Embed Size (px)
Citation preview

J. Cell Set. 43, 379-39O (1980)Printed in Great Britain © Company of Biologiits Limited igSo
CYTOSKELETON CHANGES IN FIBROBLAST
ADHESION AND DETACHMENT
R. ANDREW BADLEY, ANNE WOODS, LINDA CARRUTHERSAND DAVID A. REESUnilever Research, Cokuorth Laboratory, Sharnbrook, Bedford MK44 iLQ, U.K.
SUMMARY
The organization of the cytoskeleton in several anchorage-dependent fibroblast types hasbeen compared with the pattern of adhesions to a glass substratum which will support eithertheir growth or just their spreading. Components were stained separately for immuno-fluorescence microscopy using specific antisera against actin, tubulin, and gizzard 10-nmfilament protein, and the adhesions were visualized by interference-reflexion microscopy.
Of the cytoskeleton features, only stress fibres could be related to the pattern of focal adhes-ions; as shown before, each focal adhesion lies directly beneath a stress fibre, often near theterminus. Cells spread on fibronectin-treated glass in serum-free medium to arrest the develop-ment of focal adhesions, show correspondingly underdeveloped stress fibres. Actin geodesicdomes, microtubules, and 10-nm filaments showed no relations with the adhesion pattern.During cell rounding leading to detachment with either EGTA or trypsin, stress fibres beginto disperse in advance of shape change whereas microtubules and 10-nm filaments seem toalter their distribution as a consequence of shape change. We therefore confirm that stressfibres are the cytoskeleton features most directly related to focal adhesions and are cytoskeletontargets for 2 agents which cause rounding and hence detachment.
The sequences of events in dispersal of stress fibres by EGTA and by trypsin showed sig-nificant differences in detail. With trypsin, fibres higher in the cell and terminating at the celledge were more sensitive than most basal fibres and, during disintegration, all types of fibrewent through an intermediate ' beaded' structure. With EGTA, all stress fibres seemed to besimilarly susceptible and the beaded stage was not seen. The implications of these differencesfor our understanding of the mechanisms of dispersal of stress fibres are discussed.
INTRODUCTION
The adhesion of anchorage-dependent fibroblasts to a surface which will supporttheir growth is accompanied by the development of a visibly organized cytoskeleton,the individual components of which include microtubules (Brinkley, Fuller &Highfield, 1975; Weber, Pollack & Bibring, 1975), several forms of the so calledintermediate or 10-nm filaments (Franke, Schmid, Osborn & Weber, 1978), and actinstructures which differ from each other in composition (Lazarides, 1976 a; Heggeness,Wang & Singer, 1977) and/or in geometrical form. Examples of distinct actin structuresare the bundles of microfilaments known as stress fibres which have associatedmyosin, a-actinin and other proteins often including tropomyosin or filamin, thepolygonal nets or geodesic domes (Lazarides, 19766; Osborn, Born, Koitsch &Weber, 1978) built from short lengths of actin bundles again with other muscleproteins associated, and a meshwork in which geometrical regularity is less apparent(Buckley, 1975). Intimate adhesion in many but not all (Couchman & Rees, 1979 a, b)
25-2

380 R. A. Badley, A. Woods, L. Carruthers and D. A. Rees
types of fibroblast is confined to specialized zones known as focal contacts or focaladhesions (Abercrombie, Heaysman & Pegrum, 1971) which are convenientlyvisualized in living cells as circular or elliptical black zones of area 1-10/tm2 by thetechnique of interference-reflexion microscopy (Curtis, 1964; Abercrombie & Dunn,1975) and each of which it seems is always associated with the terminus of a stressfibre (Heath & Dunn, 1978; Badley et al. 1978). Such contacts are known to be veryclose to the substratum (Abercrombie et al. 1971; Badley, Woods, Smith & Rees,1979) and to represent a large refractive index difference between adhesion andsubstrate (Bereiter-Hahn, Fox & Thorell, 1979). They can be distinguished from areasof ' close contact' which appear as ' grey zones' by interference-reflexion microscopydue to a greater membrane substrate distance and/or refractive index change (Izzard &Lochner, 1976; Heath & Dunn, 1978; Bereiter-Hahn et al. 1979).
