Embed Size (px)
Citation preview

Deep-sea benthic foraminiferal recolonisation following avolcaniclastic event in the lower Campanian of the Scaglia
Rossa Formation (Umbria^Marche Basin, central Italy)
S. Galeotti a;b;�, M. Bellagamba a, M.A. Kaminski c;d, A. Montanari e
a Istituto di Geologia dell’Universita', Campus Scienti¢co, Localita' Crocicchia, 61029 Urbino, Italyb Centro di Palinologia dell’Universita', Campus Scienti¢co, Localita' Crocicchia, 61029 Urbino, Italy
c Research School of Geological and Geophysical Sciences, Birbeck College and University College London, Gower Street,London WC1E 6BT, UK
d KLFR, 3 Boyne Avenue, Hendon NW4 2JL, UKe Osservatorio Geologico di Coldigioco, 62020 Frontale di Apiro, Italy
Received 1 February 2000; received in revised form 23 July 2001; accepted 23 July 2001
Abstract
We studied the distribution of deep water agglutinated foraminiferal (DWAF) assemblages across a 15-cm-thickvolcaniclastic layer in the lower Campanian Scaglia Rossa limestones of the Umbria^Marche Basin. Above thevolcaniclastic layer, which is devoid of foraminifera, a remarkable pattern of recovery among DWAF has beenobserved. The complete recovery of DWAF in terms of trophic groups and complexity of assemblages is observed inthe first 5 cm above the volcaniclastic layer, representing 4.8 kyr based upon the mean sedimentation rate of theCampanian Scaglia Rossa Formation.
In its initial stage, the recovery pattern is remarkably similar to that observed following the 15 June 1991 MountPinatubo ashfall in the abyssal South China Sea where various species of Reophax, a small organically cementedspecies of Textularia, and the calcareous species Quinqueloculina seminula and Bolivina difformis are the earliestrecolonisers on top of the tephra layer.
Such similarities between modern and fossil analogues strengthens the reliability of environmental reconstructionsbased on DWAF. ß 2002 Elsevier Science B.V. All rights reserved.
Keywords: Benthic foraminifera; Volcanic ash; Substrate disturbance; Biotic recovery
1. Introduction
Information on life history and communitystructure of deep-sea benthic foraminifera is a
prerequisite for understanding the processes re-lated to disequilibrium in deep-sea benthic com-munities, which constitute a key component of theEarth’s largest biotope. Recolonisation of deep-sea substrates, in particular, is a topic that hasreceived growing interest in recent years with sev-eral studies on both modern and fossil benthicforaminiferal communities following major envi-
0377-8398 / 02 / $ ^ see front matter ß 2002 Elsevier Science B.V. All rights reserved.PII: S 0 3 7 7 - 8 3 9 8 ( 0 1 ) 0 0 0 3 7 - 8
* Corresponding author.E-mail address: [email protected] (S. Galeotti).
MARMIC 851 10-4-02
Marine Micropaleontology 44 (2002) 57^76
www.elsevier.com/locate/marmicro

ronmental disturbances (see Alve, 1999 for a re-view of the topic).
In the fossil record, attention has recently fo-cused on recolonisation patterns by benthic fora-minifera following global catastrophic events suchas the Cretaceous Oceanic Anoxic Events (Kuhnt,1992; Coccioni et al., 1995; Peryt and Lamolda,1996), the Cretaceous^Tertiary Boundary event(Kuhnt and Kaminski, 1993, 1996; Coccioni andGaleotti, 1994, 1998; Speijer and van der Zwaan,1994, 1996; Galeotti, 1998), and the Palaeocene^Eocene Boundary event (Kaminski et al., 1996;Speijer et al., 1995, 1997). Successive recolonisa-tion of the sea £oor has also been suggested toexplain the patterns observed in the vertical suc-cession of ‘£ysch-type’ foraminiferal assemblageswithin turbidite sequences in alpine and borealregions (Gru«n et al., 1964; Butt, 1981; Verdeniusand Van Hinte, 1983; Kuhnt and Kaminski,1989).
In the modern ocean, recolonisation by benthicforaminifera has been observed in shallow waterenvironments following arti¢cial disturbance (El-lison and Peck, 1983; Schafer, 1983), in the deepsea following bottom current disturbance (Kamin-ski, 1985), and after local volcanic ashfalls (Fingerand Lipps, 1981). In an experiment using recolo-nisation trays at an abyssal site in the PanamaBasin, Kaminski et al. (1988a,b) identi¢ed severalspecies of benthic foraminifera as opportunistic,including Psammosphaera, Reophax excentricusand Reophax dentaliniformis.
Deep-sea basins that experience seasonal dys-oxia also support communities of opportunisticbenthic foraminifera. In a study of the deepSan Pedro Basin o¡ southern California, Ka-minski et al. (1995) reported a faunal assem-blage consisting of Psammosphaera, Reophax,and a minute organically cemented species ofTextularia that was interpreted as opportunistic.Many of the species were the same as thosefound in the recolonisation trays in the PanamaBasin, suggesting that di¡erent types of distur-bance may result in similar benthic communities.The sole observations of in situ recolonisation bymodern deep-sea benthic foraminifera in a vastdisturbed habitat were carried out by Hess andKuhnt (1996), Hess (1998), and Hess et al.
(2001), who documented the initial stage of re-colonisation on top of the tephra layer depositedin the South China Sea as a result of the 1991eruption of Mount Pinatubo in the Philippines.The initial observations suggest that faunal re-covery (largely by species of Reophax, Subreo-phax, and a minute organically cemented speciesof Textularia) had begun as early as 3 years fol-lowing the eruption.
The process of faunal decimation and recoveryfollowing tephra falls must undoubtedly be acommon occurrence in the geological record, con-tributing to local di¡erences in the patch structureand successional patterns observed in deep-seabenthic foraminifera. However, a major unknownquestion is whether or not this process leaves anobservable fossil record, and if so what the time-scale of faunal recovery in the deep sea is. In thispaper we address these questions by documentingthe pattern of the recolonisation through a highresolution study of deep water agglutinated fora-miniferal (DWAF) assemblages from below, with-in, and above a 15-cm-thick volcaniclastic layer ina deep-sea sequence of the Scaglia Rossa Forma-tion exposed in the Furlo Gorge (Umbria^MarcheApennines, central Italy).
2. Geological and stratigraphical setting
The upper Turonian^middle Eocene ScagliaRossa Formation in the Umbria^Marche Basinconsists of regularly bedded pink and reddishlimestones interbedded with reddish marly layersdeposited under well oxygenated conditions in alower bathyal depositional environment (Arthurand Fischer, 1977; Kuhnt, 1990). The faunal and£oral associations of the Scaglia Rossa mostlyconsist of calcareous nannofossils and subordi-nate planktonic foraminifera along with rarebenthic foraminifera (mainly DWAF). TheDWAF assemblages in these limestones arehighly diversi¢ed and are unique within theUpper Cretaceous. They include elements ofmixed calcareous and organically cemented bathy-al assemblages, purely agglutinated ‘£ysch-type’assemblages, and a number of species knownfrom Upper Cretaceous sequences deposited be-
MARMIC 851 10-4-02
S. Galeotti et al. / Marine Micropaleontology 44 (2002) 57^7658

low the Carbonate Compensation Depth. Ac-cording to Kuhnt (1990), the agglutinated fora-miniferal assemblage of the Scaglia Rossa is in-dicative of a water depth between 1500 m and2000 m.
In outcrops of the lower Campanian ScagliaRossa sequence at Furlo (Marche region of Italy,Fig. 1), Mattias et al. (1988) described a discrete15-cm-thick bentonitic layer. According to thelitho-, magneto- and biostratigraphic investigationof the Furlo sequence by Alvarez and Lowrie(1984), this bentonitic layer falls within the Globo-truncana elevata planktonic foraminiferal zone,and within the lower part of Chron 33r of earlyCampanian age. Based on detailed geochemicaland mineralogical analyses, Mattias et al. (1988)suggested a volcanic origin for this bentonite. Thelayer is predominantly comprised of calcium dioc-tahedric montmorillonite derived from the trans-formation, in a marine environment, of ¢ne vol-canic glass originating from sub-aerial volcanicactivity. The sudden deposition of a 15-cm-thick(post-compaction thickness) volcanic ash layerwould have created a sudden disturbance of thesea £oor ecosystem, including mass mortality ofbenthic organisms. This event, therefore, repre-sents an interesting case for a high-resolutionstudy of the pattern of recolonisation of thebenthic community, which in this bathyal envi-ronment is normally characterised by a highly di-versi¢ed assemblage of DWAF.
3. Material and methods
To document changes in benthic foraminiferalassemblages and recolonisation patterns abovethe volcaniclastic layer, a quantitative analysisof DWAF was carried out on acid residues orwashed residues obtained from 18 samples. Sam-ples were collected across a 255-cm interval, from125 cm below the volcaniclastic layer (FB1 toFB6), within it (FB7a, b), to 115 cm above it(FB8 to FB17). The interval spanning the ¢rst5 cm above the volcaniclastic layer was sampledevery centimetre (FB8 to FB12).
Considering the duration of Chron 33r (Grad-stein et al., 1995) and its thickness in the Furlosection (Alvarez and Lowrie, 1984), the mean sed-imentation rate during the early Campanian atFurlo was about 8.6 m/Myr. Therefore, assuminga constant sedimentation rate and geologically in-stantaneous deposition of the volcaniclastic layer,the studied interval represents approximately 370kyr. Although a hiatus is known to occur withinthe Scaglia Rossa within the basal Palaeocene(e.g. Dingus, 1984), there is no visible evidenceof hiatuses or turbidites within the studied inter-val.
Limestone samples were soaked in 5% dilutedHCl, wet sieved with tap water and a 63-Wmscreen, and oven dried. The s 63-Wm residueswere split with a microsplitter into subsamplescontaining approximately 300^400 specimens
Fig. 1. Location map of the studied section.
MARMIC 851 10-4-02
S. Galeotti et al. / Marine Micropaleontology 44 (2002) 57^76 59
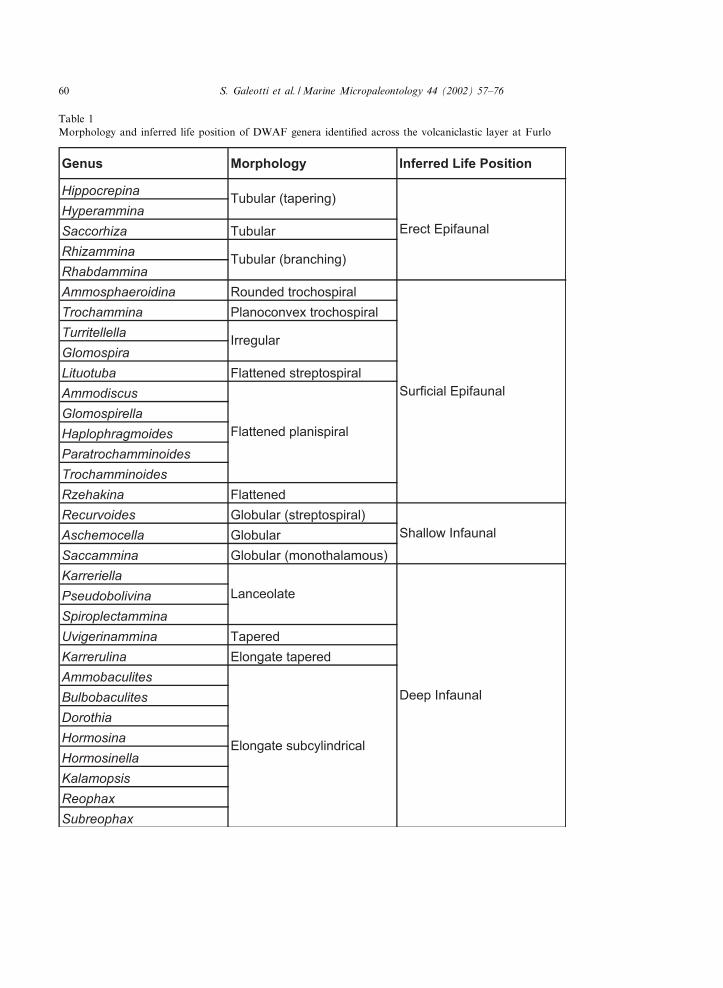
Table 1Morphology and inferred life position of DWAF genera identi¢ed across the volcaniclastic layer at Furlo
MARMIC 851 10-4-02
S. Galeotti et al. / Marine Micropaleontology 44 (2002) 57^7660

