Embed Size (px)
Citation preview

DMD # 88526
1
CYP2D1 Gene Knockout Reduces the Metabolism and Efficacy of Venlafaxine in
Rats
Hongqiu Zhoua , Li Yang
a , Changsuo Wang
a , Zhiqiang Li
a , Zhen Ouyang
a , Mangting
Shanb , Jun Gu
c,*, Yuan Wei
a ,*
aSchool of Pharmacy, Jiangsu University, Zhenjiang, Jiangsu 212013, China
bMtC BioPharma Co. Ltd, Nanjing, Jiangsu 210042, China
cWadsworth Center, New York State Department of Health, and School of Public
Health, State University of New York at Albany, Albany, NY 12201, USA
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
2
Running Title: CYP2D1-knockout rat model
*Corresponding Author: Yuan Wei*
Mailing address: School of Pharmacy, Jiangsu University, 301 Xuefu Road, Jingkou
Zone, Zhenjiang 212013, Jiangsu, China
Tel/Fax.: +86-511-85038750
Email: [email protected]
*Corresponding Author: Jun Gu*
Mailing address: Wadsworth Center, New York State Department of Health, Empire
State Plaza, Albany, NY 12201-0509, USA
Email: [email protected]
Number of Text pages: 31
Number of Tables: 5
Number of Figures: 4
Number of References: 58
Number of Words:
Abstract: 249
Introduction: 642
Discussion: 1499
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
3
Abbreviations: ANOVA, analysis of variance; AUC, area under the curve of the drug;
AUC0-t, AUC from the time of administration to the last point; AUC0-∞, time from the
start of administration to the theoretical extrapolation AUC; AhR, aryl hydrocarbon
receptor; CYPOR, NADPH-cytochrome P450 oxidoreductase; CRISPR, clustered
regularly interspaced short palindromic repeat; CMS, chronic mild stimulations; Ctrl,
control groups; CLint, intrinsic clearance; CL/F, apparent total clearance; Cmax, peak
concentration; CAR, constitutive androgen receptor; EM, extensive metabolizers;
H&E, Hematoxylin and Eosin; 5-HT, 5-hydroxytryptamine; HPLC, high-performance
liquid chromatography; Km, Michaelis constant; NE, norepinephrine; ODV,
O-desmethyl venlafaxine; PM, poor metabolizer; PXR, Pregnane X receptor; UCSC,
University of California Santa Cruz; VLF, venlafaxine; Vd/F, apparent volume of
distribution; WT, wild-type.
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
4
Abstract
Rat CYP2D1 has been considered as an ortholog of human CYP2D6. To
assess the role of CYP2D1 in physiological processes and drug metabolism, a
CYP2D1-null rat model was generated with a clustered regularly interspaced
short palindromic repeat (CRISPR)/Cas9 method. Seven base-pairs were deleted
from the exon 4 of CYP2D1 of Sprague-Dawley wild-type (WT) rats. The
CYP2D1-null rats were viable and showed no abnormalities in general
appearance and behaviors. Metabolism of venlafaxine (VLF) was further studied
in CYP2D1-null rats. Vmax and CLint of the liver microsomes in vitro from
CYP2D1-null rats were decreased (decreased by ~46% and ~57% in males,
~47% and ~58% in females), while Km was increased (increased by ~24% in
males, ~25% in females) compared to those of WTs. In the pharmacokinetic
studies, compared to the WTs, VLF in the CYP2D1-null rats had significantly
lower Cl/F and Vd/F (decreased by ~36% and ~48% in males, ~23% and ~25%
in females), significantly increased AUC0-t, AUC0-∞, and Cmax (increased by ~64%,
~59% and ~26% in males, ~43%, ~35% and ~15% in females). In addition,
O-desmethyl venlafaxine formation was reduced as well in the CYP2D1-null rats
compared to that in WTs. Rat depression models were developed by feeding
separately and chronic mild stimulations with CYP2D1-null and WT rats. VLF
showed a better efficacy in the WT depression rats compared to that in the
CYP2D1-null ones. In conclusion, a CYP2D1-null rat model was successfully
generated, and CYP2D1 was found to play a certain role in the metabolism and
efficacy of venlafaxine.
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
5
Significance Statement
A novel CYP2D1-null rat model was generated using CRISPR/Cas9 technology,
and it was a valuable tool to study the in vivo function of human CYP2D6. Moreover,
our data suggested that the reduced ODV formation was associated with a lower VLF
efficacy in rats.
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
6
Introduction
Cytochrome P450 enzymes are part of an extended family of hemoproteins and
are an important family of oxidases in the microsomal mixed-function oxidase system
(Yamazaki, 2014). To date, a total of 57 cytochrome P450 isoforms have been
identified in humans, which are grouped into 18 families and 44 subfamilies by their
sequence similarity (Zanger et al., 2008). Among them, CYP1, CYP2, and CYP3
subfamilies are responsible for the biotransformation of most of the foreign
substances including 70−80% of the clinically used drugs (Zhou et al., 2012;
Guengerich, 2008).
Although being only 4% of the total cytochrome P450 enzyme in the human liver,
CYP2D6 is associated with biotransformation of approximately 30% of the
commonly used drugs (Teh and Bertilsson, 2012). Substrates of CYP2D6 include a
variety of antiarrhythmics, antipsychotics, and second-generation and tricyclic
antidepressants (Zanger and Schwab, 2013). It has been reported that the frequency of
CYP2D6 allele varies by racial/ethnic groups, about 6−10% of the Caucasians and
only 2% of the Asians are CYP2D6 poor metabolizers (PMs) (Meyer et al., 1990;
Bradford, 2002). If the amount of a drug administered to the CYP2D6 PMs is the
same as that administered to normal individuals, serious adverse reactions are more
likely to occur in the PMs because the metabolism of the drug is often unpredictable
in the PMs (De Leon et al., 2005; Hertz et al., 2015). Further research is needed in
order to improve the efficacy of the drugs, reduce the incidence of adverse reactions,
and achieve an individualized drug therapy.
Venlafaxine (VLF) is an antidepressant of selective serotonin and norepinephrine
reuptake inhibito (Bymaster, 2001). In humans, VLF was metabolized to O-desmethyl
venlafaxine (ODV) by CYP2D6 (~89%), CYP2C19 (~10%), and CYP2C9 (~1%),
venlafaxine is also metabolized by CYP3A4 to N-desmethyl venlafaxine (Magalhães
et al., 2014). ODV is the main metabolite of VLF to exert pharmacological activity
(Wellington and Perry, 2001). For rat CYP2D subfamily, it was reported that 6
members (CYP2D1, -2D2, -2D3, -2D4, -2D5, and -2D18) have been identified by
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
7
genomic analysis and CYP2D1 was considered as an ortholog of human CYP2D6
(~83% homology) (Nelson et al., 1996; Martignoni et al., 2006).
A variety of cytochrome P450 gene knockout mouse models and CYP-humanized
mouse models have been reported to illustrate the function of cytochrome P450
isoforms on drug metabolism, toxicity and carcinogenicity (Scheer et al., 2012, 2014;
Bissig et al., 2018). However, mouse model may have some limitations. For example,
the small volume of blood samples collected from mice increases the difficulty of
pharmacokinetic studies. Moreover, the mouse model is poorly tolerated in some
experiments, such as bacterial infections experiment, chronic heart failure experiment
(Robinson et al., 1968; Li et al., 2004; Handa et al., 2009). Compared to the mouse
model, the rat model could be more suitable for certain experiments.
In 2013, a knockout rat model was generated with clustered regularly interspaced
short palindromic repeat (CRISPR)/Cas9 method (Li et al., 2013), and since then
knockout rats have become an accessible tool for studying human cytochrome P450
functions. It has been reported that the metabolism of chlorzoxazone is significantly
reduced in CYP2E1 knockout rats (Wang et al., 2016), while the metabolism of
nifedipine and midazolam were reduced as well in CYP3A1/2 double knockout rats
(Lu et al., 2017). A CYP2C11 knockout rat model was successfully generated by our
group and was used to study the metabolism of tolbutamide and warfarin (Wei et al.,
2018; Ye et al., 2019).