Our earlier investigation of cell-spreading (Thorn, Powell & Rees, 1979) showed2 stages distinguished by different requirements for serum components and by differenttypes of interference-reflexion image. On a fibronectin-coated surface in serum-freemedium, spreading stops (at 'stage 1') when cells have a well developed fibroblasticmorphology but the interference-reflexion image shows a mottled pattern of greyzones instead of the well defined focal adhesions seen in cells growing under normalculture conditions in serum. In this paper we describe for several fibroblast types acomparison of the organizations of the cell undersides and the different componentsof the cytoskeleton for cells in these 2 states. We also report the sequence of alterationsoccurring in the cytoskeleton during cell detachment both with trypsin (confirmingand extending the results of Pollack & Rifkin, 1975) and with EGTA. Our resultsprovide further evidence that cell stiffening by stress fibres is responsible for theincreased adhesiveness ('grip'; Rees, Lloyd & Thorn, 1977) seen in cells which formfocal adhesions. Other cytoskeletal components seem to influence adhesion lessdirectly, and indeed to distribute within the cell in ways that follow rather than precedechanges in adhesion and cell shape.
MATERIALS AND METHODS
Cells
16C cells (Colworth strain of an established spontaneously transformed line of rat dermalfibroblasts), rat embryo fibroblasts REF (Flow), and BHK cells were cultured as previouslydescribed (Rees et al. 1977; Badley et al. 1978). Chick embryo fibroblasts, CEF, were obtainedby trypsinization of embryos 7-9 days old (Kemp, Jones, Cunningham & James, 1967) andmaintained at 37 °C in Eagle's Basal Medium with Earle's salts plus 10 % foetal bovine serumunder an atmosphere of 10 % carbon dioxide in air. New CEF cultures were started every3 weeks.
For immunofluorescence experiments REF, BHK and CEF were seeded at densities ofio4 cm~a, 1-75 x io4 cm"1, and 2-75 x io4 cm"1 respectively on 22-mm diameter coverslips andgrown 24 h at 37 °C as above.
To obtain 16C, BHK or REF spread to Stage 1 (Thorn et al. 1979), the cells were suspendedwith EGTA (see below) and seeded at a density of 10s cm"1 onto 22-mm diameter glass cover-slips precoated by incubating overnight with chick serum cold-insoluble globulin (140000Dalton fraction; see Thorn et al. 1979; 30 fig in 1 ml of Hanks' balanced salts solution on eachcoverslip). Cells were allowed to spread for 5-8 h in serum-free medium. Control, fully spread,

Cytoskeleton and adhesion 381
cultures were obtained on uncoated coverslips in medium plus 10 % foetal bovine serum culturedovernight.
Certain experiments required treatments of the cells with enzymes or drugs. Crystallinetrypsin (Sigma) was freshly dissolved at 25 fig ml"1 in Hanks' balanced salt solution (HBSS;Flow) and used for 10 min at 37 °C followed by soybean trypsin inhibitor (Sigma, 100 fig ml"1
for 2 min at 37 °C). EGTA was used at 005 % for 10 min at 37 °C in Caa+- and Mg*+-freephosphate-buffered saline (PBS; Badley et al. 1978).
Antisera
Preparations of the antibodies against chicken gizzard actin, bovine brain tubulin andchicken gizzard 10-nm filament protein (Badley et al. 1978) were further purified by affinitychromatography on columns of Sepharose 4B coupled to appropriate antigens as previouslydescribed (Lloyd et al. 1979). Antigen from pig smooth muscle was used for purifying anti-gizzard 10-nm filament protein. Some results were obtained using antibodies against BHKintermediate filaments (Starger & Goldman, 1977) raised in guinea pigs (Fig. IG).
In view of some uncertainty regarding the cross-species and cross-tissue reactivities ofantisera raised against 10-nm filament proteins (Lazarides & Balzer, 1978; Bennett et al. 1978;Kurki, Linder, Virtanen & Stenman, 1977; Lazarides & Hubbard, 1976; Hynes & Destree,1978; Gordon, Bushnell & Burridge, 1978; Starger, Brown, Goldman & Goldman, 1978;Lazarides, 1978; Davison, Hong & Cooke, 1977; Franke et al. 1978) the sera used in the presentexperiments were characterized using chicken and rat myofibrils, each of the 4 types of fibro-blast, and these same fibroblasts treated with colchicine (Starger & Goldman, 1977) which isknown to produce perinuclear coils and whorls of 10-nm filaments. The antibodies wereaffinity-purified on a column of the pig antigen. They stained all BHK cells in each population,the majority of CEF, a minority of the REF' but none of the 16C cells. Those cells in the CEFand REF populations which did not stain were otherwise indistinguishable from those that did.Myofibrils prepared from glycerinated chicken and rat muscles were stained at the Z-line asexpected (Lazarides & Hubbard, 1976). The guinea-pig antibodies gave very similar results tothe rabbit anti-chicken gizzard 10-nm filament antibodies when used on BHK cells.
