Embed Size (px)
Citation preview

D-MANNITOL l-PHOSPHATE DEHYDROGENASE FROM ESCHERICHIA COLI*
BY JOHN B. WOLFFt AND NATHAN 0. KAPLAN
(From the McCollum-Pratt Institute, The Johns Hopkins University, Baltimore, Maryland)
(Received for publication, March 29, 1955)
D-Mannitol is the most widely distributed of the naturally occurring hexahydric alcohols, and the compound has been known since the early part of the nineteenth century. It is found as a reserve material in some algae and many higher plan&. In some lower fungi it is the product of the fermentation of sugars. Finally, mannitol is utilized for growth and fermented by a large number of species of microorganisms.
Whereas some fungi can convert certain aldohexoses and aldopentoses to mannitol, the latter has never been found in the medium after growth on fructose (2-4). On the other hand, several species of bacteria can form mannitol from fructose. Peterson and coworkers (5-7) studied this phenomenon in pentose-fermenting organisms, and Liu (8) and Sebek and Randles (9) observed the oxidation of mannitol to fructose in pseudo- monads. No phosphorylated intermediates appeared to be involved in that conversion, as indicated by the absence of phosphorylation on ana- erobic incubation of whole cells with hexitol and ATPI (9). It is known that the same carbohydrate may be dissimilated by different organisms either directly or only after phosphorylation. This paper describes studies of an enzyme from Escherichia coli which catalyzes the oxidation of man-
* Contribution No. 110 from the McCollum-Pratt Institute. The data in this paper are taken from a dissertation submitted by John B. Wolff to the Biology De- partment, The Johns Hopkins University, in partial fulfilment of the requirement,s for the degree of Doctor of Philosophy. A preliminary report of part of this work was presented at the meeting of the Society of American Bacteriologists at Pitts- burgh, May 5, 1954 (1). This work was aided in part by grants from the American Cancer Society, as recommended by the Committee on Growth of the National Re- search Council, and the Rockefeller Foundation.
t Predoctoral Fellow of the National Institutes of Health, United States Public Health Service, 1952-54. Present address, Smithsonian Institution, Division of Radiation and Organisms, Washington 25, D. C.
1 The following abbreviations will be employed in this paper: DPN for diphospho- pyridine nucleotide; DPNH for reduced diphosphopyridine nucleotide; DPNase for diphosphopyridine nucleotidase; ATP for adenosine triphosphate; TPN and TPNH for oxidized and redured triphosphopyridine nucleotide; F-6-P for fructose R-phosphate; mannitol 1-I’ for mannitol l-phosphate; sorbitol l-I’ for sorbitol l-phos- phate; glucose 6-P for glucose 6-phosphate; and mannose 6-l’ for mannose 6-phos- phate.
849
by guest on April 16, 2020
http://ww
w.jbc.org/
Dow
nloaded from

850 D-MANNITOL 1-P DEHYDROGENASE
nitol phosphate in the presence of DPN, but will not act on the non- phosphorylated hexitol.2
EXPERIMENTAL
Materi&---DPN was purchased from either the Sigma Chemical Com- pany or the Pabst Laboratories and was approximately 90 per cent pure. DPNH was prepared according to the method of Pullman et al. (10). The concentration and purity of solutions of oxidized and reduced nucleotides were assayed by the procedure of Racker (11) with the extinction coeffi- cient 6.22 X lo6 sq. cm. mole-’ (12). Deamino DPN was prepared as described by Kaplan et al. (13). TPN of approximately 80 per cent purity was prepared froM DPN by the method of Wang, Kaplan, and Stolzen- bath (14). The oxidized and reduced 3-acetylpyridine analogues of DPN were prepared according to the method of Kaplan and Ciotti (15).
The barium salts of fructose 6-phosphate, glucose 6-phosphate, and fructose 1,6-diphosphate were purchased from t’he Schwarz Laboratories. A sample of fructose l-phosphate was generously supplied by Dr. G. T. Cori. We wish to thank Dr. B. L. Horecker for donations of ribulose 5-phosphate and sedoheptulose 7-phosphate, Dr. M. W. Slein for mannose &phosphate, Dr. H. A. Lardy for sorbose 6-phosphate and tagatose 6-phos- phate, and Dr. R. D. Hotchkiss for mannitol phosphate prepared by the method of Seegmiller and Horecker (16). Larger quantities of manirose 6-phosphate were synthesized in the course of this study from mannose by the chemical method of Posternak and Rosselet (17). All the bar- ium salts were converted to potassium salts before use by addition of an equivalent amount of KzSOh. Potassium borohydride for the reduction of phosphate esters was obtained from Metal Hydrides, Inc., Beverly, Massachusetts.
Crystalline yeast alcohol dehydrogenase was prepared by the method of Racker (11). Purified DPNase from Neurospora was prepared as the fraction precipitating with 60 per cent acetone at pH 2.7 according to the procedure of Kaplan et al. (18). In the preparation of lactic dehydro- genase, the directions of Kornberg and Pricer (19) were followed. Bovine intestinal alkaline phosphatase was obtained from the Armour Labora- tories. Dr. L. Shuster kindly furnished samples of prostatic acid phos- phomonoesterase prepared according to the procedure of Markham and Smith (20).
Methods-All spectrophotometric measurements were made with a Beck-
2 In the course of these studies, Dr. R. D. Hotchkiss and Dr. J. Marmur informed us that a similar system had been found in mutants and transformants of Diplo- coccus pneumoniae. We wish to thank Dr. Hotchkiss and Dr. Marmur for making this information available.
by guest on April 16, 2020
http://ww
w.jbc.org/
Dow
nloaded from

J. B. WOLFF AND N. 0. KAPLAN 851
man spectrophotometer, either model DU or B, with plastic cuvettes (3.0 ml. capacity) or quartz cuvettes (1.5 ml. capacity), all having a 1.00 cm. light path. In some of the calorimetric determinations a Klett-Summer- son photoelectric calorimeter was employed.
Reducing sugar was determined according to the alkaline copper-arseno- molybdate method of Somogyi (21) or by the anthrone procedure later published by Mokrasch (22). Fructose and F-6-P were estimated by the met’hod of Roe (23). Hexitols were determined by estimation of for- maldehyde produced after mild periodate oxidation, according to the pro- cedure of West and Rapoport (24), except that for the acid SnClz re- agent a more stable Asz03 reagent (1.125 M NaOH and 0.55 M Asz03) was substituted to destroy excess of periodate before addition of the chro- mogenic solution. Inorganic phosphorus was measured by the method of Fiske and Subbarow (25), and total phosphorus after sulfuric acid di- gestion of the deproteinized sample. Protein was estimated according to the procedure of Lowry et al. (26).
For paper chromatograms Whatman No. 1 or No. 4 filter paper was used. Phosphate est,ers were developed in the solvent systems of Ban- durski and Axelrod (27), Hanes and Isherwood (28), and Mortimer (29). Sugars and hexitols were run in t,he developer (I-butanol (5 volumes)- glacial acetic acid (1 volume)-water (2 volumes)) of Hough (30). Phos- phate ester spots were detected with the method of Burrows, Grylls, and Harrison (31), aldohexoses with the aniline hydrogen oxalate reagent of Horrocks and Manning (32), aldoketoses with the acid resorcinol spray reagent of Bryson and Mitchell (33), and hexitols with the alkaline silver reagent of Trevelyan et al. (34).
For ion exchange chromatography of sugar phosphates the procedure of Khym and Cohn (35) was followed, while the met’hods of Khym and Zill (36) and of Zill, Khym, and Cheniae (37) were used for sugars. It should be point,ed out that the separation of fructose, sorbitol, and mannitol was accomplished independent,ly in these studies just prior to publication of the paper last mentioned.
Growth 01 Bacteria-Large amounts of E. coli B were grown on a medium containing salt,s, yeast extract, and glucose. The cells were harvested by centrifugation, washed, and lyophilized. These dried cells were kept at room temperature without loss of enzymatic activity.
Aerobacler aerogenes (ATCC 8724) was grown by Dr. R. M. Burton. The cells were ground with alumina and extracted with phosphate buffer, and the extract was extensively dialyzed against distilled water. An extract of Clostridium kluyveri was also provided by Dr. R. M. Burton. Lactobacillus plantarum and Lactobacillus brevis (ATCC 10068) were grown by Mr. F. E. Stolzenbach. Dr. R. DeMoss kindly furnished extracts of
by guest on April 16, 2020
http://ww
w.jbc.org/
Dow
nloaded from

