Embed Size (px)
Citation preview

1
Shared transcriptional signature in C. elegans dauer larvae and long-lived daf-2 mutants implicates detoxification system in longevity
assurance
Joshua J. McElwee*, Eugene Schuster†, Eric Blanc†, James H. Thomas‡ and David Gems*,§
*Department of Biology, University College London, WC1E 6BT, UK; †European Bioinformatics
Institute, Hinxton, Cambridge CB10 1SD, UK; ‡Department of Genome Science, University of
Washington, Seattle WA 98195, USA.
Running title: Shared longevity-associated transcriptional signatures
§To whom all correspondence should be addressed: Department of Biology, University College London,
Gower Street, London WC1E 6BT, UK.
Tel.: +44 (0) 207 679 4381; Fax.: +44 (0) 207 679 7096; E-mail: [email protected].
The abbreviations used are: CTLD, C-type lectin-like domain; CYP, cytochrome P450; DBE, daf-16-
binding element; DAE, daf-16-associated element; DUF, domain of unknown function; GST,
glutathione S-transferase; HSE, heat shock element; HRE, hif-1 response element; HSAS, heat shock
associated site; IIS, insulin/IGF-1 signaling; SDR, short-chain dehydrogenase/reductase; smHSP, small
heat shock protein; TRP, transthyretin-related protein; UGT, UDP-glucuronosyltransferase.
JBC Papers in Press. Published on August 11, 2004 as Manuscript M406207200
Copyright 2004 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

2
SUMMARY
In the nematode Caenorhabditis elegans, formation of the long-lived dauer larva, andadult aging are both controlled by insulin/IGF-1 signaling (IIS). Potentially, increased
adult lifespan in daf-2 insulin/IGF-1 receptor mutants results from mis-expression in
the adult of a dauer larva longevity program. Using oligonucleotide microarrayanalysis we identify a dauer transcriptional signature in daf-2 mutant adults. By means
of a non-biased statistical approach we identify gene classes whose expression isaltered similarly in dauers and daf-2 mutants, which represent potential determinants
of lifespan. These include known determinants of longevity: the small heat shock
protein (smHSP)/α-crystallins are up-regulated in both milieus. The cytochrome P450,
short-chain dehydrogenase/reductase, UDP-glucuronosyltransferase and (in daf-2
mutants) glutathione S-transferase gene classes were also up-regulated. These four
gene classes act together in metabolism and excretion of toxic endobiotic and
xenobiotic metabolites. This suggests that diverse toxic lipophilic and electrophilicmetabolites, disposed of by phase 1 and phase 2 drug metabolism, may be major
determinants of the molecular damage that causes aging. In addition, we observeddown-regulation of genes linked to nutrient uptake, including nhx-2 and pep-2. These
work together in uptake of dipeptides in the intestine, implying dietary restriction in
daf-2 mutants. Some gene groups up-regulated in dauers and/or daf-2 were enrichedfor certain promoter elements: the daf-16-binding element, the heat shock response
element, the heat shock-associated sequence or the hif-1 response element. Bycontrast, the daf-16-associated element was enriched in genes down-regulated in
dauers and daf-2 mutants. Thus, particular promoter elements appear longevity or
aging associated.
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

3
INTRODUCTION
The biological processes that determine adult lifespan remain largely unknown. Over
the last decade, analysis of mutants with altered rates of aging has led to the discoveryof many genes and molecular pathways which act as determinants of lifespan. For
example, the insulin/insulin-like growth factor 1 (IGF-1) signaling system is a
powerful regulator of aging in C. elegans: lowered insulin/IGF-1 signaling (IIS) canlead to more than a doubling of lifespan (the Age phenotype)(1-3). Similar effects
have been observed in the fruitfly D. melanogaster (4,5). Recent findings suggest thatlifespan in the mouse is influenced by both insulin and IGF-1 signaling (6,7). Thus,
the role of IIS in the control of lifespan shows evolutionary conservation. However,
the lifespan-determining processes that IIS regulates remain to be identified, and arethe topic of this report.
The IIS system is one of several molecular signal transduction pathways which
regulate larval diapause in C. elegans (8). Under adverse environmental conditions(e.g. high temperature, low food and high population density) developing larvae can
form a long-lived, non-feeding, stress-resistant form: the dauer larva (9). Thisdiapausal form can survive for more than three months, during which time it can
resume development if dauer-inducing conditions are reversed; by contrast, adult C.
elegans die of old age after only 2-3 weeks (10). Many mutations which reduce IISresult in constitutive dauer larva formation (the Daf-c phenotype), even under non-
dauer inducing conditions (8). For example, mutations affecting the genes daf-2
(insulin/IGF-1 receptor)(3) and age-1 (phosphatidylinositol 3-kinase)(11) have this
effect.
Potentially, the large increase in lifespan seen in many IIS mutant adultsreflects expression in the adult of dauer-associated longevity assurance processes (2).
A number of observations support this view: for example, dauer larvae and long-liveddaf-2 and age-1 mutant adults have increased activity levels of the antioxidant enzyme
superoxide dismutase (SOD)(12-14), and the mitochondrial MnSOD gene sod-3 shows
increased mRNA levels in both dauer larvae and daf-2 mutants (15-17). Additionally,severe daf-2 mutant adults exhibit a number of dauer-like behavioural characteristics,
including cessation of feeding, and adoption of an immobile, dauer-like posture (18).
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

4
Extensive information about gene expression patterns in dauer larvae and IIS
mutant adults has recently been generated by transcriptional profile studies.Differences in mRNA abundance between dauer larvae and mixed stage growing
populations have been measured using serial analysis of gene expression (SAGE)(19).More recently, microarray analysis was used to examine gene expression changes
occurring during dauer exit, and identified 1,984 genes showing significant expression
changes (20).The increase in adult lifespan resulting from reduced IIS is suppressed by loss of
function of the gene daf-16 (2,21,22). This gene encodes a FOXO class forkheadtranscription factor (23,24), and it is therefore likely that the action of genes
differentially regulated by DAF-16 is a major determinant of the effects of IIS on
aging. Previously, several gene classes were identified which are regulated by IIS in adaf-16-dependent manner. For example, increases have been reported in age-1 and
daf-2 mutants of enzyme activity or gene expression levels of antioxidant proteins
such as catalase, Cu/ZnSOD (cytosolic) and MnSOD (13,15,25). Also up-regulated area number of heat shock proteins (26), and the heat shock transcription factor (hsf-1) is
required for the Age phenotype (27).Genome-wide surveys of transcriptional changes resulting from reduced IIS
have been performed using spotted DNA microarrays (16,17). These have led to broad
observations of correlations between genes regulated by IIS and particular functionalprocesses or gene groups. An earlier comparison of data from over 500 microarray
experiments identified a number of clusters of genes with correlated expression (28).These clusters were represented in the form of a 3D topomap, in which co-expressed
genes form 'topomountains'. Reduced IIS increases expression of many genes
associated with topomountain (Mt.) 15, and down-regulation of genes associated withMts. 8, 19, 27 and 22 (16). Moreover, reduced IIS leads to up-regulation of several
functional gene classes (e.g. antibacterial peptides), and down-regulation of others(e.g. vitellogenins)(17).
Although insights have emerged from these studies, interpreting such
transcriptional profiles remains difficult. Microarray experiments often generate listsof genes which are difficult to interpret in an unbiased fashion. Given a long enough
list of differentially expressed genes, proof may be found for whatever theory ischerished by the investigator, or fashionable in the research community. To
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

5
circumvent this problem, we have taken a novel approach. We make the following
assumptions: a) that some common mechanisms increase lifespan in dauer larvae anddaf-2 mutant adults; and b) that these shared mechanisms are reflected by common
changes in gene transcription. We have compared previously reported expressionprofiles of genes altered during dauer exit (20) with profiles for genes regulated by
daf-2 in a daf-16-dependent manner, newly generated using whole genome
oligonucleotide microarray analysis. We find that transcriptional changes occurring indauer larvae are partially recapitulated in long-lived daf-2 adults. Using a non-biased
statistical analysis, we have identified a number of gene categories that aresignificantly enriched for members with altered expression in both dauer and daf-2
adult animals. This provides information about dauer-specific processes, which are
mis-expressed in long-lived daf-2 mutant adults, and which potentially controllongevity and aging. We also identify several promoter elements that are enriched
among dauer- and daf-2-regulated genes.
EXPERIMENTAL PROCEDURES
Strains and Media --- The following strains were used for strain constructions:
SS104 glp-4(bn2ts) I (29); DR1563 daf-2(e1370) III; DR1567 daf-2(m577) III (18);
and GR1307 daf-16(mgDf50) I (23). From these, the following strains wereconstructed by standard means: glp-4 I; daf-2(m577) III, glp-4 daf-16; daf-2(m577),
glp-4; daf-2(e1370), glp-4 daf-16; daf-2(e1370). glp-4 is included in all strains toprevent egg and progeny production, thereby limiting mRNA analysis to that of
somatic adult tissues.
For routine strain maintenance, strain constructions and lifespan analysis,nematodes were maintained on NGM agar plates, and Escherichia coli strain OP50
was used as a food source for all aspects of this study (30). For preparation of largenumbers of nematodes for RNA isolation, 9-cm diameter enriched peptone NGM
plates were used to maximize growth (31).
Analysis of lifespan --- For lifespan analysis, animals were raised at 15˚C andtransferred to 25˚C at L4 stage. Lifespan of populations was measured on NGM plates
as previously described (18). Kaplan Meier analysis was conducted on survival data,and survival of populations of different genotypes compared using the non-parametric
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

6
log rank test, using the statistical analysis package JMP 3.2.2 (SAS Institute Inc. Cary
NC, USA).Isolation of RNA for microarray analysis --- Nematodes were prepared for RNA
extraction as follows. Eggs from each strain were first isolated from three large (9 cm)plates of just-starved, mixed-stage populations by alkaline hypochlorite treatment (30).
These were allowed to hatch overnight in 50 ml S-media in 250ml Erlenmeyer flasks,
at 20ºC with rotary shaking (200 rpm). The resulting synchronized L1 larvae wereconcentrated by centrifugation and pipetted onto eight large plates (subsequently
pooled to give one biological replicate), and incubated at 15º C. When most animalshad reached L4 stage, plates were shifted up to 25ºC. This temperature shift at L4
served a dual purpose: it prevented dauer formation in the daf-16(+) strains (which are
past the developmental point at which they can form dauers), and it ensured sterility inall strains (via glp-4). Worms were harvested for RNA extraction the following day (1
day old, sterile adults), by washing worms off plates with M9, followed by three
additional washes with ice-cold M9 to remove residual bacteria. At the time ofharvesting, cultures were sampled to check for the presence of dauer larvae, males,
and carcasses which might still remain from the hypochlorite treatment. Details of thisand other aspects of this work are available at AgeBase
(haldane.biol.ucl.ac.uk/publications.html). While no dauer larvae were seen in daf-
16(0) strains, a very small number were seen in one of the daf-16(+) samples. Smallnumbers of males and non-dauer larvae were observed in some samples to varying
degrees; however we did not observe any systematic contamination of particularsample genotypes. We did not observe any carcasses remaining from hypochlorite
treatment in any samples. Finally, total RNA was isolated using TRIzol reagent
(Invitrogen, www.invitrogen.com), followed by purification and concentration usingRNEasy columns according to manufacturers' instructions (Qiagen,
www1.qiagen.com). The quality of RNA samples was confirmed on an Agilent
Bioanalyser 2100 (Agilent technologies, USA; www.chem.agilent.com).Oligonucleotide microarray analysis --- Changes in transcript abundance was
measured using C. elegans whole genome oligonucleotide microarrays (Affymetrix).
We performed five biological replicates of each genotype. All Affymetrix protocolswere performed at the UCL Institute of Child Health Gene Microarray Centre. cRNA
probe generated using standard Affymetrix protocols (www.affymetrix.com).
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

