Embed Size (px)
Citation preview

DEFECTIVE PHOTOCHEMICAL REPAIR IN EPITHELIUM PREDISPOSED T O
FIELD CANCERIZATION
DANIEL ROTH, MD,* 4ND HAROLD H. SAGE, M D ~
Capability for repairing ultraviolet radiation damage was studied in the desquamated buccal cells of 34 individuals with histories of cancer of the oral cavity and upper respiratory tract, and of 3 asymptomatic smokers. Repair was evaluated in terms of the cellular uptake of a n acridine marker for thymine dimer, in response to visible light and an enzyme derived from baker's yeast possessing photoreactivating ability (cytochrome b2). A repair deficit was dis- played by the tested cells regardless of disease stage, treatment, or cure. Cultured diploid human cells which had been incubated with calf thymus DNA or Simian virus 40 developed a comparable repair deficit showing both dose-relationship and specificity for the incubated DNA. I t is proposed that incorporated DNA, by failing to respond adequately to repair mechanisms, may increase the cell's sensitivity to mutagenic agents.
WO RECENT DEVELOPMENTS HAVE HEIGHT- T ened the general interest in the phenom- ena of the damage inflicted on cells by ul- traviolet light and of its repair. One is the concept that the repair mechanism is non- specific and is capable of correcting various forms of genetic injury in addition to that produced by irradiation.ss13,14 The other is the observation by Cleaver that fibroblasts cultured from cases of xeroderma pigmento- sum are deficient in the repair replication of DNA following irradiation.3 The questions that follow naturally are whether the defec- tive repair of errors in DNA structure plays a significant role in a broader range of human cancers than those which might be genetically determined and, if it does, what mechanisms might operate.
We have investigated this problem with a methodology which uses acriflavine in a selec- tive bond with the DNA of intact cells.16 We have recently observed that this dye binds preferentially between thymine residues, with near-stoichiometry, and that the uptake of dye by predetermined numbers of cells is
Supported by the Division of Cancer Control and Research, New York City Department of Health.
Associate Professor, Pathology; t Associate Pro- fessor. Surgery, New York University Medical Center, New York.- '
Address for reprints: Daniel Roth, MD, Department of Pathology, University Hospital, 560 First Ave., New York, N.Y. 10016.
Received for publication April 11, 1969.
proportional to the extent of irradiation damage inflicted upon them.18 The procedure which we use offers the dual advantages of being applicable to population surveys and to inactive cells such as are shed or easily harvested from accessible human squamous epithelia, the latter resulting from the fact that the acridine dye marks a photochemical transformation in the DNA molecule caused by the absorption of light at a wavelength of 2537 A predominantly. Clinical as well as experimental studies are thus made feasible. For the former we used the desquamated buccal cells of individuals with a history of squamous cancer of the laryngeal and oral mucosa, a region chosen for its strong pro- clivity to undergo multicentric, or field, can- cerization.24 We also included a case of leukoplakia and 3 heavy smokers with no le- sions, because of the clinical associations they bear to cancer of the oral cavity and larynx. For experimental models we selected WI-38 cells, representing a transformable hu- man diploid strain, and HeLa cells, repre- senting a polyploid malignant line.
We examined all these cells for their abil- i ty to recover from the photochemical effects, in terms of dye-binding, of UV irradiation by combined treatment with visible light and a photoreactivating enzyme derived from aero- bic yeast.15 We found that the control cells recovered almost completely in all instances, while all the test cells studied under equiva-
511

512 CANCER September 1969 VO~. 24.
lent conditions exhibited varying degrees of deficient recovery.
MATERIALS AND METHODS
Cells: Buccal cells were obtained by swab- bing normal-appearing areas of the buccal mucosa of 34 consecutively seen subjects with histories of cancer of the lip, gingiva, tongue, palate, pharynx, or larynx. Included indis- criminately were recently diagnosed, un- treated cases, as well as cases previously treated by surgery and/or x-rays. There were some cases with recurrences; 2 had distant metastases. Some had remained apparently cured for as long as 15 years. Their pertinent clinical data are listed in Table 2. T h e cells were transferred by swirling into 0.05 M phosphate-buffered saline solution, pH 6.0- 6.2, then washed 3 times in buffer solution, and used directly or stored u p to 2 weeks at 4°C until used.