Fluorescein (FITC) conjugated goat anti-rabbit antibodies were obtained from Nordic(Maidenhead, U.K.) or Miles (Slough, U.K.) and FITC-conjugated rabbit anti-guinea pigantibodies from Miles.
Fluorescent labelling of cells
Fixation (3-5 % formaldehyde) and staining procedures have been described (Badley et al.1978; Thorn et al. 1979). Controls included substitution of specific antibodies by preimmunesera, absorption of sera with specific antigen and use of the fluorescent antibody alone. All theseprocedures yielded negative results. For the correlation of focal adhesions with cytoskeletalcomponents, cells were fixed and photographed using interference-reflexion optics prior toextraction and antibody staining.
Stained cells were viewed on a Leitz Orthulux II microscope fitted with epi-illumination andphase and interference-reflexion optics. Photographs were on Ilford FP4 and HP5 films usingratings between 400 and 1000 ASA, processed in Promicrol (May & Baker) or ID 11 (Ilford)developers.
RESULTS
Relationships betewen cytoskeletal structures and cell underside organization innormally cultured and stage 1 spread cells
Indirect immunofluorescence on the spread fibroblasts using our antitubulinpreparation showed a characteristic lacy pattern similar to that reported by others(Weber, Rathke & Osborn, 1978) but no correlations could be seen between anyaspect of this pattern and the distribution of focal adhesions in any of the four celltypes tested (Fig. 1 c, D).

R. A. Badley, A. Woods, L. Carruthers and D. A. Rees

Cytoskeleton and adhesion 383
The relation between 10-nm filaments and the distribution of focal adhesions wasexamined in those cells which stained with our antibodies (BHK, CEF and a fractionof REF). In none of these could any correlation be found between the filaments andthe adhesions. Preliminary results with the guinea-pig antibodies confirmed theseresults for all the above cell types (Fig. 1 G, H).
In contrast to this lack of correlation between focal adhesions and patterns of eithermicrotubules or 10-nm filaments, there was a very obvious codistribution with actinstress fibres in that focal adhesions always occurred under stress fibres, most fre-quently near the termini (Fig. 1 A, B). Despite variations in size and shape of theadhesions and stress fibres which are often present in characteristic patterns thisconnexion was found in all the cell types. 16C cells tend to show small, black, circularadhesions separated by white areas. REF and CEF display larger and more elongatedadhesions with the white parts between less distinct and a greater tendency for stressfibres to appear as long, dark grey lines.
Another distinct form of actin, less frequently seen in fibroblast cultures, is the'geodesic dome' (Lazarides, 1976b) or 'polygonal net' (Osborn et al. 1978) which hasbeen suggested to be an intermediate stage towards the fully organized cytoskeleton(Lazarides, 19766) or a special form that develops in some cells only (Osborn et al.1978). We found that some cultures of rat embryo cells showed a high proportion ofthese structures. The immunofluorescence image could not be correlated with anypattern of focal contacts although underneath was often a grey zone or area of closecontact (in the sense defined in the Introduction) (Fig. IE, F). The absence of anyclear relationship with cell contacts to substratum is perhaps not surprising sincedifferential focusing confirms the result from stereo microscopy (Osborn et al. 1978)that these structures are mainly organized at the upper cell surface. Likewise it ispossible to find stress fibres high up in the cells which do not appear to be associatedwith a focal contact.
We have previously characterized cells spread on fibronectin in serum-free mediumas stage 1 by morphology and interference-reflexion microscopy. Such cells lack welldefined focal adhesions (Fig. 2G) compared to those normally cultured (Fig. 2H).By contrast the cell shape and the distribution of microtubules and 10-nm filamentsin the 2 states were found to be very similar (Fig. 2C-F). However, the actin frameworkas revealed by immunofluorescence with anti-actin was clearly different; actinbundles were often present but were much finer than usual (Fig. 2 A) compared to thethicker bundles usually seen in normally spread cells (Fig. 2B). Thus, the usualdevelopment of actin structures is incomplete where the development of focaladhesions is also incomplete, whereas the microtubule and 10-nm filament systemsorganize with cell shape in the usual way.
Fig. 1. Comparison of immunofluorescent cytoskeletal (A, C, E, G) and interference-reflexion (B, D, F, H) patterns of rat embryo (A-F) and BHK (G, H) cells, A, B, actinbundles; arrows indicate a single stress fibre attaching to several distinct adhesionzones along its length, c, D, microtubules. E, F, actin in geodesic domes; arrowsindicate no adhesion zone under geodesic dome. G, H, 10-nm filaments visualizedusing guinea-pig antibodies.

