Embed Size (px)
Citation preview

CxGELSIX: A Novel Preparation of Type VI Collagenwith Possible Use as a Biomaterial
Manapon Lekskul, M.D., Robert Burrows, Ph.D., Claire L. Kublin, B.S.,and Charles Cintron, Ph.D.
Purpose. This study was initiated to evaluate tissue acceptanceand stability of a novel type VI collagen preparation (CxGelsix)as a biomaterial in the rabbit corneal stroma. We hypothesizedthat CxGelsix, embedded intrastromally, does not have anyadverse affect on surrounding corneal tissues, and remains in-tact in the presence of an acute inflammatory reaction duringcorneal wound healing. Methods. Type VI collagen was ex-tracted and purified from rabbit corneal stroma under nonde-naturing conditions. This preparation, Gelsix, was concentratedand cross-linked with polyethylene glycol to produce a trans-parent film (CxGelsix). Discs of CxGelsix, 4.0-mm diameter,9- to 35-�m thick were implanted intrastromally and clinicallyexamined periodically for 4 months. In another experiment,implantation of CxGelsix, 2.0-mm-diameter, was followed bycorneal wounding adjacent to the implant and examined clini-cally for 30 weeks. At the end of these periods, the tissues fromthese experiments were processed for light and transmissionelectron microscopy. Results. An intralamellar 4.0-mm-diameter disc of CxGelsix does not alter the structure of cornealepithelium above the implant, suggesting normal transport ofnutrients through CxGelsix. Moreover, no structural abnormali-ties were seen in the rest of the cornea, and the cornea remainstransparent. Although the cornea accepts the presence ofCxGelsix disc as judged by clinical criteria, gradual degrada-tion of the implant is seen ultrastructurally. CxGelsix is re-markably stable despite its exposure to endogenous enzymesduring inflammation and wound healing. Partial degradation ofthe implant occurs only after many months, and it is graduallyreplaced with bundles of fine collagen fibrils reminiscent ofnormal cornea. Conclusion. The results of this study suggestthat CxGelsix is potentially useful as a biomaterial. KeyWords: Type VI collagen—�ig—Cornea—Stroma—Biomaterial—Gelsix—CxGelsix.
The filamentous type VI collagen, a ubiquitous struc-tural protein, is an integral component of the interfibrillarmatrix (1,2), and a measurable fraction of connectivetissue collagens. This collagen is a heterotrimer com-
posed of three genetically distinct polypeptides. Eachpolypeptide contains globular domains at the amino andcarboxyl termini separated by a short triple-helical do-main. The dumbell-shaped monomers assemble into te-tramers by lateral association. End-to-end association ofthese tetramers forms a beaded filamentous structure (3).
The 100-nm periodic beaded filament of type VI col-lagen is involved in cell–matrix interactions, and be-lieved to interact with extracellular matrix componentsincluding collagens, hyaluronan, and proteoglycans(4–16). As a structural protein, this collagen may play arole in anchoring basal-lamina–containing organs withinconnective tissues and restrict lateral movement of col-lagen fibrils. By virtue of its distribution in cornealstroma (9), its synthesis during morphogenesis (17), andits interaction with both corneal stromal cells and extra-cellular matrix proteins, type VI collagen may also playan important role in assembling and maintaining propercollagen fibril spacing so necessary for corneal transpar-ency.
Native type VI collagen copurifies with a protein iden-tified as �ig-h3 (18), a protein induced in a human ad-enocarcinoma cell line after treatment with TGF-�. Thisprotein in rabbit, named �ig, is synthesized in cornealstroma during morphogenesis of normal and healing tis-sue, indicating that it plays a role in these processes (18).The 683-amino acids sequence of �ig has some similar-ity with fasciculin I, a possible surface-recognition mol-ecule involved in nerve growth cone guidance, and OSF-2, a protein that has been suggested to function as anadhesion molecule in bone formation (19–21).
Recent observations suggest �ig-h3 may be an integralcomponent of extracellular matrix structures (22–24).In corneal stroma, �ig is associated with the globulardomain of native type VI collagen requiring disulfide-dependent linkages (unpublished data). Similar prepara-tions of type VI collagen, containing �ig, promote adhe-sion and spreading of corneal fibroblasts and smooth-muscle cells in vitro (11,25,27). Adhesion of cells to type
Submitted December 3, 1998. Revision received April 29, 1999.Accepted May 3, 1999.
From the Schepens Eye Research Institute, and Department of Oph-thalmology, Harvard Medical School, Boston, Massachusetts, U.S.A.
Address correspondence and reprint requests to Dr. C. Cintron, P.O.Box 210, East Randolph, VT 05041, U.S.A.
Proprietary interest category: None.
Cornea 19(2): 194–203, 2000. © 2000 Lippincott Williams & Wilkins, Inc., Philadelphia
194

VI collagen and subsequent cell spreading may be par-tially mediated by �ig (26,27). As an integral componentof the stromal interfibrillar matrix during morphogenesis,and its possible function in cell-matrix interactions, �igmay play a role in development of an ordered fibrillarmatrix, necessary for corneal transparency.
We have shown that rabbit corneal type VI collagencan be extracted efficiently with phosphate-buffered sa-line and purified under nondenaturing conditions (un-published observations; Burrows, El-Shabrawi, Kublin,Cintron, 1998). The purified preparation, containing typeVI collagen associated with �ig, forms a viscous sub-stance when concentrated. Unpublished studies in ourlaboratory have shown that this preparation from corneais in the form of a gel (Gelsix). On chemical cross-linking with activated polyethylene glycol, the gel be-comes a transparent film (CxGelsix) that is mechanicallystrong, suggesting that it may serve as a biomaterial incornea.
Numerous studies and patents have described the useof collagens alone or in combination with other compo-nents as a biomaterial (28–39). In all of these studies,fibrillar collagens have been used exclusively. To ourknowledge no studies have examined the potential oftype VI collagen as a biomaterial.