T h e WI-38 cells were obtained from Balti- more Biological Laboratories and were main- tained in Basal Eagle medium supplemented with 10% fetal calf serum. The HeLa S3 cells were the gift of Dr. William Kuhns, New York University Medical Center; they were maintained in MEM Eagle medium supple- mented with 10% fetal calf serum. Passage and subculture were carried out as confluent growth was attained. Then the harvested, trypsinized cells were divided into 2 portions -one being used to continue the culture, the other for experiments. These were washed 3 times in buffered saline solution and used immediately, unfixed, in experiments.
Simian virus 40 (SV40) was incubated with the WI-38 cells at a multiplicity of 1.8 x 103 plaque-forming units of the freshly thawed virus per cell, for one hour at 37°C. The supernate was then pipetted off, the monolayer washed 3 times with buffered sa- line solution, and fresh medium added. The cultures were then continued to confluency, at which time cells were harvested for pas- sage and for experiments.
Other WI-38 cells, which had been allowed to monolayer for 48 hours, were incubated with freshly dissolved calf thymus DNA (Sigma) at a concentration of 30 pg per flask for 48 hours. The supernatant medium was then decanted, fresh medium was added, and the culture continued to confluency be- fore harvesting for subculture and experi- ments. A long-lasting augmentation of dye-
uptake developed in cells so treated, directly related to the amount of DNA in the incuba- tion mixture, up to a limiting 30 pg.’7
Measurements: These have been described in detail elsewhere and will only be summa- rized here.18 Acriflavine was freshly diluted into buffered saline solution to a working concentration of 0.011 pg/ml. Aliquots of 50,000 cells, obtained by hemacytometer count, were exposed to 5 ml of the dilute dye in the dark at 35°C with occasional stir- ring, for 10 min. The loss of dye from the supernatant solution was determined fluoro- metrically, and the results were converted, by virtue of the stoichiometry of the binding re- action, into terms expressing the amount of dye taken u p per average cell. A reproduci- bility of k57” was obtained with this tech- nique.
UV irradiation: A single procedure was followed for all experiments. The cells were suspended, with frequent stirring, in 2 ml of buffered saline solution within cylindrical quartz cuvettes measuring 1 cm ID, while be- ing irradiated. This was carried out for 60 min using a 4-watt GE G4T4/I mercury lamp. T h e total irradiation dose to the cells, as measured by a YSI model 65 radiometer, was 2.5 x 104 ergs/mm2. At this dose, a maximal reduction in dye uptake was attained for the cells which were used in these experiments, amounting to a little below 50% of the initial, basal, levels. Dye-binding determinations were made on cell aliquots prior to, and directly after, irradiation.
Photoreactivation treatment: Cell aliquots were drawn u p after irradiation and treated with visible light and a photoreactivating enzyme derived from baker’s yeast (cyto- chrome b2, Sigma)lb at 35”C, pH 6.0-6.2. The light was supplied by a GE Photoflood lamp’ standing 30 cm from the cells, through plate glass 1 cm thick. Enzyme dosage and reac- tion time were varied for different experi- ments. The minima1 dose required for com- plete restoration of initial dye-binding levels (“recovery”) of the control buccal and cul- tured cells was found to be 0.5 “Sigma” unit of enzyme for 10 min at 35°C. The enzyme dose for test cells was calculated from this value to cover the varying amounts of sub- strate. I n this report, the term substrate re- fers to the specific loss of dye-binding by cells after maximal irradiation, under the condi- tions described above. A significant part, if not all, of this loss of dye-binding represents

DEFECTIVE PHOTOCHEMICAL REPAIR * Roth and Sage 513 No. 3
the exclusion of dye molecules from their pre- vious binding sites between adjacent thymine molecules as a result of thymine dimer for- mation, and therefore bears a quantitative equivalence to the number of UV photodim- ers of thymine, generally considered to be the substrate for the photoreactivating enzyme.26 After being treated, the cells were washed 3 times and their uptake of dye determined.
RESULTS
Table 1 lists the dye-binding responses of buccal cells from 18 nonsmoker controls. The difference between initial (basal) and post- irradiation levels is tabulated in column 3 under the heading of UV-specific loss. This is noted to range up to 50% of initial dye- binding values, a change consistent with the theoretical extent to which thymine residues can be dimerized.1911 Column 4 lists the total uptake of dye per cell, after irradiated cells were treated with visible light and enzyme at a constant dose of 0.5 unit for 10 min. Columns 5 and 6 are expressions of the spe- cific, or net, recovery of dye-binding from those levels produced by irradiation to those obtained by the photoreactivating treatment. These values are seen to range between 86% and 100% of complete “recovery,” 11 of the 18 displaying 100% reversal of those UV pho- toproducts which can be marked by the acri- dine method.