852 D-MANNITOL 1-P DEHYDROGENASE
Tkuconostoc mesenteroides, strain 39, grown on glucose or on arabinose, and cells of E. coli, Crookes strain (ATCC 8730). Saccharomyces cerevisiae was grown by Mr. C. DeLuca, Pseudomonas jztorescens (ATCC 6009-l) by Dr. H. Lenhoff, Neurospora crassa 5297a by Dr. D. ,J. D. Nicholas, and Lactobacillus casei (ATCC 7469 and 9595) by Dr. 1,. Shuster. jV;f are grateful to all of the above colleagues for their preparations. When- ever whole cells were provided, they were suspended in 10 volumes of 0.01 M NaHC03 buffer and disintegrated by sonic oscillation for 10 min- utes. Cell debris was removed by centrifugation, and t’he clear super- natant fluid was tested for activity.
Synthesis and Chemical Properties of Hexitol Phosphates-In analogy with the chemical reduction of fructose, which yields a mixture of sorbitol and mannitol, the chemical reduction of F-6-P would be expected to pro- duce two corresponding phosphorylated hexitols. This was shown t.o occur and will be discussed below. When the secondary alcohol group formed from the carbonyl group of fructose on carbon atom 2 lies trans with respect to that of carbon atom 3, the compound is n-sorbitol 6-phos- phate. Since carbon atoms 1 and 6 in n-mannitol have been proved to be equivalent and Rule 64 of the International Union of Chemist’ry concern- ing numbering of positions requires that “if the chain can be numbered in more than one way, the lowest numbers are given,” the second dia- stereoisomer resulting from the chemical reduction of F-6-P should be named n-mannitol l-phosphate.
CHzOH CHzOH CHzOH
I I I c=o H-C-OH HO-C-H
I I I HO-C-H
I + 2H HO-C-H HO-C-H
I + I H-C-OH H-C-OH H-C-OH
I I I H-C-OH H-C-OH H-C-OH
I I I CHzOPO,’ CH20POa’ CHzOP03’
D-Fructose 6-P Sorbitol 6-P o-Mannitol 1-P
As the reduction of a ketohexose always yields a mixture of two hexitols, it was necessary to use aldohexose phosphates for the synthesis of pure hexitol phosphates. Sorbitol 6-phosphate can be prepared from glucose 6-phosphate, and mannose 6-phosphate can be reduced to mannitol l-phosphate. Similarly, dulcitol l-phosphate should be obtained by re- ducing galactose 6-phosphate.
A solution of known concentration of hexose phosphate was added at room temperature to a solution containing one-half of the concentration
by guest on April 16, 2020
http://ww
w.jbc.org/
Dow
nloaded from

J. B. WOLFF AND N. 0. KAPLAN 853
H-C=0 CHzOH
I I II-C-OH H-C-OH
I I HO-C-H
I + 2H HO-C-H
I H-C-OH H-C-OH
I H-C-OH
I CH,OPOz-
I H-C-OH
I CHzOP03’
Glucose 6-P Sorbitol 6-P
H-C=0 CHzOH
I I HO-C-H HO-C-H
I I HO-C-H HO-C-H
I + 2% I H-C-OH H-C-OH
I I H-C-OH H-C-OH
I I CH20POs” CHzOPOa-
Mannose 6-P Mannitol 1 -P
of potassium borohydride in dilute n’aOH, pH 9 to 10. The course of the reaction was followed by testing for reducing sugar. At the end of the reaction the pH was lowered to about 1 by adding 1 N HCl in order to destroy excess KBHJ, and the solution was t.hen neutralized by adding 1 N NaOH. A small volume of Dowex 50 resin (H+ form) was added to remove excess alkali ions. After centrifugation, the supernatant fluid was freed of borate, which was found to inhibit enzymatic assays and to interfere with chromatographic experiments.
Chemical Properties of Hexitol Phosphates-Repeated attempts to sepa- rate the products of F-6-P reduction by paper chromatography or by ion exchange chromatography with either Dowex 1 (formate) and elution with 0.1 M sodium formate containing a gradually increasing concentrat,ion of formic acid or with Dowex 1 (chloride) and the eluents of Khym and Cohn (35) proved unsuccessful. This indicated a close similarity in chemical properties of the two hexitol phosphates.
The new compounds were routinely estimated by periodate oxidation by the method of West and Rapoport (24). Table I shows that the hexitol phosphat’es formed 1 mole of formaldehyde per mole, while the non- phosphorylated hexitols yielded 2 moles of formaldehyde, and the hexose phosphates formed only negligible amounts. However, t’he hexitol phos-
by guest on April 16, 2020
http://ww
w.jbc.org/
Dow
nloaded from

