Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1985 by The American Society of Biological Chemists, Inc.
Vol ,260, No. 11, Issue of June 10, pp. 6761-6768,1985 Printed in U. 5’. A .
Coupling of the Guanine Nucleotide Regulatory Protein to Chemotactic Peptidekeceptors in Neutrophil Membranes and Its Uncoupling by Islet-activating Protein, Pertussis Toxin A POSSIBLE ROLE OF THE TOXIN SUBSTRATE IN Ca2+-MOBILIZING RECEPTOR-MEDIATED SIGNAL TRANSDUCTION*
(Received for publication, December 12, 1984)
Fumikazu Okajima, Toshiaki Katada, and Michio Ui From the Department of Physiological Chemistry, Faculty of Pharmaceutical Sciences, Hokkaido University, Sapporo 060, Japan
A chemotactic peptide stimulated the high-affinity GTPase activity in membrane preparations from guinea pig neutrophils. The enzyme stimulation was inhibited by prior exposure of the membrane-donor cells to islet-activating protein (IAP), pertussis toxin, or by direct incubation of the membrane preparations with its A-protomer (the active peptide) in the presence of NAD. The affinity for the chemotactic peptide bind- ing to its receptors was lowered by guanyl-5”yl @,r- imidodiphosphate (Gpp(NH)p) reflecting its coupling to the guanine nucleotide regulatory protein in neutro- phils. The affinity in the absence of Gpp(NH)p was lower, but the affinity in its presence was not, in the A-protomer-treated membranes than in nontreated membranes. The inhibitory guanine nucleotide regu- latory protein of adenylate cyclase (Ni) was purified from rat brain, and reconstituted into the membranes from IAP-treated cells. The reconstitution was very effective in increasing formyl-Met-Leu-Phe-depend- ent GTPase activity and increasing the chemotactic peptide binding to membranes due to affinity increase. The half-maximal concentration of IAP to inhibit GTPase activity was comparable to that of the toxin to inhibit the cellular arachidonate-releasing response which was well correlated with ADP-ribosylation of a membrane M, = 41,000 protein (Okajima, F., and Ui, M. (1984) J. Biol. Chem. 259, 13863-13871). It is proposed that the IAP substrate, Ni, couples to the chemotactic peptide receptor and mediates arachidon- ate-releasing responses in neutrophils, as it mediates adenylate cyclase inhibition in many other cell types.
Islet-activating protein (IAP’), pertussis toxin, is a specific modifier of the guanine nucleotide regulatory protein (Ni) that is involved in receptor-mediated inhibition of adenylate cyclase. The GTP-binding a subunit of Ni with M, = 41,000
* This work was supported by research grants from the Scientific Research Fund of the Ministry of Education, Science, and Culture, Japan. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisernent” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
The abbreviations used are: IAP, islet-activating protein; N e t - Leu-Phe, N-formyl-methionyl-leucyl-phenylalanine; Hepes, 4-(2-hy- droxyethy1)-1-piperazineethanesulfonic acid; EGTA, ethylene glycol bis(P-aminoethyl ether)-N,N,N’,N’-tetraacetic acid; IBMX, 3-iso- butyl-1-methylxanthine; Gpp(NH)p, guanyl-5”yl P,y-imidodiphos- phate; App(NH)p, adenyl-5”yl P,y-imidodiphosphate; N. and Ni, the guanine nucleotide regulatory proteins responsible for stimulation and inhibition of adenylate cyclase, respectively.
is the substrate of IAP-catalyzed ADP-ribosylation. Since Ni loses its function to communicate between receptors and the adenylate cyclase catalyst after the ADP-ribosylation, recep- tor-mediated inhibition of adenylate cyclase is reversed, and activation is often enhanced, by IAP treatment of a variety of cell types (see Ref. 1 for review).
In the case of guinea pig neutrophils, however, the IAP- sensitive Ni (or the Ni-like protein with M , = 41,000) behaved as if it were a communicator in another biosignaling system in which adenylate cyclase was not involved directly (2). Incubation of neutrophils with Met-Leu-Phe caused a rapid increase in 0; generation, which was associated with arachi- donate release and Ca2+ mobilization. These effects of the chemotactic peptide were markedly attenuated by prior ex- posure of the cells to IAP. The degree of the IAP-induced attenuation was well correlated with the degree of ADP- ribosylation of a M, = 41,000 membrane protein, but was not accompanied by a reasonable change in the cAMP content (2). We have then proposed that Ni is also involved in signal transduction via the formyl-peptide receptors leading to Ca2+ mobilization and arachidonate release in neutrophils.