For a better understanding the role of human CYP2D6 in drug metabolism, in this
study, a unique CYP2D1-null rat model was generated using the CRISPR/Cas9
technology. The general phenotypes were characterized and the hepatic expression of
major P450s was determined in the knockout rat model. Venlafaxine, an inhibitor of
serotonin-norepinephrine reuptake and a substrate of human CYP2D6, was used as a
probe drug to study the metabolism and efficacy function of CYP2D1 in rats.
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
8
Materials and methods
Chemicals and reagents. NADPH (> 98% purity) was purchased from Aladdin
Industrial Co. Ltd. (Shanghai, China). Carvedilol (> 99% purity) was purchased from
Aladdin Bio-Chem Technology Co. Ltd. (Shanghai, China). Venlafaxine
hydrochloride and O-desmethyl venlafaxine (> 98% purity) were purchased from Sam
Chemical Technology Co., Ltd. (Shanghai, China). Acetonitrile and methanol, as well
as all other reagents, were purchased from Sinopharm Chemical Reagent Co. Ltd.
(Shanghai, China).
Generation of the CYP2D1-null rats by CRISPR/Cas9. Three specific sgRNA
target sequences were selected from the target site in exon 4 of the rat CYP2D1 gene
(ID: 266684) using Gene Knock-Out with Cas9 software from Xunqi Biotechnology
Co. Ltd. (Nanjing, China). Three pairs of guiding primers were designed: sgRNA1
(forward primer: 5′–GCAGCATGGCCTTGGGATTGA–3′, reverse primer:
5′–TCAATCCCAAGGCCATGCTG–3′),sgRNA2 (forward primer:
5′–AGACCCTTACCTCATCAGGA–3′, reverse primer:
5′–TCCTGATGAGGTAAGGGTCT–3′), and sgRNA3 (forward primer:
5′–CTAGTTTCACCATCCTGATG–3′, reverse primer:
5′–CATCAGGATGGTGAAACTAG–3′). The sgRNAs were transcribed in vitro using
the T7 Quick High Yield RNA Synthesis Kit, recovered by phenol-chloroform
extraction and alcohol precipitation, and finally dissolved in nuclease-free water. The
pST1374-cas9 vector was linearized by Agel (New England Bioscience, Ipswich, MA,
USA), transcribed into Cas9 mRNA, and purified with RNeasy Mini Kit (Qiagen,
Valencia, CA). The constructed Cas9 mRNA was co-injected with sgRNA1, sgRNA2,
and sgRNA3 (Cas9 mRNA: 50 ng/μL, sgRNA1: 100 ng/μL, sgRNA2: 100 ng/μL, and
sgRNA3: 100 ng/μL) into zebrafish zygotes to determine their activities, respectively.
The sgRNA3 set primer (30 ng/μL) was injected into the wild-type (WT)
Sprague-Dawley rat monocytic embryos with Cas9 mRNA (50 ng/μL) as sgRNA3
showed the highest activity in zebrafish (Supplementary Table 1). The obtained
CYP2D1+/-
rat (F0, 1♀) was hybridized with WT rat (1♂) and the pups were subjected
to PCR amplification and genomic DNA sequencing to select the heterozygous
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
9
CYP2D1+/-
rats (F1). The F1 generation rats were inbred (1♂: 2♀) to produce F2
generation rats, and then the homozygous CYP2D-/-
rats were screened out from the F2
generation rats by the same method. The homozygous CYP2D-/-
rats were transferred
from the Model Animal Research Center of Nanjing University to the Laboratory
Animal Center of Jiangsu University for further breeding and animal experiments.
The animal facilities of Nanjing University and Jiangsu University have passed the
certification of the Association for Assessment and Accreditation of Laboratory
Animal Care. All animal breeding and experimental procedures were performed in
accordance with the Guide for the Care and Use of Laboratory Animals and were
approved by the Institutional Animal Care and Use Committees of Nanjing University
and Jiangsu University.
Off-target analysis. The sgRNA3 sequence (CTAGTTTCACCATCCTGATG)
was submitted to TagScan (http://ccg.vital-it.ch/tagger) to predict the possible
off-target sites. The detailed sequences around the potential off-target sites were
obtained from the University of California, Santa Cruz (UCSC) database
(http://genome.ucsc.edu/). The corresponding primers were designed for the potential
off-target sites and PCR was performed. The PCR products were then sequenced by
Sangon Biotech Co. Ltd. (Shanghai, China) and compared to those from UCSC
database.
General characterization of the CYP2D1-null rats. The appearance, fertility, and
general behaviors of CYP2D1-null rats were observed. CYP2D1-null rats at
12-week-old (6 rats per group, male and female) were sacrificed by exposure to an
ascending concentration of CO2 and dissected for further study. The viscera indices
(defined as the ratio of tissue weight to body weight) were calculated and compared to
those of WT rats.
Analysis of the expression levels of major P450s. The rat liver microsomes were
prepared by differential centrifugation using a previously described method (Egorin et
al., 1997). The concentration of liver microsomes was determined using the Enhanced
Bicinchoninic Acid Protein Assay Kit (Beyotime, Nantong, China) and the expression
of major subtypes of cytochrome P450 in the CYP2D1-null and WT rats were
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
10
determined by western blotting. The common loading controls such as GAPDH or
β-actin could not be used in the experiment since most of them had been removed
during microsomal preparation. Therefore, we used Ponceau S stain on the duplicate
gel to guarantee the loading amount of protein per lane (5 µg/lane) was consistent (Gu
et al., 2003). The blots were incubated with CYP1A2 (sc-53241, mouse monoclonal
IgG1, Santa Cruz Biotechnology, CA, USA), CYP2A (PAB19502, rabbit polyclonal,
Abnova, Taipei, Taiwan, China), CYP3A (sc-25845, rabbit polyclonal, Santa Cruz
Biotechnology, CA, USA), CYP2B (sc-73546, mouse monoclonal IgG1, Santa Cruz
Biotechnology, CA, USA), CYP2C11 (PA3-034, rabbit polyclonal, Thermo Fisher
Scientific, Waltham, MA, USA), CYP2D1 (AB1271, rabbit polyclonal, EMD
Millipore Corporation, Single Oak Drive, Temecula, CA 92590, USA), CYP2E1
(BML-CR3271-0100, rabbit polyclonal, Enzo, NY, USA) and CYPOR (sc-55477,
mouse monoclonal, Santa Cruz Biotechnology, CA, USA) antibodies, respectively, at
room temperature for 2 h, followed by incubation with the corresponding secondary
antibodies for 1 h. Chemiluminescence solution was added to the blot and the
ChemiDoc XRS imaging system (BioRdad, Hercules, CA, USA) was used to
visualize the images.
mRNA analysis of CYP2D genes. Total RNA was isolated from livers of WT and
CYP2D1-null rats using Trizol reagent (CW0580S, CWBIO, Beijing, China)
according to the instruction of the manufacturer. To detect selected CYP expression in
mRNA level, cDNA was prepared from total RNAs using the RevertAid First Strand
cDNA Synthesis kit (K1622, Thermo Fisher Scientific, MA, USA). β-actin was set as
the internal reference. Selected genes for quantification in livers of WT and
CYP2D1-null rats were CYP2D1, -2D2, -2D3, -2D4/18,-2D5 and primers designed for
quantification were listed in Supplementary Table 2.