This study was initiated to evaluate tissue acceptanceand stability of the type VI collagen preparation in thecorneal stroma. This study was not designed to testCxGelsix as an intralamellar lens. We hypothesized thatCxGelsix is accepted by corneal tissues and remains in-tact in the presence of an acute inflammatory reactionduring corneal wound healing.
MATERIAL AND METHODS
Type VI collagen from the rabbit cornea was ex-tracted, purified, and prepared as a gel (Gelsix) by anondenaturing method. Details of the extraction and pu-rification of type VI collagen are described elsewhere(unpublished observations, Burrows, El-Shabrawi, Kub-lin, Cintron, 1998). The purity of the type VI collagenwas assessed by sodium dodecyl sulfate–polyacrylamidegel electrophoresis under reducing and nonreducing con-ditions.
In brief, corneal stromas obtained from rabbit corneas(PelFreez, Rogers, AK, U.S.A.) were frozen and pulver-ized in a Wiley mill. Powdered corneal stroma was ex-tracted several times with phosphate-buffered saline(PBS), pH 7.0 containing a protease inhibitor cocktail.The extracts were pooled, filtered, and concentrated onan Amicon Diafiltration apparatus with a YM10 filter(Amicon Inc., Beverly, MA, U.S.A.). The concentratewas centrifuged, and the pellet discarded. The supernatewas then put through a CL2B (Sigma, St. Louis, MO,U.S.A.) column, and eluted with PBS containing 0.02%
sodium azide. The eluate was monitored spectrophoto-metrically at 280 nm, and type VI collagen eluted nearthe void volume followed by a broad peak of lower-molecular-weight material. Fractions containing type VIcollagen were pooled, concentrated, and recycledthrough the gel-filtration column two additional times.The purity of the type VI collagen was assessed by so-dium dodecyl sulfate–polyacrylamide gel electrophoresisunder reducing and nonreducing conditions.
Approximately 0.15 mg of purified type VI collagen/�ig was placed in a Centricon concentrator (Amicon)and centrifuged in a swinging bucket at 5,000 g for ∼1 h,depending on the initial volume of the sample, until aminiscus was formed on the filter. The gel (Gelsix) wascross-linked on the surface of the filter with 0.1-ml ofactivated disuccinimidyl glutarate polyethyleneglycol(di-SG-PEG; MW 3,400; Shearwater Polymers, Inc.Huntsville, AL, U.S.A.), 50 mg/ml H2O, for 15 minutesbefore centrifugation for 30 minutes. The cross-linkedGelsix (CxGelsix) was then washed with 0.2 ml PBS 3times and centrifuged after each wash. To view the im-plant easily, CxGelsix was stained with 0.2 ml of 5-([4,6-dichlorotrazin-2-YL]amino)-fluorescein hydrochloride(Sigma Chemical Co.; DTAF), 5 mg/ml in 0.2 M sodiumbicarbonate, a nontoxic fluorescent dye that covalentlybinds to collagens. CxGelsix was stained for 15 minutes,excess dye removed, and the gel washed with PBS asearlier. The Centricon tube was disassembled, and thegel floated off the filter in a bath of PBS.
Discs of CxGelsix, 2.0 and 4.0 mm in diameter, werecut out from the gel with a trephine. Stained CxGelsix,sterilized with sodium azide during isolation and purifi-cation of type VI collagen, was extensively washed insterile PBS, pH 7.2, and kept in 3 ml of sterile PBScontaining 6 drops of 0.3% ciprofloxacin HCl eyedrop(Alcon Laboratories, Inc., Fort Worth, TX, U.S.A.) for�1 hour, followed by UV exposure for 15 minutes in atissue-culture hood to ensure sterility before implantationin the corneas. The CxGelsix discs implanted in corneasranged in thickness of from 9- to 35-�m, as determinedby electron microscopy. In a separate preparation, thewater content of the gel was determined to be ∼87% byweight.
Adult rabbits were prepared for this study by usingprocedures in accordance with the Association for Re-search in Vision and Ophthalmology Statement for theUse of Animals in Ophthalmic and Vision research. Todetermine the response of corneal tissues to the presenceof CxGelsix intrastromally, 10 adult rabbits were anes-thetized with intramuscular injection of a “cocktail” ofketamine, xylazine, and acepromazine, 0.5–0.7 mL/kgbody weight, and received topical anesthesia with 0.5%proparacaine hydrochloride ophthalmic solution (SightPharmaceuticals, Inc, Tampa, FL, U.S.A.) on their cor-neas. The eyes were proptosed for immobilization, and
CxGELSIX: BIOMATERIAL 195
Cornea, Vol. 19, No. 2, 2000

the ocular surface was flushed with a topical antibioticeyedrop (0.3% ciprofloxacin HCl) before surgery. An8.0-mm-diameter corneal circle in the left eye was de-lineated with a trephine. A groove, deepened to ∼0.2 mm,was made in a small area of the temporal quadrant toform a corneal lip. This tissue was then slightly elevatedwith Colibri forceps, and an interlamellar dissection wasstarted with a Martinez double-ended corneal dissectorknife (Storz Instrument Co., St. Louis, MO, U.S.A.). Theplane was enlarged laterally toward the corneal apex byoscillating horizontal movements. After the temporal twothirds corneal flap was made, a 4.0-mm-diameter CxGel-six disc was carefully placed on the stromal bed andcentered over the pupil. The flap was replaced over theimplant without sutures. Surgical placement of all Cx-Gelsix implants was performed by the same surgeon byusing a surgical microscope (Moller Microsurgical, Can-ton, OH, U.S.A.). The right eye of each rabbit served asa sham control to evaluate the effects of freehand lamel-lar dissection.
The rabbits were divided into two groups. Group A,consisting of five rabbits, were not exposed to steroidbefore or after CxGelsix implantation. In group B, alsoconsisting of five rabbits, the CxGelsix disc was washedwith 0.1% fluorometholone acetate (FML; AllerganMedical Optic, Irvine, CA, U.S.A.) before implantation,and both eyes were treated with this topical steroid eye-drop during the postoperative period, once a day for 7days. In groups A and B, topical antibiotic (0.3% cipro-floxacin HCL) was instilled immediately on all corneasafter the procedure and in the early postoperative period3 times a day for 7 days. Three days postoperatively,neomycin-polymyxin B sulfates–bacitracin zinc eyeointment (Akorn, Abita Springs, LA, U.S.A.) was giventopically once a day for 7 days.