each, which were all picked, counted, mounted onmicropalaeontological slides for quantitative anal-ysis, and then stored in the collections of the Paly-nology Center at the University of Urbino. Thetwo samples representing the bentonite layer werewashed with tap water through a 63-Wm sieve.
We largely adopted species concepts of Kamin-ski et al. (1988a,b), Kuhnt (1990), Kuhnt andKaminski (1990, 1993), Wightman and Kuhnt(1992), Kaminski and Geroch (1993), and Kuhntet al. (1998). Specimens not identi¢ed to the ge-neric level are reported as indeterminate forms.For each sample, the Fisher-K index and the per-centages of epifaunal, shallow infaunal, and deepinfaunal forms were calculated. The identi¢cationof epifaunal v. infaunal groups was carried outusing the morphogroup classi¢cation of Jonesand Charnock (1985), Tyszka (1994), Nagy et al.(1995), and Van Den Akker et al. (2000). Theinferred mode of life for each taxon is reportedin Table 1.
Tubular forms occurring in the studied material(mainly Rhizammina indivisa, Rhizammina algae-formis and Rhabdammina sp.) have been combinedinto a single group. Since only small fragments ofthese delicate forms occur in the studied material,counting them as individual specimens wouldoverestimate their relative abundance. At thesame time, considering the numbers of fragmentsas counts, as is often the case, a¡ects the relativeabundance of the non-tubular taxa. As a compro-mise, we subdivided the number of fragments by aconstant value of 15. This number was derived byconsidering the minimum number of unbranchedfragments obtainable from the holotype of R. al-gaeformis (which comprises the largest proportionof the tubular fragments). Although arbitrary, thismethod enables us to obtain an estimate of tubularspecimens that is closer to reality than countingsingle fragments as specimens.
Statistical tests, including R-mode principal com-ponent analysis (PCA) and Q-mode cluster analy-sis, have been performed on the untransformeddata set after exclusion of rare species (those thatoccur as one or two specimens in fewer than threenon-adjacent samples) and grouping species thathave a discontinuous, scattered distribution to thegeneric level. For cluster analysis, groups of com-
positionally similar samples were identi¢ed usingthe Bray^Curtis dissimilarity index (Bray and Cur-tis, 1957) in association with the nearest neighbour,or single linkage, clustering technique.
4. Results
Results are summarised in Table 2 togetherwith the mean distance of each sample from thebentonitic layer. DWAF assemblages from thesurveyed interval are rich and highly diversi¢ed,consisting of not fewer than 70 species represent-ing 33 genera. A minor degree of uncertainty con-cerning the number of occurring species is due tothe presence of specimens not identi¢ed to thegeneric level in some samples. Preservation is gen-erally good, though one of the samples above thebentonitic layer (FB10) exhibits a relatively poorstate of preservation. The most common compo-nents of the DWAF assemblage are Paratrocham-minoides spp., Trochamminoides dubius, Trocham-mina spp., Subreophax spp., Saccammina spp.,and tubular forms (mainly Rhizammina algaefor-mis). Other important components are Haplo-phragmoides spp., Reophax spp., and Ammos-phaeroidina pseudopauciloculata.
The relative abundance of each species £uctu-ates greatly in the surveyed interval, and the larg-est £uctuations are observed just above the volca-niclastic layer (see Table 2 and Fig. 2). Thebentonitic layer itself is virtually devoid of fora-minifera (both benthic and planktonic). Our ex-amination of a large volume of material collectedfrom the lower half of the ash layer revealed justone specimen of Glomospira serpens, while thesamples collected from the upper half of the layerwere completely barren.
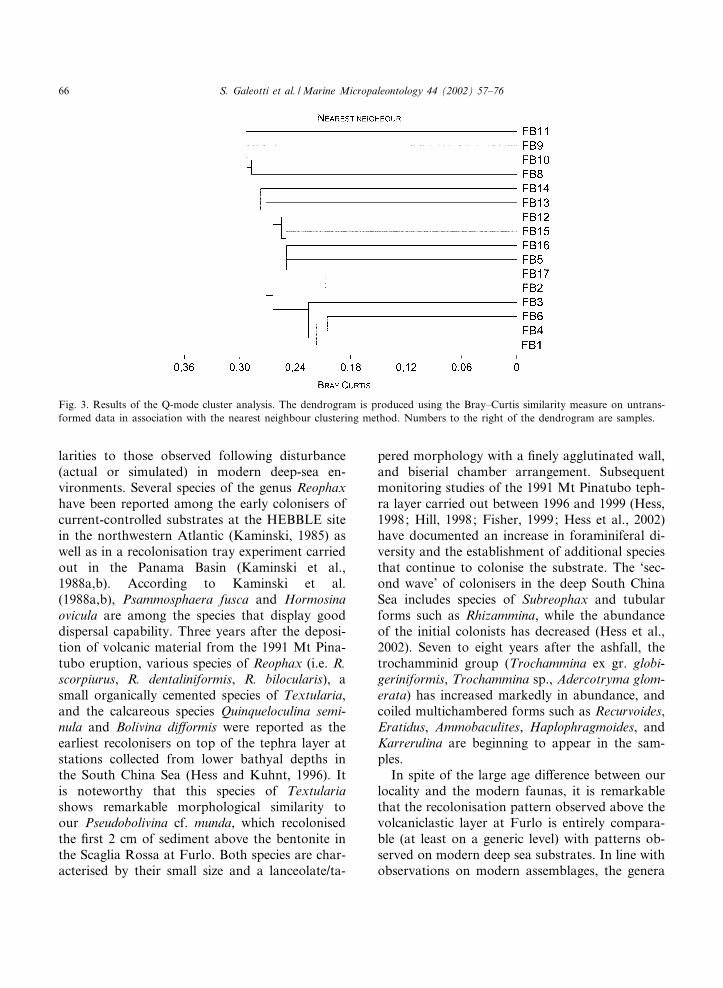
Major di¡erences in diversity, communitystructure, feeding and habitat preferences are ob-served among benthic foraminiferal assemblagesbelow and above the volcaniclastic layer. Clusteranalysis clearly separates two major branches at aBray^Curtis index value of 0.3 (Fig. 3). The ¢rstgroup consists of four samples, representing the¢rst 4 cm above the volcaniclastic layer (the ‘re-colonisation interval’), whereas the second groupincludes all the remaining samples. A further sub-
MARMIC 851 10-4-02
S. Galeotti et al. / Marine Micropaleontology 44 (2002) 57^76 61