7
Fragmented biotinylated probe was then hybridised to C. elegans whole genome
arrays. Washing, labelling (streptavidin-phycoerythrin) and scanning followedstandard procedures at the ICH Gene Microarray Centre.
Statistical analysis --- To calculate gene expression measures, the datasets werenormalized as follows. Raw image files were converted to probe set data (.cel files) in
M i c r o a r r a y S u i t e ( M A S 5 . 0 ) (32)
(www.stat.berkeley.edu/users/terry/zarray/Affy/GL_Workshop/genelogic2001.html).The 20 probe set data files were normalized together, and expression values were
determined, using the Robust Multi-chip Average (RMA) method (33), implementedin the Affymetrix package (version 1.4.14) of the free statistical programming
language R (www.r-project.org). Full array gene expression measures are available on
the Gene Expression Omnibus (www.ncbi.nlm.nih.gov/geo), accession numbers XX-XX). To identify differential gene expression between genotypes, we used
Significance Analysis of Microarrays (SAM), implemented in Microsoft Excel. A
delta value of 0.819 was chosen, which called 2,274 significant probe sets (1,348 up-regulated, 926 down-regulated), with a median false discovery rate (FDR) of 5.06%
(median number of falsely called probe sets = 116).To create the list of genes differentially regulated during dauer recovery, we re-
analyzed the raw data from a previous study (20). Expression differences between
time-0 and time-12 hours after dauer recovery were used to identify probes whichwere differentially expressed. Additionally, we calculated a new t test value for all
gene changes, using a more stringent two-tailed Student's t test and assuming unequalvariance. We then selected probes that showed at least a two-fold (up or down)
difference in expression, and had a p value of <0.005. This resulted in a list of 2,535
differentially regulated genes (1,160 up-regulated, 1,375 down-regulated).To insure the reliability of our data, we also performed several quality control
procedures for both microarray formats. Briefly, we verified the accuracy of themicroarray probes, and identified probes which are predicted to hybridize to more than
one gene (promiscuous probes) or to no gene (orphan probes) (For full analysis, see
haldane.biol.ucl.ac.uk). Probes which could unambiguously be assigned to a singlegene were used for EASE analysis (see below), while promiscuous probes were
analyzed by hand for any potential functional significance.
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

8
EASE analysis --- For our EASE analysis (see Results section for explanation of
the EASE application), we annotated every probe set on the C. elegans Affymetrixmicroarray and the Stanford cDNA microarray using information from a number of
different databases and datasets. We first associated each oligonucleotide probe set (orspotted array DNA probe) with its corresponding current curated gene entry. Next, the
gene entry of each probe set was used to annotate the probe set or cDNA probe with
functional information from Gene Ontology (GO) (34) (www.geneontology.org) andInterpro (35) (www.ebi.ac.uk/interpro). We also added functional information from
several previous microarray and SAGE studies (16,17,19,20), as well as the yeast two-hybrid map of C. elegans (36). For a full description of source information and the
files used in our complete EASE analysis, see haldane.biol.ucl.ac.uk.
Analysis of promoter sequences --- Genes with a given gene promoter motif in adefined region were identified using the program DNA Motif Searcher. This program
uses a set of user-supplied DNA sequence motifs to search for additional motif
instances in the genome of C. elegans. The set of motifs supplied by the user areinterpreted into a position specific score matrix (PSSM) that is used in the search step.
Using this PSSM, the program can then identify the closest matching occurrences ofthe motif based on either a score cut-off, or it can identify the top X number of
occurrences of the motif. For download or for a detailed explanation of how Motif
S e a r c h e r w o r k s , p l e a s e s e ecalliope.gs.washington.edu/papers/mcelwee2004a/software.html.
For the analyses presented here, we identified the top 10,000 occurrences of eachmotif using local background nucleotide frequencies, then filtered the hits to identify
genes which contained motif occurrences within 1,000 base pairs upstream of the
translational start site and/or within intron 1. The PSSM for each search were derivedfrom: DBE (37), DAE (17), HSE and HSAS (38), HRE (J. A. Powell-Coffman,
personal communication).Analysis of frequency of Drosophila orthologues in topomountains --- Fly gene
orthologues were identified using an all-vs.-all BLAST search for both worm and fly
using a cut-off value of 1x10-8. These results from these searches were then used toidentify reciprocal best BLAST hi ts using RBH Assigner
(calliope.gs.washington.edu/papers/mcelwee2004a/software.html). The resulting listof RBHs were used for this analysis, using a score cut-off of 100.0 or greater (2,623 of
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

9
3,609 RBHs). Using this list of RBHs, the fly orthologues were placed into
orthologous mountains. The total number of C. elegans genes in each mountain wascalculated, and the percent representation was determined as the fraction of fly genes
found in the orthologous mountain.
RESULTS
Genes that are transcriptionally regulated by the DAF-16 transcription factor must
include some that are direct determinants of lifespan. DAF-16-regulated genes havebeen identified using DNA microarray analysis (spotted arrays)(16,17). Here we have
taken the approach of comparing lists of genes whose expression is altered in daf-2
mutants, and in dauer larvae, performing a non-biased search for classes of genes thatare enriched in both cases. To this end we prepared a new list of daf-16-regulated
genes, using whole genome oligonucleotide (Affymetrix) arrays. The dauer-regulated
gene list was derived from a previous study (20).daf-2 mutants are all long-lived, but show variable degrees of pleiotropy (18).
To reduce the probability of identifying allele-specific targets unlinked to aging, weused two daf-2 alleles for the microarray analysis: daf-2(m577) (class 1) and daf-
2(e1370) (class 2, more pleiotropic). We studied age-synchronised one day old adults,
in which reproduction was blocked using the glp-4(bn2ts) mutation (29). Thistemperature-sensitive mutation blocks mitotic proliferation of the germline at 25˚C,
but is fertile at 15˚C. Nematodes were shifted from 15˚C to 25˚C at the L4 stage. In C.
elegans, an increase in lifespan results from removal of the germline, whether by
microsurgery (39), or by mutation (40). We therefore checked the effect of glp-
4(bn2ts) on lifespan in animals raised as above, in two trials. While glp-4 alone did notincrease lifespan, it causes a slight but significant enhancement of the daf-2 Age
phenotype (p < 0.05 in all cases, log rank test)(Supplemental Data Fig. 1).To identify differentially expressed genes, we pooled data for the two daf-2
alleles and compared gene expression between daf-2 and daf-16(0); daf-2 (ten
replicates each). We identified probe sets where transcript abundance is significantlydifferent between daf-2 and daf-16(0); daf-2 with a 5% false-discovery rate (FDR).
This identified 1,348 probe sets up-regulated and 926 probe sets down-regulated indaf-2 compared to daf-16; daf-2. Of these probe sets, 2,052 could be unambiguously
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

10
assigned to a single gene. For the dauer dataset, we identified differentially expressed
genes as those that showed at least a two fold (up or down) difference in expressionduring dauer recovery, with a p value of <0.005. This resulted in a list of 2,535 genes
differentially regulated probes (1,160 up-regulated, and 1,375 down-regulated). Ofthese probes, 2,348 could be unambiguously assigned to a single gene. Both gene sets
are listed at haldane.biol.ucl.ac.uk/publications.html.
Comparison with spotted array results --- We first compared our lists of daf-
16-regulated genes with lists from two previous analyses of the effects of reduced IIS
on gene transcription in C. elegans (16,17). In the two earlier studies DNAmicroarrays spotted with PCR-amplified DNA were employed. In principal the use of
oligonucleotide microarrays is more sensitive, since they employ multiple
oligonucleotides designed from each gene to be unique in the genome, thus greatlyreducing the problem of cross hybridisation between closely related genes. Moreover,
there appears to be significantly lower variance among replicates when using
oligonucleotide arrays (Affymetrix) compared to spotted arrays (Thomas E. Johnson et
al., personal communication). Although there is considerable overlap between genes
identified in the three analyses (Fig. 1A), a high proportion of genes are uniquelyidentified in each study.
Comparison of daf-2 and dauer-regulated genes with other gene sets --- We
compared changes in gene expression between daf-2 vs. daf-16; daf-2 (the daf-2 geneset) with gene expression differences between dauer larvae and larvae 12 hr after the
onset of dauer recovery (the dauer gene set)(20). Despite the difference in themethods used to generate the dauer and daf-2 gene sets, an overlap between gene sets
is evident (Fig. 1B). For example, 21% of genes up-regulated in dauers are also up-
regulated in daf-2 adults (219 genes), and 11% of genes down-regulated in dauers arealso down-regulated in daf-2 adults (145 genes). It is probable that genetic
determinants of longevity and aging are enriched among these 364 overlapping genes.The relatively large-number of changes in both dauer and daf-2 gene sets
means that identifying biologically relevant changes on a gene-by-gene basis would
likely prove very difficult, if not impossible. To circumvent this problem and identifypotentially important biochemical processes in a statistically rigorous way, we have
made use of the freely-avai lable software package EASE(david.niaid.nih.gov/david/ease.htm)(41). EASE is a user-customizable application
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

11
which performs a statistical analysis to identify significant over-representation of
functional gene classifications within gene lists. Essentially, it compares theprevalence of particular gene categories within our gene lists (for instance, cytochrome
P450 genes), to the prevalence of that category in the genome as a whole. Iftranscriptional up or down-regulation of a particular gene category or biological
process is important for dauer or daf-2 phenotypes, we would expect to find it over-
represented within our lists of differentially expressed genes.We have used EASE to examine our lists of differentially-expressed genes for
over-representation of functional categories from a number of biological databases,including Gene Ontology (GO) and Interpro. Additionally, we have created several
customized databases which contain microarray data and functional classifications
from previously published studies, allowing us to compare our gene lists to otherbiological conditions which may show similar patterns of gene expression.
Using EASE, we have identified many gene classes which show significant
over-representation in the dauer and daf-2 gene sets. We first used EASE to examinethe dauer and daf-2 gene sets separately, and identified gene groups which were over-
represented (p < 0.1) in either the up-regulated or down-regulated genes. We thencombined these separate analyses, and identified gene groups that were similarly over-
represented in both dauer and daf-2 gene sets.
This initially identified 35 gene classes over-represented among genes up-regulated in dauer and daf-2 gene sets, and 26 gene classes over-represented among
genes down-regulated in dauer and daf-2 gene sets. Initial examination of these listsrevealed that a number of lists contained identical or near identical members. Where
redundant lists were identified, one representative member was selected for further
analysis, and the rest set aside. Excluding redundant lists, there remained 24 geneclasses over-represented among genes up-regulated in dauer and daf-2 gene sets, and
23 gene classes over-represented among genes down-regulated in dauer and daf-2
gene sets (Table I). Gene classes showing over-representation in daf-2 mutants alone
(p < 0.001) are shown in Table II.
The dauer gene set was highly represented in the daf-2 gene set: genes up-regulated in dauers were strongly over-represented among genes up-regulated in daf-2
mutants (p = 4.33e-26), and genes down-regulated in dauers were over-representedamong genes down-regulated in daf-2 (p = 6.19e-08). There was also a weaker overlap
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

12
between genes up-regulated in daf-2 mutants and SAGE dauer tags (Table I). An
earlier study of IIS-regulated genes identified sets of genes either up-regulated (class1) or down-regulated (class 2) in long-lived IIS mutants (17). In that study, RNA
interference tests confirmed the expectation that, overall, class 1 genes promotelongevity, while class 2 genes promote aging. We observed a large overlap between
class 1 genes and genes up-regulated in dauers, and class 2 genes and genes down-
regulated in dauers (Table I). Thus, a strong dauer transcriptional signature is presentin daf-2 mutant adults. This is consistent with action of the same genes to control
longevity in dauer larvae and daf-2 mutant adults.A number of gene expression topomountains were associated with dauer and
daf-2 gene sets. Mounts 6, 15 and 21 were over-represented in up-regulated dauer/daf-
2 gene sets, and are therefore potentially longevity associated. Mounts 19, 20, 24, 27,30 and 31 were over-represented in down-regulated dauer/daf-2 gene sets, and are
therefore potentially aging associated. Mt. 8 was over-represented in both up and
down-regulated gene sets.All genes in Mt. 15 are up-regulated in dauer larvae (20). Interestingly, the
overlap between daf-2 up-regulated genes and this 'dauer mountain' was the mostsignificant of all the topomountains (p = 6.16e-54), further evidence of a dauer
transcriptional signature in daf-2 adults. Mt. 15 is discussed in more detail below. (See
Supplemental Data for further discussioon of gene expression topomountains).The gene classes identified (Table I) provide indicators of varying informative
value of processes altered in both dauer larvae and daf-2 mutants. To identify suchprocesses, we studied the contents of these gene lists, focusing on the individual genes
within each gene category that were represented in both dauer and daf-2 gene groups.
Full lists of genes within each gene class that are up or down regulated in dauer larvae,in daf-2 mutants, or in both contexts are available at haldane.biol.ucl.ac.uk.
Up-regulated gene groups --- Among these gene groups is IPR002068: Heatshock protein Hsp20. These are the small heat shock protein (smHSP)/α-crystallin
gene family of molecular chaperones. Six genes of this class are up-regulated in dauers
and/or daf-2 mutants (three in both). The six genes span the full size range of smHSPs:
there are two hsp-12's (hsp-12.3, hsp-12.6), two hsp-16's (F08H9.3, F08H9.4), sip-1
(stress induced protein, F43D9.4, an Hsp20), and hsp-43 (C14F11.5). The smHSPs are
the only gene class where a role in longevity assurance in C. elegans is well
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