R. A. Badley, A. Woods, L. Carruthers and D. A. Rees
Fig. 2. Actin (A, B), tubulin (c, D), io-nm filament protein (E, F) and interference-reflexion patterns (G, H) in rat embryo (A-D, G, H) and BHK (E, F) cells spread onfibronectin in serum-free medium (A, C, E, G) and complete medium (B, D, F, H).

Cytoskeleton and adhesion 385
Cytoskeleton changes in detachment
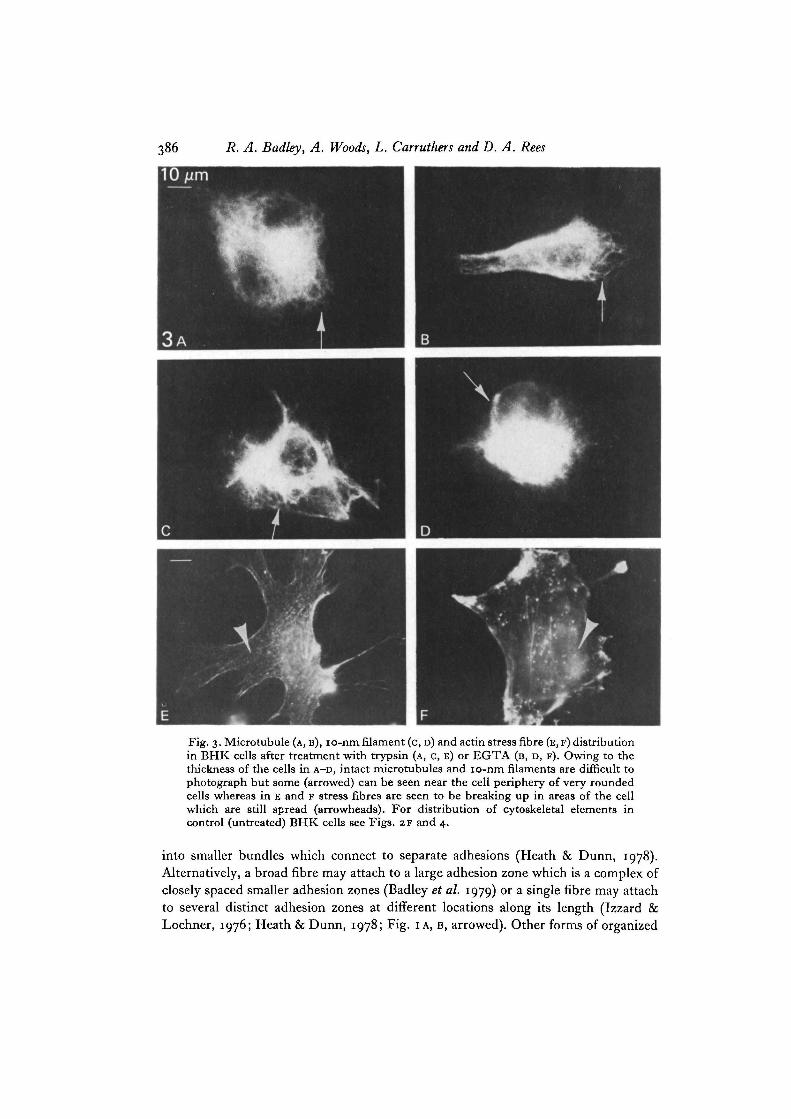
For these experiments we used BHK cells so that the results could be correlatedwith an earlier study of cell detachment by trypsin and EGTA (Rees et al. 1977).On treatment for 10 min with either agent the speed of withdrawal of cell marginsleading to cell rounding and detachment varied from cell to cell within the populationbut was accompanied by a characteristic alteration of each of the components of thecytoskeleton visualized independently by indirect immunofluorescence (Figs. 3, 4).The pattern of changes in microtubules and 10-nm filaments were similar to eachother and similar for both detachment agents. They draw back into the cell bodyfollowing cell shape changes. Fig. 3 A-D show cells in a late stage of rounding where theextent of cell spreading has been reduced by approximately half. Although cellrounding obviously distorts the microtubules, many of those closer to the nucleusremain intact (Furcht & Wendelschafer-Crabb, 1978; Osborn & Weber, 1977). Dueto the much greater cell thickness individual microtubules become difficult to resolveexcept at the edges of cells. Previous ultrastructural work, which concentrated on suchthinner regions, suggested that microtubules are lost after trypsin treatment (Reeset al. 1977) but the immunofluorescence studies now show that many of them remainunder our conditions. Much of the same behaviour is seen for the 10-nm filaments.They do not appear to disintegrate but, like the microtubules, become more convolutedand folded around the nucleus although the pattern becomes increasingly difficultto distinguish clearly as the optical thickness increases.