Koo et al. (3) showed that the binding of the formyl-peptide to its receptor in neutrophil membranes is regulated by guan- ine nucleotides; the addition of GTP analogues in the binding assay medium shifted the formyl peptide binding curve to the right. Furthermore, the formyl-peptide is capable of stimulat- ing GTPase activity in homogenates of neutrophils (4). These results suggest that a guanine nucleotide regulatory protein may interact with the formyl-peptide receptor rather directly. The purpose of the present paper is to show that the guanine nucleotide regulatory protein implied to couple to formyl- peptide receptors by the guanine nucleotide effect on receptor agonist binding or by receptor-mediated GTPase activity is an entity indistinguishable from the IAP substrate. It is probable that the IAP substrate plays an additional role in intracellular signaling processes arising from Ca2+ mobiliza- tion receptors in neutrophils.
EXPERIMENTAL PROCEDURES
Materials-IAP was purified from the 2-day culture supernatant of Bordetella pertussis (5). Its A-protomer, the peptide displaying ADP-ribosyltransferase activity, was dissociated from the residual subunits by maintaining the protein solution in 5 M urea a t 4 “C for 4 days and then purified according to the procedure described else- where (6). N-Formyl-Met-Leu-Phe was purchased from Sigma, [5,6,8,9,11,12,14,15-3H]arachidonic acid (70 Ci/mmol) and [y-”PI GTP (15 Ci/mmol) from Amersham, [w3*P]NAD (29 Ci/mmol) and flMet-Le~-[~H]Phe (50 Ci/mmol) from New England Nuclear. Re- agents for radioimmunoassay of cAMP were obtained from a Yamasa cAMP assay kit which was a kind gift from Yamasa Shoyu Co.
6761

6762 Uncoupling of Formyl-peptide Signal Transduction by IAP
(Choshi, Chiba). The sources of all other materials used are those described in the previous paper (2).
Isolation and Pretreatment of Neutrophils-Male Hartley guinea pigs (500 to 700 g, body weight) were injected intraperitoneally with 15 ml of 3% sodium casein 18 h before killing (7). Neutrophils were isolated from the peritoneal exudate of these animals as described in the previous paper (2). Where indicated, neutrophils were exposed to 100 ng/ml (unless otherwise specified) of IAP for 4 h a t 37 "C in TCM 199 culture medium (Earle's salt, pH 7.4) under a humidified atmosphere containing 5% COZ as described previously (2). [3H] Arachidonic acid (1 pCi/ml) was also added to the medium to prelabel the cellular phospholipid fraction with the radioactivity. These IAP- treated, 3H-labeled cells or nontreated or nonlabeled cells were sus- pended in the Ca'+-free Hepes-buffered medium after twice washing with the same medium. This medium was prepared by omitting CaC1, from, and further adding 0.1 mM EGTA to, the regular Hepes-buffered medium consisting of 10 mM Hepes, 134 mM NaCl, 4.7 mM KCl, 1.2 mM KHzPO,, 1.2 mM MgSO4, 2.5 mM CaCl,, 5 mM NaHC03, and 0.1% bovine serum albumin (Fraction V).
Arachidonic Acid Release from PHIArachidonate-labeled Cells- The cell suspension prepared as above (2 X lo6 cells in 0.1 ml) was incubated for 10 min at 37 "C with the same volume of the regular Hepes-buffered medium containing 0.1 p~ fMet-Leu-Phe. The reac- tion was terminated by the addition of 0.5 ml of ice-cold 0.9% NaCl, 2 mM EGTA followed by immediate centrifugation at 10,000 X g for 1 min. Radioactivity in the supernatant was measured as a reflection of the phospholipase A, activity as previously described (2).
Cyclic AMP Content-The cell suspension (2 X lo6 cells in 0.1 ml) was quickly mixed with the same volume of the regular Hepes- buffered medium containing 0.2 mM IBMX and 0.1 p~ Met-Leu- Phe and/or 10 p~ prostaglandin E2 then incubated for 2 min at 37 "C. Incubation was stopped by the addition of 0.1 ml of 0.4 N HCI followed by boiling for 2 min. Cyclic AMP in the supernatant was measured by a sensitive radioimmunoassay procedure described by Honma et al. (8).