13
demonstrated. Over-expression of hsp-16.1 and hsp-16.48 increases stress resistance
and lifespan (42). Expression of smHSPs and other HSPs is regulated by the hsf-1
(heat shock factor) transcription factor. hsf-1 expression is required for the daf-2 Age
phenotype, and over-expression increases longevity (27), while loss of hsf-1 results inprogeria (43). RNAi studies demonstrate that smHSPs are important for the Age
phenotypes resulting from either mutation of daf-2 or over-expression of hsf-1 (27).
Thus, the presence of the smHSP gene class in Table I demonstrates that our analysisis capable of identifying gene classes linked to longevity. This suggests that other gene
groups represented may also be linked to longevity.Three other gene classes in Table I are potentially linked to stress resistance: the
cytochrome P450's (CYPs), the short-chain dehydrogenase/reductases (SDRs) and the
UDP-glucuronosyltransferases (UGTs). There are 77 genes in the C. elegans genomethat encode putative CYP proteins (correctly, heme thiolate protein P450's)(44). This
number is typical of metazoan species (45); by contrast the S. cerevisiae genome
contains only three. In humans, many CYPs are monooxygenases (mixed functionoxidases), catalysing the reaction
RH + O2 + 2e- + 2H+ + NADPH or NADH -> ROH + H2OThey metabolise a range of endobiotic lipophilic substrates, including steroids, fatty
acids and prostaglandins, and lipophilic xenobiotics (46). In C. elegans, the DAF-9
CYP is believed to metabolise a steroid hormone critical to the regulation of dauerlarva formation, and with some influence on lifespan (47,48). Six genes encoding
CYPs showed increased expression in both dauer larvae and daf-2 adults, twelve indaf-2 mutants alone, and three in dauers alone. In 4/12 of the CYP genes up-regulated
only in daf-2 mutants, there is also a trend towards increased expression in dauer
larvae (p < 0.05). Expression changes in all 77 CYP genes are summarised inSupplemental Data Table II.
What biochemical and cellular processes might these CYPs be influencing?Unfortunately, little is known about the biochemical function of most C. elegans
CYPs. The possible role of CYP genes in xenobiotic detoxification has been examined
by screening for induction of transcription in response to a range of xenobioticcompounds (44). 15 CYP genes showed some degree of induction, four of which are
among the dauer and/or daf-2 up-regulated genes. These are B0213.15 (CYP34A9),which is up-regulated in daf-2 mutants and dauer larvae, and C49C8.4 (CYP33E1),
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

14
K07C6.3 (CYP35B2) and K07C6.4 (CYP35B1) which are up-regulated in daf-2
mutants only. Expression of 35B1 and 35B2 was induced by a range of xenobiotics;for example, both were induced by ethanol (44). Notably, K07C6.4 (CYP35B1) was
the most strongly up-regulated CYP gene in daf-2 mutants (9.4-fold increase).Interestingly, RNAi of this gene leads to partial suppression of the daf-2 Age
phenotype (17); this is also the case for B0213.15 (34A9)(xenobiotic induced) and
T10B9.1 (13A4)(not xenobiotic induced). This suggests the possibility that processesthat assure xenobiotic resistance also promote longevity. Increased expression of
CYPs accounts for the presence of the GO:0006118 electron transport gene category.29 genes encode putative short-chain dehydrogenase/reductases are up-regulated
in dauers and/or daf-2 adults (6 in both). SDRs function in xenobiotic detoxification,
typically catalysing the reduction of carbonyl groups in aldehydes and ketones,consuming energy from NADH or NADH in the process (49,50).
A total of 22 genes encoding predicted UDP-glucuronosyltransferases (UGTs)
are up-regulated in dauers and/or daf-2 adults (six in both). UGTs act in the smoothendoplasmic reticulum to glucuronidate (or glucosylate in insects)(51) small lipophilic
molecules, thereby rendering them water soluble and enabling their excretion (52).The substrates for UGTs are numerous, including diverse dietary constituents, such as
fatty acids and steroids, as well as xenobiotics and unwanted endobiotics (53). In
mammals, this occurs mainly in the liver. Up-regulation of UGT expression haspreviously been noted in daf-2 mutants (17) and dauer larvae, where it has been
suggested that they might affect longevity via xenobiotic detoxification, or affectdauer formation by glucuronidating a dauer steroid hormone (20).
A total of 29 genes encoding transthyretin-like proteins are up-regulated in daf-2
adults and/or dauer larvae (four in both). In vertebrates, transthyretin (TTR) is atransport protein found in extracellular fluids which carries both thyroid hormones as
well as vitamin A complexed with retinol binding protein. However, the C. elegans
transthyretin-like proteins that have been studied are members of a related but distinct
protein family, the transthyretin-related proteins (TRP)(54). TRPs are found in
vertebrates, invertebrates, prokaryotes and plants. While their function is unknown, thepredicted ligand-binding site is almost entirely conserved, suggesting that, like TTR,
they may function as carriers of lipophilic moieties.
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

15
A further clue as to TRP function is that all four TRP genes up-regulated in both
dauers and daf-2 mutants (C40H1.5, H14N18.3, Y51A2D.10, Y51A2D.11) are in Mt.8, which is associated with intestinal function, and enriched for UGTs. Most TRPs are
cytoplasmic (54); we tested for the presence of a predicted signal peptide (signifyingprobable secretion) using the signal V2.0 program (www.cbs.dtu.dk/services/SignalP-
2.0/)(55). All four overlapping TRP genes are predicted to be secreted. We postulate
that these TRPs play an active role in transporting unknown lipophilic substances fromthe intestine. None of these genes have close homologues outside the Nematoda. For
commentary on other up-regulated gene classes, and on down-regulated gene classes,see Supplemental Data.
While studying these gene lists, we noticed several individual genes that were
up-regulated in daf-2 mutants that are worthy of note. For example, there was a 2.8-fold increase in transcript levels of C50F7.10, one of two C. elegans putative glycosyl
hydrolases, homologous to mammalian klotho. Loss of function of this gene results in
a progeroid phenotype in mice (56). Expression of ges-1 was also increased (3.7-fold).The ges-1 promoter has been used in a number of previous studies to force expression
of transgenes in the nematode intestine, for example, of daf-16(+) in a daf-16; daf-2
mutant background (57). Potentially, this could have resulted in a positive feedback
loop of daf-16 expression.
Gene groups altered in daf-2 mutants exclusively --- A number of gene groupsoverlapped only with the daf-2 gene group (p < 0.001, Table II). Gene groups altered
in daf-2 mutants but not dauers might reflect lifespan determinants unique to adults.After removal of redundant groups, there were four groups up-regulated only in daf-2
mutants, and eight groups down-regulated.
In daf-2 adults, there was up-regulation of genes in Mts. 1 (muscle, neuronal),17 and 22 (collagen). The most significant overlap was with Mt. 17. This
topomountain is enriched with collagen genes (28); however, only 3/40 Mt. 17 genesup-regulated in daf-2 encode collagens. There was also up-regulation of 16 glutathione
S-transferases (GSTs)(Table II). These proteins act in the cytosol, catalysing the
addition of the tripeptide glutathione to endogenous and xenobiotic electrophilicsubstrates, effecting the detoxification and metabolism of a range of compounds. The
resulting glutathione adducts are more water soluble. In this respect their function issimilar to that of UGTs. GSTs also act in detoxification through their peroxidase
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

16
activity, and by passive binding of toxins. Additionally, GSTs play a role in oxidative
stress resistance. GST-p24 (K08F4.7) is up-regulated in response to oxidative stress(58), and over-expression of K08F4.7 increases resistance to oxidative stress (59). We
find K08F4.7 transcript levels to be increased in daf-2 mutants (3.5-fold), but not indauer larvae. GSTs also play a role in transport of specific hydrophobic ligands.
In daf-2 adults there was down-regulation of genes associated with Mt. 21
(lipid metabolism) and Mt. 22 (collagen), as previously observed (16). There was alsodown-regulation of UGTs; thus there is over-representation of this gene class both in
genes up-regulated and down-regulated in daf-2 mutants. There was also down-regulation of two overlapping gene groups linked to transport (IPR005828: General
substrate transporter; GO:0006810: Transport). One possible interpretation is that
nutrient uptake is reduced in daf-2 mutants. This is consistent with the down-regulation of several gene groups linked to protein degradation in dauers and daf-2
mutants (Table I).
Potential concerted regulation of highly orthologous gene groups --- BecauseEASE analysis requires a one-to-one correspondence between microarray probes and
gene targets, it cannot be used to analyse probes that are predicted to hybridize tomultiple genes. While both the spotted array and oligonucleotide arrays described
here were designed to reduce promiscuity, a small percentage (~7-10%) of the
differentially regulated probes for both arrays cannot be unambiguously assigned to asingle gene. There are at least two reasons for this: a probe may be poorly designed
and cross-hybridize to unrelated genes, or the probe may be specific for a gene familyin which members are highly homologous. In the latter case, these probes can be used
to assay the expression of homologous gene families as a whole. We find that two
such gene families, transposases and histones, are potentially differentially expressedin both dauers and daf-2 adults.
A large number of promiscuous probes for transposase genes on both arrays arehighly up-regulated (11 probes in the dauer dataset, and 13 probe sets in the daf-2
dataset). This may indicate general activation of transposition within the C. elegans
genome in dauers and daf-2 adults. While increased transposition would not beexpected to result in increased longevity, there are two possible explanations for this
observation. First, the chromatin architecture in long-lived worms may differ fromthat of normal-lived worms. In addition to activated transposases, we also find a large
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

17
number of differentially expressed promiscuous probes for histone genes, another
highly homologous gene family. In daf-2 adults, 7 probes for histone H2 familymembers are down-regulated, and in both dauers and daf-2 adults, probes for histone
H4 are up-regulated (1 in daf-2, 5 in dauers). Notably, increased activity of sir-2.1, aputative histone deacetylase, increases lifespan in C. elegans in a daf-16-dependent
fashion (60). Additionally, daf-16-dependent alterations in chromatin content has been
observed previously in dauers, caloric restriction, and IIS mutants (O. Ilkaeva, C.-K.Ea and J. A. Waddle, personal communication), and it may be a that altered chromatin
packing in these conditions leads to a permissive environment for transposaseexpression. The second possible explanation for the increased expression of
transposase genes is that both dauers and daf-2 adults show increased levels of heat-
shock gene induction, and we find that the promoters of transposase genes are highlyenriched for heat-shock elements (described below).
Gene regulatory elements in daf-2- and dauer-regulated genes --- We have
shown the presence of a dauer transcriptional signature in daf-2 mutant adults. Thisindicates that similar transcriptional control factors are at work in each milieu, perhaps
involving specific gene promoter elements. To explore this, we examined thefrequency in daf-2- and dauer-regulated genes of five promoter elements potentially
associated with IIS: the DAF-16-binding element (DBE)(37), the daf-16-associated
element (DAE)(17), the heat-shock response element (HSE), the heat-shock associatedsequence (HSAS)(38), and a potential hypoxia induction factor (hif-1) response
element (HRE)(J.A. Powell-Coffman, personal communication).To do this, we first identified genes which contain these regulatory elements
using DNAmotif Searcher which uses a position-specific score matrix (PSSM) to
identify potential regulatory motifs throughout the genome. These can then bematched against predicted coding regions to identify motifs within specified regulatory
regions (such as promoters). Using this approach, we have identified regulatory motifsin promoter regions (0-1000 bp upstream of the predicted translational start sites) as
well as in the first intron, which in C. elegans often contain regulatory sequences (for
full lists of genes and PSSM matrices, see haldane.biol.ucl.ac.uk). Using these lists ofgenes containing particular regulatory motifs, we again used EASE to identify
significantly over-represented functional categories which these transcription factorsmay regulate.
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