Table 2 lists the results obtained in 34 can- cer cases and a single case of leukoplakia. The clinical data include tumor site, patient’s age and sex, modality of treatment, and time elapsed since therapy when this was apprecia- ble. None of these seemed to affect the dye- binding responses.
The table is divided into 3 sections accord- ing to the photoreactivation dosage. In most instances, this was either 0.75 or 1 unit of enzyme, at a reaction time extended to 30 min, thus providing an excessive dose in corn- parison with the control cases when the UV- specific loss in dye-binding is taken as the substrate for the enzyme. Only in 3 cases was there more than 80% recovery, the single in- stance of a recovery of 90% occurring when the reaction time had been inadvertently pro- longed to 40 min. Therefore, with better than equivalent conditions for photoreactivation, in comparison with controls, the buccal cells of tested subjects exhibited a “repair” deficit. Even when the enzyme dose was quadrupled in one group of cases, complete recovery was not achieved. That enzyme dose does play a definite role, however, is to be seen in the real but limited difference in recovery registered among the final 3 cases when 0.75 unit of enzyme was used instead of 1.0 unit for 10 min.
Fig. 1 represents the foregoing data in gra- phic form. Four curves are displayed. Curve 1 represents the initial dye-binding level of
TABLE 1. Photo Repair in Control Buccal Cells
DYE-BINDING DATA (picograms per cell)
(5 1 (6) Net recovery % recovery
(3) (1) (2) UV-spec. loss (4)
0.24 0 15 0.09 0.25 0 09 100% 0.22 0.15 0.07 0.22 0.07 100 0.20 0.12 0.08 0.19 0.07 87 0.19 0.13 0.06 0.20 0.07 100 0.24 0.16 0.08 0.25 0.09 100 0.22 0.17 0.05 0.22 0 05 100 0.21 0.14 0.07 0.22 0 08 100 0.22 0 15 0.07 0.22 0 07 100 n 2.1 0 14 0 09 0.22 0 08 88
Basal After UV (1) minus (2) After PR* (4) minus (2) (5) + (3)
PR @ 0.5 enzyme unit/lO min
0.22 0.15 0.21 0.14 0.23 0 . 1 5
0.07 0.07 0.08
0.22 0.20 0.22
0 07 0.06 0.07
100 -86 87
0.19 0.12 0.07 0.19 0.07 100 0.20 0.13 0.07 0.19 0 06 86 n 33 0 13 0 09 0.22 0.09 100 0.21 0.09 0 : i 2 0.24 0.14 0.10 0.22 0.10 0.12
0.20 0.23 0.22
0.11 0.09 0.12
99 90
100
* PR: Photoreactivating treatment

514 CANCER September 1969 Vol. 24
TABLE 2. Photo Repair in Cancer-Sensitized Buccal Cells
Clinical Data Dye-Binding Data (picograms per cell)
Therapy (3) (4) (5) (6) (0 = none) (1) (2) UV sp loss After Net recovery % recovery
Site Age sex op X-ray Basal After UV ( I ) minus (2) PR* (4) minus (2) (5) t ( 3 )
PR @ 0.75 enzyme unit/30 rnin Tongue 5 0 M X O 0.32 0.20 0.12 0.26 0.06 50 %
Tongue 8 5 M X O 0.28 0.15 0.13 0.25 0.10 77 Tongue 6 7 M X X 0.25 0.15 0.10 0.20 0.05 50 Tongue (12 yrs) 6 5 F X X 0.26 0.08 0.18 0.22 0.14 77
Tongue 5 4 M X O 0.29 0.19 0.10 0.25 0.06 60