Tab
le2
Dis
trib
utio
nof
DW
AF
acro
ssth
evo
lcan
icla
stic
laye
rat
Fur
lo
Sam
ple
FB
1F
B2
FB
3F
B4
FB
5F
B6
FB
7aF
B7b
FB
8F
B9
FB
10F
B11
FB
12F
B13
FB
14F
B15
FB
16F
B17
Mea
ndi
stan
cefr
omth
ebe
nton
ite
laye
r(c
m)
312
53
853
403
153
53
0.5
00
+0.
5+
1.5
+2.
5+
3.5
+4.
5+
10+
19,5
+27
+36
+10
0
Am
mob
acul
ites
sp.
^0.
5^
^^
0.5
^^
^^
1.5
0.5
^^
^0.
92.
00.
5A
mm
odis
cus
cret
aceu
s^
^0.
5^
^0.
5^
^^
^^
^^
^1.
0^
^^
Am
mod
iscu
sgl
abra
tus
1.7
^^
1.9
0.9
1.5
^^
1.4
0.4
0.5
0.5
2.8
1.5
1.0
^0.
5^
Am
mod
iscu
spe
ruvi
anus
^^
^^
^^
^^
^^
^^
^^
^^
^0.
5A
mm
odis
cus
plan
us2.
6^
0.5
4.9
1.8
2.9
^^
1.4
1.8
^1.
51.
45.
82.
40.
40.
5^
Am
mod
iscu
ssp
.^
1.0
^^
^^
^^
^^
^^
^^
^^
^^
Am
mos
phae
roid
ina
pseu
dopa
ucilo
cula
ta7.
73.
010
.08.
78.
57.
8^
^7.
18.
30.
55.
86.
23.
42.
96.
712
.710
.2
Asc
hem
ocel
lasp
.^
^^
^^
^^
^^
^^
^^
^0.
5^
^^
Bul
boba
culit
espr
oble
mat
icus
2.1
1.0
11.
01.
8^
^^
2.8
1.3
1.0
1.5
1.9
3.4
1.4
4.5
1.0
0.9
Dor
othi
asp
.0.
4^
^^
^^
^^
^^
^^
^^
^^
^^
Glo
mos
pira
char
oide
s^
^^
0.5
^^
^^
0.5
^^
^0.
9^
^^
^0.
5G
lom
ospi
ragl
omer
ata
^0.
52.
01.
00.
40.
5^
^^
^^
^^
^^
^0.
5^
Glo
mos
pira
gord
ialis
^2.
02.
51.
06.
31.
0^
^1.
4^
1.0
2.9
3.8
3.4
2.4
^2.
41.
9G
lom
ospi
rase
rpen
s0.
40.
5^
0.5
0.9
^^
^^
0.4
^0.
5^
0.5
^0.
4^
0.5
Glo
mos
pira
spp.
3.0
0.5
^1.
00.
92.
0^
^^
^^
^^
^^
^0.
50.
4G
lom
ospi
rella
gaul
tina
2.6
1.0
^2.
91.
32.
0^
^1.
42.
20.
51
^4.
91.
40.
91.
01.
9H
aplo
phra
gmoi
des
subo
rbic
ular
is^
^^
^^
^^
^^
^^
^^
^^
0.4
^^
Hap
loph
ragm
oide
sw
alte
ri5.
111
.44.
52.
93.
62.
4^
^2.
47.
07.
00.
5^
12.6
6.3
4.5
6.3
10.2
Hap
loph
ragm
oide
ssp
.^
^^
^0.
4^
^^
^0.
41.
5^
1.4
^^
0.4
0.5
0.5
Hip
pocr
epin
ade
pres
sa^
^0.
5^
^^
^^
^^
0.5
^^
^^
0.4
^^
Hor
mos
ina
exce
lsa
^^
1.5
1.0
0.4
0.5
^^
^^
^^
^^
^0.
9^
0.5
Hor
mos
ina
sp.
1.3
0.5
^^
^0.
5^
^^
^^
^^
^^
^^
^H
orm
osin
ella
cf.
dist
ans
3.0
2.5
5.5
1.0
4.0
2.4
^^
5.7
5.3
7.0
2.4
6.2
4.9
4.3
4.5
1.5
2.3
Hyp
eram
min
adi
lata
ta^
^^
0.5
^^
^^
^^
^^
^^
^^
^^
Hyp
eram
min
asp
.0.
41.
5^
^^
0.5
^^
^0.
4^
^0.
5^
^0.
41.
51.
4K
alam
opsi
sgr
zybo
wsk
ii^
^1.
52.
40.
42.
0^
^^
^^
^^
^^
^1.
0^
Kar
reri
ella
conv
ersa
0.4
^^
^^
^^
^^
0.4
^^
^^
^^
0.5
^K
arre
rulin
asp
.0.
94.
54.
51.
05.
42.
0^
^2.
40.
94.
510
.21.
44.
45.
34.
54.
46.
9L
ituo
tuba
litui
form
is0.
4^
^1.
9^
1.0
^^
0.5
^^
^2.
8^
0.5
^^
0.5
Lit
uotu
basp
.^
^^
^^
0.5
^^
^^
^^
^^
^0.
4^
^P
arat
roch
amm
inoi
des
hete
rom
orph
us^
^0.
5^
^^
^^
^^
^1
^^
^^
0.5
^
Par
atro
cham
min
oide
ssp
.1
(Kuh
nt,
1990
)7.
29.
43.
06.
310
.75.
4^
^3.
35.
73.
04.
410
.46.
313
.09.
44.
96.
0
Par
atro
cham
min
oide
ssp
.2
(Kuh
nt,
1990
)6.
05.
92.
53.
96.
32.
0^
^2.
44.
43.
53.
45.
27.
212
.55.
88.
86.
0
MARMIC 851 10-4-02
S. Galeotti et al. / Marine Micropaleontology 44 (2002) 57^7662

Tab
le2
(con
tinu
ed)
Sam
ple
FB
1F
B2
FB
3F
B4
FB
5F
B6
FB
7aF
B7b
FB
8F
B9
FB
10F
B11
FB
12F
B13
FB
14F
B15
FB
16F
B17
Mea
ndi
stan
cefr
omth
ebe
nton
ite
laye
r(c
m)
312
53
853
403
153
53
0.5
00
+0.
5+
1.5
+2.
5+
3.5
+4.
5+
10+
19,5
+27
+36
+10
0
Par
atro
cham
min
oide
s/T
roch
amm
inoi
des
juve
nile
0.9
^0.
5^
^1.
0^
^4.
74.
87.
54.
9^
^^
^1.
0^
Pse
udob
oliv
ina
lage
nari
a^
^^
^^
^^
^^
^^
0.5
^^
^^
^^
Pse
udob
oliv
ina
cf.
mun
da0.
40.
5^
1.0
0.4
0.5
^^
6.1
6.1
3.0
3.9
4.3
1.9
1.0
0.4
1.5
0.5
Pse
udob
oliv
ina
vari
abili
s0.
9^
^^
^^
^^
0.5
^^
^^
^^
0.9
^^
Rec
urvo
ides
nucl
eosu
s^
^1.
00.
5^
0.5
^^
^0.
9^
^^
^2.
4^
0.5
^R
ecur
void
essp
.0.
4^
2.0
0.5
0.4
1.0
^^
0.9
0.4
1.0
1^
1.0
^^
1.0
0.5
Reo
phax
min
utus
3.8
5.4
3.5
3.4
3.6
2.9
^^
14.6
7.9
8.5
8.7
6.6
2.9
10.1
4.9
6.3
5.6
Reo
phax
subn
odul
osus
^2.
52.
0^
^1.
0^
^0.
5^
0.5
^0.
9^
1.0
^0.
50.
5R
eoph
axsp
.5
(Kuh
nt,
1990
)^
^^
1.0
^1.
0^
^2.
4^
1.0
^1.
4^
^^
1.0
0.9
Reo
phax
spp.
^^
^^
^^
^^
0.5
^^
^^
^^
^0.
5^
Rze
haki
naep
igon
a^
^^
^^
^^
^^
0.5
^^
^^
^^
^S
acca
mm
ina
grzy
bow
skii
3.0
2.5
2.0
2.9
0.9
2.0
^^
0.9
3.9
1.5
7.3
3.3
2.4
2.4
8.5
0.5
2.8
Sac
cam
min
apl
acen
ta5.
56.
95.
04.
95.
84.
4^
^4.
74.
43.
05.
35.
210
.26.
36.
74.
93.
2S
acca
mm
ina
spha
eric
a^
^^
^^
^^
^^
^^
^^
^^
0.4
^^
Sac
corh
iza
ram
osa
^^
1.0
1.0
^1.
0^
^^
^^
^^
^^
^^
0.9
Spi
ropl
ecta
mm
ina
dent
ata
0.4
^1.
00.
5^
^^
^^
^^
^^
^^
0.4
^^
Spi
ropl
ecta
mm
ina
laev
is0.
9^
^1.
0^
1.5
^^
^3.
11.
0^
^^
^0.
41.
5^
Spi
ropl
ecta
mm
ina
spp.
^0.
5^
^0.
9^
^^
^^
1.0
2.4
0.5
^^
^^
^S
ubre
opha
xgu
ttif
er0.
92.
52.
52.
93.
13.
4^
^2.
46.
13.
01.
90.
91.
50.
50.
93.
42.
3S
ubre
opha
xps
eudo
scal
aris
0.4
^^
^^
^^
^0.
51.
3^
^^
1.0
^^
0.5
0.5
Sub
reop
hax
scal
aris
0.4
2.0
1.0
2.4
3.6
0.5
^^
4.7
2.2
6.5
2.9
1.4
2.4
3.8
1.3
1.5
1.4
Sub
reop
hax
sple
ndid
us2.
18.
44.
06.
34.
03.
4^
^6.
12.
27.
09.
25.
23.
46.
76.
33.
97.
4S
ubre
opha
xsp
p.^
^1.
00.
50.
42.
4^
^^
^1.
50.
5^
^^
^^
^T
roch
amm
ina
defo
rmis
7.2
5.9
8.5
3.9
6.3
5.4
^^
2.4
1.3
3.0
2.4
4.3
2.4
1.4
3.6
6.3
4.6
Tro
cham
min
acf
.gy
roid
inif
orm
is^
^0.
50.
5^
^^
^^
0.4
^^
^^
^^
1.0
^
Tro
cham
min
asp
p.2.
14.
04.
06.
84.
55.
9^
^1.
40.
43.
0^
^^
0.5
^0.
50.
5T
roch
amm
inoi
des
dubi
us9.
43.
55.
54.
43.
68.
3^
^3.
88.
84.
52.
410
.42.
91.
49.
41.
07.
9T
roch
amm
inoi
des
sept
atus
0.4
^^
^0.
4^
^^
^^
^^
^^
^^
^^
Tro
cham
min
oide
ssu
bcor
onat
us^
^^
^^
^^
^^
^^
^^
0.5
^^
^^
Tro
cham
min
oide
ssp
p.3.
82.
53.
53.
9^
2.4
^^
4.2
^2.
00.
5^
0.5
1.4
0.9
2.0
0.5
Tub
ular
form
s6.
05.
97.
04.
94.
06.
3^
^4.
75.
76.
56.
38.
55.
34.
36.
37.
36.
0T
urri
telle
llasp
.^
^^
^^
0.5
^^
^^
^0.
5^
^^
^^
^U
vige
rina
mm
ina
jank
oi4.
31.
04.
01.
01.
36.
3^
^0.
5^
1.0
10.
90.
51.
40.
4^
0.5
Inde
term
inat
efo
rms
1.7
0.5
0.5
1.9
2.7
1.0
^^
0.9
0.9
1.5
2.4
0.9
2.9
0.5
2.2
2.4
1.4
Num
ber
ofsp
ecim
ens
235
202
201
206
224
205
00
212
228
200
206
211
206
208
223
205
216
MARMIC 851 10-4-02
S. Galeotti et al. / Marine Micropaleontology 44 (2002) 57^76 63