18
The DAF-16 transcription factor binds to the DAF-16 binding element (DBE),
identified from 29 random oligonucleotides which bound to DAF-16 protein (37).These defined a core sequence, TTGTTTAC, which is common to all forkhead
binding sites (61,62). DBE-containing gene groups which overlap with the daf-2 geneset are listed in Tables I and II (p < 0.05). The 14 gene classes with a strong overlap (p
< 0.01) with the DBE gene set are listed in Supplemental Data Table II. These include
genes up-regulated in daf-2 mutants and dauers, plus two other genes groups that areenriched among genes up-regulated in dauers and daf-2 mutants, including Mt. 15
('mount dauer'). This provides strong evidence for the importance of the DBE in DAF-16-regulated gene expression, and suggests that these gene classes are directly
regulated by DAF-16. Some of the remaining gene groups may be over-represented
due to possession of binding sites for other classes of forkhead transcription factor.Interestingly, none of the groups were those identified as enriched with genes down-
regulated in dauers or daf-2 mutants. This implies that DBEs are more important for
DAF-16-dependent up-regulation of gene expression. Thus, the DBE appears to be alongevity-associated promoter element.
The daf-16-associated element (DAE)(CTTATCA) was identified from acomparison of promoter sequences of daf-2-regulated genes (17). This sequence
resembles the reverse complementary GATA binding element (TTATCAGT). Of 29
gene classes strongly enriched (p < 0.01) for DAE-containing genes, 14 were alsoenriched for genes differentially expressed in dauers and/or daf-2 mutants
(Supplemental Data Table II). Interestingly, 13/14 classes show some down-regulation. These include Mts. 8 and 21, which are enriched with both daf-2 up-
regulated and daf-2 down-regulated genes (Table I, II). Moreover, the sets of genes
down-regulated in daf-2 mutants are themselves enriched for DAE-containing genes.These results strongly imply that presence of the DAE leads to DAF-16-dependent
down-regulation of transcription. Thus, DAE appears to be an aging-associatedpromoter element.
The heat-shock response element (HSE)(TTCTAGAA), and the heat-shock
associated sequence (HSAS)(GGGTGTC) are both associated with increasedexpression of chaperonin genes in response to heat shock (38). daf-16-dependent
increases in expression of heat shock proteins are important for the IIS-associated Agephenotype (27,42). This suggests that these promoter elements might be important for
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

19
the transcriptional response to altered IIS. Several dauer/daf-2 associated gene groups,
all of them up-regulated, did prove to be enriched for these elements (Table I , II,Supplemental Data Table I). Mt. 6 (neuronal gene enriched) was enriched for both
HSE and HSAS containing genes. The most strongly HSE gene-enriched group wasIPR001888: Transposase, type 1. This suggests that transposons might be activated by
heat shock; the presence of an HSE was previously noted in the Tc2 transposase gene
(63).hif-1 regulates the transcriptional response to oxygen starvation (hypoxia). A
sequence resembling the mammalian hif-1-response element (HRE) was recentlyidentified as an enriched motif among the promoter regions of C. elegans genes whose
expression is up-regulated by C. elegans HIF-1 (J.A. Powell-Coffman, personal
communication). Since daf-2 mutants are resistant to hypoxia (64), we predicted thatgenes with this HRE-like sequence might be regulated by daf-2. We detected over-
representation of HRE genes among genes up-regulated in dauers but not in daf-2
mutants. Mt. 6 (dauer/daf-2 up-regulated) was also enriched with HRE genes. Therewas also a slight enrichment of hif-1 genes in daf-2 down-regulated genes. Potentially,
the HIF-1 and DAF-16 proteins act together on genes in these clusters to effect thehypoxia resistance of DAF-2 mutants (64). In conclusion, this analysis shows that
overall the DBE, HSE, HSAS and HRE elements are enriched in genes up-regulated in
dauers and/or daf-2 mutants, and therefore represent potential longevity-associatedpromoter elements. By contrast, the DAE is associated with down-regulated genes,
and therefore represents a potential aging-associated promoter element.Poor evolutionary conservation of Mt. 15 --- Several observations suggest a
possible link between Mt. 15 and the longevity of dauer larvae and daf-2 adults.
Firstly, all genes in Mt. 15 are up-regulated in dauer larvae (20). Secondly, it is by along shot the topomountain with the strongest over-representation of genes up-
regulated in daf-2 adults (p = 6.16e-54); the next most over-represented is Mt. 08(9.19e-35). Thirdly, it is the only dauer/daf-2 regulated topomountain that shows
enrichment of DBE elements (Supplemental Data Table I). Analysis of microarray
data from orthologous genes in a range of species has demonstrated evolutionaryconservation of a number of topomountains (65). To explore the possibility that Mt. 15
might show evolutionary conservation of genes associated with longevity assurance,
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

20
we compared the frequency in each C. elegans topomountain of genes with
orthologues in Drosophila.The topomountains over-represented in daf-2 mutant-associated gene lists (p <
0.001), or both daf-2 mutants and dauers (p < 0.1) are listed in Supplemental DataTable III. Percent orthology varied among topomountains from 0-88%, mean ~18%.
With the exception of Mt. 1, all of the potential longevity-associated mountains
showed lower than average orthology, with Mt. 15 showing the lowest value (6.1%).Thus, evolutionary conservation of Mt. 15 is unlikely. Overall, the aging-associated
mountains showed a higher degree of orthology: mean, 31%, compared to 9% for thelongevity-associated mountains. This implies that there is better evolutionary
conservation among genes that promote aging than among those that promote
longevity.
DISCUSSION
Microarray analysis has shown that large numbers of genes show differential transcript
abundance in dauers (20) and daf-2 mutant adults (16,17)(this study). Both dauers anddaf-2 mutant adults exhibit increased longevity. Thus, by comparing the transcription
profiles of dauers and daf-2 mutants we have identified a number of gene groups and
processes potentially contributing to longevity and aging.A dauer transcriptional signature in daf-2 adults --- We observed over-
representation of dauer up-regulated genes among genes up-regulated in daf-2 mutantadults, and of dauer down-regulated genes among genes down-regulated in daf-2
mutants (Table I). This provides support for the idea that daf-2 mutant longevity
reflects expression of a dauer longevity program in the adult (2). It also provides somevalidation for our daf-2 mutant/dauer comparative approach. Further, the up-regulation
in daf-2 mutants and dauers of small heat shock proteins, which have a demonstratedrole in longevity assurance, demonstrates that our analysis is capable of identifying
lifespan-associated gene groups.
Increased expression of detoxification genes in daf-2 adults and dauer larvae ---
We observed up-regulation of genes encoding cytochrome P450’s (CYPs), short-chain
dehydrogenase/reductases (SDRs) and UDP-glucuronosyltransferases (UGTs) in daf-2
adults and dauer larvae (Table I). In mammals these three enzyme classes act in
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

21
concert to dispose of toxic endobiotic or xenobiotic compounds (e.g. toxins, drugs,
carcinogens). Moreover, in daf-2 adults (but not dauer larvae) we saw up-regulation ofa fourth gene group linked to detoxification, the glutathione S-transferases
(GSTs)(Table II).In pharmacological parlance, drug metabolism entails two successive phases:
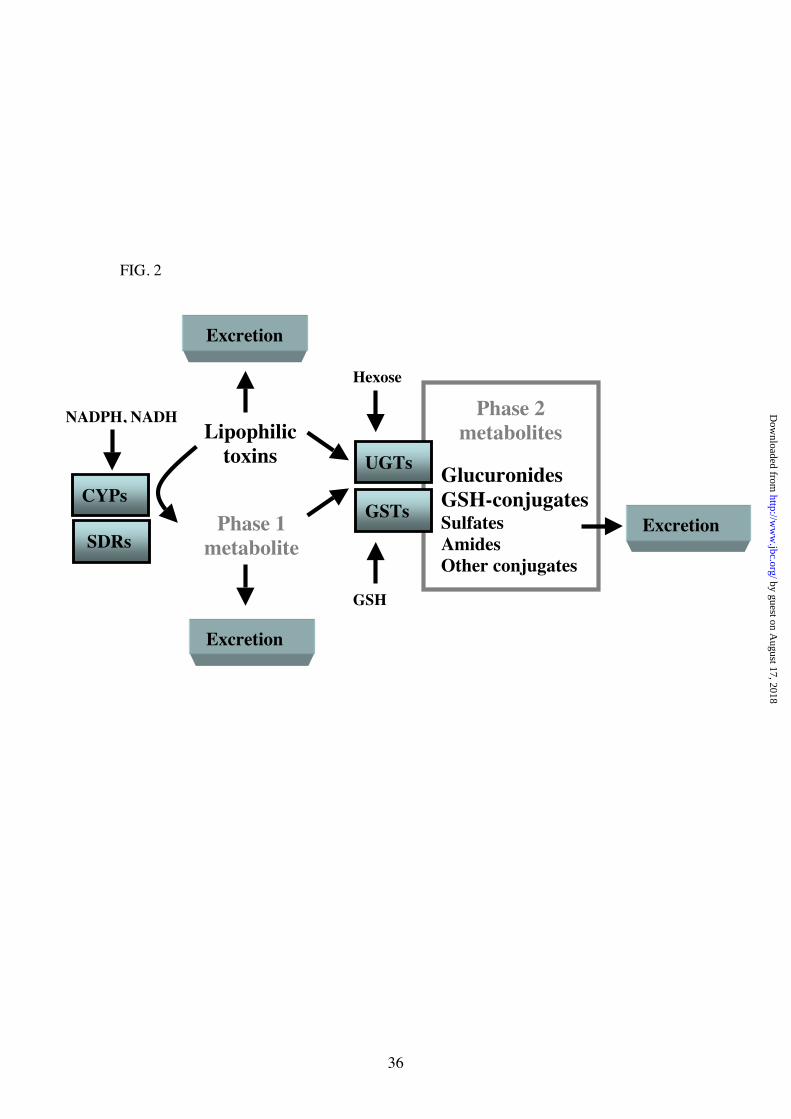
phase 1 (functionalisation reactions), and phase 2 (conjugative reactions)(Fig. 2)(51).
Phase 1 reactions result in chemically reactive functional groups, which are often (butnot always) required for phase 2 reactions, the addition of side groups which increase
solubility, aiding excretion. Phase 2 reactions represent the actual detoxificationreactions.
Both CYPs and SDRs play a role in phase 1 metabolism in mammals. CYPs are
the major effectors, acting mainly as mixed function oxidases, bioactivatingxenobiotics or stable metabolites, usually by hydroxylation. The major effectors of
phase 2 metabolism are the UGTs, but GSTs, sulphotransferases and acetyltransferases
are also important (51). Both CYPs and UGTs act in the smooth endoplasmicreticulum (ER), while GSTs act in the cytosol, and SDRs in both locations. Taken
alone, the significance of the up-regulation of many CYP genes in dauers and daf-2
mutants is difficult to interpret, since CYPs function not only in drug metabolism but
also in a range of biosynthetic processes (e.g. of steroids, fatty acids, prostaglandins,
vitamin D). Yet several observations suggest that CYP action in phase 1 metabolism isactivated in dauers/daf-2 mutants. Firstly, the concurrent up-regulation of SDRs,
UGTs and GSTs. Secondly, a number of the up-regulated CYP genes have previouslybeen shown to be up-regulated in response to xenobiotics (44)(Supplemental Data
Table II).
A new theory of aging --- The up-regulation of these four gene groups stronglyimplies that the capacity for phase 1 and phase 2 metabolism is up-regulated in daf-2
mutants and dauer larvae. In the case of three CYPs and one UGT, a role in longevityassurance has been demonstrated by RNAi (17). These findings suggest a new picture
of the biology of aging: that a major cause the molecular damage that gives rise to
aging is a wide range of toxic compounds (endobiotic and perhaps also xenobiotic),particularly those that are lipophilic. We suggest that the enzymes involved in phase 1
and phase 2 detoxification metabolism are a major mechanism of longevity assurance(Fig. 3).
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