Tongue ( 5 yrs ) 6 8 M X O 0.31 0.18 0.13 0.23 0.05 40 Tongue & cheek 5 2 F X O 0.27 0.15 0.12 0.22 0.07 60 Floor of mouth 4 8 F 0 0 0.32 0.25 0.07 0.27 0.02 30 Floor of mouth 4 8 M X X 0.27 0.15 0.12 0.25 0.10 83 Floor of mouth 6 0 M X O 0.33 0.23 0.10 0.27 0.04 40 Floor of mouth 3 8 M X X 0.31 0.15 0.16 0.21 0.06 37 Floor of month (1/2 yr) 60 M 0 X 0.29 0.17 0.12 0.23 0.06 50 Gingiva 5 4 M 0 0 0.31 0.17 0.14 0.25 0.08 57 Tonsil (3/4 yr) 6 5 M X O 0.28 0.12 0.16 0.19 0.07 44 Nasopharynx (5 yr) 6 1 F O X 0.32 0.20 0 . 1 2 0.26 0.06 50 Larynx 7 2 M 0 0 0.27 0.15 0.12 0.24 0.09 75 Larynx (2 yrs) 6 1 F X 0 0.31 0.19 0.12 0.26 0.07 59 Larynx (15 yrs) 4 9 F I 0 0.32 0.22 0.10 0.28 0.06 60
PK @ 0.75 enzyme unit/40 min Tongue 7 9 M O X 0.35 0.26 0.09 0.31 0.05 55 Floor of mouth 6 2 F X O 0.29 0.17 0.12 0.26 0.09 75 Gingiva 7 1 F X O 0.30 0.20 0.10 0.29 0.09 90
PR @ 1 enzyme unit/30 min Tongue 5 0 M X X 0.34 0.19 0.15 0.29 0.10 66 P; Tongue 5 0 M O X 0.33 0.25 0.08 0.29 0.04 50 Tongue 5 0 F 0 0 0.29 0.20 0.09 0 .25 0.05 55 Tongue 2 4 M 0 0 0.35 0.17 0.18 0.23 0.06 33 Lip 7 8 M 0 0 0.29 0.20 0.09 0.23 0.03 33 Palate 5 0 M 0 0 0.33 0 . 2 4 0.09 0.27 0.03 33 Larynx 6 5 M 0 0 0.27 0.17 0.10 0.23 0.06 60
PR @ 1 enzyme unit/lO min Pyriform fossa 7 0 M X O 0.28 0.17 0.11 0.23 0.05 45 Larynx (1 yr) 6 0 M X O 0.29 0.19 0.10 0.25 0.06 60 Tongue (2 yrs) 6 1 F X O 0.29 0.19 0. 10 0.25 0.06 60
Floor of mouth 5 5 M X O 0.25 0.14 0.11 0.23 0.09 81 Leukoplakia 7 0 M X O 0.27 0.19 0.08 0.23 0.04 50
(0.19)t (0.05) (45) Floor of mouth (5 yrs) 55 M X 0 0.28 0 . 1 5 0.13 0.25 0.10 77
(0.22) (0.07) (34)
(0.25) (0.06 ) (50) Palate 7 3 M X O 0.31 0.19 0.12 0 . 2 7 0.08 66
* Photoreactivating treatment The second set of data (in parentheses) for the final 3 cases represent results obtained @ 0 75 enzyme unit/lO min.
each control and test specimen arranged in progressively increasing order. Curve 2 repre- sents the total dye-binding level for each case after irradiation followed by photoreactiva- tion, paired with its initial value. C,urve 3 represents the UV-specific loss for each, and curve 4, the specific amount of dye-binding restored by the action of the enzyme and visi- ble light. The shaded areas express the gross deficit for total dye-binding obtained by pho- toreactivation treatment when matched against initial levels, in the upper pair, and the more precise deficit for specific restora- tion of dye-binding against the UV-specific
loss, in the lower pair of curves, respectively. I t is pointed out that no significant deficit is displayed by the control cases while signifi- cant deficits are invariable in all the test sub- jects (save for the one instance of prolonged enzyme treatment mentioned earlier). Also to be noted is the absence of an upward slope in curves 3 and 4 to parallel that of curve 1, thus suggesting a sufficiency of ultraviolet energy to cause maximal loss of dye-binding sites despite increases in the initial, DNA- specific uptake of dye represented in curve 1. The overall upward slope of curve 2, roughly parallel to that of curve 1, is taken
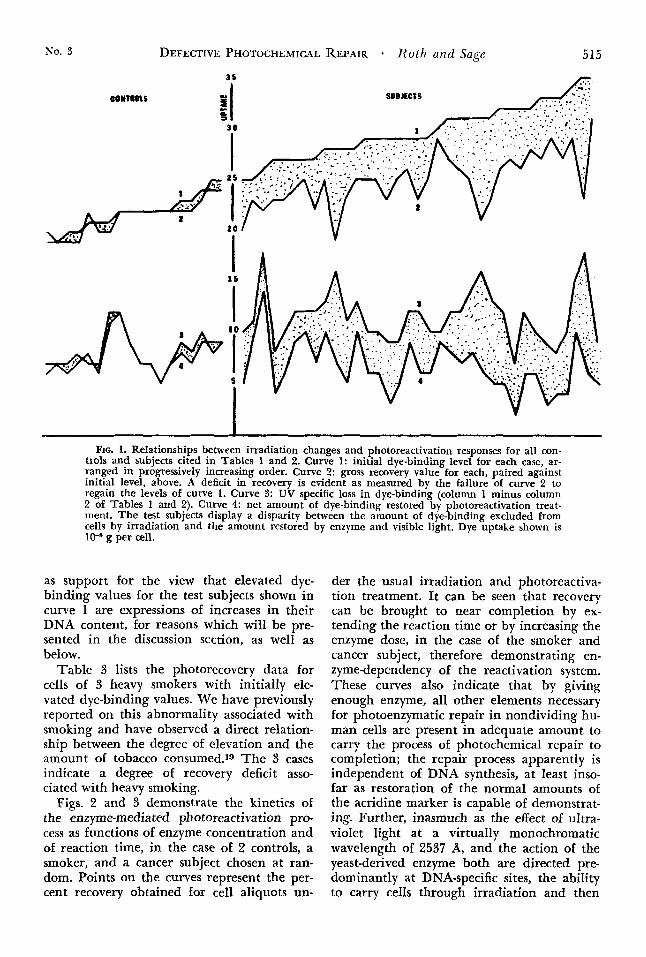
No. 3 DEFECTIVE PHOTOCHEMICAL REPAIR * Roth and Sage 515
contans
3s
SUBJECfS
V
I FIG. 1. Relationships between irradiation changes and photoreactivation responses for all con-
