Embed Size (px)
DESCRIPTION
molecular division
Citation preview

CELL DIVISION
Growth, repair and renewal in all multicellullar organisms depends on formation of new cells by division of pre-existing ones. This is mainly by mitosis in somatic cells and meiosis confined to development of germ cells.
Differentiation in plants refers to the processes by which distinct cell types arise from precursor cells and become different from each other. Plants have about a dozen basic cell types that are required for everyday functioning and survival. Additional cell types are required for sexual reproduction. Other cells such as the water-conducting vessel elements undergo cell death as part of their differentiation pathway.
Meristem Origins
Despite the differences among mature cells, all are ultimately derived from the apical meristems , populations of embryonic cells at the tips of the shoots and roots. Meristem cells are uniform in appearance: they are small and cuboidal in shape, and have a thin, flexible cell wall, a high nucleus to cytoplasm ratio, and dense cytoplasm with numerous ribosomes . The first step in the differentiation pathway is the formation of the precursors of the three tissue systems: protoderm (dermal tissue system), ground meristem (ground tissue system), and procambium (vascular tissue system).
Plant Tissue Systems
Each of the three tissue systems is found in a predictable location and consists of one or more multicellular tissues that carry out a unique function. For instance, the dermal tissue system is found at the surface of the plant and consists of a single tissue type, the epidermis. The epidermis functions to protect the plant from water loss, to permit gas exchange, and to provide a barrier to the invasion of harmful fungi and other microorganisms. The ground tissue system occurs internally to dermal tissue and may consist of three tissue types: parenchyma tissue, collenchyma tissue, and sclerenchyma tissue.

Parenchyma tissue is a multipurpose tissue that functions in photosynthesis and storage, while both collenchyma and sclerenchyma function to support the aboveground parts of the plant against the pull of gravity. The vascular tissue system occurs at the center of roots, stems, and leaves and functions in the long-distance transport of water and solutes. It consists of two tissues: xylem tissue and phloem tissue. Xylem provides a conduit for the movement of water and dissolved mineral elements from the roots to the shoot system. In contrast, phloem transports an aqueous solution of the products of photosynthesis from the green photosynthetic shoots to the roots and other parts of the plant that are using or storing food energy.
Overview of Plant Cell Types
Epidermal Cell Types. Epidermal cells are the most common cell type in the epidermis. These cells are often called "pavement cells" because they are flat polygonal cells that form a continuous layer, with no spaces between individual cells. Epidermal cells secrete the waxy hydrophobic substance cutin that polymerizes on the surface, forming a barrier to water evaporation. Epidermal cells are transparent because their plastids remain small and undifferentiated; hence light readily penetrates through to the photosynthetic tissues beneath the epidermis.
Two more specialized cell types are also found in the epidermis: guard cells and trichomes. Guard cells are kidney-shaped cells that are filled with chloroplasts. They always occur in pairs and form a small pore between them. The pair of guard cells and their pore is called a stomate and functions in gas exchange. Typically, the guard cells open the pore during daylight hours to allow CO 2 to diffuse into the photosynthetic tissues below. At night, however, the guard cells close the pore, preventing the diffusion of water vapor from internal tissues. The green chloroplasts of the guard cells function to provide the energy that fuels the opening and closing process.

Trichomes are long, narrow epidermal cells that grow perpendicular to the surface. Trichomes are either unicellular or multicellular and come in an amazing array of shapes. Some are branched and some are shield- or umbrella-shaped. Trichomes form a hairlike covering on the surface of leaves, stems, and roots and perform several important functions. The simple, unbranched trichomes of roots are called root hairs and function in the absorption of water from the soil. Trichomes on the surface of leaves and stems function primarily to retain water vapour and reduce the evaporative loss of water. Some trichomes secrete defense compounds that repel insect herbivores.
Ground Tissue Cell Types. Parenchyma cells are relatively unspecialized cells that make up the bulk of the soft internal tissues of leaves, stems, roots, and fruits. Parenchyma cells have thin, flexible cell walls and their cytoplasm typically contains a large, water-filled vacuole that fills 90 percent of the cell's volume. The vacuole may also contain compounds such as sugars (in