In contrast, the actin system is affected by trypsin and EGTA prior to cell shapechanges; the stress fibres appeared to break up rapidly in a process that startedindependently at a series of points along their length and those cells that had barelybegun to round were seen to have lost most of their stress fibres in areas of the cellwhich were still well spread (arrowheads in Fig. 3E, F). Many fibroblast types possess asubset of stress fibres located higher in the cell than those typically found close to thelower membrane (see, for example, Osborn et al. 1978). In BHK cells they are easilyidentified by differential focussing. EGTA appeared to cause the break up of boththese sets of fibres at similar rates. With trypsin, however, the stress fibres higher inthe cell or at the cell edge appeared more sensitive than those near the base. Manyexamples of cells lacking the upper set were found (compare Fig. 5 A and B) ; at a latestage of break up of either set of fibres with trypsin, a distinct beaded appearance canfrequently be seen (Fig. 5 c). Thus, trypsin and EGTA produce slightly differentsequences of changes in cytoskeleton patterns which are summarized in Table 1.
DISCUSSION
Our comparison of the pattern of focal adhesions with the organization of the 3components of the cytoskeleton - microtubules, 10-nm filaments and actin - confirmsthat it is the actin and specifically the stress fibres, which have the most direct relation-ship with the adhesive contacts of fibroblasts to substratum. A single stress fibre maybe associated with multiple adhesion points in several ways. A single fibre may split
386 R. A. Badley, A. Woods, L. Carruthers and D. A. Rees
Fig. 3. Microtubule (A, B), io-nm filament (c, D) and actin stress fibre (E, F) distributionin BHK cells after treatment with trypsin (A, C, E) or EGTA (B, D, F). Owing to thethickness of the cells in A-D, intact microtubules and io-nm filaments are difficult tophotograph but some (arrowed) can be seen near the cell periphery of very roundedcells whereas in E and F stress fibres are seen to be breaking up in areas of the cellwhich are still spread (arrowheads). For distribution of cytoskeletal elements incontrol (untreated) BHK cells see Figs. 2F and 4.
into smaller bundles which connect to separate adhesions (Heath & Dunn, 1978).Alternatively, a broad fibre may attach to a large adhesion zone which is a complex ofclosely spaced smaller adhesion zones (Badley et al. 1979) or a single fibre may attachto several distinct adhesion zones at different locations along its length (Izzard &Lochner, 1976; Heath & Dunn, 1978; Fig. IA, B, arrowed). Other forms of organized
Cytoskeleton and adhesion 387
10/im
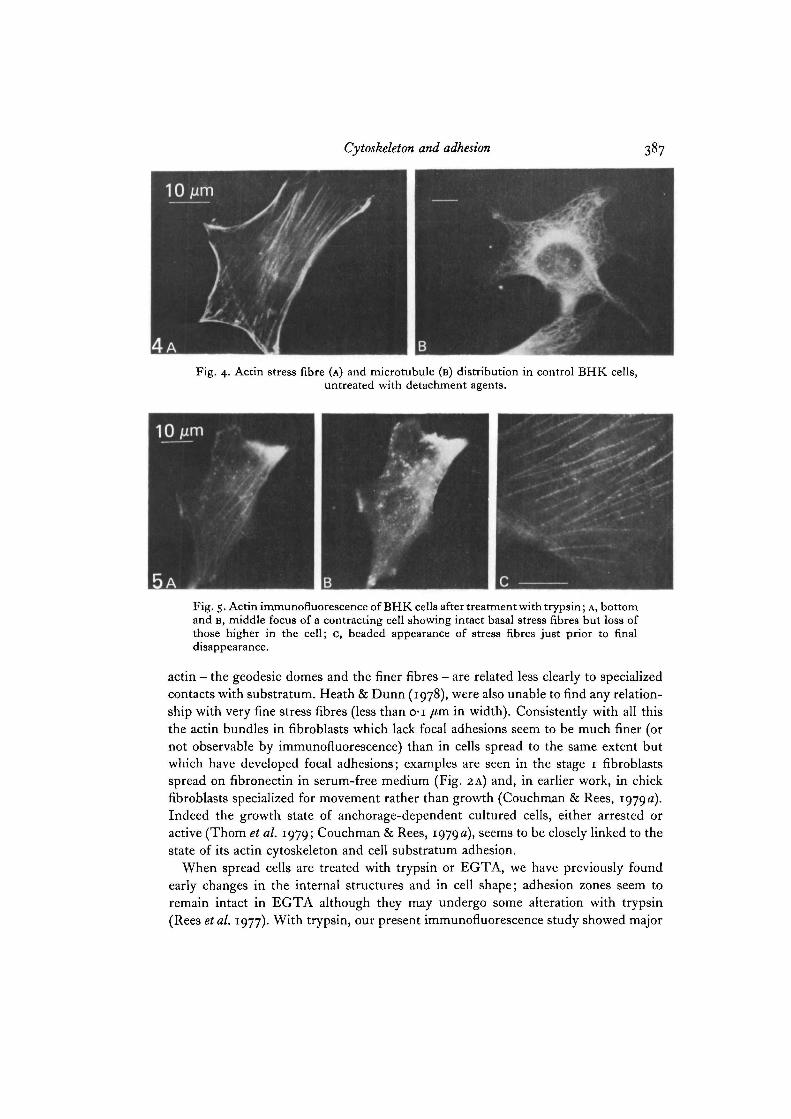
Fig. 4. Actin stress fibre (A) and microtubule (B) distribution in control BHK cells,untreated with detachment agents.