VLF metabolism in the CYP2D1-null rats. Liver microsomes (1 mg/mL) and
various concentrations of VLF (7.5 μM, 10 μM, 15 μM, 20 μM, or 25 μM) were
mixed and pre-incubated at 37 °C water bath for 5 min, followed by adding
β-NADPH (1 mM) and MgCl2 (5 mM) to initiate the reaction in a total volume of 200
μL and incubated at 37 °C water bath for 50 min. The reaction was then stopped with
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
11
800 μL of ice-cold ethyl acetate containing 50 μg/mL carvedilol, which was used as
internal standard solution. The upper organic phase was collected by vigorous
vortexing for 3 min and centrifugation at 9, 600 g for 10 min at 4 °C. Then 400 μL of
ethyl acetate was used to extract the precipitate for the second time. The combined
organic phase was dried with nitrogen and reconstituted with 100 μL acetonitrile for
high-performance liquid chromatography (HPLC) analysis.
For determination of the pharmacokinetic parameters, CYP2D1-null and WT rats
were divided into 4 groups (6 rats per group, 12-week-old, male and female). The rats
were fasted for 12 h before the experiment but were provided with water. After
administration of a single dose of VLF (20 mg/kg, intragastric administration),
approximately 0.3 mL blood was collected from the orbital vein at different time
points (0.25, 0.5, 1, 2, 4, 8, 12, 24, and 36 h) in both CYP2D1-null and WT rats. The
blood samples were centrifuged at 1,500 g for 10 min to obtain plasma. 50 μL of 1.0
M hydrochloric acid solution and 10 μL of 50 μg/mL carvedilol (internal standard)
were added into 100 μL plasma and thoroughly mixed. Then 200 μL of ethyl acetate
was added and vortexed for 3 min. The supernatant was collected by centrifugation at
9, 600 g for 10 min. The precipitate was re-extracted with 200 μL ethyl acetate. The
combined supernatant was dried with nitrogen, and re-dissolved in 100 μL acetonitrile
for HPLC analysis.
Agilent Eclipse plus C18 (5 μm, 4.6 250 mm, Agilent Technologies, Santa Clara,
CA, USA) column was used at room temperature. The flow rate was 0.8 mL/min and
the wavelength for detection was set at 228 nm. The mobile phase consisted of 60%
acetonitrile and 40% 0.01 mol/L ammonium acetate buffer (pH 4.8), and the injection
volume was 20 μL.
Development of rat model of depression. The development of the depression
model was based on previous literatures and in accordance with our laboratory
conditions (Pucilowski et al., 1993; Zhao et al., 2008). Briefly, the depression models
in WT rats and CYP2D1-null rats were developed by feeding separately and chronic
mild stimulations (CMS). In the first and second week, the CMS groups were given 8
kinds of stimulations, including tail suspension for 5 min, rat cage tilting at 45°,
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
12
humid padding, overcrowding, electric shock (36 V, 2 times/min), forced swimming,
alternated light and dark, and fasting and water deprivation for 24 h. From day 15 to
day 20, ice water (4 °C, 5 min) was added in the bath coupled with heat stress (45 °C,
5 min). The same kind of stimulation was intermittently used to prevent the rats from
anticipating the upcoming stimulation (Supplementary Table 3). During the
development of rat model of depression, all CMS rats were kept feeding with 1 rat per
cage except in the overcrowding stimulation experiment. The control groups were
bred normally with 5 rats per cage.
Body weight test. The effects of depression on body weight in rats were
investigated with reference to a previous literature (Liu et al., 2011). Rats in each
group were weighed at 9:00 am on day 21 of depression modeling.
Locomotor activity test. The experiment was performed according to the method
from a previous report (Zhao et al., 2008). On day 21 of depression modeling, rats
were individually placed in an open box of 80 cm 80 cm 50 cm. A TSE
multi-function conditioning system (TSE Systems, Thuringia, Germany) was used to
automatically record the trajectory of the movement within 8 min and calculate the
total distance of locomotor activity.
1% sucrose water consumption test. The experiment was performed according to
a previous literature (Kim et al., 2003). After 4 h of locomotor activity test, the rats
were fasted with no food or water for 24h, and then 1% sucrose water was provided to
rats for another 24 hours. The 1% sucrose water consumption for 24 h was determined
with the ratio of intake volume (mL) to rat body weight (100 g).
Efficacy of venlafaxine hydrochloride in the rat model of depression. After CMS
modeling, the rats (5 rats per group, 12-week-old, male and female) were
administered with venlafaxine hydrochloride (CMS VLF groups, 20 mg/kg,
intragastrical administration) or 2 mL 0.9% saline (CMS Saline groups) every day
for 14 days. Then body weight, locomotor activity and 1% sucrose water consumption
were also evaluated. After those analyses, approximately 1.0 mL of blood was taken
from the orbital vein of each rat, and plasma was prepared by 1,500 g centrifugation
for 20 min. The levels of norepinephrine (NE) and 5-hydroxytryptamine (5-HT) in
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
13
plasma were determined by an ELISA kit (SenBeiJia, Nanjing, China). The rats were
sacrificed by CO2 after blood sampling.
Statistical analyses. The experimental data were analyzed using Prism 5.0
statistical software (GraphPad, La Jolla, CA, USA). The experimental data were
presented as the mean ± standard deviation (SD). One-way analysis of variance
(ANOVA) was performed and a P value of < 0.05 was considered statistically
significant.
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
14
Results
Generation of CYP2D1-null rats. Two rat founders with different genotypes, 7-base
pairs (ACCTCAT) deletion or 1-base pair (A) insertion in exon 4, were generated.
Due to the infertility of the other one (with 1-bp insertion), only the founder with 7-bp
deletion was used for further studies. The generation scheme of CYP2D1-null rat
model was shown in Fig. 1A. The PCR amplification results of the CYP2D1-null and
WT rats are shown in Fig. 1B. The DNA fragments showed bright bands around 586
bp. The homozygous CYP2D1-null rats lacked the 7 bp (ACCTCAT) at 270 ~ 280 bp
as compared to the WT rats, and the heterozygous CYP2D1-null rats presented mixed
peaks in corresponding area (Fig. 1C). Based on the genotyping results, CYP2D1-null
rats were successfully generated and the genotype-stable offspring were obtained.
Off-target analysis. Nine potential off-target sites of sgRNA3 (Supplementary
Table 4) were predicted using the off-target detection software and the corresponding
primers (Supplementary Table 5). The sequencing results showed that those potential
off-target sites were not mutated in the CYP2D1-null rats.
General characterization of the CYP2D1-null rats. There were no significant
differences in the appearance, fertility, and general behaviors between the
CYP2D1-null and WT rats, in either males or females (Supplementary Table 6-7). The
data of viscera indices were shown in Table 1. The viscera indices of CYP2D1-null
rats also showed no significant difference compared to those of WTs.
Hepatic expression levels of major P450s. Hepatic expression levels of major
P450s in CYP2D1-null and WT rats were shown as Fig. 2. CYP2D1 protein was only
detected in the WTs, but not in the CYP2D1-null rats. The expression levels of the
other P450s showed no significant differences, except for the expression of CYP2A,
which was significantly higher in the CYP2D1-null rats compared with that in the WT
rats, for both males and females. Notably, CYP2C11 was detected neither in the WT
females nor in CYP2D1-null females.
mRNA expression levels of CYP2D genes. Greatly reduced mRNA level of
targeted CYP2D1 gene was detected in the livers of the CYP2D1-null model
(Supplementary Fig. 1). Among the mRNA expression of other CYP2D genes,
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
15
CYP2D2 and CYP2D3 were up-regulated, CYP2D4/18 and CYP2D5 were
down-regulated (Supplementary Fig. 2).
Venlafaxine hydrochloride metabolism in the CYP2D1-null rats. The VLF in vitro
activities were determined with liver microsomes from both WTs and the
CYP2D1-null rats. The kinetic parameters of Michaelis constant (Km), maximum
velocity (Vmax), and intrinsic clearance (CLint) were calculated and shown in Table 2.
There is no sex difference in the model, the Vmax and the CLint of CYP2D1-null males
decreased by 46% (P < 0.05) and 57% (P < 0.05), respectively, while the Km increased
by 24% (P < 0.05), compared to those in WT males. The Vmax and the CLint of the
CYP2D1-null females decreased by 47% (P < 0.05) and 58% (P < 0.05), respectively,
while Km increased by 25%, compared to those in WT females.
