassociated with an upregulation of expression of inflammatory cytokines in theintestine of piglets. Journal of Nutrition 134, 641–647.
Pittman, M.A., Rovira, A., 2011a, Porcine peri-weaning failure to thrive syndrome(PFTS), Part II: Gross lesions, histopathology and diagnostic analysis. In:
Proceeding from the 2011 American Association of Swine Veterinariansmeeting, Phoenix, AZ, pp. 367–377.
Pittman, M.A., Rovira, A., 2011b, Porcine peri-weaning failure to thrive syndrome(PFTS), Part II: Gross lesions, histopathology and diagnostic analysis. In:Proceedings from the 2011 American Association of Swine Veterinariansmeeting, Phoenix, AZ, pp. 367–368.
Shan, T., Li, L., Simmonds, P., Wang, C., Moeser, A., Delwart, E., 2011. The fecalvirome of pigs on a high-density farm. Journal of Virology 85, 11697–11708.
Smith, F., Clark, J.E., Overman, B.L., Tozel, C.C., Huang, J.H., Rivier, J.E., Blikslager, A.T.,Moeser, A.J., 2010. Early weaning stress impairs development of mucosal barrierfunction in the porcine intestine. American Journal of PhysiologyGastrointestinal Liver Physiology 298, G352–363.
Spreeuwenberg, M.A., Verdonk, J.M., Gaskins, H.R., Verstegen, M.W., 2001. Smallintestine epithelial barrier function is compromised in pigs with low feed intakeat weaning. Journal of Nutrition 131, 1520–1527.
Tafazoli, F., Zeng, C.Q., Estes, M.K., Magnusson, K.E., Svensson, L., 2001. NSP4enterotoxin of rotavirus induces paracellular leakage in polarized epithelialcells. Journal of Virology 75, 1540–1546.
Vellenga, L., Wensing, T., Egberts, H.J., van Dijk, J.E., Mouwen, J.M., Breukink, H.J.,1988. Intestinal permeability to macromolecules during colibacillosis in piglets.Veterinary Research Communications 12, 305–311.