Clinical postoperative examinations were performedon days 3, 7, 14, and 21 after surgery, and monthly for 4months. The rabbits were then killed with an intravenousoverdose of pentobarbital, and the corneas were removedto determine the status of the implants and the cornealcells by light and transmission electron microscopy. Thesamples were fixed in 2% paraformaldehyde and 2.5%glutaraldehyde in 0.1 M cacodylate buffer, pH 7.4, de-hydrated in a graded ethanol series, and embedded inPoly/Bed 812 (Polysciences, Warrington, PA, U.S.A.) toobtain ultrathin sections on grids. Grids were stained in4% uranyl acetate in 50% ethanol for 25 minutes, fol-lowed by Reynolds’ lead citrate for 5 minutes, and ex-amined with a Philips 410 transmission electron micro-scope.
To determine the stability of CxGelsix in the presenceof a healing corneal wound, six adult rabbits were anes-thetized and prepared for surgery as described earlier. Ineach corneal stroma of both eyes, a pocket extending 2–3
mm from the limbus and ending near the center of thecornea was made with a Troutman knife (Weck/Xomed-Treace, Jacksonville, FL, U.S.A.). A 2.0-mm-diameterbutton of CxGelsix was introduced into each pocket andallowed to heal in place near the center of the cornea.The CxGelsix button was often folded on itself duringinsertion of the gel. After 1 week of healing, each rabbitwas prepared for surgery as before, and a 2.0-mm-diameter full-thickness trephination wound was made ineach eye adjacent to the CxGelsix implant (40). Each eyereceived antibiotics each day for 3 days as describedearlier and watched carefully to make sure no infectionsdeveloped. The eyes were examined each week by slit-lamp microscopy and photographed. At 11 and 30 weeksafter wounding, rabbits were killed with an overdose ofsodium pentobarbital, the eyes enucleated, and immedi-ately fixed and processed as earlier in preparation fortransmission electron microscopy.
RESULTS
Corneal Response to CxGelsix:Clinical Observations
Except for three eyes, the corneal epithelia of bothgroups were clinically normal, based on negative stain-ing with rose bengal. The experimental eye of one rabbitfrom group A had a minor corneal erosion at day 50, butrecovered again at day 60. In group B, the steroid-treatedanimals, the experimental eyes of two rabbits had mildsuperficial keratitis only during the first week after sur-gery. Minimal subepithelial infiltrate in these eyes waspresent anterior to the implant area, in an otherwise clearcornea. Fifty-five percent of all eyes showed corneal-flapretraction, six eyes in group A (no steroid treatment) andfive eyes in group B (steroid treatment). Of these, sixeyes had implants versus four sham controls. Three eyesin group A, one with the implant and two in the shamcontrols, developed mild superficial corneal neovascu-larization that failed to extend beneath the flaps. Two, ingroup A with implants, developed corneal flap thinning,in the midperiphery area near the original incision, atdays 45 and 50. A summary of the complications for bothgroups indicates that the steroid had some amelioratingeffects on several complications (Table 1). Althoughthere are no significant differences noted, there appearsto be a trend. Throughout the follow-up time, the implantremained transparent and survived intact with no sign ofinfection in any of the eyes. Taken together, the presenceof the CxGelsix implant failed markedly to alter thestructural integrity of the cornea, based on clinical ob-servations. The DTAF-stained CxGelsix implant re-mained clear and intact during the 4-month period of
M. LEKSKUL ET AL.196
Cornea, Vol. 19, No. 2, 2000

study (Fig. 1). No obvious changes in corneal thicknesswere evident by slit-lamp microscopy.
Corneal Response to CxGelsix: Light Microscopy
Although the histologic appearance varied from eye toeye, the general pattern of results is represented by thefigures shown. The structure of the cornea 4 months afterthe sham surgical procedure without postsurgical steroidtreatment showed minimal changes (Fig. 2A). The epi-thelium was composed of four to five layers of cells witha distinct line representing the basement membrane. Insome cases the location of the original lamellar dissec-tion was indicated by a slight difference in the stainingintensity of the stroma. The stroma and Descemet’smembrane posterior to the original incision appearednormal. The contralateral eye, containing the CxGelsixcorneal implant for 4 months, showed a homogeneouslystained structure located parallel to the surface about onethird to one half the depth of the stroma from the surface
of the cornea (Fig. 2B). In some cases, the implant con-tributed to the overall thickness of the cornea. This, how-ever, did not have any effect on the appearance or thick-ness of the epithelium, stroma, or Descemet’s membranein comparison to the sham control. In all cases examined,the CxGelsix implants were associated with numerouscells having rounded nuclei and dark-stained cytoplas-mic granules. The number of cells associated with im-plants varied considerably along their length. Eyestreated with steroid before the surgical procedure werehistologically indistinguishable from those not treatedwith steroids (not shown). As with the nonsteroidalsamples, a variable number of cells containing dark-stained granules were seen adjacent to the implant.Again, the epithelium, the stroma above and below theimplant, and Descemet’s membrane appeared normal.Moreover, the epithelium and stroma failed to show anyabnormalities associated with the presence of the implantother than the presence of large granule-containing cellsadjacent to the implant.
FIG. 1. Rabbit cornea with 4.0-mm-diameter CxGelsix implant. (A) The implant (arrow) hasbeen stained with DTAF to visualize the material. The transparency of CxGelsix is evidentby the appearance of the pupil and iris behind the implant. (B) Slit-lamp photograph ofcornea showing that the peripheral corneal thickness is similar to the central cornea thick-ness containing CxGelsix (arrow).