trols and subjects cited in Tables 1 and 2. Curve 1: initial dye-binding level for each case, ar- ranged in progressively increasing order. Curve 2: gross recovery value for each, paired against initial level, above. A deficit in recovery is evident as measured by the failure of curve 2 to regain the levels of curve 1. Curve 3: UV specific loss in dye-binding (column 1 minus column 2 of Tables 1 and 2). Curve 4: net amount of dye-binding restored by photoreactivation treat- ment. The test subjects display a disparity between the amount of dye-binding excluded from cells by irradiation and the amount restored by enzyme and visible light. Dye uptake shown is 10-8 g per cell.
as support for the view that elevated dye- binding values for the test subjects shown in curve 1 are expressions of increases in their DNA content, for reasons which will be pre- sented in the discussion section, as well as below.
Table 3 lists the photorecovery data for ceIIs of 3 heavy smokers with initially ele- vated dye-binding values. We have previously reported on this abnormality associated with smoking and have observed a direct relation- ship between the degree of elevation and the amount of tobacco consumed.~O The 3 cases indicate a degree of recovery deficit asso- ciated with heavy smoking.
Figs. 2 and 3 demonstrate the kinetics of the enzyme-mediated photoreactivation pro- cess as functions of enzyme concentration and of reaction time, in the case of 2 controls, a smoker, and a cancer subject chosen at ran- dom. Points on the curves represent the per- cent recovery obtained for cell aliquots un-
der the usual irradiation and photoreactiva- tion treatment. It can be seen that recovery can be brought to near completion by ex- tending the reaction time or by increasing the enzyme dose, in the case of the smoker and cancer subject, therefore demonstrating en- zyme-dependency of the reactivation system. These curves also indicate that by giving enough enzyme, all other elements necessary for photoenzymatic repair in nondividing hu- man cells are present in adequate amount to carry the process of photochemical repair to completion; the repair process apparently is independent of DNA synthesis, at least inso- far as restoration of the normal amounts of the acridine marker is capable of demonstrat- ing. Further, inasmuch as the effect of ultra- violet light at a virtually monochromatic wavelength of 2537 A, and the action of the yeast-derived enzyme both are directed pre- dominantly at DNA-specific sites, the ability to carry cells through irradiation and then

516 CANCER September 1969 Vol. 24
TABLE 3. Photo Repair in Buccal Cells of Smokers
DYE-BINDING DATA (picograms per cell)
(1) (2) (3) (4) ( 5 ) (6)
Basal After UV UV sp loss After PR Net recovery % recovery
PR @ 1 enzyme unit/30 min 0 .28 0.15 0.13 0.27 0.12 92 % 0 .28 0 .23 0.05 0 .25 0.02 40 0.27 0 . 2 3 0.04 0 . 2 6 0.03 75
back to their initial dye-binding levels by the enzymatic treatment is an indication of the specificity of the DNA-dye bond as well as of the probability that the elevated dye-bind- ing exhibited initially by the test cells is a DNA-dependent event.