854 D-MANNITOL 1 -P DEHYDROGENASI?
phates were completely inert in the tests for reducing sugar and ketohexosc by which F-6-P was estimated.
The rates of chemical hydrolysis at 100” in 1 N HCI and 0.1 N KaOH of hexitol phosphates and hexose phosphates are illustratjed in Fig. I. The stability of hexitol phosphaks to alkaline hydrolysis, as \vell as lo acid hydrolysis, can be compared with the same property possessed by a-glycerol phosphate (38).
As further evidence that the hexitols were phosphorylated at the primary alcohol group, the ratio of inorganic phosphat,e liberated by acid phos- phomonoesterase to the increased amount. of formaldehyde produced after periodate oxidat’ion was determined throughout the course of the
TABLE I
Analysis of Hexitol Phosphates by Periodate Oxidation, As Compared with Other Compounds
Compound Moles formaldehyde formed per mole compound
Mannitol............................................. 2.0 Sorbitol.. ............................................ 2.0 Fructose ............................................. 1.0 Glucose .............................................. 0.2-0.3 Mannitol l-phosphate ................................ 1.0 Sorbitol 6-phosphate ................................. 1.0 Fructose 6-phosphate ................................. 0.05-0.10 Glucose 6-phosphate ................................. 0.05
Oxidation by the method of West and Rapoport (24) for 6 minutes at 23-25”.
enzymatic hydrolysis. From Table II it will be seen that initially each mole of hexitol phosphate gave rise to 1 mole of formaldehyde after perio- date oxidation and that at the end of the enzymatic dephosphorylation each mole of hexitol produced 2 moles of formaldehyde when oxidized by periodate. The ratio of inorganic phosphate to additional formaldehyde formed remained close to 1; that is, the primary alcohol group with which the phosphate had been esterified could, after periodate oxidation, form a second mole of formaldehyde per mole of hexitol as soon as the phos- phate group had been split off.
Results
Discovery of Enzyme and Assay-In the course of testing crude extracts of E. coli for phosphohexose isomerase activity, it was observed that the extracts catalyzed the oxidation of DPNH in the presence of F-6-P. There was a slight oxidation of DPNH in the absence of added substrate.
by guest on April 16, 2020
http://ww
w.jbc.org/
Dow
nloaded from
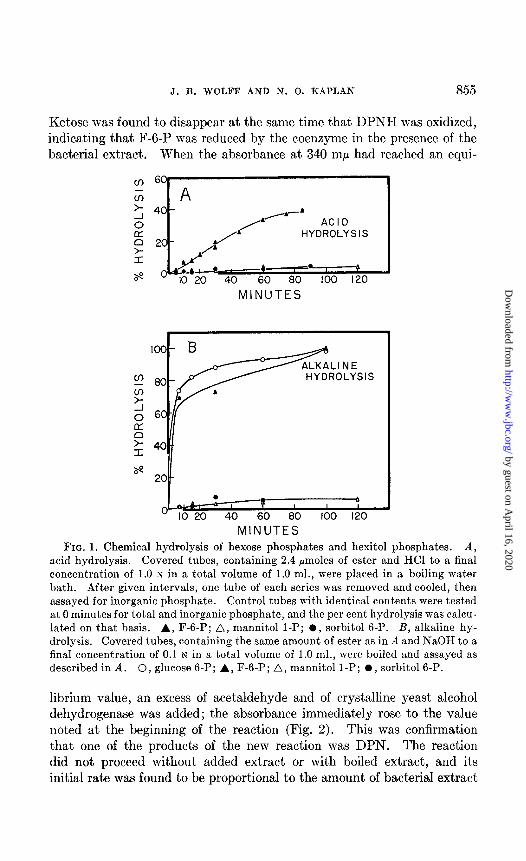
J. B. WOLFF AND N. 0. KAPLAN 855
Ketose was found to disappear at the same bime that DPNII was oxidized, indicating that F-6-P was reduced by the coenzyme in the presence of the bacterial extract,. When the absorbance at 340 rnp had reached an equi-
m 60 v, A 5 40- 0
,/-+ ACID
E5 2c- HYDROLYSIS
% ,A/
' &y, 16'20 l . . ! 0
ae 40 60 80 100 120 MINUTES
HYDROLYSIS
8 20
F . L-h-- ' f a ' I I I IO 20 40 60 80 100 120
MINUTES FIG. 1. Chemical hydrolysis of hexose phosphates and hexitol phosphates. A,
acid hydrolysis. Covered tubes, containing 2.4 pmoles of ester and HCl to a final concentration of 1.0 IV in a total volume of 1.0 ml., were placed in a boiling water bath. After given intervals, one tube of each series was removed and cooled, then assayed for inorganic phosphate. Control tubes with identical contents were tested at 0 minutes for total and inorganic phosphate, and the per cent hydrolysis was calcu- lated on that basis. A, F-6-P; A, mannitol 1-P; l , sorbitol 6-P. B, alkaline hy- drolysis. Covered tubes, containing the same amount of ester as in A and NaOH to a final concentration of 0.1 N in a total volume of 1.0 ml., were boiled and assayed as described in A. 0, glucose 6-P; A, F-6-P; A, mannitol l-P; 0, sorbitol6-P.
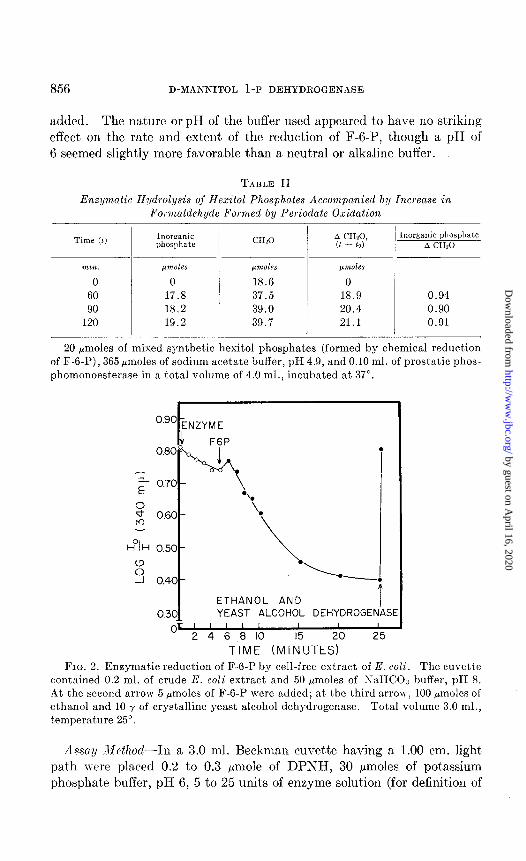
librium value, an excess of acetaldehyde and of crystalline yeast alcohol dehydrogenase was added ; the absorbance immediately rose to the value noted at the beginning of t,he reaction (Fig. 2). This was confirmation that one of the products of the new reaction was DPN. The reaction did not proceed without added extract or with boiled extract, and its initial rate was found to be proportional to the amount of bacterial extract
by guest on April 16, 2020
http://ww
w.jbc.org/
Dow
nloaded from
856 D-MANNITOL 1 -P DEHYDROGENASE
added. The nature orpH of the buffer used appeared to have no striking effect on the rate and extent of the reduction of F-6-P, though a pH of 6 seemed slightly more favorable than a neut.ral or alkaline buffer.
TABLE II
Enzymatic Hydrolysis of Hexitol Phosphates Accompanied by Increase in Formaldehyde Formed by Periodate Oxidation
Time (I) Inorganic CH20 A C&O, Inorganic phosphate phosphate (1 - to) A CHSO
n,ilz. pmoles &Lnaoles pmoles
0 0 18.6 0 60 17.8 37.5 18.9 0.94 90 18.2 39.0 20.4 0.90
120 19.2 39.7 21.1 0.91
20 pmoles of mixed synthetic hexitol phosphates (formed by chemical reduction of F-6-P), 365 pmoles of sodium acetate buffer, pH 4.9, and 0.10 ml. of prostatic phos- phomonoesterase in a total volume of 4.0 ml., incubated at 37”.
“go ENZYME
. .
ETHANOL AND ETHANOL AND YEAST ALCOHOL DEHYDROGENASE YEAST ALCOHOL DEHYDROGENASE
FIG. 2. Enzymatic reduction of F-6-P by cell-free extract of E. coli. The cuvette contained 0.2 ml. of crude E. coli extract and 50 hmoles of iVaHCOa buffer, pH 8. At the second arrow 5 pmoles of F-6-P were added; at the third arrow, 100 pmoles of ethanol and 10 y of crystalline yeast alcohol dehydrogenase. Total volume 3.0 ml., temperature 25”.
Asssa~/ M~fhod-In a 3.0 ml. Beckman cuvette having a 1.00 cm. light path n-err placed 0.2 to 0.3 pmole of DPNH, 30 pmoles of potassium phosphat,e buffer, pH 6, 5 to 25 units of enzyme solution (for definit,ion of
by guest on April 16, 2020
http://ww
w.jbc.org/
Dow
nloaded from