Figure; Distinct plant cell types are all ultimately derived from apical meristems.
sweet fruits such as apples, organic acids (as in oranges), or defense compounds such as the tannins found in tea leaves. Other specialized parenchyma cells may have starch-containing plastids such as those of potato tubers.
Collenchyma cells are long narrow cells with thick, strong, yet extensible, cell walls. Collenchyma cells are found in strands or sheets beneath the epidermis and function to provide support while a stem or leaf is still expanding. When growth is complete, sclerenchyma cells take over the role of providing mechanical support. Typical sclerenchyma cells are also long narrow cells (called fibers) with thick strong walls. Unlike collenchyma, the cell walls of sclerenchyma fibers are hard and rigid due to the deposition of lignin within the wall.
Fibers contribute more than half the volume of woody tissues and are found in all stems and leaves that are hard and tough. Other specialized sclerenchyma cells are shorter (sclereids) and have a protective function such as those of seed coats, walnut shells, or peach pits. Since the supportive or protective functions of sclerenchyma are carried out by the lignified cell walls, there is no requirement for living cytoplasm and these cells typically die as part of the differentiation process.
Vascular Tissue Cell Types. The vascular tissues are complex tissues, each consisting of a number of distinct cell types. The xylem contains conducting cells called vessel elements, as well as sclerenchyma fibers and parenchyma cells.
Vessel elements are highly specialized cells. Like sclerenchyma fibers, they form a thick cell wall that is impregnated with lignin. This cell wall can take many forms, depending on the time and location of its formation within the plant. In tissues that are growing, the lignified part of the cell wall is formed in rings or a helix, allowing the vessel elements to extend as the plant grows. In nonexpanding parts of the plant the cell wall forms as a netlike or pitted structure; these patterns contribute to the mechanical support of the plant and prevent cell collapse. Vessel elements are aligned end to end within the xylem. The part of the cell wall

between adjacent cells is degraded, so that the interior of all the vessel elements in a file becomes continuous, forming a vessel. Vessel elements are dead at maturity, leaving a hollow tube for the flow of water upward from the roots to the shoot system.
Phloem is also a complex tissue, containing two unique cell types, the sieve tube elements and companion cells, as well as parenchyma and sclerenchyma cells. Sieve tube elements are elongate cells with thick flexible cell walls. Adjacent cells are aligned end to end, forming the sieve tube. The end walls between adjacent cells have numerous pores; the sievelike appearance of these end walls led early microscopists to give these cells their name.
Sieve tubes transport the products of photosynthesis from the leaves to all parts of the plant where energy is needed or stored. The mechanism of transport requires that the sieve tube elements have a living cell membrane, but not other large components of the protoplast that might block the pores on the sieve plate. Thus mature sieve tube elements lack a vacuole, a nucleus, rough endoplasmic reticulum , and golgi. Sieve tube elements often live for years, but only because each is associated with a specialized parenchyma cell called a companion cell. The nucleus and cytoplasm of the companion cell must do the work for two cells, making and exporting the proteins required for sieve tube element function

CELL CYCLE
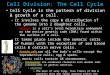
This is an ordered set of events, culminating in cell growth and division into two daughter cells. There are two main events; copying and cleaving. Somatic cells divide by mitosis or M-Phase which takes only a small segment the cell cycle, most of the cell’s life period is spent in interphase. Interphase is made up of cellular growth, DNA synthesis and duplication of other important cellar constituents.
Cell cycle is made of the following stages, G1, S-Phase, G2 and M-Phase (mitosis).
G1 PHASE
This is an intermediate phase occupying the time between the end of cell division in mitosis and the beginning of DNA replication during the S phase. During this time the cell grows in preparation for DNA replication, and certain intracellular components such as the centromeres undergo replication.
G1 phase is where cell monitoring occurs to ensure that the cell is biologically ready to take on such a process. The cell reviews the cellular environment and size to ensure that the conditions are appropriate to support DNA replication. If not the cell can pause during G1 and enters G0. G0 can last days , weeks or even years.