22
This theory provides a possible explanation for a number of other gene groups
found in this study to be linked to daf-2 mutant and dauer states. For example, thegene class IPR000379: Esterase/lipase/thioesterase: a range of esterases play a role in
reductive drug metabolism, e.g. the hydrolysis of the anaesthetic procaine by plasmaesterase (51). Likewise, the TRPs seem likely to be carriers of lipophilic compounds:
possibly they also play a role in disposal of toxic lipophilic moieties. An alternative
possibility is that TRPs act as carriers for lipophilic hormones involved in regulationof dauer formation and lifespan, for example the predicted steroid hormone associated
with the DAF-9 CYP (47,48).Our analysis detected GST up-regulation, and down-regulation of UGTs in daf-2
mutants but not dauers. This could reflect differences in the targets of detoxification in
each milieu: more lipophilic moieties in dauers, and more electrophilic ones (perhapsxenobiotic) in adults. Such differences in gene expression may reflect differences in
gene regulation by signaling pathways, such as IIS; alternatively, they may reflect
differential induction of gene expression due to the presence of different detoxificationtarget molecules.
Detoxification is energetically costly: links to the disposable soma theory of
aging --- The potential role of phase 1 and phase 2 metabolism in longevity assurance
may be linked to the evolutionary theory of aging. According to Kirkwood's
disposable soma theory, natural selection does not usually favor non-aging organismsbecause somatic maintenance processes that assure longevity are energetically costly
(66). A notable feature of both phase 1 and phase 2 metabolism of toxins is that theyare energetically costly (in contrast to the action of SOD and catalase). The mixed
function oxidase reactions of phase 1 metabolism, catalysed by CYPs, consume
NADPH or NADH, as do SDR reactions. Similarly, each UGT glucuronidationreaction consumes a molecule of glucose (an extraordinary expense), and
glutathionylation by GSTs a molecule of glutathione. Potentially, increased phase 1and 2 metabolism in daf-2 mutant adults consume energy that could otherwise be
expended on reproduction, thus reducing fitness.
Insulin/IGF-1 signaling regulates genes linked to nutrient uptake --- A numberof gene classes linked to transport are down-regulated in dauers and daf-2 mutant
adults (Table I). These include the gene nhx-2 (B0495.4), which is expressed in theapical membranes of intestinal cells, and appears to prevent intracellular acidification
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

23
by catalysing the electroneutral exchange of extracellular sodium for an intracellular
proton. Such acidification of intestinal cells occurs as the result of proton gradient-driven uptake of nutrients, e.g. by the oligopeptide transporter PEP-2 (OPT-
2)(KO4E7.2)(67). Interestingly, reduced expression of nhx-2, slows development andextends lifespan in C. elegans, probably by inducing dietary restriction (67). Deletion
of the pep-2 gene does not increase lifespan per se, but enhances the daf-2(e1370) Age
phenotype (68). We find that in daf-2 mutants, nhx-2 and pep-2 show 5.6- and 7.7-foldreductions in transcript levels, respectively. Reduced expression of pep-2 with reduced
IIS was previously noted (17). In dauer larvae nhx-2 and pep-2 also show reducedtranscript abundance, by 2.0- and 3.7-fold, respectively. This suggests that dietary
restriction due to reduced nutrient uptake in the intestine contributes to the daf-2 Age
phenotype.In conclusion: the aim of this work was to generate new ideas about
mechanisms of aging from microarray data, using a novel approach: comparison of
transcript profiles of dauer larvae and daf-2 mutants. This approach was successful, inthat it generated a number of new hypotheses about classes of genes and biochemical
processes that determine lifespan, for example, the determinative role in lifespan of thedetoxification system. Whether or not this theory is true is currently being tested in our
laboratory.
ACKNOWLEDGEMENTS
We thank Jo Anne Powell-Coffman for providing the hif-1 response element sequence,
Thomas E. Johnson and James Waddle for communication of unpublished results, and
Linda Partridge, Matt Piper and members of the Gems lab and Wellcome TrustFGAA-funded partner labs for useful discussion. Some strains were provided by the
Caenorhabditis Genetics Center, which is funded by the National Institutes of HealthNational Center for Research Resources. This work was supported by grants from the
Wellcome Trust (Functional Genomic Analysis of Ageing) to J. J. M., E. S., E. B. and
D. G. and from the Royal Society to D. G.
REFERENCES
1. Friedman, D. B., and Johnson, T. E. (1988) Genetics 118, 75-86
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

24
2. Kenyon, C., Chang, J., Gensch, E., Rudener, A., and Tabtiang, R. (1993)Nature 366, 461-464
3. Kimura, K. D., Tissenbaum, H. A., Liu, Y., and Ruvkun, G. (1997) Science277, 942-946
4. Clancy, D., Gems, D., Harshman, L. G., Oldham, S., Hafen, E., Leevers, S. J.,and Partridge, L. (2001) Science 292, 104-106
5. Tatar, M., Kopelman, A., Epstein, D., Tu, M.-P., Yin, C.-M., and Garofalo, R.S. (2001) Science 292, 107-110
6. Holzenberger, M., Dupont, J., Ducos, B., Leneuve, P., Geloen, A., Even, P.,Cervera, P., and Le Bouc, Y. (2003) Nature 421, 182-187
7. Bluher, M., Kahn, B., and Kahn, C. (2003) Science 299, 572-5748. Riddle, D. L., and Albert, P. S. (1997) in C. elegans II (Riddle, D. L.,
Blumenthal, T., Meyer, B. J., and Priess, J. R., eds), pp. 739-768, Cold SpringHarbor Laboratory Press, Plainview, N.Y.
9. Cassada, R. C., and Russell, R. L. (1975) Dev. Biol. 46, 326-34210. Klass, M. R., and Hirsh, D. I. (1976) Nature 260, 523-52511. Morris, J. Z., Tissenbaum, H. A., and Ruvkun, G. (1996) Nature 382, 536-53812. Anderson, G. L. (1982) Can. J. Zool. 60, 288-29113. Vanfleteren, J. R. (1993) Biochem. J. 292, 605-60814. Vanfleteren, J. R., and De Vreese, A. (1995) FASEB J. 9, 1355-136115. Honda, Y., and Honda, S. (1999) FASEB J. 13, 1385-139316. McElwee, J., Bubb, K., and Thomas, J. (2003) Aging Cell 2, 111-12117. Murphy, C. T., McCarroll, S. A., Bargmann, C. I., Fraser, A., Kamath, R. S.,
Ahringer, J., Li, H., and Kenyon, C. J. (2003) Nature 424, 277-28418. Gems, D., Sutton, A. J., Sundermeyer, M. L., Larson, P. L., Albert, P. S.,
King, K. V., Edgley, M., and Riddle, D. L. (1998) Genetics 150, 129-15519. Jones, S., Riddle, D., Pouzyrev, A., Velculescu, V., Hillier, L., Eddy, S.,
Stricklin, S., Baillie, D., Waterston, R., and Marra, M. (2001) Genome Res.11, 1346-1352
20. Wang, J., and Kim, S. (2003) Development 130, 1621-163421. Dorman, J. B., Albinder, B., Shroyer, T., and Kenyon, C. (1995) Genetics 141,
1399-140622. Larsen, P. L., Albert, P. S., and Riddle, D. L. (1995) Genetics 139, 1567-158323. Ogg, S., Paradis, S., Gottlieb, S., Patterson, G. I., Lee, L., Tissenbaum, H. A.,
and Ruvkun, G. (1997) Nature 389, 994-99924. Lin, K., Dorman, J. B., Rodan, A., and Kenyon, C. (1997) Science 278, 1319-
132225. Larsen, P. L. (1993) Proc. Natl. Acad. Sci. USA 90, 8905-890926. Walker, G., White, T., McColl, G., Jenkins, N., Babich, S., Candido, E.,
Johnson, T., and Lithgow, G. (2001) J Gerontol A Biol Sci Med Sci. 56, B281-B287
27. Hsu, A., Murphy, C., and Kenyon, C. (2003) Science 300, 1142-114528. Kim, S., Lund, J., Kiraly, M., Duke, K., Jiang, M., Stuart, J., Eizinger, A.,
Wylie, B., and Davidson, G. (2001) Science 293, 2087-209229. Beanan, M., and Strome, S. (1992) Development 116, 755-76630. Sulston, J., and Hodgkin, J. (1988) in The nematode Caenorhabditis elegans
(Wood, W. B., ed), 0 Ed., pp. 587-606, Cold Spring Harbor, N.Y.31. Lewis, J., and Fleming, J. (1995) in Caenorhabditis elegans: Modern
Biological Analysis of an Organism (Epstein, H., and Shakes, D., eds) Vol. 48,pp. 4-29, Academic Press, San Diego
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

25
32. Hubbell, E. (2001) in Gene Logic Workshop on Low Level Analysis ofAffymetrixGeneChip data
33. Irizarry, R., Hobbs, B., Collin, F., Beazer-Barclay, Y., Antonellis, K., Scherf,U., and Speed, T. (2003) Biostatistics 4, 249-264
34. Ashburner, M., Ball, CA, Blake, JA, Botstein, D, Butler, H, Cherry, JM,Davis, AP, , and Dolinski K, D. S., Eppig JT, Harris MA, Hill DP, Issel-Tarver L, Kasarskis A, Lewis S, Matese JC, Richardson JE, Ringwald M,Rubin GM, Sherlock G. (2000) Nat Genet 25, 25-29
35. Mulder, N., Apweiler, R., Attwood, T., Bairoch, A., Barrell, D., Bateman, A.,Binns, D., Biswas, M., Bradley, P., Bork, P., Bucher, P., Copley, R.,Courcelle, E., Das, U., Durbin, R., Falquet, L., Fleischmann, W., Griffiths-Jones, S., Haft, D., Harte, N., Hulo, N., Kahn, D., Kanapin, A., Krestyaninova,M., Lopez, R., Letunic, I., Lonsdale, D., Silventoinen, V., Orchard, S., Pagni,M., Peyruc, D., Ponting, C., Selengut, J., Servant, F., Sigrist, C., Vaughan, R.,and Zdobnov, E. (2003) Nucleic Acids Res. 31, 315-318
36. Li, S., Armstrong, C., Bertin, N., Ge, H., Milstein, S., Boxem, M., Vidalain,P., Han, J., Chesneau, A., Hao T, DS, G., Li N, M. M., Rual JF, Lamesch P,Xu L, Tewari M, Wong SL, Zhang LV, Berriz GF, Jacotot L, V. P., J, R., andHirozane-Kishikawa T, L. Q., Gabel HW, Elewa A, Baumgartner B, Rose DJ,Yu H, Bosak S, Sequerra R, Fraser A, Mango SE, Saxton WM, Strome S, VanDen Heuvel S, Piano F, Vandenhaute J, Sardet C, Gerstein M, Doucette-Stamm L, Gunsalus KC, Harper JW, Cusick ME, Roth FP, Hill DE, Vidal M.(2004) Science 303, 540-543
37. Furuyama, T., Nakazawa, T., Nakano, I., and Mori, N. (2000) Biochem J 349,629-634
38. GuhaThakurta, D., Palomar, L., Stormo, G., Tedesco, P., Johnson, T., Walker,D., Lithgow, G., Kim, S., and Link, C. (2002) Genome Research 12, 701-712
39. Hsin, H., and Kenyon, C. (1999) Nature 399, 362-36640. Arantes-Oliveira, N., Apfeld, J, Dillin, A, Kenyon, C. (2002) Science 295,
502-50541. Hosack, D., Dennis, G. J., Sherman, B., Lane, H., and Lempicki, R. (2003)
Genome Biol. 4, R7042. Walker, G. A., and Lithgow, G. J. (2003) Aging Cell 2, 131-13943. Garigan, D., Hsu, A., Fraser, A., Kamath, R., Ahringer, J., and Kenyon, C.
(2002) Genetics 161, 1101-111244. Menzel, R., Bogaert, T., and Achazi, R. (2001) Arch Biochem Biophys. 395,
158-16845. Nelson, D. (1999) Arch Biochem Biophys. 369, 1-1046. Mansuy, D. (1998) Comp Biochem Physiol C Pharmacol Toxicol Endocrinol
121, 5-1447. Jia, K., Albert, P., and Riddle, D. (2002) Development 129, 221-23148. Gerisch, B., Weitzel, C., Kober-Eisermann, C., Rottiers, V., and Antebi, A.
(2001) Dev Cell 1, 841-85149. Kallberg, Y., Oppermann, U., Jornvall, H., and Persson, B. (2002) Eur J
Biochem. 269, 4409-441750. Oppermann, U., and Maser, E. (2000) Toxicology 144, 71-8151. Gibson, G. G., and Skett, P. (2001) Introduction to drug metabolism, 3rd Ed.,
Nelson Thornes, Bath, UK52. Tukey, R., and Strassburg, C. (2000) Annu Rev Pharmacol Toxicol. 40, 581-
616
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