J. B. WOLFF AND N. 0. KAPLAN 857
the mlit see below), and wat,er to a total volume of 2.90 ml. Absorbance was read at 340 rnp in the spectrophotometer before and at 15 to 30 second intervals after mixing with 0.1 ml. of 0.1 n[ F-6-P solution. 1 unit of enzyme is defined as that amount which c+auses an initial rat,e of change in absorhancc (A RUO) of 0.01 per minute under t,he above conditions al, 25”. Specific activity is expressed as units per mg. of protein. When crude bacterial ext,racts mere assayed, it was often necessary to dilute the extract with buffer so that its absorbance at 340 rnp was small enough to permit accurate spectrophotometric readings. The rate of endogenous oxidation of DPNH must be subt>ract’ed from the rate obtained in the presence of F-6-P.
Preparation of Crude CelWree Extract-15 gm. of freshly harvested or lyophilized E. coli, strain B, were ground in a chilled mortar with an equal weight of powdered alumina (Alcoa A-301) and then extracted with 150 ml. of ice-cold 0.02 M ISaHCOs buffer. The extract was centrifuged 10 minutes at 2300 X g at 0” and the residue of alumina and cell debris discarded. The supernatant liquid was dialyzed overnight against 4 liters of 0.02 M NaHC03 solution in the cold room (4”). Alt.ernatively, the extract may be prepared by subjecting a suspension of bacteria in 10 volumes of cold buffer to sonic oscillation for 10 minutes, followed by centrifugation and dialysis.
PuriJication Procedure--The dialyzed crude extract was acidified to pH 4.7 with cold 1 N acetic acid. After centrifugation the supernatant fluid was neutralized and brought to 0.55 sat.uration (2.66 M) with solid ammonium sulfate. After centrifugation for 10 minutes at 20,000 X g (0”) the ammonium sulfate concentration of the supernatant fluid was raised to 0.65 saturation (3.16 M). After further high speed centrifugation for 10 minutes the residue containing most of the activit,y was dissolved in a minimal amount of water. The preparation had no detectable DPNH oxidase, alcohol dehydrogenase, lactic dehydrogenase, or cY-glycerol phos- phate dehydrogenase activity, but still contained some phosphohexoiso- merase activity.3 This procedure resulted in a 25- to 30-fold purification of the mannitol-phosphate dehydrogenase (see Table III).
Xtability of Enzyme-The purified preparation may be kept at -20” for several months without loss of activity. It may be further dialyzed in the cold, but this is invariably attended by progressive loss of activity with time of dialysis. Thus, dialysis for 21 hours at 4’ against 0.1 M
NaHC03-0.05 M cysteine solution, pH 7, resulted in 40 to 50 per cent loss of activity. The activity could not be restored by adding boiled enzyme or 3.3 X 1O-3 M reduced glutathione, and the loss appears to be due to protein denaturation. The enzymatic activity can be completely de-
3 The amount of phosphohexoisomerase varied in different preparations.
by guest on April 16, 2020
http://ww
w.jbc.org/
Dow
nloaded from

858 D-MANNITOL 1-l’ DEHYDROGENASE
st,royed by heat,ing for 10 minutes at 60”. Heat denaturation is more rapid at pH 5 than at pH 8.
Stoichiometry of Reaction-Preliminary experiments had shown that, in order to drive the reaction as far as possible toward completion, it was necessary to use an excess of DP;lc‘H over F-6-P (2 DPNH: 1 F-G-P). Furthermore, a preparation of DPXase from N. crassa was included in order to remove DPN as it was formed by severing its nicotinamide riboside linkage and thus draw t’he equilibrium further to the reduction of F-6-P. DPNase had little effect on the rate of the reaction, but in- creased the extent of DPNH oxidation considerably in the presence of the E. coli enzyme. Equilibrium was considered to have been reached when there was no further decrease in absorbance at 340 mp. After the addition of perchloric acid to a final concentration of 10 per cent, fresh activated
TABLE III
Summary of Purification Procedure
Crude extract.............................. 30,800 1034 30 Supernatant fluid, pH 4.7 15,600 134 117 Ammonium sulfate, 0.55-0.65 saturation. 4,900 5.7 860
Assay conditions as described in the text.
units per mg. WT.
protein
51 16
vegetable charcoal (Pfanstiehl Chemical Company, 100 mg. per ml.) was added to the protein-free solution and the mixture allowed to stand at room temperature, with frequent shaking, for 20 to 30 minutes. The charcoal was intended to remove all oxidized and reduced DPN which would otherwise have interfered in the subsequent calorimetric det,er- minations. About 80 per cent of t,he nucleotides was adsorbed in the first cycle, and the charcoal treatment was repeated as often as necessary, fresh charcoal being employed each t,ime, to remove the nucleotides com- pletely as judged by the lack of absorbance at 260 rnp of the supernatant fluid. The charcoal was then washed several times with water to recover any phosphate ester that had been adsorbed. The combined solution was neutralized with 5 N KOH and t)he resultant precipitate of KC104 removed by centrifugation. Representative data for bhe disappearance of DPNH, reducing sugar, and ketose are presented in Table IV. In spit’e of t,he variation inherent in the method of obtaining the results, there a,ppears t,o bc a fairly close relationship between the amount, of DPNH oxidized and the amount of reducing sugar disappearing.
by guest on April 16, 2020
http://ww
w.jbc.org/
Dow
nloaded from

J. B. WOLFF AND N. 0. KAPLAN 859
The reaction yields a product which, on periodate oxidation, gave a considerable increase in formaldehyde over the starting material, F-6-P. This formaldehyde is the oxidation product of a primary alcohol group which is adjacent to a secondary alcohol group. The results of Table IV indicate that for every mole of DPNH oxidized 1 mole of F-6-P is reduced, with the formation of a secondary alcohol group adjacent to an existing primary one. This suggested that the product of the reaction was ap- parently a hexitol phosphate.
F-6-P + DPNH + H+ ---f DPN+ + hexitol phosphate
Isolation and Identification oj’ Product of F-6-P Reduction--Ion exchange chromatography according to the principles developed by Khym and Cohn (35) was employed. At the end of the F-6-P reduction, an excess of both
TABLE IV Stoichiometry of F-6-P Reduction
A DPNH, /moles A reducing sugar, pmoles F-6-P
Ketose disappearance, Increase in CHzO after pm&s F-6-P periodate oxidation,
pIIdeS
2.3 2.2 2.5 4.6 3.5 4.1 5.7 6.3 7.0 2.8 3.3
10 pmoles of F-6-P, 20 pmoles of DPNH, 0.7 to 1.3 mg. of E. coli enzyme, NaHCOt buffer, pH 8.5 to 9.0; total volume 1.5 ml.; temperature 37”; time 1 to 3 hours.
acetaldehyde and yeast alcohol dehydrogenase was added to the boiled mixture in order to oxidize any residual DPNH. The solution was aerated to remove acetaldehyde, and a preparation of DPNase from N. crassa was added to the mixture to split the nicotinamide portion from the adenosine diphosphoribose moiety of DPN. The reaction was followed spectrophotometrically by the cyanide procedure of Colowick et al. (39). After completion of this reaction, the mixture was deproteinized by boiling for 10 minutes, and the supernatant fluid was concentrated under reduced pressure. The concentrate was brought to pH 9 with NaOH solution and adsorbed on a column (0.8 sq. cm. X 9 cm.) of Dowex 1 (formate) resin when solutions of sodium formate-formic acid of decreasing pH were used as eluents, or on Dowex 1 (chloride) when the eluents of Khym and Cohn were used. All 10 to 15 ml. fractions were tested for absorbance at 260 and 340 rnp, total and inorganic phosphate, ketohexose, and perio- date-oxidizable material. Nicotinamide could be washed from the col- umn with water. The product, assumed to be a hexitol phosphate, was
by guest on April 16, 2020
http://ww
w.jbc.org/
Dow
nloaded from