I n a possibly analogous fashion, incubation
r------
60 - f E IAI
5 40 - Ly
m
20 -
I i I I 0 0.5 1 1.5 1
FIG. 2. Plot of photochemical repair of buccal cells, in terms of percent recovery of initial dye-binding, as a function of enzyme dose. Complete recovery of 2 con- trols with photoreactivating treatment (0.5-0.75 Sigma units enzyme for 10 min). Incomplete recovery in cells of cancer subject at 2.5 x the enzyme dose despite only a 1 3 1 ratio in UV-specific loss of dye-binding. Intermediate response for cells of smoker with a 1.4:1 ratio in UV-specific loss. Initial dye uptake by con- trols: 0.21 and 0.25 picogramjcell; cancer subject: 0.26; smoker: 0.28.
ENZYME UNITS
of the WI-38 cells with either calf thymus DNA or SV40 led to an increased level of their dye-binding. This new level remains constant over an indeterminate number of cell generations and has been shown to be proportional to the concentration of the DNA in the incubation mixture.17 Table 4 lists the enzyme-catalyzed recovery from irra- diation injury, of these cells and of HeLa cells. T h e enzyme dose was again adjusted between 0.5 and 1.5 units, at a constant re- action time of 10 min to meet the increased substrate requirements which were antici- pated. As it turned out, only the HeLa cells exhibited an increased substrate (UV-specific loss) level. Yet the WI-38 cells incubated with DNA required twice as much enzyme as un- treated cells to attain complete recovery, and the SV40-incubated cells required 3 times as much. The HeLa cells showed the lowest re- covery rate, attaining a level of only 83% recovery at the tripled dose of enzyme. Thus, in the cases of the model cells and the de- squamated buccal cells of cancer subjects, an elevated initial dye-binding level was as- sociated with a disproportionately high en- zyme dose relative to the substrate value, for carrying photochemical repair to completion.
DISCUSSION
It must be stressed that this study treats irradiation injury and photoreactivation (re- pair, recovery) in terms of those photochemi- cal transformations, specific for these events, which can be labelled by the binding of acriff avine. Those which involve sites not “tagged” by the dye, such as purine bases, DNA-protein complexes, enzymes and other proteins, are essentially unlabelled and un- recognized. Until there is more complete un- derstanding of biopolymer-acridine interac- tions, some impreciseness in our conceptions of the bonding sites for dye must remain. Yet there is a fair amount of theoretical and

DEFECTIVE PHOTOCHEMICAL REPAIR - Roth and Sage 517 No. 3
experimental support for the view that the dye associates preferentially between intra- strand and possibly interstrand thymine re% idues.6,18,25 It is the coincidence of this pref- erence and the site of the known major pho- tobiological lesion caused by ultraviolet light that forms the basis for this study: the act of exclusion or of intercalation of dye at sites of thymine dimerization and dimer-splitting, respectively, comprises the major source of the quantitative shifts in dye-binding by the cells. The possibility exists, additionally, that peroxides, formed by the action of ultravio- let light on DNA, may affect dye-binding by virtue of their attack on the phospho-diester backbone, a radical playing an important role in the binding of cationic dyes to nucleic acids.2,
The following points are cited to complete the background for considering the DNA- acriflavine bond as a marker for photoreacti- vation:
1. According to Jagger,g any function of a cell that is photoreactivable is closely linked to nucleic acid activity, since nucleic acid, and only nucleic acid, possesses photoreactiv- able sites; 2. Thymine dimer is the specific photoreactivable lesion in irradiated DNA;22,23 3. The photoreactivating enzyme extracted from baker’s yeast binds only to UV-induced lesions on pyrimidine bases;20 and 4. Cyto- chrome b2 extracted from baker’s yeast has photoreactivating activity.5~ 26
While it has been claimed that cytochrome b2 is not the photoreactivating enzyme, and that any such activity demonstrated in prepa- rations of cytochrome b2 is due to a con- taminant21 we have found that the com- mercial product does act specifically in our experimental system and has displayed a high degree of consistency in this activity through many batches purchased over a 9-month pe- riod.
100
c 8 0 E U U u 0
60 E U u >
40 nl U
20
0
CONTROLS .
I 10 20 30 40
TIME CMIN)
FIG. 3. Photochemical repair in buccal cells of sub- jects shown in Fig. 2, as a function of photoreactivat- ing time at constant enzyme dose of 1 Sigma unit. Again, a disproportion is seen between the recovery from UV-induced levels of dye-binding, in relation to time required for complete recovery, for the cancer and smoker subjects compared to the controls.
From all the foregoing we derived our ra- tionale that the acridine dye serves with ac- ceptable specificity and sensitivity as a marker for thymine photodimer and its repair.
We offer the following conclusions and pos- tulation in final interpretation of the data obtained in this study:
1. The initially elevated dye-uptake by the buccal cells of cancer subjects (Curve 1, Fig. 1) is an expression of their DNA content.