S PHASE
Synthesis phase, is the phase of the cell cycle when DNA packaged into chromosomes is replicated. This is an essential aspect of the cell cycle which allows for each cell created by cell division to have the same genetic make-up.
Other events occurring during this phase include, cell growth, synthesis of a number of proteins and enzymes involved in DNA synthesis. Once DNA replication is complete the cell contains twice its normal number of chromosomes and becomes ready to enter the G2 phase.
G2 PHASE
Similar to G1, G2 is an intermediate phase for the cell to ensure that it is ready to proceed to the next stage of the cell cycle which is Mitosis or M-phase.
G2 occurs between the end of DNA replication at S phase and the beginning of cell division at Mitosis. G2 is thought of as a safety gap during which a cell can check to make sure that the entire DNA and other intra cellular components have been properly duplicated. G2 also represents the cell’s final chance to grow before it splits into two independent cells.
MITOSIS
This is a process which involves nuclear and cytoplasmic division resulting into two indentical daughter cells.
General process; culture cells round up, chromosomes condense, mitotic spindle fibres form and the chromosomes separate to opposite spindle poles. It involves a five step process, and then a final culminating sixth step called cytokinesis( actual cleavage event/ splitting the cell into two).
The five steps of mitosis are prophase, prometaphase, metaphase, anaphase and telophase.
Prophase and prometaphase

Before cells enter M-Phase they must meet certain cellular requirements. Among these is appropriate cell size and cellular environment. Following DNA replication in S phase, cells contain twice their normal number of chromosomes because cell that undergo mitosis are diploid this means their number of chromosomes can be represented as 2N, where N equals the number of distinct chromosomes in the cell. Cells about to enter M phase i.e. those which have passed through S phase and replicated their DNA have 4N chromosomes.
Entry into the M phase is allowed by the formation of the mitotic cyclin-Cdk complex known as M phase promoting factor that occurs as a cell cycle regulatory mechanism in the G2 phase.
Prophase follows G2, at this point chromosomes condense or thicken and are visible under the microscope, still enclosed in the cell nucleus within the nuclear envelope. Microtubules begin to sprout from replicating centrosomes. Kinetochres assemble on the centromeres, kinetochore microtubules latter to these sites. Duplicated centrosomes (organizing centers of microtubules) begin to break down and the mitotic spindle forms. Mitotic spindle is an arrangement of microtubules that is responsible for aligning duplicated chromosomes in later phases.

Prometaphase
There Is a breakdown of nuclear envelope into small vesicles. Kinetochores become fully matured, disruption of the nuclear envelope allows for the mitotic spindles to have access to the mature kinetochores.As the microtubules of the mitotic spindle enter the nuclear region, some attach to the kinetochores making the kinetochore microtubules. Once they have captured chromosomes, the kinetochores microtubule begin to exert force on the chromosomes, moving them.
Metaphase and anaphase
Alignment of chromosomes at the center of the cell occurs and subsequent sister chromatids separate to opposite mitotic spindle poles.
Metaphase
Marked by alignment of the chromosomes at the center of the cell, half way between each of the mitotic spindle poles. Movement is mediated by kinetochore microtubules which pull and push on the chromosomes to align them into what is called the metaphase plate.