TABLE 1. Summary of corneal response to CxGelsix:Clinical observations
Experimental eye Sham eye
Group A Group B Group A Group B
Subepithelial infiltrate 3 — — —Corneal neovascularization 1 — 2 —Corneal erosion 1 — — —Corneal flap retraction 4 2 2 2Corneal flap thinning 2 — — —Corneal flap edema 1 — — 1Anterior chamber inflammation 1 — 1 —Superficial punctate keratopathy 2 — — —Implant thinning 1 — — —
Experimental eye contained CxGelsix implant.Sham eye, freehand lamellar dissection without CxGelsix implant; Group A, no
steroid treatment; Group B, steroid treatment.
CxGELSIX: BIOMATERIAL 197
Cornea, Vol. 19, No. 2, 2000

Corneal Response to CxGelsix: TransmissionElectron microscopy
Corneal ultrastructure at 4 months confirmed clinicaland histologic observations, showing that the epitheliumabove the implant was normal in comparison to the shamcontrol (Fig. 3). The epithelial structure in both tissuesconsisted of basal, wing, and squamous cell layers, withno indication of epithelial edema. Furthermore, an intactand uniform basement membrane failed to indicate any
previous damage. The stroma above and below the Cx-Gelsix implant contained bundles of collagen fibrils andoccasional stromal cells, with minimal cell organellesindicative of normal quiescent cells (Fig. 4). The char-acteristic short intertwining lamellae of the subepithelialstroma, and the long, uniform bundles of collagen fibrilsof the posterior stroma in corneas with CxGelsix im-plants gave no indication of extracellular matrix alter-ations. No stromal differences were seen between ste-roid- and non–steroid-treated corneas (not shown). Cx-Gelsix implants appeared ultrastructurally as densehomogeneous structures having a uniform thickness foreach sample (Fig. 5). The thickness of the implant, how-ever, varied from one preparation to another, rangingfrom 9 to 35 mm. In some instances, small electron-lucent pores were seen within the implant. Along theupper and lower surfaces of the implant, the structure ofthe matrix and cellular profile varied. Some regions
FIG. 2. Histologic section of central cornea of rabbit 4months after implantation of a 4.0-mm-diameter disc ofCxGelsix. (A) Cornea after lamellar dissection serves as asham control. (B) Stromal implantation of CxGelsix showsimplant (arrow) located about one third the depth of thecornea. The implant is associated with numerous darkcells (arrowhead). Other than an increase in corneal thick-ness, due to the implant thickness, no histologic differ-ences could be seen between the cornea with the implantand its sham control. Bar = 50 mm.
FIG. 3. Electron microscopy of corneal epithelium 4months after implantation of a 4.0-mm-diameter disc ofCxGelsix. (A) Corneal epithelium after lamellar dissectionserves as a sham control. The epithelium sits on a normalsubepithelial stroma. (B) Epithelium from cornea contain-ing CxGelsix implant. No marked differences can be seenbetween the cornea with implant and its sham control. Bar= 10 mm.
FIG. 4. Electron microscopy of corneal stroma 4 monthsafter implantation of a 4.0-mm-diameter disc of CxGelsix.(A) Subepithelial stroma (S) showing small tightly inter-woven lamellae characteristic of normal rabbit cornea. E,epithelium. (B) Stroma between the CxGelsix implant andDescemet’s membrane showing normal long uniform la-mellae. Bar = 1.0 mm.
FIG. 5. Electron microscopy of 4.0-mm-diameter disc ofCxGelsix 4 months after implantation. The implant (I), ∼20mm thick, is homogeneous except for small holes (arrow)dispersed throughout the implant. Corneal stromal lamel-lae (S) are arranged parallel to the implant. Bar = 10 mm.
M. LEKSKUL ET AL.198
Cornea, Vol. 19, No. 2, 2000

showed no discernible alteration of the implant structureor adjacent stromal cells. Fibroblastic cells near the im-plant were associated with a lamellar matrix adjacent tothe implant and distinct from the normal stromal matrix,suggestive of encapsulation (Fig. 6A). In other regions,irregularities in the implant surface and pockets contain-ing cellular material within the implant could be seenassociated with the presence of macrophages or fibro-blasts undergoing cell death (Fig. 6B). In some cases,macrophages were numerous, and these cells were asso-ciated with marked structural alterations in the implant(Fig. 7).
Stability of CxGelsix Implant in the Presence ofHealing Wound: Clinical Observations
Formation of a stromal pocket to accommodate a 2.0-mm disc of CxGelsix was intentionally crudely done tostimulate inflammation. Edema, associated with the for-mation of the stromal pocket, was noted a day after sur-gery. Nevertheless, all eyes, except for one, accepted theimplant without vascularization. The animal with a vas-
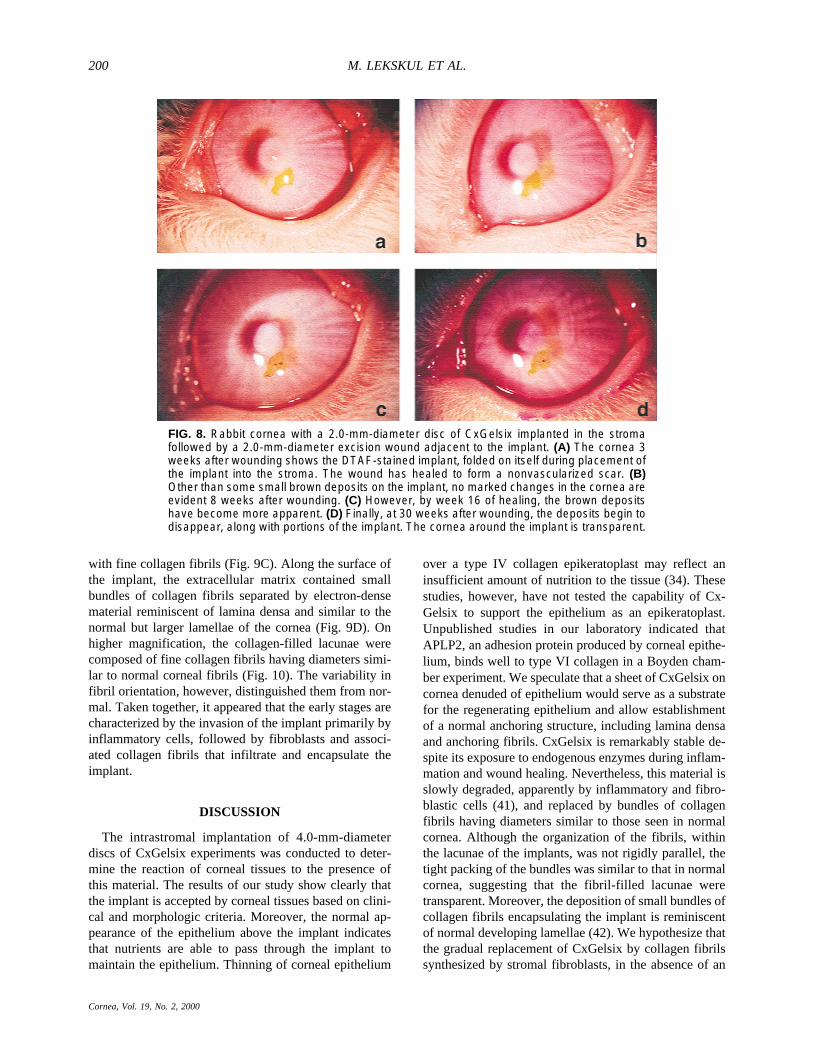
cularized cornea was eliminated from the study. A 2.0-mm-diameter excision wound was then made adjacent tothe CxGelsix 1 week after implantation and allowed toheal (Fig. 8). After 1 week of healing, all eyes wereclinically progressing well. A white, avascular scar tissuewas deposited along the edge of the wound, leaving thecenter occupied by a fibrin clot. The wound was subse-quently filled completely by avascular scar tissue by thesecond to third week. One week after the excision woundwas made, two eyes showed neovascularization at thelimbus, extending toward the implant. By the third weekafter implantation (2 weeks after wounding), the vascu-larization subsided in the two eyes and subsequently re-gressed. No apparent changes could be seen in the im-plants during the first 3 weeks after wounding (Fig. 8A).On week 8, small brown deposits were seen along theedge of the implants (Fig. 8B). These deposits progres-sively became larger by week 16 (Fig. 8C). In some eyes,CxGelsix-associated brown deposits were noticeably de-creased by week 30 (Fig. 8D).