In the pharmacokinetic study, the plasma levels of VLF in the CYP2D1-null rats
were significantly higher than those in the WT rats (at the marked time points, Fig. 3A,
B), while the levels of O-desmethyl venlafaxine (ODV) were significantly lower than
those in the WT rats (at the marked time points, Fig. 3C, D). Compared to the WT
males, VLF in the CYP2D1-null males had significantly lower Cl/F (decreased by
~36%, P < 0.05) and Vd/F (decreased by ~48%, P < 0.05), significantly increased
AUC0-t (increased by ~64%, P < 0.05) and AUC0-∞ (increased by ~59%, P < 0.05),
Cmax (increased by ~26%, P < 0.05). VLF in the CYP2D1-null females also had
significantly lower Cl/F (decreased by ~23%, P < 0.05) and Vd/F (decreased by ~25%,
P < 0.05), and had a significantly higher AUC0-t (increased by ~43%, P < 0.05),
AUC0-∞ (increased by ~35%, P < 0.05), and Cmax (increased by ~15%, P < 0.05), as
compared to those of the WTs (Table 3).
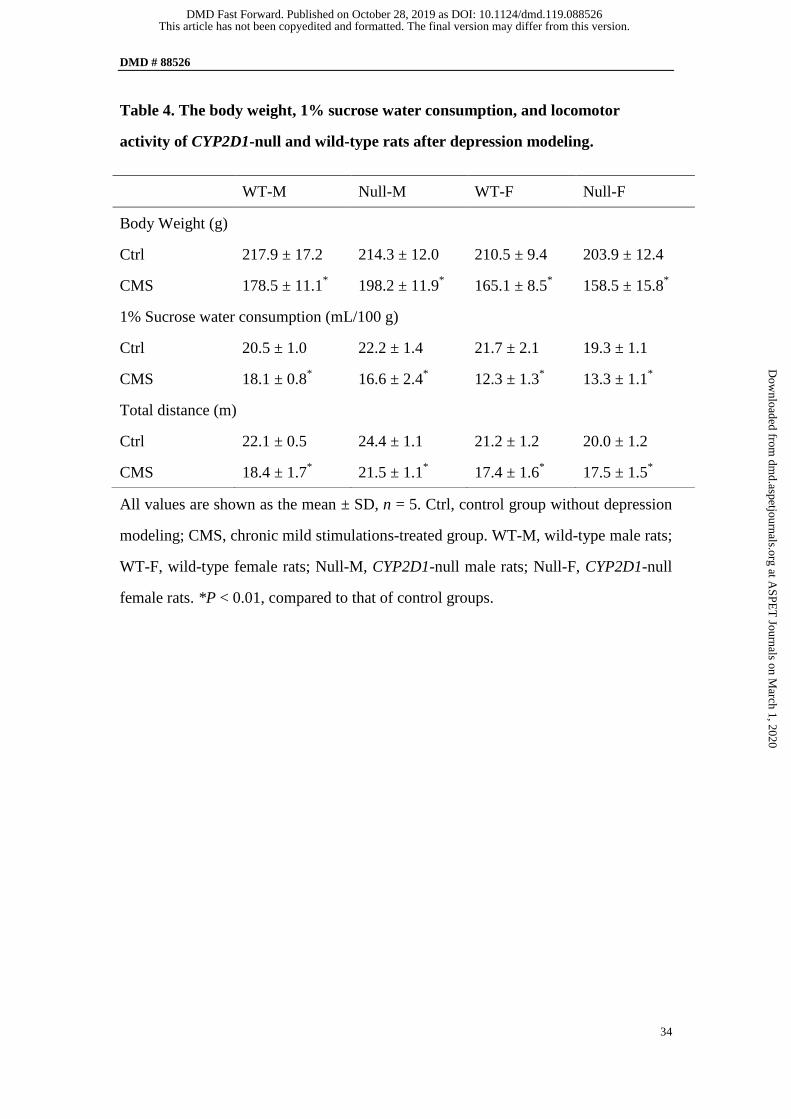
Body weight, 1% sucrose water consumption, and locomotor activity after
depression modeling. There were significant differences (P < 0.05) in body weight,
1% sucrose water consumption, and locomotor activity in CMS groups as compared
to those in control groups (Table 4). This result suggested that the depression
modeling in both strains was successful.
Efficacy of venlafaxine hydrochloride in the rat depression model. After 14 days
of administration of VLF, three depression indicators (e.g. body weight, 1% sucrose
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
16
water consumption, and locomotor activity) were recorded and shown in Table 5. All
the three indicators in VLF-treated groups (CMS + VLF groups) were significantly
increased than those saline-treated ones (CMS + Saline groups) for WT rats (P <
0.05). Those indicators were slightly improved (P > 0.05) in VLF-treated
CYP2D1-null rats compared with those of saline-treated groups, except for the 1%
sucrose water consumption in CYP2D1-null females. With VLF treatments, those
indicators in WTs were significantly higher (P > 0.05) than those in the CYP2D1-null
rats. Furthermore, with VLF treatments, plasma NE and 5-HT levels in WTs were
significantly higher than those in the CYP2D1-null rats, in either males or females
(Fig.4).
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
17
Discussion
Rat and human CYP2D isoforms share a good homology (> 70%), and rat CYP2D1
(~83% homology) is known as an ortholog of human CYP2D6 (Venhorst et al., 2003;
Martignoni et al., 2006). Rat CYP2D1 shares high homology with other CYP2D genes
(e.g. CYP2D2, ~82%; -2D3, ~85%; -2D4, ~80%; -2D5, ~98% and -2D18, ~80%),
which makes it difficult to develop a CYP2D1 knockout model in rats. In this study,
the exon 4 was found to be a suitable target for the CRISPR/Cas9 technology. We
analyzed the coding sequence of CYP2D1 gene and found that the deletion of
ACCTCAT would lead to truncated protein with only 213 amino acids, while the WT
CYP2D1 had 511 amino acids.
Previous studies using CRISPR/Cas9 have reported that off-target effects may
occur in the sgRNAs of mice and rats, which could also be inherited by the offspring
(Tan et al., 2015; Anderson et al., 2018). We used TagScan to predict the potential
off-target sites of sgRNA3 sequences in the CYP2D1-null rats and confirmed there
were no mutations in those potential off-target sites. Whether the mutation of
CRISPR/Cas9 occurred in non-predicted sites was necessary for further study due to
various factors.
In this study, the expression of CYP2C11 protein was detected only in male
CYP2D1-null rats (same as WT), so the knockout of CYP2D1 had little effects on the
male-specifically expression of CYP2C11. There was no significant difference in the
expression levels of other P450s, except the expression levels of CYP2As were higher
in the CYP2D1-null rats as compared to those in the WT rats. The rat CYP2A family
consisted of CYP2A1, -2A2 and -2A3 (Su and Ding, 2004). In the liver of rats,
CYP2A1 and CYP2A2 were dominantly expressed in females and males, respectively
(Haduch et al., 2005). CYP2A1 is responsible for the 7α-hydroxylation of
testosterone, while CYP2A2 catalyzes the 15α- and 7α-hydroxylation of testosterone
(Martignoni et al., 2006). The induction mechanism of CYP2A genes is still not very
clear. To the best of our knowledge, very few endogenous substances were identified
to regulate the expression of CYP2A genes in rats, except sexually dimorphic growth
hormone had been reported to up-regulate the expression of CYP2A1 and CYP2A2
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
18
(Waxman et al., 1995; Thangavel et al., 2004). Since the expression pattern of
CYP2C11 was not changed in the CYP2D1-null rat model, it was unlikely that the
induction of CYP2A was caused by growth hormone. Foreign compounds, including
phenobarbital, dexamethasone, and 3-methylcholanthrene, were found to up-regulate
CYP2A1 (Ryan and Levin, 1990), and pregnane X receptor (PXR), constitutive
androstane receptor (CAR), and aryl hydrocarbon receptor (AhR) could also be
involved in the regulation of CYP2As (Wallace and Redinbo, 2013). The specific
roles of those nuclear receptors in CYP2D1-null rats need to be further studied.