division of the second cluster enables us to distin-guish samples FB12^FB16.
The ¢rst and second axes of the PCA accountedfor 78% and 6% of total variance, respectively.Loading scores for the ¢rst two axes are reportedin Table 3. Based on PCA scores, we identi¢ed¢ve assemblages denominated by the dominanttaxa (Fig. 4). These display a distinctive distribu-tion across the volcaniclastic layer. A comparisonof the relative abundance of faunal groups iden-ti¢ed by the PCA in the pre-ashfall, ‘recolonisa-tion’, and post-recovery intervals is given in Fig. 5.
Pre-ashfall and post-ashfall assemblages arequite similar, despite displaying some minor dif-ferences in the species composition of the DWAF.In particular, the post-ashfall assemblages show
slightly higher proportions of the Subreophaxand Reophax minutus assemblage, and lower rela-tive abundance of the Paratrochamminoides^Tro-chamminoides^Trochammina assemblage. Reophaxminutus, which forms a separate assemblage, is arelatively common taxon throughout the studiedinterval, but shows a peak in relative abundancejust above the volcaniclastic layer. The Pseudo-bolivina cf. munda assemblage, which consists ofP. munda and small specimens of Paratrocham-minoides and Trochamminoides, is found almostexclusively in the ‘recolonisation interval’. Athird PCA assemblage comprised of Subreophaxscalaris, Subreophax splendidus, Karrerulina spp.,and Hormosinella cf. distans shows highest rela-tive abundance in the ‘recolonisation interval’,
Fig. 2. Vertical distribution of selected DWAF taxa across the volcaniclastic layer at Furlo. Notice the scale break between 0and 5 cm above the volcaniclastic layer.
MARMIC 851 10-4-02
S. Galeotti et al. / Marine Micropaleontology 44 (2002) 57^7664

although not immediately above the volcaniclasticlayer.
Below the volcaniclastic layer, DWAF assem-blages are characterised by a rather stable epifau-nal/infaunal ratio, with an average relative abun-dance of infaunal forms around 22% (Fig. 6). Inthis stratigraphic interval, DWAF diversity (Fish-er-K) ranges from a minimum value of 10.7 insample FB2 to a maximum of 16.5 just belowthe ash layer (FB6), with an average value of13.3. The changes recorded by the PCA assem-blages across the volcaniclastic layer are clearlyre£ected in the record of faunal parameters (epi-faunal vs. infaunal abundance and diversity). The‘bloom’ in the elongate morphotypes that domi-nate the Reophax minutus, Pseudobolivina cf. mun-da, and Subreophax assemblages is re£ected in apeak of infaunal forms occurring within the ‘re-colonisation interval’, with a maximum value of46.2% recorded in the ¢rst centimetre above thevolcaniclastic layer. Just above the volcaniclasticlayer, diversity shows average values slightly low-er than those observed in the pre-ashfall interval,dropping to its lowest value in sample FB12, justabove the ‘recolonisation interval’.
Although based on few samples, a gradual re-covery to average values comparable to that ofthe pre-ash interval can be observed upsection.Unlike the rest of the surveyed interval, a clearcovariance between diversity and the proportionof infaunal forms is observed in the ‘recolonisa-tion interval’.
5. Discussion
The DWAF assemblages studied at Furlo aregenerally similar to those reported by Kuhnt(1990) from the Upper Cretaceous Scaglia seriesof the classic Bottaccione section, near Gubbio.The taxonomic composition of these assemblagesis indicative of a lower bathyal depositional set-ting at a palaeodepth of 1500^2000 m.
Prior to the deposition of the volcaniclasticlayer, the DWAF assemblage is indicative of acertain environmental stability as indicated bythe relatively minor £uctuations in the measuredfaunal parameters. This stability was disrupted
catastrophically by the deposition of over 15 cmof volcanic ash, an event that would have oc-curred within a day or two at the lower bathyalwater depths represented by the Scaglia Rossa(Wiesner et al., 1995). It is possible that somesediment-dwelling foraminifera reacted to burialby attempting to exhume themselves, and the spe-cies found within the basal centimetre of the ben-tonite, Glomospira serpens, does possess the typi-cal morphology of an active burrower. However,the ability of foraminifera to burrow upwardthrough volcanic ash is apparently quite limited(see also Hess and Kuhnt, 1996, ¢gs. 3 and 4).
A remarkable pattern that can be interpretedas evidence of faunal recovery characterised thesea £oor following the deposition of the volcanicash. Species such as Reophax minutus and Pseu-dobolivina cf. munda, which behaved as oppor-tunists in the recolonisation process, are inter-preted as infaunal deposit feeders. Accordingly,a special feature of infaunal elongate/lanceolatemorphotypes and, in particular of the genusReophax and its relatives seems to be their highcapability for dispersal. Forms belonging to theSubreophax assemblage (Subreophax scalaris,Subreophax splendidus, Hormosinella cf. distans,and Karrerulina sp.), although bene¢ting fromconditions following the deposition of the volca-niclastic layer, apparently represent ‘secondwave’ colonisers.
The stratigraphic distribution of PCA assem-blages reveals that the highest abundances of ‘re-colonisers’ is restricted to the ¢rst 4 cm above thevolcaniclastic layer. This interval corresponds to4.8 kyr, assuming a constant sedimentation rateof 8.6 m/Myr. However, this value for the timesigni¢cance of the disturbed fauna is certainly anoverestimate, owing to ubiquitous presence of bi-oturbational mixing in the Scaglia Rossa. Thegroup most adversely a¡ected by the depositionof the volcaniclastic layer appears to be the Para-trochamminoides^Trochamminoides^Trochamminaassemblage.
5.1. Comparison with Recent analogues
The recovery pattern of the DWAF in the Sca-glia Rossa at Furlo displays remarkable simi-
MARMIC 851 10-4-02
S. Galeotti et al. / Marine Micropaleontology 44 (2002) 57^76 65
larities to those observed following disturbance(actual or simulated) in modern deep-sea en-vironments. Several species of the genus Reophaxhave been reported among the early colonisers ofcurrent-controlled substrates at the HEBBLE sitein the northwestern Atlantic (Kaminski, 1985) aswell as in a recolonisation tray experiment carriedout in the Panama Basin (Kaminski et al.,1988a,b). According to Kaminski et al.(1988a,b), Psammosphaera fusca and Hormosinaovicula are among the species that display gooddispersal capability. Three years after the deposi-tion of volcanic material from the 1991 Mt Pina-tubo eruption, various species of Reophax (i.e. R.scorpiurus, R. dentaliniformis, R. bilocularis), asmall organically cemented species of Textularia,and the calcareous species Quinqueloculina semi-nula and Bolivina di¡ormis were reported as theearliest recolonisers on top of the tephra layer atstations collected from lower bathyal depths inthe South China Sea (Hess and Kuhnt, 1996). Itis noteworthy that this species of Textulariashows remarkable morphological similarity toour Pseudobolivina cf. munda, which recolonisedthe ¢rst 2 cm of sediment above the bentonite inthe Scaglia Rossa at Furlo. Both species are char-acterised by their small size and a lanceolate/ta-
pered morphology with a ¢nely agglutinated wall,and biserial chamber arrangement. Subsequentmonitoring studies of the 1991 Mt Pinatubo teph-ra layer carried out between 1996 and 1999 (Hess,1998; Hill, 1998; Fisher, 1999; Hess et al., 2002)have documented an increase in foraminiferal di-versity and the establishment of additional speciesthat continue to colonise the substrate. The ‘sec-ond wave’ of colonisers in the deep South ChinaSea includes species of Subreophax and tubularforms such as Rhizammina, while the abundanceof the initial colonists has decreased (Hess et al.,2002). Seven to eight years after the ashfall, thetrochamminid group (Trochammina ex gr. globi-geriniformis, Trochammina sp., Adercotryma glom-erata) has increased markedly in abundance, andcoiled multichambered forms such as Recurvoides,Eratidus, Ammobaculites, Haplophragmoides, andKarrerulina are beginning to appear in the sam-ples.
In spite of the large age di¡erence between ourlocality and the modern faunas, it is remarkablethat the recolonisation pattern observed above thevolcaniclastic layer at Furlo is entirely compara-ble (at least on a generic level) with patterns ob-served on modern deep sea substrates. In line withobservations on modern assemblages, the genera
Fig. 3. Results of the Q-mode cluster analysis. The dendrogram is produced using the Bray^Curtis similarity measure on untrans-formed data in association with the nearest neighbour clustering method. Numbers to the right of the dendrogram are samples.
MARMIC 851 10-4-02
S. Galeotti et al. / Marine Micropaleontology 44 (2002) 57^7666