Metaphase can take up a large portion of the total time of mitosis because chromosome alignment at the center of the cell on the metaphase plate acts as a check point for progression to the next stage, anaphase.
Anaphase
Entrance into anaphase is triggered by the inactivation of M phase-promoting factor that allows mitotic cyclin degreadtion. During this stage the kinetochore microtubules retract, increasing the separation of the sister chrmatids as they are moved further towards the opposite spindle poles.
Anaphase has two distinct phases;
Anaphase A, where chromosomes move poleward away from the metaphase plate with the retraction of microtubules.
Anaphase B, the mitotic poles marked by centrosomes themselves separate by the elongation of a specific type of non-kinetochore microtubule called a polar microtubule.

Telophase and cytokinesis
The final two stages of M phase are the re-forming of the nuclear envelope around the separated sister chromatids and the cleavage of the cell. These events occur in telophase and cytokinesis, respectively.
Telophase
Technically the final stage of mitosis, during this phase the sister chromatids reach the opposite poles. The small nuclear vesicles in the cell begin to re-form around the group of chromosomes at each end, two nuclei are created in one cell. Telophase marks the dissolution of the kinetochore microtubules and the continued elongation of the polar microtubules. As the nuclear envelopes re-form, the chromosomes begin to decondense and become more diffuse.

Cytokinesis
Process in which the cell actually didvdes into two. With the two nuclei already at the opposite poles of the cell, cytoplasm seperates and cell pinches in the middle ultimately leading to cleavage. Cleavage occur by contraction of a thin ring of actin filaments that forms a contractile ring.

The passage through cell cycle is driven by a small group of proteins mainly of two type, i.e. cycline and cycline dependent kinases Cdks. The Cdk driving the cell through M-phase is Cdk 1 and through the S-phase is Cdk 2. These proteins Cdk 1 & 2 are present at constant amounts at all phases of the cell cycle but enzymatic activites of the two proteins are turned on at key cell cycle transition points by a small family of proteins called cyclins.


FERTILIZATION AND EMBRYONIC DEVELOPMENT
Fertilization is a complex mechanism that has evolved in flowering plants. This process involves the joining of a female gametophyte (embryo sac) with two male gametes (sperm). It begins when a pollen grain adheres to the stigma of the carpel, the female reproductive structure of a flower. After a pollen grain has landed on an accessible stigma, the pollen grain takes in moisture and begins to germinate, forming a pollen tube that extends down toward the ovary through the style. The tip of the pollen tube then enters the ovary and penetrates through the micropyle. The micropyle is an opening in the protective layers of the ovule. The pollen tube proceeds to release the two sperm in or near the embryo sac.
One sperm fertilizes the egg cell and the other sperm combines with the two polar nuclei of the large central cell of the embryo sac. The sperm and haploid egg combine to form a diploid zygote, while the other sperm and two haploid polar nuclei form a triploid nucleus (some plants may form polyploid nuclei). The large cell of the embryo sac will then form the endosperm, a nutrient-rich tissue which provides nourishment to the developing embryo. The ovary, surrounding the ovules, develops into the fruit, which is used for protection and dispersion of the seeds.
The two central cell maternal nuclei (polar nuclei) that contribute to the endosperm, arise by mitosis from a single meiotic product. Therefore, maternal contribution to the genetic constitution of the triploid endosperm is different from that of the embryo.


REFFERENCES (17th May 2011)
commons.wikimedia.org/wiki/File:Cell_cycle_an...
en.wikipedia.org/wiki/Double_fertilization
www.ba-education.com/for/science/dnabiology.html
www.oncolink.org/.../article.cfm?c=2&s=9&id=319
http://www.school.net.th/library/create-web/10000/science/10000-6584/pic1.jpeg

COURSE: MSC. MOLECULAR BIOLOGY AND BIOTECHNOLOGY
COURSE UNIT: CROP BIOTECHNOLOGY MBS 7221
NAME: NASSIMBWA FLORENCE
REG NO: 2010/HD17/317U
COURSE WORK TOPICS;
Cell division, differentiation and development.
Cell cycle- mitosis and meiosis.
Fertilization and embryonic development.
LECTURER: PROF. RUBAIHAYO
MAY 2010