Preparation of Neutrophil Membranes-Freshly prepared or 4-h treated (as above) neutrophils were homogenized with a Polytron homogenizer (PT 10-20-3500 type) for 30 s in a cold hypotonic medium consisting of 25 mM Tris buffer (pH 7.6), 5 mM MgC12 and 50 units/ml of aprotinin. The homogenate was then centrifuged a t 150 X g for 5 min and the resultant supernatant was collected. The procedure was repeated the pellet was once again homogenized in the same buffer, and the homogenate was centrifuged. The pooled supernatant was centrifuged a t 10,000 X g for 5 min. The pellet was suspended in the same buffer (1-2 mg of protein/ml) then stored a t -80 "C until use.
Treatment of Membranes with the Preactiuated A-protomer of IAP or the Al-fragment of Cholera Toxin-The A-protomer of IAP was used for experiments after being preactivated at 30 "C for 10 min with 20 mM dithiothreitol (9). In Table I only was used the native IAP similarly preincubated with dithiothreitol. Cholera toxin was also incubated a t 30 "C for 10 min with 20 mM dithiothreitol to release the enzymatically active Al-fragment (10). For the treatment with the A-protomer of IAP (or the native IAP in some experiments shown in Table I), the membranes (0.5-2 mg of protein) were incu- bated with 20 pg/ml of the A-protomer of IAP (or 50 pg/ml of the native IAP) a t 30 "C for 15 min in 1 ml of the ADP-ribosylation reaction mixture consisting of 25 mM Tris buffer (pH 7.6), 10 mM NAD, 1 mM ATP, 1 mM EGTA, 10 mM thymidine, 2 mM dithio- threitol, 2.5 mM MgC12, 3 mM potassium phosphoenolpyruvate and 10 pg/ml of pyruvate kinase. For cholera toxin treatment, membranes were similarly incubated with the preactivated cholera toxin (100 pg/ ml) in the same medium as used for IAP treatment except that 25 mM Tris was replaced by 100 mM potassium phosphate buffer (pH 7.5) and 0.1 mM GTP was further added. In experiments shown in Fig. 3, where the effects of both toxins were compared, treatment of membranes with the A-protomer was done under the same conditions as those employed for cholera toxin treatment. These toxin treat- ments were terminated by dilution with 3 ml of the assay buffer for GTPase or fMet-Le~-[~H]Phe binding (see below) followed by cen- trifugation a t 10,000 X g for 5 min; the pellet was washed once by repeating dilution and centrifugation in the same buffer.
Purification of N;-N, was purified to near homogeneity from rat brain by the procedures adapted for purification of Ni from rabbit liver (11) with a slight modification. Detailed procedures will be
described elsewhere.' Ni (234 pg/ml) dissolved in 50 mM Hepes buffer (pH 8.0) containing 1 mM EDTA, 1 mM dithiothreitol, and 0.1% Lubrol (henceforth referred to as the HEDL medium) was used for reconstitution experiments.
Reconstitution of Ni into Neutrophil Membranes-Membranes, which were prepared from neutrophils treated or not treated with IAP, were used in the reconstitution experiments. Various concentra- tions of Ni (1.17-7.02 pg of protein in 30 pl of the HEDL medium) were added to the membranes (approximately 200 pg of protein in 450 pl) suspended in 25 mM Tris buffer (pH 7.6) containing 1 mM ATP, 5 mM phosphocreatine, 50 units/ml of creatine phosphokinase, 5 mM MgC12, and 50 units/ml of aprotinin. After incubation for 10 min at 30 "C, the mixture was diluted with 6 volumes of the ice-cold buffer used for the following GTPase, fMet-Le~-[~H]Phe binding, or ADP-ribosylation assay. The mixture was then centrifuged a t 10,000 X g for 5 min to obtain the membrane pellet, which was washed only by repeated dilution and centrifugation in the same buffer prior to further manipulation.
Radiolabeling of Neutrophil Membranes with /a-32PlNAD-Mem- branes reconstituted with Ni (approximately 100 pg of protein) were incubated for 60 min at 30 "C with the A-protomer of IAP (20 pg/ml) in 100 p1 of the ADP-ribosylation reaction mixture (see above) containing 10 p M of [w3'P]NAD (5 pci) in place of 10 mM NAD. Reaction was terminated by adding Laemmli sample buffer (12) followed by boiling for 3 min. The radiolabeled membranes were submitted to sodium dodecyl sulfate-polyacrylamide slab gel electro- phoresis and autoradiography as described previously (13, 14).