10jum
Fig. 5. Actin immunofluorescence of BHK cells after treatment with trypsin; A, bottomand B, middle focus of a contracting cell showing intact basal stress fibres but loss ofthose higher in the cell; c, beaded appearance of stress fibres just prior to finaldisappearance.
actin — the geodesic domes and the finer fibres - are related less clearly to specializedcontacts with substratum. Heath & Dunn (1978), were also unable to find any relation-ship with very fine stress fibres (less than o-i /tm in width). Consistently with all thisthe actin bundles in fibroblasts which lack focal adhesions seem to be much finer (ornot observable by immunofluorescence) than in cells spread to the same extent butwhich have developed focal adhesions; examples are seen in the stage 1 fibroblastsspread on fibronectin in serum-free medium (Fig. 2A) and, in earlier work, in chickfibroblasts specialized for movement rather than growth (Couchman & Rees, 1979 a).Indeed the growth state of anchorage-dependent cultured cells, either arrested oractive (Thorn et al. 1979; Couchman & Rees, 1979a), seems to be closely linked to thestate of its actin cytoskeleton and cell substratum adhesion.
When spread cells are treated with trypsin or EGTA, we have previously foundearly changes in the internal structures and in cell shape; adhesion zones seem toremain intact in EGTA although they may undergo some alteration with trypsin(Rees etal. 1977). With trypsin, our present immunofluorescence study showed major

388 R. A. Badley, A. Woods, L. Carruthers and D. A. Rees
Table i. The sequences of events during trypsin and EGTA detachment of BHKfibroblasts (earlier events listed at the top)
Trypsin EGTA
Upper stress fibres give beadedappearance
Upper stress fibres disappear andlower stress fibres begin to breakup to give beaded appearance
Lower stress fibres disappearMicrotubules and io-nm filamentsdraw back, some break up; cellmargins draw back
Cell roundsCell detaches
Upper and lower stress fibres breakup and disappear
Microtubules and io-nm filamentsdraw back, some break up; cellmargins draw back
Cell roundsCell detaches
changes in the stress fibres, consistent with the earlier results of Pollack & Rifkin(1975) but showing a number of additional features. The stress fibres at the base ofthe cell which we know to be associated with focal adhesions (Heath & Dunn, 1978;Badley et al. 1978), are relatively resistant and some may persist in shortened formeven in cells which have progressed to a rounded morphology before eventuallybreaking up. In contrast, those higher in the cell are relatively sensitive. Both sets areseen to break up with trypsin at points along their length to convert to an intermediatebeaded state. EGTA appears to affect both sets of stress fibres equally and the beadedintermediate state is not seen. These distinct patterns of disintegration would beconsistent with different mechanisms of action of the 2 agents and particularlysuggest that: (i) the action of trypsin is initiated by cell surface cleavages and that therelative rates of dispersal of individual fibres reflect the relative accessibilities of theirtarget sites since basal stress fibres seem more resistant than those higher in the cell;(ii) following proteolysis at the membrane site, disintegration of each fibre seems to bepropagated by preferential loss of material along its length giving rise to the beadedappearance seen in this immunofluorescence study; and (iii) EGTA acts more generallyon the stress fibre system rather than on individual fibres, probably as a result of thedepletion of intracellular Ca2+ so that we do not see sequential dispersal of one set offibres relative to another or the propagated dispersal of individual fibres from themembrane-attachment sites. We do not exclude the possibility that some of thetrypsin-induced changes are initiated intracellularly, since it is well known that thisenzyme is taken up by fibroblasts.