860 D-MANNITOL 1-P DEHYDROGENASE
eluted in the first few fractions after development of the chromatogram had begun. Inorganic phosphate followed soon afterwards. To elute F-6-P it was necessary to use 0.03 N NH&I. Adenosine diphosphoribose was tightly bound, but could be eluted with 1 M HCl. Total recovery in terms of total phosphate was approximately 95 per cent.
In ascending paper chromatograms a new phosphate ester was detected at RF 0.61 which gave a positive test for total carbohydrate but a nega- tive test for reducing sugar or ketohexose.
The reaction mixture after enzymatic reduction of F-6-P was freed of pyridine nucleotide by charcoal adsorption, and the phosphate esters were dephosphorylated with intestinal or prostatic phosphatase (20). After deproteinization by boiling, a portion of t.he mixture was assayed
TABLE V
Assay of Sorbitol with Liver Sorbitol (L-Iditol) Dehydrogenase
The tubes contained, in 1.0 ml. total volume, potassium phosphate (pH 7.8) 350 pmoles, DPN 0.7 pmole, C. kluyveri extract 0.1 ml., and rat liver sorbitol dehydro- genase 2.0 mg. The values in the table represent micromoles.
Sorbitol added.. 0 0.1 0.25 0.5 0.66 1.0 Mannitol added. 5.0
____-
Fructose found.. . . . 0 0.07 0.24 0.45 0.59 0.90 0
Incubated at 37” for 60 minutes; trichloroacetic acid added to 5 per cent final concentration. Aliquot of filtrate analyzed for fructose by method of Roe. Fruc- tose values corrected for a small amount of color formed in the blank by liver ex- tract.
for sorbitol with a preparation of sorbitol (L-iditol) dehydrogenase (40) from rat or beef liver. This enzyme has been shown to oxidize sorbitol to fructose, but it will not act on mannitol or phosphorylated compounds. The results are presented in Table V. The crude extract of C. kluyveri containing DPNH oxidase (41) was added to shift the equilibrium of the reaction as far as possible toward the complete oxidation of sorbitol by reoxidizing DPNH as it was formed. This assay is sensitive to about 0.1 Fmole per ml. of sorbitol, measured after the dehydrogenase reaction as fructose (Table V).
n-Sorbitol + DPN+ --j n-fructose + DPNH + H+
DPNH + 02 + H+ --f DPN+ + Hz02
Sum. n-Sorbitol + 02 --, n-fructose -I- Hz02
In none of the samples tested could any significant amount of sorbitol bc detect’ed, while controls containing known amounts of sorbitol gave an equivalent of fructose. This negative evidence indicated that t,he
by guest on April 16, 2020
http://ww
w.jbc.org/
Dow
nloaded from

J. B. WOLFF AND N. 0. KAPLAN 861
product of the enzymatic reduction of F-6-P was probably not sorbitol 6-phosphate.
The hexitol could not be identified by paper chromatography, but anion exchange chromatography, according to the principles developed by Khym and Zill (36) for the separation of sugars, led to its identification. Dowex 1 (borate) in a column 0.8 sq. cm. X 13 cm. was washed with 50 ml. of 0.005 M KzB40~. 10 ml. of 0.01 M KzB40, solution containing 10 mg. each of fructose, mannitol, and sorbitol (55 pmoles) were added, and the column was washed with 10 ml. of 0.005 M KzB407, then eluted with 0.015 M
KzB,07. Fractions (20 ml.) were tested calorimetrically for ketose and periodate-oxidizable material. As may be seen in the lower part of Fig. 3, fructose was eluted first, followed by one of t’he hexitols. The other hexitol could not be eluted until t’he borate concentration was raised to 0.030 M. Fractions of the last peak were combined, concentrated to dryness under reduced pressure, and recrystallized from pyridinc. The material melted at 160-164”, and authentic mannitol similarly recBryst,al- lized melted at 164-166”. Since sorbitol melts at about 97”, the second hexitol fraction was mannitol and the first therefore sorbitol. Recovery of fructose was quantitative; of the hexitols, only 60 to 70 per cent.
The dephosphorylated reaction product was chromatographed similarly, and the result is depicted in the upper port’ion of Fig. 3. The major portion of hexitol could not be eluted unt,il the borate concentrat’ion had been increased to 0.030 M. This corresponds to the behavior of mannit,ol. Hence it appeared that the product of the enzymatic reduction of F-6-P was indeed mannitol l-phosphate.
Reversibility of F-6-P Reducing Reaction-When the isolated hexitol phosphate was incubated in bicarbonate buffer at pH 9 wit,h DPN in the presence of the partially purified enzyme preparation, there was a rapid increase in absorbance at 340 rnp until a new equilibrium was reached. A simultaneous increase in ketose and reducing sugar was observed. When excess of both acetaldehyde and yeast alcohol dehydrogenasc was added, the absorbance at 340 rnp fell to its initial value.
As shown in Fig. 4, synthetic sorbitol 6-phosphate was inactive in this test, but both the mixture resulting from the chemical reduction of F-6-P and authentic mannitol l-phosphate prepared from synthetic mannose 6-phosphate possessed activity comparable to that found with the isolated product of the enzymatic reduction of F-6-P when all were tested at the same concentration. A preparation of mannitol l-phosphate synthesized by Dr. J. Marmur and Dr. R. D. Hotchkiss by direct phosphorylation of mannitol (16) was also found to be active. Thus the product of the reaction catalyzed by t,he enzyme from E. coli is D-mannitol I-phosphate, and t,he enzyme has been named D-mannitol l-phosphate dehydrogenase.
by guest on April 16, 2020
http://ww
w.jbc.org/
Dow
nloaded from