Reophax, Hormosinella, and Pseudobolivina areindicative of recolonisation following a majorsubstrate disturbance.
An additional indication of stressed environ-mental conditions following the deposition ofthe volcaniclastic layer comes from the highproportion of juvenile and/or dwarfed specimensof the Paratrochamminoides^Trochamminoidesgroup, which forms part of the Pseudobolivinacf. munda PCA assemblage and which is observed
in the ¢rst few centimetres above the bentoniticlayer (see Table 2 and Fig. 2). It is possible thatthese forms, which presumably had an epifaunalto shallow infaunal position in the substrate, werenot able to reach the adult stage owing to the lackof food resources on a comparatively sterile vol-caniclastic substrate or because of competition forresources from the established opportunists livingon the sediment surface. Infaunal foraminiferawould in fact have no reason to colonise the ash
Table 3Loading scores of identi¢ed taxa on the ¢rst two axes of the PCA
Taxon Loadings
First axis Second axis
1 Ammobaculites sp. 0.018 0.0212 Ammodiscus spp. 0.135 30.1113 Ammosphaeroidina pseudopauciloculata 0.333 30.2814 Bulbobaculites problematicus 0.083 0.0375 Glomospira spp. 0.160 30.2046 Glomospirella gaultina 0.076 30.0427 Haplophragmoides walteri 0.263 30.0058 Haplophragmoides sp. 0.016 0.0199 Hormosina spp. 0.021 30.07610 Hormosinella cf. distans 0.187 0.15811 Hyperammina spp. 0.022 30.03012 Kalamopsis grzybowskii 0.019 30.08113 Karrerulina sp. 0.188 0.15114 Lituotuba lituiformis 0.022 30.03315 Paratrochamminoides sp. 1 (Kuhnt, 1990) 0.337 30.11616 Paratrochamminoides sp. 2 (Kuhnt, 1990) 0.265 30.00617 Paratrochamminoides/Trochamminoides juvenile 0.071 0.34118 Pseudobolivina cf. munda 0.094 0.28819 Recurvoides spp. 0.045 0.00720 Reophax minutus 0.296 0.52321 Reophax subnodulosus 0.026 30.01522 Reophax sp. 5 (Kuhnt, 1990) 0.024 0.03723 Saccammina placenta 0.262 30.03724 Saccammina sphaerica 0.145 0.06925 Saccorhiza ramosa 0.011 30.04126 Spiroplectammina spp. 0.050 0.02527 Subreophax guttifer 0.113 0.02328 Subreophax pseudoscalaris 0.013 0.01329 Subreophax scalaris 0.111 0.22430 Subreophax splendidus 0.254 0.20131 Subreophax spp. 0.016 30.02032 Trochammina deformis 0.207 30.30433 Trochammina spp. 0.101 30.22034 Trochamminoides dubius 0.270 30.17935 Trochamminoides spp. 0.087 30.06436 Tubular forms 0.285 30.01737 Uvigerinammina jankoi 0.070 30.188
The ¢rst axis explains 78% of the total variance and the second axis explains 6% of the total variance.
MARMIC 851 10-4-02
S. Galeotti et al. / Marine Micropaleontology 44 (2002) 57^76 67

layer itself because of a lack of food (in the formof bacteria).
The unchanged proportion of tubular formsacross the volcaniclastic layer seems to be atodds with the ¢ndings of Kaminski (1985) andKaminski et al. (1988a,b), who found that tubularepifaunal forms such as Dendrophrya have thelowest dispersal capability. Likewise, these formsare slow at recolonising the 1991 Mt Pinatubo ashlayer in the South China Sea (see data tables inHess, 1998). The proportion of tubular forms inthe ‘recolonisation interval’ (with the exception ofsample FB9) is only slightly lower than that ob-served in the pre-ashfall and post-recovery fauna.However, the relative abundance of tubular fora-minifera is certainly overestimated in these sam-ples because of poorer preservation and increased
fragmentation just above the volcaniclastic layer.Fragments of Rhizammina indivisa collected in the¢rst few centimetres above the volcaniclastic layerare smaller than those observed in the rest of thestudied stratigraphic interval. A possible explan-ation for the increased fragmentation of Rhizam-mina above the volcaniclastic layer is scavengingby bioturbators, which following the deposition ofthe sterile volcaniclastic layer must have been con-centrated near the sediment^water interface. Tub-ular foraminifera are also more likely to be scav-enged by surface-grazing macrofaunal burrowersthan are infaunal species. Burrows ¢lled with tub-ular foraminifera have been observed in boxcoresfrom the South China Sea (Kaminski and Wetzel,unpublished data).
Five centimetres above the volcaniclastic layer
Fig. 4. Plot of taxa scores on the ¢rst two axes of the PCA. The ¢rst and second axes explain 78% and 6% of total variance, re-spectively. Numbers refer to taxa as shown in Table 3. Five assemblages (encircled) are identi¢ed and denominated by the domi-nant taxa.
MARMIC 851 10-4-02
S. Galeotti et al. / Marine Micropaleontology 44 (2002) 57^7668

at Furlo, we observe a return to background val-ues in the relative abundance of the initial recol-onisers (i.e. Reophax minutus, Hormosinella cf.distans, and Pseudobolivina cf. munda), togetherwith the disappearance of juvenile forms of theParatrochamminoides^Trochamminoides group.This change is here interpreted as the ¢nal phaseof the recolonisation process. Based on an estima-tion of the average sedimentation rate for Chron33r in the Scaglia Rossa at Furlo, this return topre-ashfall comparable assemblages occurredsome 5 kyr after the deposition of the volcaniclas-tic material. This estimate is at odds with the ¢nd-ings of Hess et al. (2002), who document the re-turn of trochamminids to the 1991 Mt Pinatubotephra layer some 5^8 years after the volcaniceruption. As mentioned above, this estimate ofthe recovery time at Furlo is certainly a¡ectedby relatively intense bioturbation, which wouldhave displaced part of the original recolonisationassemblage upward.
It is most likely that the deposition of a c. 15-cm-thick volcaniclastic layer (whose original
thickness prior to compaction was probablymuch higher) sealed o¡ the entire benthic fauna.Studies of the 1991 Mt Pinatubo ash layer revealsthat an ash thickness in excess of about 4 cmresults in total mortality of the benthic foramin-ifera within a few years, as oxygen becomes de-pleted beneath the ash layer. The bentonite layerat Furlo contains dispersed pyrite crystals, indi-cating a reducing environment (Mattias et al.,1988). At Furlo, the mechanism responsible fortotal mass mortality would have been burial andsubsequent oxygen depletion of the pore waters,which would have killed o¡ any surviving deepinfaunal species.
In terms of the recolonisation mechanism, thelateral migration of initial recolonisers from areasuna¡ected by the ashfall seems most appropriate.It is interesting to note that the species whichrecolonised the substrate after the ashfall all be-long to the deep infaunal morphogroup, implyingthat they might not be easily transported by bot-tom currents or similar mechanisms. However,the elongate/lanceolate morphology might bemore an adaptation for mobility in the mixedlayer of the sediment. In dysoxic environments,species belonging to the deep infaunal mor-phogroup are found living at the sediment surface(Kaminski et al., 1995). An obvious mechanismfor dispersal would be the drifting of gametes orzygotes during/after reproduction, since numerousspecies of benthic foraminifera possess £agellatedgametes (see table 1 in Lee et al., 1991). Addition-ally, resuspension and transport of the ¢rst recol-onisers by bottom currents is another mechanismfor dispersion. Living specimens of Reophax scottihave been found in plankton tows collected fromthe North Sea (John, 1987), although these organ-isms may have gone into suspension in shallowwater areas during a major storm.
6. Conclusions
Signi¢cant di¡erences in diversity, communitystructure, feeding and habitat preferences are ob-served among benthic foraminiferal assemblagesbelow and above the volcaniclastic layer, whichis virtually barren of foraminifera.
Fig. 5. Distribution of PCA assemblages across the volcani-clastic layer. Pre-ashfall values are calculated as a simpleaverage of data from sample below the volcaniclastic layer.Likewise, post-recovery values represents an average of rela-tive abundances recorded from samples above the ‘recoloni-sation interval’ (i.e. the ¢rst 5 cm above the volcaniclasticlayer). P-T-T = Paratrochamminoides^Trochamminoides^Tro-chammina assemblage. S-R-H = Saccammina^Rhizammina^Haplophragmoides assemblage. The relative abundance ofrare and indeterminate species (reported as R/I) is also re-ported.
MARMIC 851 10-4-02
S. Galeotti et al. / Marine Micropaleontology 44 (2002) 57^76 69