Stability of CxGelsix Implant in the Presence ofHealing Wound: Transmission Electron Microscopy
Corneal epithelium and subjacent stroma appearednormal over the CxGelsix implant 11 weeks afterwounding. Deeper in the cornea, however, numerouscells were congregated around the implant (Fig. 9A).Based on their morphologic appearance, most of the cellswere identified as macrophages and fibroblasts. A finegranular–filamentous electron-dense material, presum-ably fibrin, was interspersed between the stroma and cel-lular infiltrate. By 30 weeks, most of the cells associatedwith the implant were fibroblastic with few macrophages(Fig. 9B). The fibroblasts were characterized by theabundance of rough endoplasmic reticulum and elon-gated nuclei. Spaces within the implant (lacunae), pre-sumably produced by enzymatic digestion, were filled
FIG. 6. Electron microscopy of 4.0-mm-diameter disc ofCxGelsix 4 months after implantation. (A) In some re-gions, the stromal–implant interface shows very little cel-lular activity except for an occasional keratocyte with anabundance of rough endoplasmic reticulum (arrow). (B) Inother regions of the cornea, numerous cells, probablymacrophages, are associated with irregular surfaces ofthe implant. Cell processes appear to be extending intothe implant to form lacunae within the implant (arrows). S,stroma; I, implant. Bar = 5.0 mm.
FIG. 7. Electron microscopy of 4.0-mm-diameter disc ofCxGelsix 4 months after implantation. In some cases, theinfiltration of cells into the implant (I) is extensive, sug-gesting that these macrophages (arrows) are digestingthe implant. Bar = 5.0 mm.
CxGELSIX: BIOMATERIAL 199
Cornea, Vol. 19, No. 2, 2000
with fine collagen fibrils (Fig. 9C). Along the surface ofthe implant, the extracellular matrix contained smallbundles of collagen fibrils separated by electron-densematerial reminiscent of lamina densa and similar to thenormal but larger lamellae of the cornea (Fig. 9D). Onhigher magnification, the collagen-filled lacunae werecomposed of fine collagen fibrils having diameters simi-lar to normal corneal fibrils (Fig. 10). The variability infibril orientation, however, distinguished them from nor-mal. Taken together, it appeared that the early stages arecharacterized by the invasion of the implant primarily byinflammatory cells, followed by fibroblasts and associ-ated collagen fibrils that infiltrate and encapsulate theimplant.
DISCUSSION
The intrastromal implantation of 4.0-mm-diameterdiscs of CxGelsix experiments was conducted to deter-mine the reaction of corneal tissues to the presence ofthis material. The results of our study show clearly thatthe implant is accepted by corneal tissues based on clini-cal and morphologic criteria. Moreover, the normal ap-pearance of the epithelium above the implant indicatesthat nutrients are able to pass through the implant tomaintain the epithelium. Thinning of corneal epithelium
over a type IV collagen epikeratoplast may reflect aninsufficient amount of nutrition to the tissue (34). Thesestudies, however, have not tested the capability of Cx-Gelsix to support the epithelium as an epikeratoplast.Unpublished studies in our laboratory indicated thatAPLP2, an adhesion protein produced by corneal epithe-lium, binds well to type VI collagen in a Boyden cham-ber experiment. We speculate that a sheet of CxGelsix oncornea denuded of epithelium would serve as a substratefor the regenerating epithelium and allow establishmentof a normal anchoring structure, including lamina densaand anchoring fibrils. CxGelsix is remarkably stable de-spite its exposure to endogenous enzymes during inflam-mation and wound healing. Nevertheless, this material isslowly degraded, apparently by inflammatory and fibro-blastic cells (41), and replaced by bundles of collagenfibrils having diameters similar to those seen in normalcornea. Although the organization of the fibrils, withinthe lacunae of the implants, was not rigidly parallel, thetight packing of the bundles was similar to that in normalcornea, suggesting that the fibril-filled lacunae weretransparent. Moreover, the deposition of small bundles ofcollagen fibrils encapsulating the implant is reminiscentof normal developing lamellae (42). We hypothesize thatthe gradual replacement of CxGelsix by collagen fibrilssynthesized by stromal fibroblasts, in the absence of an
FIG. 8. Rabbit cornea with a 2.0-mm-diameter disc of CxGelsix implanted in the stromafollowed by a 2.0-mm-diameter excision wound adjacent to the implant. (A) The cornea 3weeks after wounding shows the DTAF-stained implant, folded on itself during placement ofthe implant into the stroma. The wound has healed to form a nonvascularized scar. (B)Other than some small brown deposits on the implant, no marked changes in the cornea areevident 8 weeks after wounding. (C) However, by week 16 of healing, the brown depositshave become more apparent. (D) Finally, at 30 weeks after wounding, the deposits begin todisappear, along with portions of the implant. The cornea around the implant is transparent.