862 D-MANNITOL 1-P DEHYDROGENASE
Specijicity-Table VI indicates the remarkable substrate specificity of the enzyme. Neither free fructose nor glucose is reduced, and, within the limits of the availability of sugar phosphate esters, the substrate specificity of mannitol phosphate dehydrogenase appears to be absolute. The slight activity of the preparation with glucose 6-phosphate can be ascribed to contamination of the enzyme with phosphohexoisomerase which was not completely removed during purification.3 The specificity for the electron donor in the enzyme system is also shown.
m Y I .o
iii - c
5.0
4.0 m
Y 0 3.0 2
- 2.0
I .o
0
0
)
:
ENZYMATIC PRODUCT
!015 M K,B,O, L
, I I I 1000 1200 1400 1600 180
ML. THROUGH COLUMN
MANN ITOL
FRUCTOSE
SORB ITOL
I”..-: , - 0.015 M 0.030 M
WW7 2B407 1 I I A,
150 300 450 600 750 900 1050
ML. THROUGH COLUMN
.O
1
5.0
1.0
3.0
CO
1.0
FIG. 3. Ion exchange chromatogram (Dowex 1 resin, 200 to 400 mesh, borate form) of fructose, mannitol, and sorbitol solution and of dephosphorylated enzymatic re- action product solution.
Saturation Curves-Values of the Michaelis con&ants for half saturation of the enzyme, calculated by the met’hod of Lineweaver and Burk (42) and plotted by the method of least squares, are 1.17 X 10m3 M F-6-P, 1.40 X 1OW M mannitol l-phosphate, 2.49 X 1O-4 M DPNH, and 2.29 X 1 CF4 M DPN.
Effect of Inhibitors on Mannitol i-Phosphate Dehydroyenase-Semicarba- zide, hydroxylamine, and sodium fluoride (all at 5 X low2 M) did not in- fluence the enzymahic oxidation of mannitol phosphate. The absence of inhibition by disodium et,hylenediaminetetraacetate (5.4 X 10W4 nr), I<lic?I’, or Sax:< (lO--zl u), considered togcthrr with the lack of stimulation
by guest on April 16, 2020
http://ww
w.jbc.org/
Dow
nloaded from
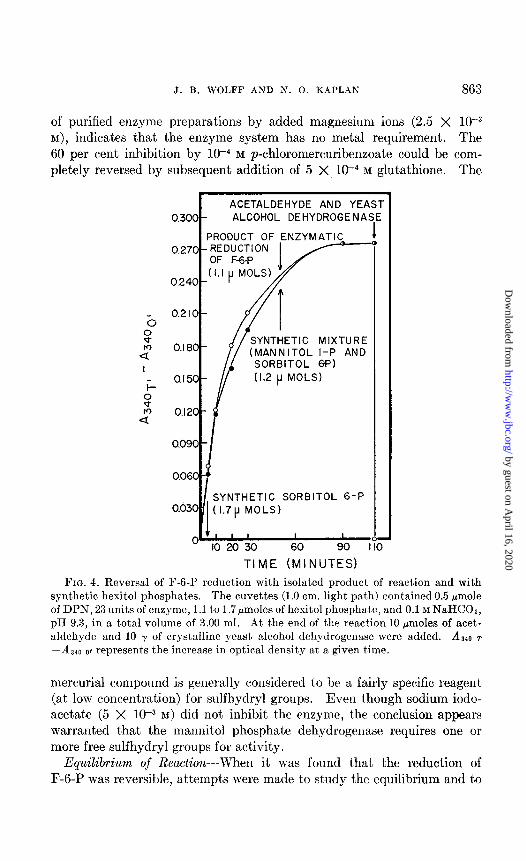
J. B. WOLFF AND S. 0. KAPLAN 863
of purified enzyme preparations by added magnesium ions (2.5 X 10e3 M), indicat,es t,hat the enzyme system has no metal requirement. The 60 per cent inhibition by 1O-4 M p-chloromercuribenzoate could be com- pletely reversed by subsequent addition of 5 X 10v4 M glut,athione. The
c ACETALDEHYDE AND YEA 0.300 ALCOHOL DEHYDROGENA’
PRODUCT OF ENZYMATL REDUCTION
SYNTHETIC MIXTURE (MANNITOL I-P AND
SORBITOL 6f’) (1.2 p MOLS)
SYNTHETIC SORBITOL 6-P
O 11, , , I I
IO 20 30 60 90 I
TIME (MINUTES)
.
FIG. 4. Reversal of F-6-P reduction with isolated product of reaction and with synthetic hexitol phosphates. The cuvettes (1.0 cm. light path) contained 0.5 pmole of DPN, 23 units of enzyme, 1.1 to 1.7pmoles of hexitol phosphate, and 0.1 M NaHCOa, pH 0.3, in a total volume of 3.00 ml. .4t the end of the reaction 10 pmoles of acet- aldehyde and 10 y of crystalline yeast, alcohol dehydrogenase were added. Az40 T -A 340 0, represents the increase in optical density at a given time.
mercurial compound is generally considered to be a fairly specific reagent> (at low concentration) for sulfhydryl groups. Even though sodium iodo- acetat.e (5 X 1O-3 M) did not inhibit t,he enzyme, t~he conclusion appears warranted that the mannitol phosphate dehydrogenase requires one or more free sulfhydryl groups for activity.
Equilibrium of Reaction--When it was found t,hat, the reduction of F-6-P was reversible, attempts were made to st,udy the equilibrium and to
by guest on April 16, 2020
http://ww
w.jbc.org/
Dow
nloaded from

864 D-MANNITOL 1-P DEHYDROGENASE
calculate an equilibrium constant for the reaction
n-Mannitol 1-P + DPN+ + F-6-P + DPNH + H+
(F-6-P)(DPNH)(H+) Ke = (Mannitol l-P)(DPN+)
In these experiments the relative concentrations of F-6-P and DPNH were varied, as well as the pH between 6 and 10. The values for DPN and mannitol l-phosphate were calculated by subtracting the measured
TABLE VI
Substrate + Coenzyme Specijicity of Mannitol l-Phosphate Dehydrogenase
Substrate*
D-Fructose B-phosphate 100 n-Glucose B-phosphate 10 D-Fructose 0 o-Glucose 0 D-Fructose l-phosphate 0
‘I 1,6-diphosphate 0 n-Mannose B-phosphate 0 L-Sorbose 6-phosphate 0 n-Tagatose 6-phosphate 0 D-Ribulose 5-phosphate 0 n-Sedoheptulose 7-phosphate 0
Per cent
rctiv- ity
Coenzymet
DPNH Deamino DPNH 3-Acetyl pyridine analogue of DPNH TPNH
P.3 cent .ctiv- ity
100
22 0 0
* 550 pmoles of phosphate buffer, pH 7.5, 0.07 @mole of DPNH, 3 pmoles of sub- strate, and 15 units of partially purified enzyme, in a total volume of 1 .O ml. Tem- perature 23”.
t 300 pmoles of NaHC03 buffer, pH 8, 15 ,umoles of F-6-P, 4 units of partially purified enzyme, 0.2 pmole of coenzyme, in a total volume of 3.0 ml. Temperature 23".
changes in concentrat,ion of DPNH and F-6-P from the known concentra- t,ions added. Some data were also obtained, starting with known con- centrations of mannitol l-phosphate and DPN and varying the pH. Here the amount of DPN reduwd was taken as equal to the F-6-P formed by oxidation of mannitol I-phosphate. From the average K, at pH 0 of 4.9 f 3.6 X 10W", the change in free energy for the oxidation of mannitol l-phosphate has been calculated as A F” m -12,700 calories.
Eflect of pH on Enzyme Activity-As mentioned earlier, pH or the nature of the buffer (with phosphate, pyrophosphate, glycylglycine, or bicarbonate) had no marked tffect] on the initial rate of F-6-P reduction by the enzyme. The only effect that could be observed in numerous
by guest on April 16, 2020
http://ww
w.jbc.org/
Dow
nloaded from