26
53. Wells, P., Mackenzie, P., Chowdhury, J., Guillemette, C., Gregory, P., Ishii,Y., Hansen, A., Kessler, F., Kim, P., Chowdhury, N., and Ritter, J. (2004)Drug Metab Dispos. 32, 281-290
54. Eneqvist, T., Lundberg, E., Nilsson, L., Abagyan, R., and Sauer-Eriksson, A.(2003) Eur J Biochem. 270, 518-532
55. Nielsen, H., Engelbrecht, J., Brunak, S., and von Heijne, G. (1997) ProteinEngineering 10, 1-6
56. Kuro-o, M., Matsumura, Y., Aizawa, H., Kawaguchi, H., Suga, T., Utsugi, T.,Ohyama, Y., Kurabayashi, M., Kaname, T., Kume, E., Iwasaki, H., Iida, A.,Shiraki-Iida, T., Nishikawa, S., Nagai, R., and Nabeshima, Y. (1997) Nature390, 45-51
57. Libina, N., Berman, J., and Kenyon, C. (2003) Cell 115, 489-50258. Tawe, W., Eschbach, M., Walter, R., and Henkle-Duhrsen, K. (1998) Nucleic
Acids Research 26, 1621-162759. Leiers, B., Kampkotter, A., Grevelding, C., Link, C., Johnson, T., and Henkle-
Duhrsen, K. (2003) Free Radic. Biol. Med. 34, 1405-141560. Tissenbaum, H. A., and Guarente, L. (2001) Nature 410, 227-23061. Pierrou, S., Hellqvist, M., Samuelsson, L., Enerback, S., and Carlsson, P.
(1994) EMBO J. 13, 5002-501262. Kaufmann, E., and Knochel, W. (1996) Mech Dev. 57, 3-2063. Ruvolo, V., Hill, J., and Levitt, A. (1992) DNA and Cell Biology 11, 111-12264. Scott, B., Avidan, M., and Crowder, C. (2002) Science 296, 2388-239165. Stuart, J., Segal, E., Koller, D., and Kim, S. (2003) Science 302, 249-25566. Kirkwood, T. B. L. (1977) Nature 270, 301-30467. Nehrke, K. (2003) J Biol Chem. 278, 44657-4466668. Meissner, B., Boll, M., Daniel, H., and Baumeister, R. (2004) J Biol Chem. In
press by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

27
Figure legends
Fig. 1. A, Comparison of genes identified as daf-2 regulated in three microarraystudies (16,17)(this study). B) Overlap between genes differentially expressed indaf-2 mutants and in dauer larvae.
Fig. 2. Phase 1 and phase 2 drug detoxification [adapted from (51)]. CytochromeP450’s (CYPs), short-chain dehydrogenase/reductases (SDRs), UDP-
glucuronosyltransferases (UGTs), and glutathione S-transferases (GSTs) act in concertto dispose of unwanted lipophilic and electrophilic molecules. See text for further
explanation.
Fig. 3. A new theory of aging. We postulate that a major contributor to the aging
process is molecular damage resulting from toxins (mainly endobiotic and lipophilic)
which are targets of the phase 1/phase 2 drug detoxification system. According to thisview, the smooth ER functions as a longevity organelle, derivatising and excreting
lipophilic toxins. However, this process is energetically costly, and most cells makethe energy saving step of down-regulating the detoxification system, and dumping
such toxins within lipofuscin depots. CYP, cytochrome P450 (mainly oxidases); GCE,
glutathionyl; Gluc, glucuronosyl/glucosyl; SDR, short-chain dehydrogenase/reductase;UGT, UDP-glucuronosyl- (or glucosyl-) transferases.
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

28
TABLE I
Gene groups where both the daf-2 and dauer gene sets are over-represented (p < 0.1)
Common Promoter element over-representation2
daf-2 gene set Dauer gene set overlapping ------------------------------------------------
Gene classes Score Score genes1 DBE DAE HSE HSAS HRE--------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------
Up-regulated genesDauer-, daf-2-associated gene lists
daf-2 up-regulated (this study) - 7.29e-29 (219/990) 219 *** *daf-2 up-regulated, McElwee et al. (2003) 5.15e-73 (213/489) 3.51e-38 (158/513) 86 ***Class 1 genes 2.13e-52 (109/186) 2.03e-21 (72/201) 47 *2-fold up in dauer 4.33e-26 (219/916) - - *** * *** * **SAGE dauer specific tags 3.00e-02 (47/291) 4.85e-08 (63/293) 18 *Gene expression topomountains
Mount 06 3.67e-20 (125/807) 6.44e-116 (250/803) 52 * ** * **Mount 08 9.19e-35 (139/685) 5.37e-22 (114/708) 38 ***Mount 15 6.16e-54 (94/214) 1.23e-57 (97/224) 42 **Mount 21 4.46e-02 (15/131) 1.41e-19 (44/140) 8 ***Gene classesIPR002401:E-class P450, group I 1.15e-08 (20/67) 1.29e-04 (15/74) 6
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

29
IPR002198:Short-chain dehydrogenase/reductase 6.97e-05 (17/87) 8.41e-06 (18/83) 6
IPR000379:Esterase/lipase/ thioesterase, active site 4.52e-03 (15/104) 9.32e-06 (20/101) 6
IPR001534:Transthyretin-like 1.44e-03 (10/45) 4.75e-15 (23/47) 4 **IPR000010:Cysteine protease inhibitors 6.74e-03 (4/7) 4.73e-02 (3/6) 1
IPR002068:Heat shock protein Hsp20 8.67e-03 (5/14) 7.01e-02 (4/16) 3
IPR002213:UDP-glucuronosyl/glucosyl transferase 2.29e-02 (10/68) 6.86e-07 (18/70) 6
IPR003703:Acyl-CoA thioesterase 3.34e-02 (3/5) 2.06e-03 (4/5) 2
IPR000873:AMP-dependent synthetase and ligase 3.45e-02 (6/30) 4.97e-05 (10/30) 3
IPR002018: Carboxylesterase, type B 4.65e-02 (7/43) 9.60e-04 (10/43) 2
GO:0008152:metabolism 5.49e-09 (59/427) 1.15e-15 (74/406) 20 *GO:0016491:oxidoreductase activity 1.45e-06 (35/229) 8.09e-08 (37/213) 12
GO:0006118:electron transport 2.67e-04 (44/403) 2.02e-03 (42/385) 13
GO:0003824:catalytic activity 7.57e-04 (36/322) 1.90e-07 (45/299) 11 *GO:0016787:hydrolase activity 1.06e-02 (19/161) 3.55e-02 (18/163) 5
Down-regulated genesDauer-, daf-2-associated gene lists
daf-2 down-regulated (this study) - 4.04e-11 (145/667) - *** * *daf-2 down-regulated, McElwee et al. (2003) 7.73e-99 (277/881) 1.69e-22 (220/916) 80 ***Class 2 genes 1.95e-17 (56/185) 2.24e-3 (42/205) 22 **2-fold down in dauer 6.19e-08 (145/1123) - -
Gene expression topomountains
Mount 08 4.88e-13 (74/685) 7.45e-7 (88/708) 15 ***
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

30
Mount 19 3.25e-48 (69/171) 6.13e-23 (58/176) 21 ***Mount 20 4.46e-3 (15/147) 8.7e-27 (57/146) 10
Mount 24 1.16e-30 (47/128) 1.52e-07 (29/130) 17 ***Mount 27 2.56e-17 (27/75) 1.96e-14 (30/76) 13 **Mount 30 0.0182 (6/36) 0.012 (8/35) 3
Mount 31 1.82e-5 (8/21) 0.0267 (6/24) 3
Gene classes
IPR003366:Protein of unknown function DUF141 2.70e-22 (28/50) 7.34e-6 (16/51) 11
IPR000560:Histidine acid phosphatase 2.79e-3 (6/20) 0.043 (5/17) 2
IPR006025: Neutral zinc metallopeptidases 0.0278 (11/98) 0.0388 (14/94) 3
IPR000859:CUB domain 0.0344 (7/48) 0.0154 (10/49) 5 ***IPR003582:Metridin-like ShK toxin 0.0396 (11/104) 3.16e-5 (23/106) 2
GO:0005529:sugar binding 1.21e-3 (23/207) 0.0911 (21/207) 7 ***GO:0006508: Proteolysis and peptidolysis 0.0186 (30/371) 3.81e-03 (46/337) 7
GO:0016491:oxidoreductase activity 0.0559 (19/229) 7.67e-03 (26/213) 6
GO:0003824:catalytic activity 0.0810 (24/322) 1e-5 (43/299) 8 *GO:0006730: One-carbon compound metabolism 0.093 (3/10) 0.0126 (5/11) 1
GO:0006865: Amino acid transport 0.096 (5/34) 0.0469 (7/30) 2
WI5 Y2H NW132 0.0141 (7/40) 2.04e-4 (14/37) 1
------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------
1This is the number of genes in common between the differentially expressed genes in the column “daf-2 gene set” and “dauer gene set”.
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from
31
2DBE, daf-16-binding element; DAE, daf-16-associated element; HSE, heat shock element; HSAS, heat shock-associated site; HRE, hif-1 (hypoxia-
induced factor 1) response element. * 0.05 > p > 0.01; ** 0.01 > p > 0.001; *** p < 0.001.
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

32
TABLE II
Gene categories where only the daf-2 gene set is over-represented (p < 0.001)
Promoter element over-representation1
------------------------------------------------------
Gene classes Score DBE DAE HSE HSAS HRE-------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------
Gene groups up-regulated in daf-2 mutants, not dauers
Mount 01 9.06e-06 (136/1456) *** *** ** *** ***Mount 17 5.32e-12 (40/174)
Mount 22 9.55e-05 (21/121)
IPR004045:Glutathione S-transferase, N-terminal 7.35e-08 (15/41)
Gene groups down-regulated in daf-2 mutants, not dauers
Mount 21 4.21e-15 (32/131) ***Mount 22 2.16e-04 (16/121)
IPR002213: UDP-glucuronosyl/UDP-glucosyl transferase 3.44e-08 (18/68)
IPR005071: Protein of unknown function DUF274 6.83e-09 (11/19) ***IPR005828: General substrate transporter 5.16e-06 (17/85) **IPR004119: Protein of unknown function DUF227 7.48e-06 (9/22)
GO:0006810: transport 1.00e-04 (45/472)
Stanford: Lipid metabolism genes 4.08e-04 (41/274) **
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

33
--------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------1DBE, daf-16-binding element; DAE, daf-16-associated element; HSE, heat shock element; HSAS, heat shock-associated site; HRE, hif-1
(hypoxia-induced factor 1) response element. * 0.05 > p > 0.01; ** 0.01 > p > 0.001; *** p < 0.001.
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

34
FIG. 1 A
90
McElwee et al 2003
This studyMurphy et al 2003
414
78
69
182
891
1308
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

35
FIG. 1 B
Up in dauer Up in daf-2
Down in dauerDown in daf-2
219
145
33 49
842
602
768
1072 by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from
36
FIG. 2
NADPH, NADH
CYPs
Lipophilictoxins
Phase 1metabolite
Excretion
Excretion
Excretion
Phase 2metabolites
GlucuronidesGSH-conjugatesSulfatesAmidesOther conjugates
UGTs
GSTs
Hexose
SDRs
GSH
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

37
FIG. 3
CYPs
LIPOPHILICTOXIC
METABOLITES
Excretion
Smooth endoplasmic reticulum
Diversemetabolic
&stochasticprocesses
Molecular damage Aging
Xenobiotics
OHOH
OH
OHOH
OH
OH
OH
OH
OH
OH
UGTs GSTs
GLUC
GLUC
GLUC
GLUC GLUC
GLUC
GSTGST
GST
GST
GST
Phase 1metabolism
Phase 2metabolism
OH
ROS
ROS
Plasma membrane
SDRs
OH
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