Below the volcaniclastic layer, DWAF assem-blages are diverse with 28^34 species, and only mi-nor £uctuations in the relative abundance of taxo-nomic groups and faunal parameters are observed.Common taxa are Paratrochamminoides spp., Tro-chamminoides dubius, Trochammina spp., Subreo-phax spp., Saccammina spp. and tubular forms.
The recolonising assemblage above the bento-nitic layer is signi¢cantly di¡erent in terms ofboth diversity and relative abundances of taxaand faunal parameters, showing reduced diversityand an increased proportion of infaunal forms.High relative abundances of Reophax minutusand Pseudobolivina cf. munda (the latter being al-most exclusively represented just above the volca-niclastic layer) are observed in the ¢rst centimetre
above the bentonitic layer, in association with ju-venile/dwarfed specimens of Paratrochamminoidesand Trochamminoides. Reophax minutus and P.munda probably represent the initial recolonisersthat appeared following the deposition of the vol-caniclastic material. Between 2 and 4 cm abovethe volcaniclastic layer, the highest relative abun-dances of Subreophax scalaris, Subreophax splen-didus, Hormosinella cf. distans, and Karrerulina sp.are observed. These taxa represent the next stagein the recolonisation process, and although ex-ploiting niches opened up by the mass mortalityof the original fauna, are not as opportunistic asR. minutus and P. munda. The recolonisation fau-na is comprised mainly of morphologies belong-ing to the infaunal morphogroup (Plate I), which
Fig. 6. Relative abundance of inferred infaunal and epifaunal taxa, and DWAF diversity (Fisher-K index) across the volcaniclas-tic layer at Furlo. The proportion of indeterminate species is also reported. Notice the scale break between 0 and 5 cm above thevolcaniclastic layer.
MARMIC 851 10-4-02
S. Galeotti et al. / Marine Micropaleontology 44 (2002) 57^7670

apparently live at the sediment surface in dis-turbed conditions, and are dispersed by deep-seacurrents. The succession of species above the vol-caniclastic layer at Furlo displays interesting sim-ilarities to the succession of species found colonis-ing the 1991 Mt Pinatubo tephra layer in theabyssal South China Sea. Profound di¡erencesbetween the calculated duration of the recolonisa-tion process at Furlo and that observed in theSouth China Sea are attributed to bioturbationalmixing in the Scaglia Rossa, which resulted in anexpanded record of the fossil ‘recolonisation as-semblage’. Despite the passage of 80 million yearsof Earth History, broad similarities at the genericlevel between modern and fossil analogues
strengthens the reliability of environmental recon-structions based on DWAF.
Acknowledgements
We thank J. Erbacher and F. Jorissen for theircritical review that greatly improved the manu-script. This is Contribution No. 66 of the DeepWater Agglutinated Foraminiferal Project.
Appendix A. Taxonomic reference list
All DWAF species identi¢ed in the samples of
Plate I. Typical specimens of the recolonisation fauna above the volcaniclastic layer.
a. Hormosinella cf. distans Brady; sample FB8, U250b. Subreophax splendidus Grzybowski; sample FB8, U250c. Pseudobolivina cf. munda Krasheninnikov; sample FB10, U350d. Reophax minutus Tappan; sample FB8, U226
MARMIC 851 10-4-02
S. Galeotti et al. / Marine Micropaleontology 44 (2002) 57^76 71

this study are alphabetically arranged with refer-ence to their original description.Ammobaculites sp.Test consisting of 3^4 chambers in the coiled
stage later uncoiling to form a rectilinear stagecomposed of 2^3 chambers. The wall is com-posed of medium to ¢ne grain-sized particlesresulting in a somewhat smooth aspect. Aper-ture is a round, simple opening at the end ofthe last chamber.
Ammodiscus cretaceus (Reuss)Operculina cretacea Reuss, 1845, pl. 13, ¢gs. 64, 65.Ammodiscus glabratus Cushman and Jarvis, 1928
pl. 12, ¢gs. 6, 6a.Ammodiscus peruvianus Berry, 1928, pl. 27.Ammodiscus planus Loeblich, 1946, pl. 22, ¢g. 2.Loeblich, 1946. Foraminifera from the type Pep-
per shale of Texas. J. Paleontol., 20, 130^139.Ammodiscus sp.We place in this species specimens showing
a slightly irregular planispiral coiling. Ourspecimens are, therefore, comparable tothose reported as Ammodiscus sp. 1 fromthe Campanian and Maastrichtian ScagliaRossa of the Gubbio section by Kuhnt(1990), who regarded them as intermediateforms between the genera Ammodiscus andTrochamminoides.
Ammosphaeroidina pseudopaciloculata (Mjatliuk)Cystamminella pseudopaciloculata, Mjatliuk, 1966, pl.
1, ¢gs. 5^8; pl. 2, ¢g. 6; pl. 3, ¢g. 3.Aschemocella sp.This species resembles A. carpathica (Neagu) in
possessing large chambers with a relativelycoarsely agglutinated wall. However, in ourmaterial we found only large fragments, whichprevented us from classifying it at the specieslevel.
Bulbobaculites problematicus (Neagu)Ammobaculites agglutinans sp. problematicus Neagu,
1962.Dorothia sp.Test ¢nely agglutinated, consisting of sub-
rounded, in£ated chambers arranged in troco-chospiral coil, later becoming biserial. Theaperture is a low interiomarginal arch, at thebase of the last chamber.
Glomospira charoides (Jones and Parker)
Trochammina squamata var. charoides Jones andParker, 1860, p. 304.
Glomospira glomerata (Grzybowski)Ammodiscus glomeratus Grzybowski, 1898, pl.11,
¢g. 4.Glomospira gordialis (Jones and Parker)Trochammina squamata var. gordialis Jones and
Parker, 1860, p. 304.Glomospira serpens (Grzybowskii)Ammodiscus serpens Grzybowski, 1898, pl. 11, ¢gs. 2,
3.Glomospirella gaultina (Berthelin)Ammodiscus gaultinus Berthelin, 1880, pl. 1, ¢gs. 3a,b.Haplophragmoides suborbicularis (Grzybowski)Cyclammina suborbicularis Grzybowski, 1896, pl. 9,
¢gs. 5^6.Haplophragmoides walteri (Grzybowski)Trochammina walteri Grzybowski, 1898, pl. 11, ¢g. 31.Hippocrepina depressa Vasicek, 1947, pl. 11, ¢gs.
1, 2.Hormosina excelsa (Dylazanka)Hyperammina excelsa Dylazanka, 1923, pl. 1, ¢g. 3.Hormosina velascoensis (Cushman)Nodosinella velasconsis Cushman, 1926, pl. 20, ¢g. 9.Hormosina sp.Test coarsely agglutinated, composed of large
subrounded chambers linked by thick connec-tions.
Hormosinella cf. distans (Brady)cf. Reophax distans Brady, 1881, p. 50.Specimens in our material di¡er from typical H.
distans in having a smaller test and ¢ner ag-glutination.
Hyperammina dilatata Grzybowski, 1896, pl. 8,¢g. 17.
Hyperammina sp.Large subrounded chambers composed of a
¢nely agglutinated, thick wall. Di¡ers from H.dilatata in possessing less elongate, more regu-larly subrounded, instead of sack-like cham-bers.
Kalamopsis grzybowskii (Dylazanka)Hyperammina grzybowskii Dylazanka, 1923, p. 65^66.Karreriella conversa (Grzybowski)Gaudryina conversa Grzybowski, 1901, pl. 7, ¢gs. 15,
16.Karrerulina sp.A small-sized species with a variably elongate,
MARMIC 851 10-4-02
S. Galeotti et al. / Marine Micropaleontology 44 (2002) 57^7672

tapered test. The aperture is slightly producedon a short neck.
Lituotuba lituiformis (Brady)Trochammina lituiformis Brady, 1879, pl. 5, ¢g. 16.Lituotuba sp.Test ¢nely agglutinated with an irregularly coiled
initial portion. This form resembles Glomo-spira irregularis (Grzybowski) from which it isdi¡ers in having the tendency to uncoil in the¢nal part of the test.
Paratrochamminoides heteromorphus (Grzybow-ski)
Trochamminoides heteromorphus Grzybowski, 1898, pl.11, ¢g. 16.
Paratrochamminoides sp.1 Kuhnt, 1990, pl. 5,¢gs. 12^13.
Paratrochamminoides sp.2 Kuhnt, 1990, pl. 5,¢gs. 14^16.
Paratrochamminoides/Trochamminoides juvenileWe place in this group small-sized specimens
showing an irregular coiling of a few roundedto subrounded chambers resembling the initialportion of the test in Trochamminoides andParatrochamminoides.
Pseudobolivina lagenaria Krasheninnikov, 1974,pl. 5, ¢gs. 1a,b, 2c.
Pseudobolivina munda Krasheninnikov, 1973, pl.2, ¢gs. 10^11.
Pseudobolivina variabilis Vasicek, 1947, pl. 1,¢gs. 10^12.
Recurvoides nucleosus (Grzybowski)Trochammina nucleosus Grzybowski, 1898, pl. 11, ¢gs.
28^29.Recurvoides sp.Test small, streptospiral. The 5^6 visible cham-
bers are di⁄cult to distinguish. Specimens inour material are often deformed.
Reophax minutus Tappan, 1940, pl. 14, ¢gs.4a,b.
Reophax subnodulosus Grzybowski, 1898, pl. 10,¢gs. 17^18.
Reophax sp. 5 Kuhnt, 1990, pl. 3, ¢g. 11.Rzehakina epigona (Rzehak)Silicina epigona Rzehak, 1895, pl. 6, ¢g. 1.Saccammina grzybowskii (Schubert)Reophax grzybowskii Schubert, 1902, pl. 1, ¢g. 13.Saccammina placenta (Grzybowski)Reophax placenta Grzybowski, 1898, pl. 10, ¢gs. 9^10.
Saccammina sphaerica Sars, 1872, p. 250.Saccorhiza ramosa (Brady)Hyperammina ramosa Brady, 1879, pl. 3, ¢gs. 14^15.Spiroplectammina dentata (Alth)Textularia dentata Alth, 1850, pl. 13, ¢g. 13.Spiroplectammina laevis (Roemer)Textularia laevis Roemer, 1841, pl. 15, ¢g. 17.Spiroplectammina spp.We place in this group all specimens that, due
to relatively bad preservation of Spiroplectam-mina in the HCl residues, could not be classi-¢ed at species level.
Subreophax guttifer (Brady)Reophax guttifer Brady, 1881, p. 49.Subreophax pseudoscalaris (Samuel)Reophax pseudoscalaria Samuel, 1977, pl. 3, ¢gs. 4a,b.Subreophax scalaris (Grzybowski)Reophax guttifer var. scalaria Grzybowski, 1896, pl. 8,
¢g. 26.Subreophax splendidus (Grzybowski) 1898Reophax splendida Grzybowski, 1898, pl. 10, ¢g. 16.Trochammina deformis Grzybowski, 1898, pl. 11,
¢gs. 20^22.Trochammina gyroidinaeformis Krasheninnikov,
1974, pl. 5, ¢gs. 7a^c, 8a^c, 9c.Trochamminoides dubius (Grzybowski)Ammodiscus dubius Grzybowski, 1898, pl. 8, ¢gs. 12,
14.Trochamminoides septatus (Grzybowski)Ammodiscus spetatus Grzybowski, 1898, pl. 11, ¢g. 1.Trochamminoides subcoronatus (Grzybowski)Trochammina subcoronata Grzybowski, 1896, pl.
9, ¢gs. 3a^c.Turritellella sp.A tubular form characterised by a series of close
convolutions whose diameter slightly increasesin size giving the test a somewhat conicalshape. Aperture terminal, rounded.
Uvigerinammina jankoi Majzon, 1943, pl. 2, ¢gs.15a,b.
References
Alth, A., 1850. Geognostich pala«eontologische Beschreibung d.na«schsten Umgebung von Lembereg. Haidingers Nat.wiss.Abh. 3, 171^284.
Alvarez, W., Lowrie, W., 1984. Magnetic stratigraphy appliedto synsedimentary slumps, turbidites and basin analysis: The
MARMIC 851 10-4-02
S. Galeotti et al. / Marine Micropaleontology 44 (2002) 57^76 73