J. B. WOLFF AND N. 0. KAPLAK 865
attempts to detect a pH optimum for the reaction was a very gradual decline in activit,y from pH 6 to pH 10; therefore the lowest pH (6.0) at which DPNH is stable was generally used in assays. However, the en- zymatic oxidation of mannitol l-phosphate in the presence of DPN was found to be markedly affected by pH. As a consequence of the finding that maximal activity occurred near pH 10, NaHC03 buffer of pH 9 to 10 was used routinely in assays in this direction.
Distribution of Mannitol 1 -Phosphate Dehydrogenase-Extracts of several microorganisms were tested in a mixture consisting of 2.56 pmoles of mannitol l-phosphate, 0.60 pmole of DPN, 10 pmoles of NaHC03 buffer, pH 9.0, and 0.05 ml. of extract in a total volume of 1.0 ml. 1 unit of
TABLE VII
Distribution of Mannitol I-Phosphate Dehydrogenase
Organism (ATCC No.) /
Specific activity
A. aerogenes (8724). E. coli (Crookes strain) (8739). L. casei (9595) ‘I “ (7469). ” plantarum. S. cerevisiae.. L. mesenteroides (glucose-grown). “ “ (arabinose-grown).
units per nzg. protein
1000 638 660” 350*
193,110 21 19 0.6
All units are expressed in terms of oxidat.ion of mannitol l-phosphate. * Assayed by reduction of F-6-P.
enzyme is defined as that amount which causes an initial rate of change in absorbance at 340 rnp of 0.010 per minute at 23” under the above condi- hions. The two L. casei strains were tested wihh F-6-P instead of mannitol l-phosphate. Positive results are presented in Table VII. Yeast ap- pears to resemble bacteria more than fungi in this aspect of metabolism; in the only other mold tested, N. crassa 5297a, no activity was detected. The high activity found in members of the colon-aerogenes and homo- fermentative (L. casei) and heterofermentative (L. plantarum and L. mesenteroides) lactobacillus groups indicates that mannitol l-phosphate dehydrogenase represents a major pathway of ma,nnitol and fructose ut,ilization in those organisms.
DISCUSSION
The enzyme from E. coli discussed here differs in its substrate specificity from the polyol dehydrogenase of mammalian liver discovered by Blakley
by guest on April 16, 2020
http://ww
w.jbc.org/
Dow
nloaded from

866 D-MANNITOL 1-P ‘DEHYDROGENASE
(40) and further tested by Edson and McCorkindale (43, 44). The latter enzyme is believed to be identical with that from rat accessory sexual organs found by Williams-Ashman and Banks (45). Whereas the E. coli protein will catalyze only the reduction of F-6-P or the oxidation of mannitol l-phosphate in the presence: of .pyridine ~nucle.otide, the. liver enzyme oxidizes n-sorbitol,. n-iditol, and a few. other sugar alcohols fitting the specificity rules deduced by Edson and,McCorkindale; but it will not attack mannitol or any phosphorylated compounds. The liver enzyme, however, shares with the E. coli enzyme a coenzyme specificity for DPN.
The very high degree of substrate specificity and substrate affinity of the mannitol l-phosphate dehydrogenase makes the enzyme a useful analytical tool in the study of certain phases of carbohydrate metabolism. With the partially purified enzyme preparation from E. coli it has been possible to detect concentrations of F-6-P as low as 2 X 1O-6 M, which lies below the sensitivity of most commonly used calorimetric and chromat- ographic methods.
It should be noted that, although the mannitol phosphate dehydrogenase is quite specific, an enzyme capable of oxidizing sorbitol 6-phosphate has been detected in E. coli grown on sorbitol (46). Extracts of E. coli cells grown on dulcitol have been found to reduce tagatose 6-phosphate; cells grown on sorbitol or mannitol are not able to reduce this ketose sugar
(46). It is of considerable int,erest to note that Marmur and Hotchkiss have
recently found mannitol phosphate dehydrogenase activity in Diplococcus pneumoniae cells able to grow and metabolize mannitol in media in which mannitol is the limiting energy source (47). This ability to utilize man- nitol can be transferred to non-mannitol-utilizing cells by incubating the latter for a short time with deoxyribonucleate preparations from mannitol- utilizing mut.ants.
In comparing the equilibrium constant obtained in t,his study with values for the liver n-iditol dehydrogenase, it should be pointed out that Blakley’s (40) value of 0.240 f 0.013 at 20” and at pH 8.0 does not in- clude t,he hydrogen ion concentration. On the other hand, the hydrogen ion concentration is included in the average equilibrium constant of values determined at various hydrogen ion concentrations by Williams-Ashman and Banks (45), given as 2.05 X lO+ at 25” for the oxidation of sorbit.01. These authors observed that a rise in pH displaces the equilibrium of the reaction toward the complete oxidation of sorbitol. A similar effect has been noted for mannitol l-phosphate oxidation in the E. coli system. The value of the equilibrium constant obtained with the liver sorbitol system is of the same order of magnitude as that found for the E. coli mannitol phosphate dehydrogenase, but is about 100 times larger than those re-
by guest on April 16, 2020
http://ww
w.jbc.org/
Dow
nloaded from

J. B. WOLFF AND N. 0. KAPLAN 867
ported for the oxidation of glycerol to dihydroxyacetone in A. aerogenes* or of glycerol phosphate to dihydroxyacetone phosphate by Baranowski’s muscle enzyme (48). The difference may be due to the increased stability of F-6-P in its 2,5-furanose form. Although it is presumably the acyclic form of F-6-P with a free carbonyl group that is active in the enzymatic reduction, most of the product of the oxidat,ion of mannitol l-phosphate will be immediately cyclized to the more st’able furanose form of F-6-P as soon as it is formed, thus displacing the equilibrium further toward the formation of the ketose than is possible in t,he case of 3-carbon com- pounds.
A more extensive survey of bacteria of the groups Lactobacteriaceae and Pseuddmonadaceae should be made before definite conclusions can be drawn about the presence or absence of a pathway involving the phos- phorylated mannitol as an intermediate. That this pathway exists in members of the coli-aerogenes and lacbic acid bacteria has been shown, but it is not clear whether all organisms which can interconvert fructose and mannitol utilize the phosphorylated route or whether a direct oxida- tion may occur in some species, as has been suggested by Sebek and Ran- dles and others.
SUMMARY
1. A soluble enzyme from Escherichia coli B has been found to reduce F-6-P reversibly to mannitol I-phosphate in the presence of DPNH. The enzyme has been purified some 30-fold by isoelectric precipitation of protein impurit)ies and fractionation with ammonium sulfate.
2. The enzyme has an absolute specificity for F-6-P and a high degree of specificity for DPNH; TPNH is inactive. The dissociation (Michaelis) constant,s are 1.17 X 1O-3 M F-6-P, 2.49 X lop4 M DPNH, 1.40 X 10-* M mannitol l-phosphate, and 2.29 X 1e4 M DPN.
3. The reaction product has been identified as mannitol l-phosphate by (a) identity in enzymatic and chemical properties with synthetic mannitol 1-phosphat’e, (b) anion exchange chromatography after enzy- matic dephosphorylation, and (c) lack of activit>y of the dephosphoryl- ated compound with the sorbitol (L-iditol) dehydrogenase from liver.
4. Sorbitol- A-phosphate and mannitol l-phosphate have been synthe- sized by potassium borohydride reduction of glucose 6-phosphate for t’he former and of mannose 6-phosphate for the latter. Mannose B-phosphate was also prepared from mannose by chemical synthesis.
5. The enzyme requires one or more free sulfhydryl groups for activity, as shown by its sensitivit,y to p-chloromercuribenzoat’e and reversal of this inhibitory action by glutathione. No metal cofactors are required.
4 Burton, R. M., Lamborg, M. R., and Kaplan, N. O., in preparation.
by guest on April 16, 2020
http://ww
w.jbc.org/
Dow
nloaded from