In human beings, it was reported that after administration of 150 mg VLF to 141
healthy subjects, the Cmax, AUC0-t, and AUC0-∞ of VLF in PMs were increased by 83%,
189%, and 212%, respectively as compared to those of extensive metabolizers (EMs).
The t1/2 was prolonged by 71%, and Cl/F was decreased by 68% (Kandasamy et al.,
2010). In another study, 14 healthy volunteers received 75 mg VLF extended release
formulations, the mean Cmax and AUC of VLF were increased by 149% and 331% in
the PMs compared those in the EMs, while Cl/F was decreased by 83% (Preskorn et
al., 2009). However, in our VLF metabolism study with rat models, the Cmax, AUC0-t,
and AUC0-∞ of VLF in the CYP2D1-null males were only increased by 24%, 64%, and
59%, respectively, as compared with those of the WT males; while Cl/F and Vd/F
were decreased by 36%, 48%, respectively. The difference of VLF metabolism
between CYP2D1-null and WT rats seemed much smaller than that between human
PMs and EMs. This fact suggested that other 2D isozymes might also contribute to
VLF metabolism, and the broad use of the current knockout model was limited.
Therefore, a rat model with multiple knockouts of CYP2D genes would be a better
approach to understand the VLF metabolism in humans.
The CRISPR/Cas9 system could be used to knockout multiple genes
simultaneously by co-expressing multiple sgRNAs targeting different sites (Cong et
al., 2013; Cao et al., 2016; Hashimoto et al., 2016). However, those approaches
achieved complete bi-allelic knockout animals with low efficiencies (Zuo et al., 2017).
Since there were six members in CYP2D subfamily, the deletion of those genes
simultaneously in rat genome would require an approach having very high gene
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
19
targeting efficiency. In 2017, Zuo et al. demonstrated a new “C-CRISPR” approach,
with which three genes (Tet1, Tet2, and Tet3) were completely knocked out in mouse
embryos after an one-step injection of sgRNAs in zygotes (Zuo et al., 2017). Most
recently, Campa et al. demonstrated a multiplexed genome targeting method using
Cas12a and CRISPR arrays delivered on a single plasmid (Campa et al., 2019). Those
high efficiency methods would greatly facilitate our future generation of a
CYP2D-null rat model with deletion of all the six CYP2D genes.
Depression is a serious mental disorder characterized by anhedonia, weight loss,
sleeping disturbances, a sense of worthlessness, cognitive decline, or repeated
thoughts of death (Xu et al., 2002). In the CMS rat model, 1% sucrose water
consumption reflected the euphoria of the rats, and the locomotor activity in open-box
was used to detect the state of nervousness in the new environment (Matthews et al.,
1995; Willner, 1997). Thus, body weight test, 1% sucrose water consumption test and
locomotor activity test were commonly used behavioral indicators for depression
research (Bortolato et al., 2007; Dai et al., 2010). After 20 days of depression
modeling, the body weight, 1% sucrose water consumption, and locomotor activity of
the CMS groups were changed similar to the previous reports (Pucilowski et al., 1993;
Orsetti et al., 2008; Zhao et al., 2008; Dai et al., 2010). Those results demonstrated
the reproducibility in our depression modeling. Notably, sporadic death occurred in
both of WT female and CYP2D1-null female rats during the CMS modeling (data not
shown). Whereas, the rat mortality rate our experiment was less than that of mice
under the same challenge (Goshen et al., 2008; Sun et al., 2011), indicating that the
rats are more tolerant than mice in the depression modeling.
For the depressed WT rats, after VLF-treatment for 14 days, all the three
depression indicators were significantly improved than those in saline-treated ones.
This result suggested that VLF was effective in our depression model. Based on the
plasma levels of NE and 5-TH, it was clear that VLF treatment was more effective in
WT depression rats than that in the CYP2D1-null ones, possibly because of the
reduced formation of ODV in CYP2D1-null rats. We also tried to determine ODV
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
20
concentrations in the brain, but was unable obtain convincing results due to the
limitation of the analytical instruments.
In conclusion, we have successfully developed a novel and valuable CYP2D1-null
rat model in studying human drug metabolism by CYP2D6 using the CRISPR/Cas9
technology and demonstrated that the antidepressant effects of VLF were decreased in
the depressed CYP2D1-null rats. Moreover, our data demonstrated that the reduced
ODV formation was associated with a lower VLF efficacy in rats, suggesting a
potential application of monitoring plasma ODV level in the clinical optimization of
VLF for the treatment of depression.
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
21
Authorship contributions
Participated in research design: Yuan Wei, Jun Gu, Hongqiu Zhou
Conducted experiments: Hongqiu Zhou, Yang Li, Changsuo Wang, Zhiqiang Li
Performed data analysis: Hongqiu Zhou, Mangting Shan
Wrote or contributed to the writing of the manuscript: Hongqiu Zhou, Yuan Wei, Jun
Gu, Zhen Ouyang
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
22
References
Anderson KR, Haeussler M, Watanabe C, Janakiraman V, Lund J, Modrusan Z,
Stinson J, Bei Q, Buechler A, Yu C, Thamminana SR, Tam L, Sowick M-A,
Alcantar T, O’Neil N, Li J, Ta L, Lima L, Roose-Girma M, Rairdan X,
Durinck S, and Warming S (2018) CRISPR off-target analysis in genetically
engineered rats and mice. Nat Methods 15:512–514.
Bissig K-D, Han W, Barzi M, Kovalchuk N, Ding L, Fan X, Pankowicz FP, Zhang
Q-Y, and Ding X (2018) P450-Humanized and Human Liver Chimeric Mouse
Models for Studying Xenobiotic Metabolism and Toxicity. Drug Metab Dispos
46:1734–1744.
Bortolato M, Mangieri RA, Fu J, Kim JH, Arguello O, Duranti A, Tontini A, Mor M,
Tarzia G, and Piomelli D (2007) Antidepressant-like Activity of the Fatty Acid
Amide Hydrolase Inhibitor URB597 in a Rat Model of Chronic Mild Stress.
Biol Psychiatry 62:1103–1110.
Bradford LD (2002) CYP2D6 allele frequency in European Caucasians, Asians,
Africans and their descendants. Pharmacogenomics 3:229–243.
Bymaster F (2001) Comparative Affinity of Duloxetine and Venlafaxine for Serotonin
and Norepinephrine Transporters in vitro and in vivo, Human Serotonin
Receptor Subtypes, and Other Neuronal Receptors.
Neuropsychopharmacology 25:871–880.
Campa CC, Weisbach NR, Santinha AJ, Incarnato D, and Platt RJ (2019) Multiplexed
genome engineering by Cas12a and CRISPR arrays encoded on single
transcripts. Nat Methods, doi: 10.1038/s41592-019-0508-6.
Cao J, Wu L, Zhang S-M, Lu M, Cheung WKC, Cai W, Gale M, Xu Q, and Yan Q
(2016) An easy and efficient inducible CRISPR/Cas9 platform with improved
specificity for multiple gene targeting. Nucleic Acids Res gkw660.
Cong L, Ran FA, Cox D, Lin S, Barretto R, Habib N, Hsu PD, Wu X, Jiang W,
Marraffini LA, and Zhang F (2013) Multiplex Genome Engineering Using
CRISPR/Cas Systems. Science 339:819–823.
Dai Y, Li Z, Xue L, Dou C, Zhou Y, Zhang L, and Qin X (2010) Metabolomics study
on the anti-depression effect of xiaoyaosan on rat model of chronic
unpredictable mild stress. J Ethnopharmacol 128:482–489.
De Leon J, Susce MT, Pan R-M, Fairchild M, Koch WH, and Wedlund PJ (2005) The
CYP2D6 Poor Metabolizer Phenotype May Be Associated With Risperidone
Adverse Drug Reactions and Discontinuation. J Clin Psychiatry 66:15–27.