M. LEKSKUL ET AL.200
Cornea, Vol. 19, No. 2, 2000

acute inflammatory environment, results in the formationof a transparent tissue similar to normal stroma. In ourmodel, the trephination wound heals rapidly with thedeposition of an opaque scar tissue by 2 to 3 weeks (40).The deposition of matrix around and within the CxGelsixoccurs late, perhaps after 11 weeks, well after the acuteinflammatory reaction in the trephination wound hassubsided. At this time the corneal epithelium and sur-rounding stroma has returned to its normal structure, ef-fectively sealing the implant from further interactionwith tear-borne inflammatory factors and polymorpho-nuclear leukocytes, present only during early periods ofhealing. In some ways, the de novo deposition of im-plant-associated matrix is reminiscent of fetal healing[i.e., it occurs in the absence of polymorphonuclear leu-kocytes (43)]. As in fetal wound healing, the new matrixis deposited without the formation of scar tissue.
The invasion of CxGelsix by stromal fibroblasts dur-ing wound healing is not surprising because type VI
FIG. 9. Electron microscopy of rabbit cornea with a 2.0-mm-diameter disc of CxGelsiximplanted in the stroma followed by a 2.0-mm-diameter excision wound adjacent to theimplant. (A) At 11 weeks after wounding, the implant (arrow) is associated with numerousmacrophages (M), fibroblasts (F), and and a granular/fibrous substance, probably fibrin (G).(B) In some regions, the fibroblasts and macrophages have entered the implant (I). Extra-cellular matrices, containing collagen fibrils, fill the lacuna (L) adjacent to fibroblasts. (C) By30 weeks, the implant is filled with fibroblasts and collagen-filled lacunae. No macrophagesare evident. (D) A dense extracellular matrix is p resent adjacent to the implant 30 weeksafter wounding, encapsulating the implant. The collagen fibrils are organized into smallbundles (lamellae) and separated by a dense material reminiscent of basal lamina.Collagen-filled lacunae appear to be contiguous with the extracellular matrix capsule.Bars = 5 mm.
FIG. 10. Electron microscopy of a 2.0-mm-diameter discof CxGelsix implanted in the corneal stroma 30 weeksafter a 2.0-mm-diameter excision wound was made adja-cent to the implant. (A) Bundles of collagen fibrils (arrows)embedded in the implant (I) are tightly grouped together inan apparently random fashion. (B) Collagen fibrils in la-mellae of the same cornea show highly organized fibrilsarranged parallel to each other. Bar = 1.0 mm.
CxGELSIX: BIOMATERIAL 201
Cornea, Vol. 19, No. 2, 2000

collagen, a normal constituent of corneal stroma, hasbeen shown to be an excellent substrate for corneal stro-mal cell binding in vitro (25). Furthermore, isolation andpreparation of CxGelsix from rabbit corneal stroma inthis study was accomplished by nondenaturing methods,increasing the probability that corneal cells in vivo willrecognize the native macromolecule in CxGelsix. TypeVI collagen is one of the earliest matrix proteins depos-ited during cell infiltration within a collagen tissue–polymer composite used to repair an abdominal wall de-fect, suggesting that it serves as a provisional matrix forcell migration (37).
Previous studies indicated that steroid treatment ofcorneas with intracorneal implants decreases postopera-tive complications (44). Moreover, some antiinflamma-tory agents have been shown to decrease the expressionof metalloproteinases in corneas containing intralamellarimplants (44). Although our study shows that fluoro-methalone acetate decreased postoperative complicationson a clinical level, no marked histologic or ultrastructuraldifferences in the integrity of the CxGelsix implant wereseen between steroid-treated and untreated eyes. Subtlequantitative differences or a measure of metalloprotein-ase activity, however, could not be determined with thetechniques used in this study.
Before CxGelsix can be considered as a biomaterial,rigorous immunologic studies are needed to test the an-tigenicity of this material, as well as the presence ofendotoxins. Acute inflammatory responses can be trig-gered by surrounding host proteins such as immunoglob-ulin G (IgG), complement, and fibrinogen. The latter,adsorbed fibrin on biomaterial, was found to mediate theacute response to biomaterials (45). The mechanism mayinvolve the denaturation of fibrin to expose previouslyhidden major histocompatibility complex (MHC) II–inducing epitopes. Moreover, activated macrophages,producing interleukin-1 (IL-1), may induce T-cell pro-duction of �-interferon (IFN) to stimulate the recruitmentof phagocytic macrophages and the formation of giantcells (35). Whether this mechanism is in operation in theavascular corneal stroma has yet to be determined.Complement activation provoked by surfaces on normalhuman sera is a marker able to predict the capacity ofbiomaterials to induce inflammation (46–48). Futurestudies will address the capacity of CxGelsix to inciteinflammation. Polyethylene glycol modification of anti-genic proteins abrogates the immunogenicity of the thenative protein (49), suggesting that our polyethylene gly-col–cross-linked Gelsix may render the type VI collagenmore tolerogenic. These results are encouraging and sug-gest that CxGelsix may be tolerated and useful as a bio-material.