In contrast to the apparent relationships between actin organization and both thetype of cell substrate adhesion and growth state, we find that the microtubule andio-nm filament patterns show no corresponding relationships but seem to correlatemainly with cell shape. Thus stage 1 and fully spread cells both display similar arraysof the latter 2 cytoskeleton components which seem to follow, rather than precede,cell-shape changes during cell rounding.

Cytoskeleton and adhesion 389
In summary, the developed organization of microtubules and 10-nm filamentsseems to be linked to cell shape and, for example, show little change with changingpatterns of adhesion or growth status. The actin organization, on the other hand, doesreflect such additional variables. Further, agents which cause rounding and detach-ment first disperse the stress fibres whereas the microtubules and 10-nm filamentschange later along with cell shape. With trypsin the stress fibres seem to break downin a rather regular way, those higher in the cell disappearing earlier whereas withEGTA all the fibres appear to break up at similar rates and do riot show the inter-mediate beaded forms often seen with trypsin.
We thank Dr J. V. Small for a gift of skeletin, Miss Christine Evans for photography, MrsD. S. Cox for fibronectin samples and Miss S. Ord and Mrs H. M. Muir-Howie for cellcultures.
REFERENCES
ABERCROMBIE, M. & DUNN, G. A. (1975). Adhesions of fibroblasts to substratum duringcontact inhibition observed by interference reflection microscopy. Expl Cell Res. 92, 57-62.
ABERCROMBIE, M., HEAYSMAN, J. E. M. & PEGRUM, S. M. (1971). The locomotion of fibro-blasts in culture. IV. Electron microscopy of the leading lamella. Expl Cell Res. 67, 359-367.
BADLEY, R. A., LLOYD, C. W., WOODS, A., CARRUTHERS, L., ALLCOCK, C. & REES, D. A. (1978).Mechanisms of cellular adhesion. III. Preparation and preliminary characterisation ofadhesions. Expl Cell Res. 117, 231-244.
BADLEY, R. A., WOODS, A., SMITH, C. G. & REES, D. A. (1979). Expl Cell Res. (In Press).BENNETT, G. S., FELLINI, S. A., CROOP, J. M., OTTO, J. H., BRYAN, J. & HOLTZER, H. (1978).
Differences among 100-A filament subunits from different cell types. Proc. natn. Acad. Sci.U.S.A. 75, 4364-4368.
BEREITER-HAHN, J., FOX, C. H. & THORELL, B. (1979). Quantitative reflection contrast micro-scopy of living cells. J. Cell Biol. 82, 767-779.
BRINKLEY, B. R., FULLER, G. M. & HIGHFIELD, D. P. (1975). Cytoplasmic microtubules innormal and transformed cells in culture: Analysis by tubulin antibody immunofluorescence.Proc. natn. Acad. Set. U.S.A. 72, 4981-4983.
BUCKLEY, I. K. (1975). Three dimensional fine structure of cultured cells: possible implicationsfor subcellular motility. Tissue & Cell 7, 51-72.
COUCHMAN, J. R. & REES, D. A. (1979a). Actomyosin organisation for adhesion, spreading,growth and movement in chick fibroblasts. Cell Biol. int. Rep. 3, 431-439.
COUCHMAN, J. R. & REES, D. A. (1979&). Behaviour of fibroblasts migrating from chick heartexplants. Changes in adhesion, locomotion and growth and in the distribution of actomyosinand fibronectin. J. Cell Set. 39, 149-165.
CURTIS, A. S. G. (1964). The mechanism of adhesion of cells to glass. A study by interferencereflection microscopy. J. Cell Biol. 20, 199-215.
DAVISON, P. F., HONG, B.-S. & COOKE, P. (1977). Classes of distinguishable 10 nm cytoplasmicfilaments. Expl Cell Res. 109, 471-474.
FRANKE, W. W., SCHMID, E., OSBORN, M. & WEBER, K. (1978). Different intermediate-sizedfilaments distinguished by immunofluorescence microscopy. Proc. natn. Acad. Sci. U.S.A.75. S034-S038.
FURCHT, L. T. & WENDELSCHAFER-CRABB, G. (1978). Trypsin induced coordinate alterationsin cell shape, cytoskeleton and intrinsic membrane structure of contact-inhibited cells.Expl Cell Res. 114, 1-14.
GORDON, W. E., BUSHNELL, A. & BURRIDGE, K. (1978). Characterisation of the intermediate(10 nm) filaments of cultured cells using an autoimmune rabbit antiserum. Cell 13, 249-261.
HEATH, J. P. & DUNN, G. A. (1978). Cell to substratum contacts of chick fibroblasts and theirrelation to the microfilament system. A correlated interference-reflexion and high-voltageelectron-microscope study. J. Cell Sci. 29, 197-212.