TABLE 4. Photo Repair in Cultured Cells Percent Recovery @ 10 min PR time
Enzyme dose (units) Dye-Binding Data (picograms per cell)
Cell Type Basal After UV UV sp loss 0.5 1 1.5
WI-38 WI-38 plus DNA WI-38 plus SV40
0.18 0.09 0.09 100% 100% ;;;% 100 0.22 0.13 0.09 66
0.23 0.14 0.09 33 56 89 HeLa S3 0.34 0.22 0.12 - 50 83

518 CANCER September 1969 Vol. 24
In addition to the evidence described earlier in this report, we cite previously obtained data which show that all dye-binding by ab- normal as well as normal buccal cells is abolished by pretreatment of the cells with pancreatic DNA’ase, all the specific fluores- cence that can be detected by fluorescence mi- croscopy of the stained cells is localized within their nuclei, the increase in dye-binding which follows the incubation of cultured cells with DNA is proportional to the amount of DNA in the incubation mixture, and, when cell cultures are caused to accumulate cells arrested at metaphase by using colchicine or vinblastine sulfate, the resultant slope of the increasing metaphase count is paralleled by an increase in dye-binding by the cells.l* Finally, the staining procedure itself was orig- inally deveIoped to be specific for DNA in cells.16
2. At the dose of ultraviolet light found to cause a maximal loss of dye-uptake (60 min), the maximal photochemical change capable of being labelled by our acridine marker is achieved (Curve 3, Fig. 1).
3. The deficient recovery of this UV-specific loss in dye-binding by the abnormal cells upon treatment with visible light and enzyme (Curve 4, Fig. 1) is offered as evidence for a specific defect in the repair of UV-damaged DNA which characterizes the buccal cells of subjects predisposed to oral cancer, and of cultured cells which have incorporated exog- enous DNA. It can be proposed that the DNA fraction in those cells, contributing to the increment in dye-binding may, in each case, be exogenous, and that this incorporated DNA, while demonstrating a capacity for rep- lication, may be nonfunctional in transcrib- ing ability ordinarily. Lacking the degree of organization of the cell’s intrinsic DNA, this fraction may not be linked to the essential repair enzyme of the cell. Such unprotected DNA (or nucleotide chains) might there- fore be particularly susceptible to any muta- genic agent encountered at some future date.
In an experimental model of this hypothesis, we incorporated pre-irradiated DNA into WI-38 cells and found that subsequent en- zymatic repair was defective in proportion to the amount of DNA incubated with the cells. This sugested that the specifically dam- aged DNA fraction of exogenous origin was the portion of the cellular DNA which was unable to respond to photoreactivating treat- ment.
T h e yeast enzyme we use is an exogenous substitute for the postulated human enzyme. In fact, a photoreactivating enzyme has not yet been identified as occurring naturally in mammalian cells.4~ 10 However, any enzyme which might exist in mammalian cells, with dehydrogenase activity similar to that of the yeast product, would be inactivated by the amount of ultraviolet light energy which is used experimentally to produce damage to DNA unless it were monochromatically iso- lated with care. We have completely inac- tivated any repair function of the yeast en- zyme by exposing it to the doses of UV light (admittedly high) we use against the cells.
It has been shown recently?, 12 that dehy- drogenases occur naturally within the nuclei of cells as well as in their mitochondria. There- fore, it is conceivable that an enzyme with properties similar to the yeast product could exist normally in the DNA-protein complex of chromosomes and serve to repair acquired errors in the structure of its complementary base chains. Incorporated DNA might have an impaired access to such an enzyme.
If the mutagen which attacked the cell containing incorporated DNA were carcino- genic, the persistence of the genetic error it induced could then trigger an abnormal tran- scribing function leading ultimately to its ex- pression in malignant transformation of the cell. This concept evokes the “latent virus” theory of carcinogenesis and may contribute to it by supplying a mechanism for the initial activation of a persistent viral gene into pathways leading to malignan6y.
REFERENCES
1. BolIum, F. J., and Setlow, R. B.: Ultraviolet in- 3. Cleaver, J. E.: Defective repair replication of activation of DNA primer activity. I: Effects of differ- DNA in xeroderma pigmentosum. Nature (London) ent wave lengths and doses. Biochim. Biophys. Acta 218:652-656, 1968. 68:599-607, 1963. 4. - : Photoreactivation: ,4 radiation repair 2. Butler, J. A. v.: Effects of ultra-violet light on mechanism absent from mammalian cells. Biochem.
nucleic acid and nucleoproteins and other biological BiophYs. Commun. 24:569-5769 1966. systems. Exfierimentia 11:289-293, 1955. 5. Duine, J. A., and Berends, W.: Photoreactivation

No. 3 DEFECTIVE PHOTOCHEMICAL of transforming DNA by cytochrome b2 from yeast. Biochem. Biophys. Res. Commun. 24:888-891, 1966.