The apparent resistance of CxGelsix to rapid degrada-tion by endogenous enzymes suggests that this materialmay facilitate anchorage of prosthetic devices to living
tissues, such as corneal prostheses. By virtue of its role asa substrate for fibroblasts, and resistance to rapid degra-dation, chemical cross-linking of Gelsix in vivo mayfunction as a biologic glue for healing wounds. It isanticipated that cross-links would form between the typeVI collagen and collagens at the cut edge of the wound,as well as between Gelsix filaments.
Acknowledgment: We thank Michael Martino, AnimalHealth Technologist, for his technical assistance. We also areindebted to Dr. Jose Conejo for his continuing support. Thisstudy was supported by the National Institutes of Health grantRO1-EYO1199.
REFERENCES
1. Zimmermann DR, Trueb B, Winterhalter KH, Witmer R, FischerRW. Type VI collagen is a major component of the human cornea.FEBS Lett 1986;197:55–8.
2. Trueb B, Schreier T, Bruckner P, Winterhalter KH. Type VI col-lagen represents a major fraction of connective tissue collagens.Eur J Biochem 1987;166:699–703.
3. Kuo H-J, Keene R, Glanville W. The macromolecular structure oftype VI collagen: formation and stability of filaments. Eur J Bio-chem 1995;232:364–72.
4. Doane KJ, Howell SJ, Birk DE. Identification and functional char-acterization of two type VI collagen receptors, �3�1 integrin andNG2, during avian corneal stromal development. Invest Ophthal-mol Vis Sci 1998;39:263–75.
5. Tillet E, Ruggiero F, Nishiyama A, Stallcup WB. The membrane-spanning proteoglycan NG2 binds to collagens V and VI throughthe central nonglobular domain of its core protein. J Biol Chem1997;272:10769–76.
6. Vogel KG, Paulsson M, Heinegard D. Specific inhibition of type Iand type II collagen fibrillogenesis by the small proteoglycan oftendon. Biochem J 1984;223:587–97.
7. Koller E, Winterhalter KH, Trueb B. The globular domains of typeVI collagen are related to the collagen-binding domains of carti-lage matrix protein and von Willebrand factor. EMBO J 1989;8:1073–7.
8. Bonaldo P, Russo V, Bucciotti F, Doliana R, Colombatti A. Struc-tural and functional features of the �3 chain indicate a bridging rolefor chicken collagen VI in connective tissues. Biochemistry 1990;29:1245–54.
9. Takahashi T, Cho H-I, Kublin C L, Cintron C. Keratan sulfate anddermatan sulfate proteoglycans associate with type VI collagen infetal rabbit cornea. J Histochem Cytochem 1993;41:1447–57.
10. McDevitt CA, Marcelino J, Tucker L. Interaction of intact type-VIcollagen with hyaluronan. FEBS Lett 1991;294:167–70.
11. Kielty CM, Whittaker SP, Grant ME, Shuttleworth CA. Type VIcollagen microfibrils: evidence for a structural association withhyaluronan. J Cell Biol 1992;118:979–90.
12. Bidanset DJ, Guidry C, Rosenberg LC, Choi HU, Timpl R, HookM. Binding of the proteoglycan decorin to collagen type VI. J BiolChem 1992;267:5250–6.
13. Hoylaerts MF, Yamamoto H, Nuyts K, Vreys I, Deckmyn H, Ver-mylen J. von Willebrand factor binds to native collagen VI pri-marily via its A1 domain. Biochem J 1997;324:185–91.
14. Finnis ML, Gibson MA. Microfibril-associated glycoprotein-1(MAGP-1) binds to the pepsin-resistant domain of the �3(VI) chainof type VI collagen. J Biol Chem 1997;272:22817–23.
15. Burg MA, Tillet E, Timpl R, Stallcup WB. Binding of the NG2proteoglycan to type VI collagen and other extracellular matrixmolecules. J Biol Chem 1996;271:26110–6.
16. Keene DR, Engvall E, Glanville RW. Ultrastructure of type VIcollagen in human skin and cartilage suggests an anchoring func-tion for this filamentous network. J Cell Biol 1988;107:1995–2006.
M. LEKSKUL ET AL.202
Cornea, Vol. 19, No. 2, 2000

17. Cintron C, Hong B-S. Heterogeneity of collagens in rabbit cornea:type VI collagen. Invest Ophthalmol Vis Sci 1988;29:760–6.
18. Rawe IM, Zhan Q, Burrows R, Bennett K, Cintron C. �ig: mo-lecular cloning and in situ hybridization in corneal tissues. InvestOphthalmol Vis Sci 1997;38:893–900.
19. Skonier J, Neubauer M, Madisen L, Bennett K, Plowman GD,Purchio AF. cDNA cloning and sequence analysis of �ig-h3, anovel gene induced in a human adenocarcinoma cell line aftertreatment with transforming growth factor-�. DNA Cell Biol 1992;7:511–22.
20. Zinn K, MacAllister L, Goodman CS. Sequence analysis and neu-ronal expression of fasciculin I in grasshopper and Drosophila.Cell 1988;53:577–87.
21. Takeshita X, Kikuno R, Tezuka K, Amann E. Osteoblast-specificfactor 2: cloning of a putative bone adhesion protein with homol-ogy with the insect protein fasciculin I. Biochem J 1993;294:271–8.
22. Gibson MA, Hatzinikolas G, Kumaratilake JS, et al. Further char-acterization of proteins associated with elastic fiber microfibrilsincluding the molecular cloning of MAGP-2 (MP 25). J Biol Chem1996;271:1096–103.
23. Hashimoto K, Noshiro M, Ohno S, et al. Characterization of acartilage-derived 66-kDa protein (RGD-CAP/�ig-h3) that binds tocollagen. Biochim Biophys Acta 1997;1355:303–14.
24. Gibson MA, Kumaratilake JS, Cleary EG. Immunohistochemicaland ultrastructural localization of MP78/70 (�ig-h3) in extracellu-lar matrix of developing and mature bovine tissues. J HistochemCytochem 1997;45:1683–96.