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
23
Egorin MJ, Rosen DM, Benjamin SE, Callery PS, Sentz DL, and Eiseman JL (1997)
In vitro metabolism by mouse and human liver preparations of halomon, an
antitumor halogenated monoterpene. Cancer Chemother Pharmacol 41:9–14.
Gonzalez FJ (2003) Role of Gene Knockout and Transgenic Mice in the Study of
Xenobiotic Metabolism. Drug Metab Rev 35:319–335.
Goshen I, Kreisel T, Ben-Menachem-Zidon O, Licht T, Weidenfeld J, Ben-Hur T, and
Yirmiya R (2008) Brain interleukin-1 mediates chronic stress-induced
depression in mice via adrenocortical activation and hippocampal
neurogenesis suppression. Mol Psychiatry 13:717–728.
Gu J, Weng Y, Zhang Q-Y, Cui H, Behr M, Wu L, Yang W, Zhang L, and Ding X
(2003) Liver-specific deletion of the NADPH-cytochrome P450 reductase
gene: impact on plasma cholesterol homeostasis and the function and
regulation of microsomal cytochrome P450 and heme oxygenase. J Biol Chem
278:25895–25901.
Guengerich FP (2008) Cytochrome P450 and Chemical Toxicology. Chem Res Toxicol
21:70–83.
Haduch A, Wójcikowski J, and Daniel WA (2005) Effect of short- and long-term
treatment with antidepressant drugs on the activity of rat CYP2A in the liver.
Pharmacol Rep Pr 57:774.
Handa T, Katare RG, Kakinuma Y, Arikawa M, Ando M, Sasaguri S, Yamasaki F, and
Sato T (2009) Anti-Alzheimer’s Drug, Donepezil, Markedly Improves
Long-Term Survival After Chronic Heart Failure in Mice. J Card Fail
15:805–811.
Hashimoto M, Yamashita Y, and Takemoto T (2016) Electroporation of Cas9
protein/sgRNA into early pronuclear zygotes generates non-mosaic mutants in
the mouse. Dev Biol 418:1–9.
Hertz DL, Snavely AC, McLeod HL, Walko CM, Ibrahim JG, Anderson S, Weck KE,
Rubin P, Olajide O, Moore S, Raab R, Carrizosa DR, Corso S, Schwartz G,
Peppercorn JM, Evans JP, Desta Z, Flockhart DA, Carey LA, and Irvin WJ
(2015) CYP2D6 intermediate metabolizers includes patient groups with
distinct metabolic activity. Cancer Res 75:P1-03–02.
Kandasamy M, Srinivas P, Subramaniam K, Ravi S, John J, Shekar R, Srinivas N, and
Thangam S (2010) Differential outcomes from metabolic ratios in the
identification of CYP2D6 phenotypes–focus on venlafaxine and
O-desmethylvenlafaxine. Eur J Clin Pharmacol 66:879–887.
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
24
Kim H, Whang W-W, Kim H-T, Pyun K-H, Cho S-Y, Hahm D-H, Lee H-J, and Shim I
(2003) Expression of neuropeptide Y and cholecystokinin in the rat brain by
chronic mild stress. Brain Res 983:201–208.
Li D, Qiu Z, Shao Y, Chen Y, Guan Y, Liu Meizhen, Li Y, Gao N, Wang L, Lu X,
Zhao Y, and Liu Mingyao (2013) Heritable gene targeting in the mouse and rat
using a CRISPR-Cas system. Nat Biotechnol 31:681–683.
Li M, Zheng C, Sato T, Kawada T, Sugimachi M, and Sunagawa K (2004) Vagal
Nerve Stimulation Markedly Improves Long-Term Survival After Chronic
Heart Failure in Rats. Circulation 109:120–124.
Liu J, Li Y, Bai G, Wang J, and Xue C (2011) The change of behavior and the
consumption of food and syrup of depression rats model under space
circumstances. Chin J Vet Med 47:12–14.
Lu J, Shao Y, Qin X, Liu D, Chen A, Li D, Liu M, and Wang X (2017) CRISPR
knockout rat cytochrome P450 3A1/2 model for advancing drug metabolism
and pharmacokinetics research. Sci Rep 7:42922.
Magalhães P, Alves G, Llerena A, and Falcão A (2014) Venlafaxine pharmacokinetics
focused on drug metabolism and potential biomarkers. Drug Metabol Drug
Interact 29:129–141.
Martignoni M, Groothuis GMM, and De Kanter R (2006) Species differences
between mouse, rat, dog, monkey and human CYP-mediated drug metabolism,
inhibition and induction. Expert Opin Drug Metab Toxicol 2:875–894.
Matthews K, Forbes N, and Reid IC (1995) Sucrose consumption as an hedonic
measure following chronic unpredictable mild stress. Physiol Behav
57:241–248.
Meyer UA, Skoda RC, and Zanger UM (1990) The genetic polymorphism of
debrisoquine/sparteine metabolism—molecular mechanisms. Pharmacol Ther
46:297–308.
Nelson DR, Koymans L, Kamataki T, Stegeman JJ, Feyereisen R, Waxman DJ,
Waterman MR, Gotoh O, Coon MJ, Estabrook RW, Gunsalus IC, and Nebert
DW (1996) P450 superfamily: update on new sequences, gene mapping,
accession numbers and nomenclature. Pharmacogenetics 6:1–42.
Orsetti M, Di Brisco F, Canonico PL, Genazzani AA, and Ghi P (2008) Gene
regulation in the frontal cortex of rats exposed to the chronic mild stress
paradigm, an animal model of human depression. Eur J Neurosci
27:2156–2164.
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
25
Preskorn S, Patroneva A, Silman H, Jiang Q, Isler JA, Burczynski ME, Ahmed S, Paul
J, and Nichols AI (2009) Comparison of the Pharmacokinetics of Venlafaxine
Extended Release and Desvenlafaxine in Extensive and Poor Cytochrome
P450 2D6 Metabolizers. J Clin Psychopharmacol 29:39–43.
Pucilowski O, Overstreet DH, Rezvani AH, and Janowsky DS (1993) Chronic mild
stress-induced anhedonia: Greater effect in a genetic rat model of depression.
Physiol Behav 54:1215–1220.
Robinson HJ, Phares HF, and Graessle OE (1968) Effects of indomethacin on acute,
subacute, and latent infections in mice and rats. J Bacteriol 96:6–13.
Ryan DE, and Levin W (1990) Purification and characterization of hepatic
microsomal cytochrome P-450. Pharmacol Ther 45:153–239.
Scheer N, Kapelyukh Y, McEwan J, Beuger V, Stanley LA, Rode A, and Wolf CR
(2012) Modeling Human Cytochrome P450 2D6 Metabolism and Drug-Drug
Interaction by a Novel Panel of Knockout and Humanized Mouse Lines. Mol
Pharmacol 81:63–72.
Scheer N, McLaughlin LA, Rode A, MacLeod AK, Henderson CJ, and Wolf CR
(2014) Deletion of 30 Murine Cytochrome P450 Genes Results In Viable Mice
With Compromised Drug Metabolism. Drug Metab Dispos 42:1022–1030.
Su T, and Ding X (2004) Regulation of the cytochrome P450 2A genes. Toxicol Appl
Pharmacol 199:285–294.
Sun X-L, Liu Y, Dai T, Ding J-H, and Hu G (2011) Uncoupling protein 2 knockout
exacerbates depression-like behaviors in mice via enhancing inflammatory
response. Neuroscience 192:507–514.
Tan EP, Li Y, Del Castillo Velasco-Herrera M, Yusa K, and Bradley A (2015)
Off-target assessment of CRISPR-Cas9 guiding RNAs in human iPS and
mouse ES cells: Off-Target Assessment of CRISPR-Cas9. genesis
53:225–236.