38
Supplemental Data
Further discussion of gene expression topomountains. The other daf-2/dauer up-
regulated mountains (besides Mt. 15) are linked to distinct processes: Mt. 6 to neuronal
function and Mt. 21 to lipid metabolism (1). Up-regulation of Mt. 15 genes in daf-2
mutants and dauers, and Mt. 6, 8 and 21 in dauers have been noted previously (2,3).
Of the daf-2/dauer down-regulated mountains, Mt. 19 is linked to amino acidmetabolism, lipid metabolism and cytochrome P450's, Mt. 20 to protein synthesis, heat
shock, biosynthesis and the germline, Mt. 24 to amino acid metabolism, lipid metabolism
and fatty acid oxidation, Mt. 27 to amino acid metabolism and energy generation, Mt. 30to protein expression, and Mt. 31 has no associated function. Mt. 8, which is both up- and
down-regulated is linked to the intestine. Down-regulation of Mts. 8, 19, 24 and 27 in daf-
2 mutants has been previously noted (2). In that study Mt. 21 was also found to be down-
regulated; however, in this study Mt. 21 was up-regulated in daf-2 mutants and dauers.
Other gene classes up-regulated in dauer larvae and daf-2 mutant adults --- Twogenes of the class IPR003703: Acyl-CoA thioesterase are up-regulated in both daf-2
mutants and dauers (C17C3.3, F25E2.3). Both are predicted to be peroxisomal.Peroxisomes are a major site of catabolism of endobiotic and xenobiotic lipids.
Peroxisomal acyl-CoA thioesterases (PTEs) cleave acyl-CoA, generating the free acid and
CoASH. Mammalian PTE-2 appears to be a major regulator of peroxisomal lipidmetabolism (4).
Many of the remaining up-regulated gene categories in Table I suggested no obvious
conclusions, for example, IPR000010:Cysteine protease inhibitors. Some categoriescomprised a heterogeneous mixture of genes, e.g. GO:0003824: catalytic activity. The
classes GO:0008152: metabolism, GO:0016491: oxidoreductase activity and IPR002198:
Short-chain dehydrogenase/reductase are all enriched for a common set of genes, mainly
short-chain dehydrogenases, many of which are predicted to be mitochondrial. Little is
know about the function of these genes.Gene classes down-regulated in dauer larvae and daf-2 mutant adults --- Among the
gene groups over-represented among genes down-regulated in dauers and daf-2 mutantswas a protein class bearing Interpro Domains of Unknown Function, DUF141. The
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

39
presence of DUF141 proteins among Class 2 genes was previously noted and RNAi of
four (C32H11.10, C32H11.12, F55G11.5, K10D11.1) was found to increase lifespan (5).The DUF141 proteins are a family of proteins found in C. elegans which share a
conserved domain of ~130 amino acid residues, with two conserved Cys residues; none ofthese proteins have any known function. However, 7/11 (64%) of DUF141 genes down-
regulated in both dauers and daf-2 mutants occur in Mt. 19. This topomountain is
associated with amino acid metabolism, lipid metabolism, and CYPs. Many DUF141proteins also contain a CUB domain, which is an extracellular domain of ~110 amino acid
residues. Moreover, we found that 10/11 contain a predicted N terminal signal peptide.This implies that these are secreted or transmembrane proteins.
Seven genes in the GO:0005529: sugar binding class were down-regulated in both
dauers and daf-2 adults. All seven contain C-type lectin-like domains (CTLDs). Thisdomain was originally identified in mammals as a Ca2+-carbohydrate recognition domain.
However, many vertebrate and invertebrate proteins contain diverged CTLDs which
apparently lack carbohydrate-binding activity. Some 125 C. elegans proteins contain oneor more CTLDs, mainly of the non-carbohydrate-binding type (6).
Based on the presence and order of other domains, C. elegans CTLDs have beenclassified into nine types, A - I (6). In the main, these types do not correspond to known
mammalian CTLD-containing proteins. Almost all are predicted to be secreted. The seven
down-regulated CTLDs belong to four groups: B0365.5 to group A6, F35C5.9 to groupB1, four genes to group C (F08H9.8 to C1, C03H5.1 to C2, and B0365.6 and F16H6.1 to
C3), and one gene to D3 (C14A6.1). The C1-C3 groups contain various combinations ofnon-carbohydrate-binding CTLDs, and CUB domains. The one group where the CTLD
may bind carbohydrates (possibly galactose) is D3. Down-regulation of CTLD genes is a
major reason for the presence of the IPR000859: CUB domain group in Table I. Themammalian mannose-binding proteins are secreted C-type lectins, which play a role in
innate immunity; possibly some nematode CTLD proteins have this role (6).The IPR003582: Metridin-like ShK toxin is a domain found in metridin, a toxin
from Metridium senile (the brown sea anemone). This domain is often found in
metallopeptidases. The down-regulation of this group suggests that protein up-take,digestion and/or turnover is reduced in dauers and daf-2 mutants.
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

40
Other gene classes down-regulated in daf-2 mutant adults exclusively --- Two DUF
proteins groups are also down-regulated in daf-2 adults. DUF227 proteins are found inboth C. elegans and Drosophila. The DUF227 domain contains many conserved His and
Asp residues, suggesting metal binding and possibly phosphoesterase activities. Very littleis known about DUF272 and DUF274 proteins. All of the DUF272 proteins are predicted
to be non-secreted, based on the absence of a predicted signal peptide; by contrast 9/11
DUF274 proteins are predicted to be secreted.
REFERENCES1. Kim, S., Lund, J., Kiraly, M., Duke, K., Jiang, M., Stuart, J., Eizinger, A., Wylie,
B., and Davidson, G. (2001) Science 293, 2087-20922. McElwee, J., Bubb, K., and Thomas, J. (2003) Aging Cell 2, 111-1213. Wang, J., and Kim, S. (2003) Development 130, 1621-16344. Hunt, M., Solaas, K., Kase, B., and Alexson, S. (2002) J Biol Chem. 277, 1128-
11385. Murphy, C. T., McCarroll, S. A., Bargmann, C. I., Fraser, A., Kamath, R. S.,
Ahringer, J., Li, H., and Kenyon, C. J. (2003) Nature 424, 277-2846. Drickamer, K., and Dodd, R. (1999) Glycobiology 9, 1357-13697. Menzel, R., Bogaert, T., and Achazi, R. (2001) Arch Biochem Biophys. 395, 158-
168
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

41
Supplemental Table I
Overlap between genes with selected promoter motifs, and other gene groups (p < 0.01)
Gene Classes1 Score
-----------------------------------------------------------------------------------------DBE-containing genesdaf-2 up-regulated, this study 6.39e-10 (214/1107)
Dauer up-regulated, Wang and Kim (2003) 4.39e-05 (172/989)
IPR002492:Transposase, Tc1/Tc3 5.20e-05 (18/49)
GO:0015074:DNA integration 9.28e-05 (18/51)
daf-2 up-regulated, McElwee et al. (2003) 1.45e-04 (99/526)
Mount 1 6.99e-04 (235/1521)
Mount 15 2.12e-03 (46/231)
GO:0008021:synaptic vesicle 3.05e03 (7/13)
IPR001565:Synaptotagmin 6.24e03 (5/7)
IPR001534:Transthyretin-like 7.16e-03 (14/50)
WI5 Y2H networks: NW2482 7.48e-03 (5/7)
WI5 Y2H networks: NW982 8.19e-03 (4/4)
Stanford: Biosynthesis 9.17e-03 (76/440)
Stanford: Neuronal genes 9.45e-03 (19/77)
-----------------------------------------------------------------------------------------
DBEA containing genesGO:0005529:sugar binding 5.41e-16 (77/239)
IPR001304:C-type lectin 4.31e-15 (73/223)
daf-2 down-regulated, McElwee et al. (2003) 4.07e-10 (181/935)
daf-2 down-regulated, this study 1.85e-09 (154/776)
IPR005071: DUF274 1.26e-07 (14/21)
Mount 8 1.37e-07 (134/730)
Stanford: Lectins (proteome) 5.92e-07 (53/209)
Mount 19 9.80e-07 (45/177)
IPR000859:CUB domain 1.39e-05 (20/54)
IPR002900: DUF38 5.99e-05 (52/238)
Mount 1 6.28e-05 (228/1521)
Mount 24 1.18e-04 (32/132)
IPR003326: DUF130 1.68e-04 (14/35)
IPR002083:Meprin/TRAF-like MATH 1.78e-04 (26/95)
Mount 21 4.70e-04 (32/142)
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

42
GO:0015074:DNA integration 6.06e-04 (16/51)
IPR002492:Transposase, Tc1/Tc3 6.24e-04 (16/49)
IPR001810:Cyclin-like F-box 1.14e-03 (77/434)
Mount 27 1.24e-03 (20/77)
GO:0005777: Peroxisome 1.56e-03 (6/9)
IPR002035:Von Willebrand factor, type A 1.58e-03 (13/38)
Stanford: Lipid metabolism genes 2.65e-03 (54/290)
Up-regulated hypoxia genes 2.85e-03 (24/101)
All hypoxia genes 4.21e-03 (25/110)
IPR005828:General substrate transporter 4.36e-03 (21/87)
IPR006090:Acyl-CoA dehydrogenase, C-terminal 6.57e-03 (9/24)
GO:0016829: Lyase activity 9.03e-03 (7/16)
Class 2 genes, Murphy et al. (2003) 9.24e-03 (39/207)
GO:0006635:fatty acid beta-oxidation 9.37e-03 (5/8)
-----------------------------------------------------------------------------------------
HSE containing genesIPR001888:Transposase, type 1 5.47e-27 (42/59)
Mount 25 2.67e-11 (32/83)
Dauer up-regulated, Wang and Kim (2003) 6.10e-06 (144/989)
Mount 36 1.77e-04 (6/7)
IPR000008:C2 domain 4.35e-04 (14/44)
IPR001092:Basic HLH dimerization domain bHLH 1.92e-03 (21/95)
Mount 6 3.88e-03 (108/832)
Mount 1 5.20e-03 (183/1521)
-----------------------------------------------------------------------------------------
HSAS containing genesMount1 2.64e-10 (230/1521)
GO:0007242: Intracellular signaling cascade 1.02e-03 (42/235)
IPR001849:Pleckstrin-like 3.92e-03 (14/56)
IPR000719:Protein kinase 4.39e-03 (62/423)
GO:0004713: Protein-tyrosine kinase activity 6.32e-03 (51/326)
IPR003131:K+ channel tetramerisation 7.21e-03 (12/47)
IPR000961:Protein kinase C-terminal domain 7.46e-03 (7/18)
-----------------------------------------------------------------------------------------
HRE containing genesMount 1 6.39e-04 (188/1521)
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

43
GO:0004713:Protein-tyrosine kinase activity 8.86e-04 (53/326)
Dauer up-regulated, Wang and Kim (2003) 1.34e-03 (126/989)
IPR001723:Steroid hormone receptor 2.10e-03 (14/54)
IPR000276:Rhodopsin-like GPCR superfamily 2.25e-03 (59/399)
Mount 6 2.33e-03 (108/832)
hif-1-dependent genes 2.67e-03 (15/63)
Up-regulated hypoxia genes 3.99e-03 (20/101)
All hypoxia genes 4.77e-03 (21/110)
IPR000719:Protein kinase 8.00e-03 (59/423)
GO:0006468:Protein amino acid phosphorylation 8.05e-03 (63/448)
WI5 Y2H networks NW1739 9.76e-03 (4/5)
GO:0004674:protein Ser/Thr kinase activity 9.87e-03 (58/407)
-----------------------------------------------------------------------------------------1In bold: gene groups with significant overlap with genes altered in dauers and daf-2
mutants; or underscored: in daf-2 mutants alone.
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