Scaglia Limestone at Furlo, Italy. Geol. Soc. Am. Bull. 95,324^336.
Alve, E., 1999. Colonisation of new habitats by benthic fora-minifera: a review. Earth Sci. Rev. 46, 167^185.
Arthur, M.A., Fischer, A.G., 1977. Upper Cretaceous^Paleo-cene magnetic stratigraphy at Gubbio, Italy: 1 ^ Lithostra-tigraphy and sedimentology. Geol. Soc. Am. Bull. 88, 367^371.
Berry, E.W., 1928. The smaller foraminifera of the middleLobitos shales of northwestern Peru. Ecl. Geol. Helv. 21,390^405.
Berthelin, G., 1880. Memoire sur les foraminiferes fossiles del’etage Albien de Montcley (Doubs). Mem. Soc. Geol.France 3.1, 1^84.
Brady, H.B., 1879. Notes on some of the reticularian Rhizo-poda of the ‘Challenger’ Expedition, Part 1: On new or littleknown arenaceous types. Q. J. Microsc. Sci. 19, 20^63.
Brady, H.B., 1881. Ueber einge arktische Tiefsee-Foraminife-ren gesammelt wa«hrend der oesterreichisch-ungarischenNordpol-Expedition in den Jahren 1872^1874. K. Akad.Wissensch. Denkschr. 43, 9^110.
Bray, J.R., Curtis, J.T., 1957. An ordination of the uplandforest communities of Southern Wisconsin. Ecol. Monogr.27, 325^349.
Butt, A., 1981. Depositional environments of the Upper Creta-ceous rocks in the northern part of the eastern Alps. Cush-man Lab. Foraminifer. Res. Spec. Publ. 20, 1^121.
Coccioni, R., Galeotti, S., 1994. K/T Boundary extinction:Geologically instantaneous or gradual event? Evidencefrom deep-sea benthic foraminifera. Geology 22, 779^782.
Coccioni, R., Galeotti, S., 1998. What happened to smallbenthic foraminifera at the K/T Boundary? Bull. Soc.Geol. France 169, 271^279.
Coccioni, R., Galeotti, S., Gravili, M., 1995. Latest Albian-earliest Turonian deep-water agglutinated foraminifera inthe Bottaccione section (Gubbio, Italy) ^ Biostratigraphicand paleoecologic implications. Rev. Esp. Paleontol., Nu-mero Homenaje al Dr. Guillermo Colom, 1, 135^152.
Cushman, J.A., 1926. The foraminifera of the Velasco Shale ofthe Tampico Embayment. Am. Ass. Petrol. Geol. Bull. 10,581^612.
Cushman, J.A., Jarvis, P.W., 1928. Cretaceous foraminiferafrom Trinidad. Contrib. Cushman Lab. Foraminifer. Res.4, 85^103.
Dingus, L.W., 1984. E¡ects of stratigraphic completeness oninterpretation of extinction rates across the Cretaceous^Tertiary boundary. Paleobiology 10, 420^438.
Dylazanka, M., 1923. Warstwy inocermowe z lomu x Szym-barku kolo Gorlic. Rocz. Pol. Tow. Geol. 1, 36^80.
Ellison, R.L., Peck, G.E., 1983. Foraminiferal recoloniza-tion on the continental shelf. J. Foraminifer. Res. 13,231^241.
Finger, K.L., Lipps, J.H., 1981. Foraminiferal decimation andrepopulation in an active volcanic caldera, Deception Island,Antarctica. Micropaleontology 27, 111^139.
Fisher, J., 1999. The Recolonisation of the South China Sea byBenthic Foraminifera Following the 1991 Mount Pinatubo
Eruption: Summer 1999. Unpublished M.Res. Thesis, Uni-versity College, London, 114 pp.
Galeotti, S., 1998. Crisi Biologiche e Foraminiferi BentoniciAttraverso i Limiti Cretaceo/Terziario, Paleocene/Eocene eEocene/Oligocene. Unpublished Ph.D. Thesis, University ofParma, 205 pp.
Gradstein, F.M., Agterberg, F.P., Ogg, J.G., Hardenbol, J.,Van Veen, P., Thierry, J., Huang, Z., 1995. A Triassic, Juras-sic, and Cretaceous time scale. SEPM Spec. Publ. 54, 95^126.
Gru«n, W., Lauer, G., Niedemar, G., Schnabel, W., 1964. DieKreide-Tertiar grenze im Wienerwald£ysch bei Hochstrass/Niederosterreich. Verh. Geol. Bundesanst. 2, 226^283.
Grzybowski, J., 1896. Otwornice czerwonych ilow z Wadowic.Rozpr. Akad. Umiejetnosci Krakowie, Wydz. Mat.-Przyr.Ser. 2, 30, 261^308.
Grzybowski, J., 1898. Otwornice poklado¤w naftonosnychokoligy Krosna. ozpr. Akad. Umiejetnosci Krakowie,Wydz. Mat.-Przyr. Ser. 2, 33, 257^305.
Grzybowski, J., 1901. Otwornice wartstw inoceramowych oko-licy Gorlic. ozpr. Akad. Umiejetnosci Krakowie, Wydz.Mat.-Przyr. Ser. 2, 41, 219^288.
Hess, S. 1998. Verteilungsmuster rezenter benthischer Fora-miniferen im Su«dchinesischen Meer. Ber. Geol.-Pala«ontol.Inst. Mus., Christian-Albrechts-Univ. Kiel, 91, 173 pp.
Hess, S., Kuhnt, W., 1996. Deep-sea benthic foraminiferal re-colonization of the 1991 Mt. Pinatubo ash layer in the SouthChina Sea. Mar. Micropaleontol. 28, 171^197.
Hess, S., Kuhnt, W., Hill, S., Kaminski, M.A., Holbourn, A.,De Leon, M., 2002. Monitoring the recolonisation of theMt. Pinatubo 1991 ash layer by benthic foraminifera. Mar.Micropaleontol. 43, 119^142.
Hill, S.J., 1998. The Recolonisation of the South China Sea byDeep-sea Benthic Foraminifera after the 1991 Mount Pina-tubo Volcanic Eruption. Unpublished M.Res. Thesis, Uni-versity College, London, 169 pp.
John, A.W.G., 1987. The regular occurrence of Reophax scottiChaster, a benthic foraminiferan in plankton samples fromthe North Sea. J. Micropalaeontol. 6, 61^63.
Jones, T.R., Parker, W.K., 1860. On the rhizopodal fauna ofthe Mediterranean compared with that of the Italian andsome other Tertiary deposits. Q. J. Geol. Soc. London 16,292^307.
Jones, R.W., Charnock, M.A., 1985. ‘Morphogroups’ of ag-glutinating foraminifera. Their life position and feeding hab-its and potential applicability in (paleo)ecological studies.Rev. Pale¤obiol. 4, 311^320.
Kaminski, M.A., 1985. Evidence for control of abyssal agglu-tinated foraminiferal community structure by substrate dis-turbance: Results from the HEBBLE area. Mar. Geol. 66,113^131.
Kaminski, M.A., Geroch, S., 1993. A revision of foraminiferalspecies in the Grzybowski Collection. In: Kaminski, M.A.,Geroch, S.m Kaminski, D. (Eds.), The Origins of AppliedMicropaleontology: The School of Jozef Grzybowski. Grzy-bowski Foundation Special Publication, Vol. 1, pp. 239^323.
Kaminski, M.A., Boersma, A., Tyszka, J., Holbourn, A.E.L.,1995. Response of deep-water agglutinated foraminifera to
MARMIC 851 10-4-02
S. Galeotti et al. / Marine Micropaleontology 44 (2002) 57^7674