Teh LK, and Bertilsson L (2012) Pharmacogenomics of CYP2D6: Molecular Genetics,
Interethnic Differences and Clinical Importance. Drug Metab Pharmacokinet
27:55–67.
Thangavel C, Garcia MC, and Shapiro BH (2004) Intrinsic sex differences determine
expression of growth hormone-regulated female cytochrome P450s. Mol Cell
Endocrinol 220:31–39.
Venhorst J, ter Laak AM, Commandeur JNM, Funae Y, Hiroi T, and Vermeulen NPE
(2003) Homology Modeling of Rat and Human Cytochrome P450 2D
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
26
(CYP2D) Isoforms and Computational Rationalization of Experimental
Ligand-Binding Specificities. J Med Chem 46:74–86.
Wallace BD, and Redinbo MR (2013) Xenobiotic-sensing nuclear receptors involved
in drug metabolism: a structural perspective. Drug Metab Rev 45:79–100.
Wang X, Tang Y, Lu J, Shao Y, Qin X, Li Y, Wang L, Li D, and Liu M (2016)
Characterization of novel cytochrome P450 2E1 knockout rat model generated
by CRISPR/Cas9. Biochem Pharmacol 105:80–90.
Waxman D, Ram P, Pampori N, and Shapiro B (1995) Growth hormone regulation of
male-specific rat liver P450s 2A2 and 3A2: induction by intermittent growth
hormone pulses in male but not female rats rendered growth hormone
deficient by neonatal monosodium glutamate. Mol Pharmacol 48:790.
Wei Y, Yang L, Zhang X, Sui D, Wang C, Wang K, Shan M, Guo D, and Wang H
(2018) Generation and Characterization of a CYP2C11-Null Rat Model by
Using the CRISPR/Cas9 Method. Drug Metab Dispos 46:525–531.
Wellington K, and Perry CM (2001) Venlafaxine Extended-Release: A Review of its
Use in the Management of Major Depression. CNS Drugs 15:643–669.
Willner P (1997) Validity, reliability and utility of the chronic mild stress model of
depression: a 10-year review and evaluation. Psychopharmacology (Berl)
134:319–329.
Wuttke H, Rau T, Heide R, Bergmann K, Bohm M, Weil J, Werner D, and
Eschenhagen T (2002) Increased frequency of cytochrome P450 2D6 poor
metabolizers among patients with metoprolol-associated adverse effects. Clin
Pharmacol Ther 72:429–437.
Xu S, Lu X, Ying Y, He X, Zhang J, and Ye C (2002) An controlled analysis of
clinical characters of 43 cases with chronic depression. Sichuan Ment Health
15:24–26.
Yamazaki H (ed) (2014) Fifty Years of Cytochrome P450 Research, Springer Japan,
Tokyo.
Ye H, Sui D, Liu W, Yuan Y, Ouyang Z, and Wei Y (2019) Effects of CYP2C11 gene
knockout on the pharmacokinetics and pharmacodynamics of warfarin in rats.
Xenobiotica, doi: 10.1080/00498254.2019.1579006.
Zanger UM, and Schwab M (2013) Cytochrome P450 enzymes in drug metabolism:
Regulation of gene expression, enzyme activities, and impact of genetic
variation. Pharmacol Ther 138:103–141.
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
27
Zanger UM, Turpeinen M, Klein K, and Schwab M (2008) Functional
pharmacogenetics/genomics of human cytochromes P450 involved in drug
biotransformation. Anal Bioanal Chem 392:1093–1108.
Zhao Z, Wang W, Guo H, and Zhou D (2008) Antidepressant-like effect of liquiritin
from Glycyrrhiza uralensis in chronic variable stress induced depression
model rats. Behav Brain Res 194:108–113.
Zhou X, Chan K, and Yeung JHK (2012) Herb-drug interactions with Danshen (Salvia
miltiorrhiza): a review on the role of cytochrome P450 enzymes. Drug
Metabol Drug Interact 27:9–18.
Zuo E, Cai Y-J, Li K, Wei Y, Wang B-A, Sun Y, Liu Z, Liu J, Hu X, Wei W, Huo X,
Shi L, Tang C, Liang D, Wang Y, Nie Y-H, Zhang C-C, Yao X, Wang X, Zhou
C, Ying W, Wang Q, Chen R-C, Shen Q, Xu G-L, Li J, Sun Q, Xiong Z-Q, and
Yang H (2017) One-step generation of complete gene knockout mice and
monkeys by CRISPR/Cas9-mediated gene editing with multiple sgRNAs. Cell
Res 27:933–945.
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
28
Footnotes
This study was supported by the National Natural Science Foundation of China [Grant
Nos. 81102522, 81373480, and 81573529], and the Natural Science Foundation of
Jiangsu Province [Grant No. BK2011473].
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
29
FIGURE LEGENDS
Figure 1 Confirmation of gene targeting in CYP2D1-null rats. (A) Schematic of the
exon structure of the CYP2D1 gene in rats. Exons are shown as black boxes. The 7 bp
(ACCTCAT) was deleted in exon 4. (B) Fragments of approximately 586 bp were
amplified from ten F2 pups. (n = 10) (C) Genotyping results for CYP2D1–/–
,
CYP2D1+/–
and wild-type (WT) rats; seven base pairs were deleted in exon 4 of
CYP2D1 in the knockout alleles.
Figure 2 Western blotting analyses of several cytochrome P450 isoforms (CYP2D1,
CYP1A2, CYP2B, CYP2A, CYP2C11, and CYP2E1) in the livers of CYP2D1-null
and WT rats. Each lane represents the pooled microsomes of the livers of WT or
CYP2D1-null rats. CYP2D1 was not detected in the liver microsomes of
CYP2D1-null rats. The expression of CYP1A2, CYP2B, CYP2C11, and CYP2E1 in
CYP2D1-null rats showed no marked difference, while the expression of CYP2A
showed higher levels, compared to that of WT rats. CYP2C11 protein was not
expressed in female rats. WT (M), wild-type male rats; WT (F), wild-type female rats;
Null (M), CYP2D1-null male rats; Null (F), CYP2D1-null female rats.
Figure 3 Mean plasma concentration-time curves of venlafaxine hydrochloride and
O-desmethyl venlafaxine after intragastrical administration at a dose of 20 mg/kg in
wild-type (WT) Sprague-Dawley and CYP2D1-null rats. (A) Mean plasma
concentration-time curves of venlafaxine hydrochloride (VLF) in male rats; (B) Mean
plasma concentration-time curves of venlafaxine hydrochloride (VLF) in female rats;
(C) Mean plasma concentration-time curves of O-desmethyl venlafaxine (ODV) in
male rats; (D) Mean plasma concentration-time curves of O-desmethyl venlafaxine
(ODV) in female rats. WT-M, wild-type male rats; WT-F, wild-type female rats;
Null-M, CYP2D1-null male rats; Null-F, CYP2D1-null female rats. All values are
shown as the mean ± SD, n = 6. *P < 0.05, compared to that of WTs with same sex.
Figure 4 The levels of NE (A) and 5-HT (B) in CYP2D1-null and WT rats after
administration of venlafaxine hydrochloride. WT-M, wild-type male rats; WT-F,
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
30
wild-type female rats; Null-M, CYP2D1-null male rats; Null-F, CYP2D1-null female
rats. Ctrl, control group; CMS Saline, chronic mild stimulations saline-treated group;
CMS VLF, the group of chronic mild stimulations treated-rats administered with
venlafaxine. All values are shown as the mean ± SD, n = 6. *P < 0.05, compared to
that of Ctrl group; #P < 0.05, compared to that of CMS Saline group.
$P < 0.05,
compared to that of WT group.
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
31
Table 1. Viscera indices in CYP2D1-null and wild-type rats.
All values are shown as the mean ± SD, n = 6. WT-M, wild-type male rats; WT-F,
wild-type female rats; Null-M, CYP2D1-null male rats; Null-F, CYP2D1-null female
rats.