44
Supplemental Table IIExpression of cytochrome P450 genes in daf-2 mutants and dauer larvae
Cyp RNAi
daf-2 p Dauer p Class Clan phenotype, notes1
--------------------------------------------------------------------------------------------------------------------------------------------
Up-regulated in both daf-2 mutants and dauer larvae (6/77)B0213.15 3.68 2e-05 27.84 1.57e-05 34A9 2C. e. Suppression of daf-2 Age; Class 1
βNF
C25E10.2 1.16 0.0180 17.52 7.97e-04 33B1 2C. e.
C26F1.2 1.44 5.8e-04 5.33 4.50e-03 32A 4
1.29 0.148
F02C12.5 2.83 2.14e-03 15.05 2.15e-04 13B1 3
2.36 0.0105
K09A11.2 1.58 1.9e-04 6.43 1.39e-03 14A1 2C. e.
T10B9.2 2.14 2.35e-03 19.62 2.29e-04 13A5 3
Up-regulated in daf-2 mutants alone (12/77)B0213.10 1.55 9e-05 4.71 0.0112 34A5 2C. e. N.A.
B0213.11 1.53 3.27e-03 1.30 0.286 34A6 2C. e.
B0213.12 1.29 1.75e-03 1.24 0.383 34A7 2C. e.
B0213.14 3.66 0 0.93 0.677 34A8 2C. e. N.A.
C45H4.2 1.50 3.14e-03 3.57 6.71e-03 33C1 2C. e.
C45H4.17 2.45 6.9e-04 2.88 5.80e-03 33C2 2C. e.
1.04 0.298
C49C8.4 1.81 0.0142 1.04 0.711 33E1 2C. e. βNF
F42A9.5 1.48 9e-04 0.64 0.131 33E2 2C. e.
K05D4.4 1.39 2e-05 2.04 0.0563 33D1 2C. e.
K07C6.3 2.45 3e-05 1.18 0.367 35B2 2C. e. βNF
K07C6.4 9.38 0 3.22 0.0397 35B1 2C. e. βNF,
Suppression of daf-2 Age; Class 1
K09A11.4 1.54 3.33e-03 1.98 9.17e-03 14A3 2C. e.
Up-regulated in dauer larvae alone (3/77)
T10B9.10 1.05 0.536 12.89 1.12e-03 13A7 3 N.A.
T10H4.11 1.08 0.575 18.97 3.39e-04 34A2 2C. e.
Y5H2B.5 0.939 0.763 31.16 2.43e-05 N.A. ?
Not up-regulated in daf-2 mutants or dauer larvae (41/77)B0213.16 1.26 0.0461 5.04 0.0230 34A10 2C. e. N.A.
B0304.3 1.10 0.0439 0.59 0.0482 23A 2C. e.
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

45
1.03 0.0669
B0331.1 1.25 0.204 0.67 0.0201 29A4 4
C01F6.3 1.05 0.380 0.91 0.301 31A1 4 N.A. Xenobiotic up-regulated
C03G6.14 1.08 0.32 1.36 0.222 35A1 2C. e. βNF
C03G6.15 1.01 0.916 1.55 0.0835 35A2 2C. e. βNF
C06B3.3 1.02 0.905 1.47 0.170 35C1 2C. e. βNF
0.769 0.224
C12D5.7 1.05 0.706 0.83 0.0882 33A1 2C. e.
C34B7.3 0.802 0.273 0.99 0.946 36A1 2C. e.
C41G6.1 1.05 0.352 1.42 0.106 34A3 2C. e.
C44C10.2 0.989 0.891 0.33 8.38e-03 29A1 4
C49G7.8 0.705 0.0592 0.64 0.0159 35A4 2C. e. βNF
C50H11.15 0.989 0.946 0.65 0.0254 33C9 2C. e.
E03E2.1 1.36 0.145 1.24 0.0434 43A clan 43
1.388 0.0662
1.09 0.585
F01D5.9 0.708 0.245 1.87 0.0197 37A1 4
F08F3.7 0.443 0.0162 1.59 0.0130 14A5 2C. e. βNF
F14H3.10 0.735 0.103 1.25 0.269 35D1 2C. e. βNF
F28G4.1 0.548 0.0348 0.96 0.791 37B1 4
F41B5.2 1.17 0.177 1.22 0.139 33C7 2C. e.
F41B5.7 1.03 0.181 1.17 0.0481 33C6 2C. e.
F42A9.4 1.04 0.719 0.97 0.898 33E3 2C. e.
F44C8.1 0.526 0 1.69 0.0131 33C4 2C. e.
1.09 0.274
K06G5.2 1.07 0.575 0.79 0.366 13B2 3
K07C6.2 0.884 0.520 0.93 0.802 35B3 2C. e.
K07C6.5 0.884 0.520 0.46 8.88e-03 35A5 2C. e. Xenobiotic up-regulated
K09A11.3 0.585 0.0182 2.11 0.0294 14A2 2C. e.
K09D9.2 0.331 8.39e-03 0.83 0.108 35A3 2C. e. βNF
R08F11.3 0.913 0.583 2.41 0.0262 33C8 2C. e.
T09H2.1 1.10 0.647 3.80 0.010 34A4 2C. e.
T10B9.3 1.02 0.566 8.09 6.33e-03 13A6 3
T10B9.4 0.964 0.662 4.78 0.0599 13A8 3 10% Emb lethal, Clr, sluggish
T10B9.5 0.852 0.0871 1.19 0.240 13A3 3
T10B9.7 0.349 0 1.47 0.373 13A2 3
T10B9.8 1.00 0.843 5.62 0.0192 13A1 3
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

46
T13C5.1 1.042 0.568 0.31 0.0203 22A 2 daf-9
Y17D7A.4 1.12 0.108 1.10 0.543 33D3 2C. e. N.A.
Y38C9B.1 1.00 0.964 2.68 0.0159 29A3 4
Y49C4A.9 1.23 0.199 1.13 0.0407 33C11 2C. e. N.A.
Y80D3A.5 1.07 0.330 0.42 9.12e-04 42A 4 Unc
ZK177.5 0.964 0.771 0.88 0.477 44A Mitoch- Only mitochondrial
ondrial CYP in C. elegans
ZK1320.4 0.955 0.560 5.22 0.0144 13A10 3 N.A.
Incomplete data, or imperfect probe sets (15/77)
C36A4.1* 1.26 0.170 23.69 6.53e-05 25A1 3
C36A4.2* 1.64 4.8e-04 12.30 8.47e-05 25A2 3
C36A4.6* 1.02 0.593 13.11 3.64e-06 25A4 3 Affymetrix probes identical to F42A6.4
F14F7.2* N.A. 11.23 0.0151 13A11 3
F14F7.3* 2.15 1.33e-03 11.28 0.0178 13A12 3
F41B5.3* 0.709 1e-05 1.49 0.142 33C5 2C. e.
F41B5.4* 1.16 0.238 3.85 9.88e-04 33C3 2C. e.
F42A6.4* 1.02 0.593 9.11 3.55e-04 25A5 3 Affymetrix probes identical to C36A4.6
K06B9.1* 1.22 0.0620 40.54 1.42e-05 25A6 3
H02I12.8* 0.896 0.689 0.95 0.682 31A2 4 N.A.
R04D3.1 1.51 1e-05 N.A. 14A4 2C. e.
T10B9.1* 1.05 0.435 19.73 1.13e-04 13A4 3 Unc, suppression of daf-2 Age; Class 1
T10H4.10 1.03 0.632 N.A. 34A1 2C. e.
T19B10.1 2.27 3e-05 N.A. 29A2 4 βNF
Y17G9B.3 0.921 0.685 N.A. 31A3 4 N.A. Xenobiotic up-regulated
----------------------------------------------------------------------------------------------------------------------------------------------------1No entry in this column indicates no RNAi phenotype seen. N.A. No data available. βNF up-regulated: expression increased
in response to the xenobiotic β-naphthoquinone (7). Class 1: genes up-regulated where IIS is reduced (5).
*Genes where probe sets are less than perfect, i.e. where one or more oligonucleotides hybridises to one or more other genes.
In most cases, only a minority of oligonucleotides are predicted to cross hybridise, and thus data is probably a true reflection of
transcript abundance for the named gene. For details of probe set analysis for these genes, see haldane.biol.ucl.ac.uk.
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

47
Supplemental Table IIIFrequency of orthology with Drosophila of genes in C. elegans topomountains
Orthologous Orthologous
genes in genes in
mountain mountain Fly %D. melanogaster C. elegans representation
-----------------------------------------------------------------------------------------------Total 2,623 14,667 17.88
Up-regulated dauers and daf-2 mutants (p < 0.1)Mt 06 85 771 11.02Mt 08 73 682 10.70
Mt 21 9 129 6.97
Mt 15 13 213 6.10Mean 8.70Down-regulated dauers and daf-2 mutants (p < 0.1)Mt 20 90 126 71.42
Mt 30 22 32 68.75
Mt 27 24 72 33.33Mt 24 19 125 15.2
Mt 19 22 161 13.66Mt 08 73 682 10.70
Mt 31 1 22 4.54
Mean 31.09Up-regulated in daf-2 mutants alone (p < 0.001)Mt 01 372 1371 27.13Mt 17 3 173 1.73
Mt 22 2 124 10.17MeanDown-regulated in daf-2 mutants alone (p < 0.001)
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

48
Mt 21 9 129 6.97
Mt 22 2 124 1.61Mean 4.29No over-representation in daf-2 mutantsMt 39 7 8 87.5
Mt 41 5 7 71.42
Mt 18 100 168 59.52Mt 38 4 7 57.14
Mt 23 66 117 56.41Mt 05 407 751 54.19
Mt 40 4 8 50.0
Mt 11 214 461 46.42Mt 02 550 1252 43.92
Mt 42 2 6 33.33
Mt 34 3 11 27.27Mt 36 1 4 25.0
Mt 44 1 4 25.0Mt 25 8 53 15.09
Mt 32 2 14 14.28
Mt 07 94 684 13.74Mt 13 41 339 12.09
Mt 09 71 709 10.01Mt 16 17 192 8.85
Mt 14 25 307 8.14
Mt 0 146 2330 6.26Mt 03 65 1130 5.75
Mt 28 2 38 5.26Mt 10 25 528 4.73
Mt 12 10 397 2.51
Mt 04 18 1004 1.79Mt 26 0 86 0
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

49
Mt 33 0 3 0
Mt 35 0 14 0Mt 37 0 1 0
-----------------------------------------------------------------------------------------------
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

50
Supplemental Data
FIG 1
0
20
40
60
80
100
0 10 20 30 40 50 60 70 80
+glp-4(bn2)daf-2(m577)daf-2(e1370)glp-4; daf-2(m577)glp-4; daf-2(e1370)daf-16 glp-4; daf-2(m577)daf-16 glp-4; daf-2(e1370)
Time (days)
glp-4(bn2) acts as a weak enhancer of daf-2 Age. Figure shows summed data from two
trials. The values for each individual trial are as follows: mean lifespan ± S.E. (number of
deaths scored, number of censored values). Trial 1: N2 (wild type): 12.3±0.83 days (28,4); glp-4(bn2): 10.8±0.88 days (30, 8); daf-2(m577): 25.4±1.20 days (68, 9); daf-
2(e1370): 33.0±3.23 days (15, 24); glp-4; daf-2(m577): 29.0±1.61 days (81, 0)(daf-2 vs
glp-4; daf-2: p = 0.0086, log rank test); glp-4; daf-2(e1370): 51.7±2.87 days (35, 0)(daf-2
vs glp-4; daf-2: p = 4.1e-5, log rank test); daf-16(mg50) glp-4; daf-2(m577): 8.6±0.11 days
(61, 18); daf-16(mg50) glp-4; daf-2(e1370): 10.4±0.45 days (24, 1)(single trial only).Trial 2: N2 (wild type): 13.9±0.45 days (46, 1); glp-4: 13.3±0.50 days (53, 4); daf-
2(m577): 24.3±2.47 days (16, 15); daf-2(e1370): 36.1±3.18 days (31, 33); glp-4; daf-
2(m577): 33.3±1.00 days (86, 0)(daf-2 vs glp-4; daf-2: p = 5.8e-5, log rank test); glp-4;
daf-2(e1370): 46.6±2.40 days (47, 0)(daf-2 vs glp-4; daf-2: p = 0.023, log rank test); daf-
16(mg50) glp-4; daf-2(m577): 10.7±0.30 days (74, 9).
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Joshua J. McElwee, Eugene Schuster, Eric Blanc, James H. Thomas and David Gemsmutants implicates detoxification system in longevity assurance
Shared transcriptional signature in C. elegans dauer larvae and long-lived daf-2
published online August 11, 2004J. Biol. Chem.
10.1074/jbc.M406207200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2004/08/25/M406207200.DC1
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from





