868 D-MANNITOL 1-P DEHYDROGENASE
6. The pH for optimal mannitol l-phosphate oxidation lies near 10, but the optimum for F-6-P reduction is not as sharply defined, being near pH 6.
7. The equilibrium of the reaction lies far toward reduction of F-6-P. The value of the constant is considerably higher than that for the forma- tion of triose phosphate from glycerol phosphate, the difference presumably being due to formation of the furanose form of F-6-P.
8. The enzyme has been found in crude extracts of bacteria of the colon- aerogenes group, lactobacilli, and in yeast.
BIBLIOGRAPHY
1. Wolff, J. B., and Kaplan, N. O., Bact. Proc., 110 (1954). 2. Birkinshaw, J. H., Charles, J. H. V., Hetherington, A. Cl., and Raistrick, H.,
Tr. Roy. Sot. London, Series B, 220, 153 (1931). 3. Coyne, F. P., and Raistrick, H., Rio&em. J., 26, 1513 (1931). 4. Birkinshaw, J. H., and Raistrick, H., Tr. Roy. See. London, Series B, 220, 331
(1931). 5. Peterson, W. H., and Fred, E. B., J. Biol. Chem., 41,431 (1920). 6. Peterson, W. H., and Fred, E. B., J. Biol. Chem., 42, 273 (1920). 7. Stiles, H. R., Peterson, W. H., and Fred, E. B., J. Biol. Chem., 64,643 (1925). 8. Liu, P., J. Bat., 64,773 (1952). 9. Sebek, 0. K., and Randles, C. I., J. Bat., 63, 693 (1952).
10. Pullman, M. E., Colowick, S. P., and Kaplan, N. O., J. Biol. Chem., 194, 593 (1952).
11. Racker, E., J. Biol. Chem., 164,313 (1950). 12. Horecker, B. L., and Kornberg, A., J. Biol. Chem., 175,385 (1948). 13. Kaplan, N. O., Colowick, S. P., and Ciotti, M. M., J. Biol. Chem., 194,579 (1952). 14. Wang, T. I’., Kaplan, N. O., and Stolzenbach, F. E., J. Biol. Chem., 211, 465
(1954). 15. Kaplan, N. O., and Ciotti, M. M., J. Am. Chem. Sot., 76, 1713 (1954). 16. Seegmiller, J. E., and Horecker, B. L., J. Biol. Chem., 192,175 (1951). 17. Posternak, T., and Rosselet, J. P., Helv. chim. actu, 36, 1614 (1953). 18. Kaplan, N. O., Colowick, S. P., and Nason, A., J. Biol. Chem., 191,473 (1951). 19. Kornberg, A., and Pricer, W. E., Jr., J. BioZ. Chem., 193,481 (1951). 20. Markham, R., and Smith, J. D., Biochem. J., 62,558 (1952). 21. Somogyi, M., J. BioZ. Chem., 196,19 (1952). 22. Mokrasch, L. C., J. BioZ. Chem., 208,55 (1954). 23. Roe, J. H., J. Biol. Chem., 107, 15 (1934). 24. West, C. D., and Rapoport, S., Proc. Sot. Exp. BioZ. and Med., 70, 141 (1949). 25. Fiske, C. H., and Subbarow, Y., J. BioZ. Chem., 66,375 (1925). 26. Lowry, 0. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J., J. BioZ. Chem.,
193, 265 (1951). 27. Bandurski, R. S., and Aselrod, B., J. Biol. Chem., 193, 405 (1951). 28. Hancs, C. S., and Isherwood, F. A., Nature, 164,1107 (1949). 29. Mortimer, D. C., Cunad. J. Chem., 30, 653 (1952). 30. Hough, L., Nature, 166, 400 (1950). 31. Burrows, S., Grylls, F. S. M., and Harrison, J. S., Nature, 170, 800 (1952). 32. Horrocks, R. H., and Manning, G. S., Luncet, 266,1042 (1949).
by guest on April 16, 2020
http://ww
w.jbc.org/
Dow
nloaded from

J. B. WOLFF AND N. 0. KAPLAN 869
33. Bryson, J. L., and Mitchell, T. J., Nature, 16’7,864 (1951). 34. Trevelyan, W. E., Procter, D. I’., and Harrison, J. S., Nature, 166, 444 (1950). 35. Khym, J. X., and Cohn, W. E., ./. Am. Chem. Sot., 75, 1153 (1953). 36. Khym, J. X., and Zill, L. I’., J. Am. Che,m. Sot., 74, 2090 (1952). 37. Zill, L. I-‘., Khym, J. X., and Cheniae, G. M., J. Am. Chem. Sot., 75, 1339 (1953). 38. Kaplan, N. O., and Greenberg, D. M., J. Biol. Chem., 166,511 (1944). 39. Colowick, S. I’., Kaplan, N. O., and Ciotti, M. M., J. Biol. Chem., 191,447 (1951). 40. Blakley, R. L., Bioch,em. J., 49, 257 (1951). 41. Weber, M. M., and Kaplan, N. O., Bat. P/,oc., 96 (1954). 42. Lineweaver, K., and Burk, D., J. Am. Chem. Sot., 56,658 (1934). 43. Edson, N. I,., Australian and New Zealand Association for the Advancement of
Science, Report of the 29th meeting, Sidney, 1952; 29, 281 (1953). 44. McCorkindale, J., and Edson, N. I,., Biochem. tJ., 57, 518 (1954). 45. Williams-Ashman, H. G., and Banks, J., Arch. Biochem. und Biophys., 50, 513
(1954). 46. Wolff, J. B., and Kaplan, N. O., J. Bat., in press. 47. Hotchkiss, R. D., and Marmur, J., Proc. Nat. Acad. SC., 40, 55 (1954). blarmur,
J., and Hotchkiss, R. D., J. Biol. Chem., 214,383 (1955). 45. Baranowski, T., J. Biol. Chem., 183, 535 (1949).
by guest on April 16, 2020
http://ww
w.jbc.org/
Dow
nloaded from

John B. Wolfe and Nathan O. KaplanESCHERICHIA COLI
DEHYDROGENASE FROM d-MANNITOL 1-PHOSPHATE
1956, 218:849-869.J. Biol. Chem.
http://www.jbc.org/content/218/2/849.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
alerts to choose from all of JBC's e-mailClick here
tml#ref-list-1
http://www.jbc.org/content/218/2/849.citation.full.haccessed free atThis article cites 0 references, 0 of which can be
by guest on April 16, 2020
http://ww
w.jbc.org/
Dow
nloaded from





