Viscera indices Null-M WT-M Null-F WT-F
Kidney (%) 0.9321 ± 0.0838 0.8779 ± 0.0121 0.7357 ± 0.0203 0.9419 ± 0.0550
Liver (%) 4.537 ± 0.3352 4.700 ± 0.4461 3.915 ± 0.3302 4.108 ± 0.0110
Heart (%) 0.3659 ± 0.0255 0.3646 ± 0.0271 0.4032 ± 0.0041 0.4140 ± 0.0113
Testis (%) 0.9323 ± 0.0840 0.9521 ± 0.0372 N/A N/A
Spleen (%) 0.2208 ± 0.0290 0.2518 ± 0.0091 0.1880 ± 0.0061 0.2446 ± 0.0324
Brain (%) 0.6345 ± 0.0202 0.5511 ± 0.0460 0.7624 ± 0.0650 0.8546 ± 0.0452
Lungs (%) 0.6309 ± 0.0588 0.5602 ± 0.0711 0.6343 ± 0.0649 0.7862 ± 0.0951
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
32
Table 2. Kinetics of venlafaxine hydrochloride metabolism in liver microsomes
from CYP2D1-null and wild-type rats.
All values are shown as the mean ± SD, n = 3. Km, substrate concentration at half of
the maximum reaction rate; Vmax, maximum reaction rate; CLint, intrinsic clearance;
WT-M, wild-type male rats; WT-F, wild-type female rats; Null-M, CYP2D1-null
male rats; Null-F, CYP2D1-null female rats. *P < 0.05, compared to that of WTs with
same sex.
Vmax (nmol-1
·min-1
·mg potein-1
) Km (µmol·L-1
) CLint (L·mg-1
·min-1
)
WT-M 156.7 ± 14.7 94.7 ± 8.9 1655 ± 165
Null-M 84.6 ± 5.5* 117.9 ± 16.6
* 717 ± 33
*
WT-F 125.1 ± 18.8 144.4 ± 8.7 866 ± 216
Null-F 66.2 ± 9.1* 181.1 ± 15.9
* 365 ± 57
*
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
33
Table 3. Plasma pharmacokinetic parameters in CYP2D1-null rats and wild-type
rats after treatment with venlafaxine hydrochloride at 20 mg/kg.
Parameter WT-M Null-M WT-F Null-F
AUC0-t
(μg·mL-1
·h-1
)
12.4 ± 3.3 20.3 ± 2.5* 13.6 ± 1.7 19.4 ± 1.3*
AUC0-∞
(μg·mL-1
·h-1
)
15.0 ± 2.6 23.8 ± 4.2* 15.3 ± 2.1 20.7 ± 1.5*
Cmax
(μg·mL-1
)
1.9 ± 0.1 2.4 ± 0.1* 2.6 ± 0.1 3.0 ± 0.1*
Vd/F
(103
mL·kg-1
)
25.2 ± 7.2 13.1 ± 3.4* 15.4 ± 1.4 11.5 ± 2.5*
Cl/F
(103
mL·h-1
·kg-1
)
1.4 ± 0.3 0.9 ± 0.2* 1.3 ± 0.2 1.0 ± 0.1*
All values are shown as the mean ± SD, n = 6. Cmax, peak concentration; AUC, area
under the curve of the drug; AUC0-t, AUC from the time of administration to the last
point; AUC0-∞, time from the start of administration to the theoretical extrapolation
AUC; CL/F, apparent total clearance; Vd/F, apparent volume of distribution. WT-M,
wild-type male rats; WT-F, wild-type female rats; Null-M, CYP2D1-null male rats;
Null-F, CYP2D1-null female rats. *P < 0.05, compared to that of WTs.
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from
DMD # 88526
34
Table 4. The body weight, 1% sucrose water consumption, and locomotor
activity of CYP2D1-null and wild-type rats after depression modeling.
All values are shown as the mean ± SD, n = 5. Ctrl, control group without depression
modeling; CMS, chronic mild stimulations-treated group. WT-M, wild-type male rats;
WT-F, wild-type female rats; Null-M, CYP2D1-null male rats; Null-F, CYP2D1-null
female rats. *P < 0.01, compared to that of control groups.
WT-M Null-M WT-F Null-F
Body Weight (g)
Ctrl 217.9 ± 17.2 214.3 ± 12.0 210.5 ± 9.4 203.9 ± 12.4
CMS 178.5 ± 11.1* 198.2 ± 11.9
* 165.1 ± 8.5
* 158.5 ± 15.8
*
1% Sucrose water consumption (mL/100 g)
Ctrl 20.5 ± 1.0 22.2 ± 1.4 21.7 ± 2.1 19.3 ± 1.1
CMS 18.1 ± 0.8* 16.6 ± 2.4
* 12.3 ± 1.3
* 13.3 ± 1.1
*
Total distance (m)
Ctrl 22.1 ± 0.5 24.4 ± 1.1 21.2 ± 1.2 20.0 ± 1.2
CMS 18.4 ± 1.7*
21.5 ± 1.1* 17.4 ± 1.6
* 17.5 ± 1.5
*
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
35
Table 5. The body weight, 1% sucrose water consumption, and locomotor
activity of CYP2D1-null and wild-type rats after administration.
All values are shown as the mean ± SD, n = 5. WT-M, wild-type male rats; WT-F,
wild-type female rats; Null-M, CYP2D1-null male rats; Null-F, CYP2D1-null female
rats. Ctrl, control group without depression modeling; CMS Saline, chronic mild
stimulations-treated group administered with saline; CMS VLF, chronic mild
stimulations-treated group administered with venlafaxine. *P < 0.05, compared to that
of Ctrl group; #P < 0.05, compared to that of CMS Saline group.
$P < 0.05,
compared to that of WT group.
WT-M Null-M WT-F Null-F
Body Weight (g)
Ctrl 255.8 ± 9.0 240.1 ± 15.5 228.7 ± 14.6 220.7 ± 7.3
CMS Saline 207.6 ± 24.9* 185.6 ± 14.0
* 174.0 ± 8.1
* 134.3 ± 14.5
*
CMS VLF 237.0 ± 14.4*,#
197.6 ± 15.1*,$
194.5 ± 9.7*,#
152.5 ± 11.5*,$
1% Sucrose water consumption (mL/100 g)
Ctrl 22.2 ± 1.5 20.5 ± 0.9 19.2 ± 0.9 20.3 ± 1.5
CMS Saline 17.8 ± 1.7* 15.0 ± 1.2
* 13.0 ± 1.6
* 14.6 ± 1.7
*
CMS VLF 21.2 ± 1.6# 16.2 ± 1.3
*,$ 17.1 ± 1.0
*,# 13.5 ± 1.1
*,$
Total distance (m)
Ctrl 23.8 ± 1.2 21.5 ± 0.7 19.5 ± 1.2 20.7 ± 1.3
CMS Saline 20.4 ± 0.9* 18.0 ± 1.9
* 17.4 ± 0.7
* 15.6 ± 1.5
*
CMS VLF 24.6 ± 1.6#
20.2 ± 1.5$ 21.4 ± 1.5
# 16.5 ± 1.5
*,$
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
36
Figure 1
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
37
Figure 2
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
38
Figure 3
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from

DMD # 88526
39
Figure 4
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on October 28, 2019 as DOI: 10.1124/dmd.119.088526
at ASPE
T Journals on M
arch 1, 2020dm
d.aspetjournals.orgD
ownloaded from






















![Cardio - Admera Health · PHARMACOGENOMICS TEST TO BETTER TREAT CARDIOVASCULAR DISEASES & CONDITIONS Cardio [16] Genes ABCB1 ACE ADRA2A AGTR1 APOE CYP2C19 CYP2C9 CYP2D6 CYP3A4 CYP3A5](https://img.dokumen.tips/doc/110x75/5fb27aa653d65601df68e958/cardio-admera-health-pharmacogenomics-test-to-better-treat-cardiovascular-diseases.jpg)