6. Gersch, Nerida, F., and Jordan, D. 0.: Inter- action of DNA with aminoacridines, J. Mobc. Biol.
7. Georgiev, G. P.: The Nucleus. I n Enzyme Cytol- ogy, D. B. Roodyn, Ed. New York, Academic Press,
8. Hanawalt, P. C., and Haynes, R. H.: Repair replication of DNA in bacteria: irrelevance of chemical nature of base defect. Biochem. Biophys. Res. Commun. 19:462-467, 1965.
9. Jagger, J:: Introduction to Research in Ultra- violet Photobiology. Englewood Cliffs, Prentice-Hall, 1967; p. 117.
10. Klimek, M.: Formation but n o excision of thy- mine dimers in mammalian cells after UV-irradiation. Neoplasma 12:459-460, 1965.
11. Lamola, A. A., and Yamane, T.: Sensitized photo- dimerization of thymine in DNA. Proc. Nut. Acad. Sci. U.S.A. 58:443446, 1967.
12. McEwen, B. S., Allfrey, V. G., and Mirsky, A. E.: Studies of energy-yielding reactions in thymus nuclei. I. Comparison of nuclear and mitochondria1 phos- phorylation. J. Biol. Chem. 238:758-766, 1963. 111. Participation of glycolysis and the citric acid cycle in nuclear adenosine triphosphate synthesis. Ibid. 238:
13. Patrick, M. H., and Haynes, R. H.: Dark recov- ery phenomena in yeast. Radiat. Res. 23:564579, 1964.
14. Pettijohn, D., and Hanawalt, P. C.: Evidence for repair-replication of ultraviolet damaged DNA in bac- teria. J. Molec. Biol. 9395-410, 1964.
15. Roth, D.: Photoenzymatic repair in mammalian cells revealed by an acridine marker. Biochem. Biophys. Res. Commun. 33:664-669, 1968.
13~138-156, 1965.
1962; pp. 27-102.
2579-2586, 1963.
REPAIR - Roth and Sage 519 16. - , London, M., and Manjon, M.: Binding
specificity and affinity of acriflavine for nucleic acids. Stain Techn. 42:125-132, 1967.
17. - , Manjon, M., and London, M.: Lasting changes in acriflavine binding induced in mammalian cells by exogenous DNA. Exp. Cell Res. 53:lOl-107, 1968.
18. - , and Manjon, M.: Studies of a specific association between acriflavine and DNA in intact cells. Riopolymers (In press).
19. - , Oppenheim, A., and Fredrickson, D. T.: DNA-dependent dye binding by oral epithelium. Its relation to smoking habits. Arch. Environ. Health 17:
20. Rupert, C. S.: Photoenzymatic repair of ultra- violet damage in DNA. 11. Formation of an enzyme- substrate complex. J. Gen. Physiol. 45:725-741, 1962.
21. Setlow, Jane, K.: Cytochrome b2 is not photore- activating enzyme. Biochem. Biophys. Res. Commun. 26:30-31, 1967.
22. - . and Setlow. R. B.: Nature of the Dhoto-
59-61, 1968.
reactivating ‘ ultra-violet ’ lesion in deoxyriborheic acid. Nature 197:560-562, 1963.
23. - , Boling, M. E., and Bollum, F. J.: The chemical nature of photoreactivable lesions in DNA. Proc. Nut. Acad. Sci. U.S.A. 53:143&1436, 1965.
24. Slaughter, D. P., Southwick, H. W., and Smejkal, W.: “Field cancerization” in oral stratified squamous epithelium-clinical implications of multicentric ori- gin. Cancer 6:963-968, 1983.
25. Tubbs, R. K., Ditmars, W. E., Jr., and Van Winkle, Q.: Heterogeneity of the interaction of DNA with acriflavine. J. Molec. Biol. 9:545-557, 1964.
26. Wulff, D. L., and Rupert, C. S.: Disappearance of thymine photodimer in ultraviolet irradiated DNA upon treatment with a photoreactivating enzyme from baker’s yeast. Biochem. Biophys. Res. Commun. 7:237- 240, 1962.





