39© R. A. Badley, A. Woods, L. Carruthers and D. A. Rees
HEGGENESS, M. H., WANG, K. & SINGER, S. J. (1977). Intracellular distributions of mechano-chemical proteins in cultured fibroblasts. Proc. natn. Acad. Sci. U.S.A. 74, 3883-3887.
HYNES, R. O. & DESTREE, A. T. (1978). 10 nm filaments in normal and transformed cells. Cell13, 151-163-
IZZARD, C. S. & LOCHNER, L. R. (1976). Cell-to-substrate contacts in living fibroblasts: aninterference-reflexion study with an evaluation of the technique. J. Cell Sci. 21, 129-159.
KEMP, R. B., JONES, B. M., CUNNINGHAM, I. & JAMES, M. C. M. (1967). Quantitative investiga-tion on the effect of puromycin on the aggregation of trypsin and versene-dissociated chickfibroblast cells. J. Cell Sci. 2, 323-340.
KURKI, P., LINDER, E., VIRTANEN, I. & STENMAN, S. (1977). Human smooth muscle auto-antibodies reacting with intermediate (100 A) filaments. Nature, hand. 268, 240-241.
LAZARIDES, E. (1976a). Two general classes of cytoplasmic actin filaments in tissue culturecells: the role of tropomyosin. J. supramolec. Struct. 5, 531-563.
LAZARIDES, E. (19766). Actin, a-actinin and tropomyosin interaction in the structural organisa-tion of actin filaments in non-muscle cells. J. Cell Biol. 68, 202-219.
LAZARIDES, E. (1978). The distribution of desmin (100 A) filaments in primary cultures ofembryonic chick cardiac cells. Expl Cell Res. 112, 265-273.
LAZARIDES, E. & BALZER, D. R. (1978). Specificity of desmin to avian and mammalian musclecells. Cell 14, 429—438.
LAZARIDES, E. & HUBBARD, B. D. (1976). Immunological characterisation of the subunit of the100 A filaments from muscle cells. Proc. natn. Acad. Sci. U.S.A. 73, 4344-4348.
LLOYD, C. W., SLABAS, A. P., POWELL, A. J., MACDONALD, G. & BADLEY, R. A. (1979).Cytoplasmic microtubules of higher plant cells visualised with anti-tubulin antibodies.Nature, Land. 279, 239-241.
OSBORN, M., BORN, T., KOITSCH, H. J. & WEBER, K. (1978). Stereo immunofluorescencemicroscopy. I. Three dimensional arrangement of microfilaments, microtubules and tono-filaments. Cell 14, 477-488.
OSBORN, M. & WEBER, K. (1977). The detergent-resistant cytoskeleton of tissue culture cellsincludes the nucleus and the microfilament bundles. Expl Cell Res. 106, 339-349.
POLLACK, R. & RIFKIN, D. (1975). Actin-containing cables within anchorage dependent ratembryo cells are dissociated by plasmin and trypsin. Cell 6, 495-506.
REES, D. A., LLOYD, C. W. & THOM, D. (1977). Control of grip and stick in cell adhesionthrough lateral relationships of membrane glycoproteins. Nature, Land. 267, 124-128.
STARGER, J. M., BROWN, W. E., GOLDMAN, A. E. & GOLDMAN, R. D. (1978). Biochemical andimmunological analysis of rapidly purified 10 nm filaments from baby hamster kidney(BHK-21) cells, y. Cell Biol. 78, 93-109.
STARGER, J. M. & GOLDMAN, R. D. (1977). Isolation and preliminary characterisation of 10 nm.filaments from baby hamster kidney (BHK-21) cells. Proc. natn. Acad. Sci. U.S.A. 74,2422-2426.
THOM, D., POWELL, A. J. & REES, D. A. (1979). Mechanisms of cellular adhesion. IV. Role ofserum glycoproteins in fibroblast spreading on glass. J. Cell Sci. 35, 281-305.
WEBER, K., POLLACK, R. & BIBRING, T. (1975). Antibody against tubulin: the specific visualisa-tion of cytoplasmic microtubules in tissue culture cells. Proc. natn. Acad. Sci. U.S.A. 72,
WEBER, K., RATHKE, P. C. & OSBORN, M. (1978). Cytoplasmic microtubular images in glutar-aldehyde-fixed tissue culture cells by electron microscopy and by immunofluorescencemicroscopy. Proc. natn. Acad. Sci. U.S.A. 75, 1820-1824.
(Received 10 October 1979 - Revised 12 December 1979)





