25. Doane KJ, Yang G, Birk DE. Corneal cell-matrix interactions: typeVI collagen promotes adhesion and spreading of corneal fibro-blasts. Exp Cell Res 1992;200:490–9.
26. Skonier J, Bennett K, Rothwell V, et al. �ig-h3: a transforminggrowth factor-�-responsive gene encoding a secreted protein thatinhibits cell attachment in vitro and suppresses the growth of CHOcells in nude mice. DNA Cell Biol 1994;13:571–84.
27. LeBaron RG, Bezverkov KI, Zimber MP, Pavelec R, Skonier J,Purchio AF. �ig-h3: a novel secretory protein inducible by trans-forming growth factor-�, is present in normal skin and promotesthe adhesion and spreading of dermal fibroblasts in vitro. J InvestDermatol 1995;104:844–9.
28. Lee SD, Hsiue GH, Chang PC, Kao CY. Plasma-induced graftedpolymerization of acrylic acid and subsequent grafting of collagenonto polymer film as biomaterials. Biomaterials 1996;17:1599–1608.
29. Desgrange P, Tardieu M. Loisance D, Barritault D. Extracellularmatrix covered biomaterials for human endothelial cell growth. IntJ Artif Organs 1992;15:722–6.
30. Rhee W, Wallace DG, Michaels AS, et al. Collagen-polymer con-jugates: patent number 5,162,430: date of patent: Nov. 10, 1992.
31. Kelman CD, DeVore DP. Collagen-based adhesives and sealantsand methods of preparation and use thereof: patent number:5,219,895. date of patent: June 15, 1993.
32. Kelman CD, DeVore DP. Method for bonding soft tissue withcollagen-based adhesives and sealants: patent number: 5,354,336:date of patent: Oct. 11, 1994.
33. Orgill DP, Ehret FW, Regan JF, Glowacki J, Mulliken JB. Poly-ethylene glycol/microfibrillar collagen composite as a new resorb-able hemostatic bone wax. J Biomed Mater Res 1998;39:358–63.
34. Thompson KP, Hanna KD, Gipson IK, Gravagna P, Waring GOIII, Johnson-Wint B. Synthetic epikeratoplasty in rhesus monkeyswith human type IV collagen. Cornea 1993;12:35–45.
35. van Luyn MJA, Khouw MSL, van Wachem PB, Blaauw IH, Werk-meister JA. Modulation of the tissue reaction to biomaterials: II.the function of T cells in the inflammatory reaction to crosslinkedcollagen implanted in T-cell-deficient rats. J Biomed Mater Res1998;39:398–406.
36. van Wachem PB, van Luyn MJA, Damink LO, Dijkstra PJ, FeijenJ, Neiuwenhuis P. Biocompatibility and tissue regenerating capac-ity of crosslinked dermal sheep collagens. J Biomed Mater Res1994;28:353–63.
37. Werkmeister JA, Edwards GA, Casagranda F, White JF, RanshawJAM. Evaluation of a collagen-based biosynthetic material for therepair of abdominal wall defects. J Biomed Mater Res 1998;39:429–36.
38. Chiou AG-Y, Mermoud A, Underdahl JP, Schnyder CC. An ul-trasound biomicroscopic study of eyes after deep sclerectomy withcollagen implant. Ophthalmology 1998;105:746–50.
39. Fujioka K, Maeda M, Hojo T, Sano A. Protein release from col-lagen matrices. Adv Drug Delivery Rev 1998:31:247–66.
40. Cintron C, Schneider H, Kublin C. Corneal scar formation. ExpEye Res 1973;17:251–9.
41. Everts V, Korper W, Neihof A, Jansen I, Beertsen W. Type VIcollagen is phagocytosed by fibroblasts and digested in the lyso-somal apparatus: involvement of collagenase, serine proteinasesand lysosomal enzymes. Matrix Biol 1994;14:665–76.
42. Cintron C, Covington H, Kublin CL. Morphogenesis of rabbitcorneal stroma. Invest Ophthalmol Vis Sci 1983;24:543–56.
43. Mast BA, Krummel TM. Acute inflammation in fetal wound heal-ing. In: Adzick NS, Longaker MT, eds. Fetal wound healing. NewYork: Elsevier, 1992:227–40.
44. Fitton JH, Ziegelaar BW, Hicks CR, et al. Assessment of anticol-lagenase treatments after insertion of a keratoprosthetic material inthe rabbit cornea. Cornea 1998;17:108–14.
45. Tang L, Eaton W. Inflammatory responses to biomaterials. Am JClin Pathol 1995;103:466–71.
46. Herzlinger GA, Bing DH, Stein R, Cumming RD. Quantitativemeasurement of C3 activation at polymer surfaces. Blood 1981;57:764–70.
47. Kochounian H, Maxweell A, Gupta A. Complement activation bysurface modified poly-methyl methacrylate intraocular lenses. JCataract Refract Surg 1991;17:139–42.
48. Verhagen C, Breebaart A, Kijlstra A. The effects of C� depletionon corneal inflammation in rats. Invest Ophthalmol Vis Sci 1992;33:273–9.
49. Ito H-O, Ueda T, Imoto T, Koga T. Prevention of collagen-inducedarthritis (CIA) by treatment with polyethylene glycol-conjugatedtype II collagen; distinct tolerogenic property of the conjugatedcollagen from the native one. Clin Exp Immunol 1997;108:213–9.
CxGELSIX: BIOMATERIAL 203
Cornea, Vol. 19, No. 2, 2000






![Preparation of [3H] Collagen for Studies of the Biologic ... · PDF filePreparation of [31-11 Collagen for Studies of the Biologic Fate of Xenogenic Collagen Implants In Vivo John](https://img.dokumen.tips/doc/110x75/5a9d47ca7f8b9abd058b5ce6/preparation-of-3h-collagen-for-studies-of-the-biologic-of-31-11-collagen.jpg)






