dysoxic conditions in the California borderland basins. In:Kaminski, M.A., Geroch, S., Gasinski, M.A. (Eds.), Pro-ceedings of the Fourth International Workshop on Aggluti-nated Foraminifera. Grzybowski Foundation Special Publi-cation, Vol. 3, pp. 131^140.
Kaminski, M.A., Gradstein, F.M., Berggren, W.A., Geroch,S., Beckmann, J.P., 1988. Flysch-type agglutinated foramini-feral assemblages from Trinidad: Taxonomy, Stratigraphyand Paleobathymetry. In: Gradstein, F.M., Ro«gl, F.(Eds.), Proceedings of the Second Workshop on Aggluti-nated Foraminifera, Vienna 1986. Abh. Geol. Bundesanst.41, 155^228.
Kaminski, M.A, Grassle, J.F., and Whitlatch, R.B., 1988. Lifehistory and recolonization among agglutinated foraminiferain the Panama Basin. In: Gradstein, F.M., Ro«gl, F. (Eds.),Proceedings of the Second Workshop on Agglutinated For-aminifera, Vienna 1986. Abh. Geol. Bundesanst. 41, 229^244.
Kaminski, M.A., Kuhnt, W., Radley, J., 1996. Palaeocene-Eo-cene deep water agglutinated foraminifera from the Numi-dian Flysch (Rif, Northern Morocco): their signi¢cance forthe palaeoceanography of the Gibraltar Gateway. J. Micro-palaentol. 15, 1^19.
Krasheninnikov, V.A., 1973. Cretaceous benthonic foramini-fera, Leg 20, Deep Sea Drilling Project. In: Heezen, B.C.,MacGregor, I.D. (Eds.), Init. Reports. DSDP 20, 205^219.
Krasheninnikov, V.A., 1974. Upper Cretaceous benthonic ag-glutinated foraminifera, Leg 27 of the Deep Sea DrillingProject, In: Veevers, J.J., Hertzler, J.R. (Eds.), Init. Reports.DSDP 27, 631^661.
Kuhnt, W., 1990. Agglutinated foraminifera of western Med-iterranean Upper Cretaceous pelagic limestones (UmbrianApennines, Italy, and Betic Cordillera, Southern Spain). Mi-cropaleontology 36, 297^330.
Kuhnt, W., 1992. Abyssal recolonization by benthic foramin-ifera after the Cenomanian-Turonian boundary anoxic eventin the North Atlantic. Mar. Micropaleontol. 19, 257^274.
Kuhnt, W., Kaminski, M.A., 1990. Palaeoecology of Late Cre-taceous to Paleocene deep-water agglutinated foraminiferafrom the North Atlantic and Western Tethys. In: Hemleben,C., Kaminski, M.A., Kuhnt, W., Scott, D.B. (Eds.), Palae-oecology, Biostratigraphy and Taxonomy of AgglutinatedForaminifera. NATO-ASI Series C327, Kluwer Academic,Dordrecht, pp. 433^505.
Kuhnt, W., Kaminski, M.A., 1989. Upper Cretaceous deep-water agglutinated benthic foraminiferal assemblages fromthe Western Mediterranean and adjacent areas. In: Wied-mann, J. (Ed.), Cretaceous of the western Tethys. Proceed-ings 3rd International Cretaceous Symposium, Tu«bingen1987. E. Schweizerbart’sche Verlagsbuchhandlung, Stutt-gart, pp. 91^120.
Kuhnt, W., Kaminski, M.A., 1993. Changes in the communitystructures of deep water agglutinated foraminifers across theK/T boundary in the Basque Basin (northern Spain). Rev.Esp. Micropaleontol. 25, 57^92.
Kuhnt, W., Kaminski, M.A., 1996. The response of benthicforaminifera to the K/T boundary event ^ A review. Ge¤ol.Afr. Atl. Sud Actes Colloq. Angers 1994, 433^442.
Kuhnt, W., Moullade, M., Kaminski, M.A., 1998. Upper Cre-taceous, K/T Boundary, And Paleocene Agglutinated Fora-minifers from Hole 959D (Leg 159, Co“te d’Ivoire-GhanaTransform Margin). Proc. ODP Sci. Results 159, 389^411.
Lee, J.J., Faber, W.W., Anderson, O.R., Pawlowski, J., 1991.Life cycles of Foraminifera. In: Lee, J.J., Anderson, O.R.(Eds.), Biology of Foraminifera. Academic Press, NewYork, pp. 285^334.
Loeblich, A.R., Jr., 1946. Foraminifera from the type Peppershale of Texas. J. Paleontol. 20, 130^139.
Majzon, L., 1943. Adatok egyes Ka¤rpa¤taljai Flis-re¤tegekhez,tekintettel a Globotruncana¤kra. A Magyar Kira¤lyi Fo«ltaniInte¤zet, Evko«nyve 37, 1^170.
Mattias, P., Montanari, A., Ristori, G.C., Paris, E., 1988. Seg-nalazione di un livello bentonitico nella Scaglia Rossa Cam-paniana Cretacica presso la Gola del Furlo nell’AppenninoMarchigiano (Acqualagna, Pesaro). Miner. Petrogr. Acta31, 243^258.
Mjatliuk, E.V., 1966. K voprosu o foraminiferakh s kremne-zemnym skeletom. Vopr. Mikropaleontol. 10, 255^269.
Nagy, J., Gradstein, F.M., Kaminski, M.A., Holbourn, A.E.,1995. Foraminiferal morphogroups, paleoenvironments andnew taxa from Jurassic to Cretaceous strata of Thakkola,Nepal. In: Kaminski, M.A., Geroch, S., Gasinski, M.A.(Eds.), Proceedings of the Fourth International Workshopon Agglutinated Foraminifera. Grzybowski FoundationSpecial Publication, Vol. 3, pp. 181^209.
Neagu, T., 1962. Studiul foraminiferelor aglutinante din argi-lele Cretacic Superioare de pe Valea Sadovei (C|“mpulung-Moldovenesc) si bazinul superior al Vaii Buzaului. Stud.Cercet. Geol. 7, 45^81.
Peryt, D., Lamolda, M., 1996. Benthonic foraminiferal massextinction and survival assemblages from Cenomanian^Turonian Boundary Event in the Menoyo Section, northernSpain. In: Hart, M.B. (Ed.), Biotic Recovery from MassExtinction Events. Geological Society Special Publication,Vol. 102, pp. 245^259.
Reuss, G.W., 1845. Die Versteinerungen der Bohmischen Krei-deformation. Abtheilung 1. E. Schweizerbarth’sche Verlags-buchhandlung, Stuttgart, p. 58.
Roemer, F.A., 1841. Die Versteinerungen des norddeutschenKreidegebirges. Hahn’schen Hofbuch-handlung, Hannover,145 pp.
Rzehak, A., 1895. Uº ber einige merkwu«rdige Foraminiferen ausdem o«sterrechischen Tertia«r. Ann. K.K. Nat.hist. Hofmus.10, 213^230.
Samuel, O., 1977. Agglutinated foraminifers from Paleogene£ysch formations in West Carpathians of Slovakia. Za¤padn.Karp. Ser. Paleontol. 2/3, 7^69.
Sars, G.O., 1872. UndersÖgelser over Hardangerfjordens Fau-na. Vidensk.-Selk. Christiania Forh. 1871, 246^255.
Schafer, C.T., 1983. Foraminiferal colonization of an o¡shoredump site in Chaleur Bay, New Brunswick, Canada. J. For-aminifer. Res. 12, 317^326.
Schubert, R.J., 1902. Neue und interessante Foraminiferen ausdem su«dtiroler Alttertia«r. Beitr. Palao«ntol. Geol. Oº sterr.-Ung. Orients 14, 9^26.
MARMIC 851 10-4-02
S. Galeotti et al. / Marine Micropaleontology 44 (2002) 57^76 75

Speijer, R.P., van der Zwaan, G.J., 1994. Extinction and sur-vivorship patterns in southern Tethyan benthic foraminiferalassemblages across the Cretaceous/Paleogene boundary.Geol. Ultraiectina 124, 19^64.
Speijer, R.P., van der Zwaan, G.J., 1996. Extinction andsurvivorship of southern Tethyan benthic foraminiferaacross the Cretaceous/Palaeogene boundary. In: Hart,M.B. (Ed.), Biotic Recovery from Mass Extinction Events.Geological Society Special Publication, Vol. 102, pp. 343^371.
Speijer, R.P., Schmitz, B., Aubry, M.P., Charisi, S.D., 1995.The latest Paleocene benthic extinction event: Punctuatedturnover in outer neritic foraminiferal faunas from GebelAweina, Egypt. In: Aubry, M.P., Benjamini, C. (Eds.), Pa-leocene/Eocene Boundary Events in Space and Time. Isr. J.Earth Sci. 44, 207^222.
Speijer, R.P., Schmitz, B., van der Zwaan, G.J., 1997. Benthicforaminiferal extinction and repopulation in response to lat-est Paleocene Tethyan anoxia. Geology 8, 683^686.
Tappan, H., 1940. Foraminifera from the Greyson Formationof northern Texas. J. Paleontol. 14, 93^126.
Tyszka, J., 1994. Response of Middle Jurassic benthic fora-miniferal morphogroups to dysoxic/oxic conditions in the
Pieniny Klippen Basin, Polish Carpathians. Palaeogeogr. Pa-laeoclimatol. Palaeoecol. 110, 55^81.
Van Den Akker, T.J.H.A., Kaminski, M.A., Gradstein, F.M.,Wood, J., 2000. Campanian to Palaeocene biostratigraphyand palaeoenvironments in the Foula Sub-basin, west of theShetland Islands, UK. J. Micropalaeontol. 19, 23^43.
Vasicek, M., 1947. Poznoamki k mikrobiostratigra¢i magur-skeho £yse na Morave. Vesn. Statniho Geol. Ust. Cecko-slov. Repub. 22, 235^266.
Verdenius, J.G., Van Hinte, J.E., 1983. Central Norwegian-Greenland sea: Tertiary arenaceous foraminifera, biostratig-raphy and environment. In: Verdenius, J.G., Van Hinte, J.E.,Fortuin, A.R. (Eds.), Proceedings of the First Workshop onArenaceous Foraminifera, 7^9 September, 1981. ContinentalShelf Institute Norway, Publ. No. 108, pp. 173^224.
Wiesner, M.G., Wang, Y., Zheng, L., 1995. Fallout of volcanicash to the deep South China Sea induced by the 1991 erup-tion of Mount Pinatubo (Philippines). Geology 23, 885^888.
Wightman, W.G., Kuhnt, W., 1992. Biostratigraphy and palae-oecology of late Cretaceous abyssal agglutinated foramini-fers from the Western Paci¢c Ocean (Deep Sea Drilling Proj-ect Holes 196A and 198A and Ocean Drilling Program Holes800A and 801A). Proc. ODP Sci. Results 129, 247^264.
MARMIC 851 10-4-02
S. Galeotti et al. / Marine Micropaleontology 44 (2002) 57^7676





