GTPase Assay-GTPase activity was measured based on 32Pi lib- eration from [Y-~'P]GTP by the procedure of Cassel and Selinger (15) with slight modifications as shown elsewhere (16). The neutro- phil membranes (20-30 pg of proteins) suspended in 0.1 ml of 25 mM Tris buffer (pH 7.6) containing 1 mM EGTA were incubated a t 37 "C with 0.5 mM App(NH)p, 0.5 mM MgCl,, 1 mM dithiothreitol, 0.2 mM IBMX, 0.25 mM ATP, 5 mM phosphocreatine, 50 units/ml of creatine phosphokinase, 0.1% bovine serum albumin, and [Y-~'P]GTP (at the concentration of 0.25 p M unless otherwise specified). Further addi- tions are shown in figures or tables. The reaction was terminated by the addition of 1 ml of a 5% suspension of charcoal (Norit A) containing 0.5% bovine serum albumin and 0.1% dextran (M, = 200,000) in 20 mM phosphate buffer (pH 7.5). The assay tubes were centrifuged to sediment the charcoal, and 0.5-ml aliquots of the supernatant were counted for 32Pi.
Formyl-Met-Leu-pH]Phe Binding to Neutrophil Membranes- F~rmyl-Met-Leu-[~H]Phe binding was performed by the procedure described by Koo et al. (3) with slight modifications. Neutrophil membranes (20-30 pg of protein) were incubated for 30 min a t 25 "C in 6 mM phosphate buffer (pH 7.4) containing 140 mM NaCl, 1.0 mM KH2P04, 5 mM Na2HP04, 0.5 mM MgCl,, 0.15 mM CaC12, and 0.1% bovine serum albumin in the final volume of 0.2 ml. The assay medium was further supplemented with the indicated concentration of fMet-Le~-[~H]Phe (1 nM-2 pM) and 100 p M Gpp(NH)p (where indicated). The reaction was terminated by dilution with 5 ml of the fresh ice-cold assay mixture; it was followed by immediate filtration through a glass fiber filter Whatman GF/C (2.4-cm diameter). The filter was rapidly rinsed with 5 ml of the same buffer three times. The retained radioactivity was determined after the filters were dried in an oven at 60 "C. Specific binding was assessed from the difference between bindings in the absence and presence of 1 mM cold M e t - Leu-Phe.
Since Scatchard plots (shown in Fig. 4B) of the binding were curvilinear, the data for the binding assay were fitted by a nonlinear least squares analysis to the binding equation,
for a two-site model. B = total binding (equivalent to the total number of receptors) at a given concentration of free ligand ( F ) ; B,(1) or B,,(2) = the number of a high-affinity receptor (RH) or a low-affinity receptor (RL), respectively; andKd(1) or &(2) = dissociation constant of the ligand to RH or RL, respectively.
Data Presentation-Experiments in which duplicate or triplicate observations were done with a single batch of cells or membranes were repeated at least twice. Representative results are shown as means -t S.E., when essentially the same effects were repeatedly observed.
' T. Katada, and M. Ui, manuscript in preparation.

Uncoupling of Formyl-peptide
RESULTS
Inhibition of Membrane GTPase Actiuity by Treatment of Neutrophils with ZAP-Fig. 1 shows time courses of 3zPi liberation when neutrophil membranes were assayed for GTPase activity in the presence of 0.25 FM [-p3’P]GTP. In the case of control membranes, which were prepared from neutrophils not treated with IAP, 32Pi liberated almost linearly during 2-min incubation, then the rate of 32Pi liberation declined gradually. The addition of Met-Leu-Phe to the assay mixture caused approximately an 80% increase in GTPase activity during the first 2-min incubation. There was no further effect of the addition on the liberation rate after 2 min. When membranes prepared from IAP-treated neutro- phils were assayed for GTPase activity, the basal activity (i.e.
FIG. 1. Time courses for GTP hydrolysis by membranes from IAP-treated or nontreated neutrophils. Membranes were prepared from neutrophils treated (A and A) or not treated (0 and 0) with IAP. They were assayed for GTPase in the presence (0 and A) or absence (0 and A) of 1 p~ met-Leu-Phe as described under “Experimental Procedures.” GTPase activity shown on ordinate is expressed as GTP hydrolysis (pmol)/mg of protein. Each point is the mean f S.E. of triplicate incubations.
TABLE I NAD requirement for IAP or its A-protomer to inhibit GTPase
activity Neutrophil membranes were incubated with IAP (50 Fg/ml) or its
A-protomer (20 pg/ml) in the presence or absence of 10 mM NAD as described under “Experimental Procedures.” GTPase activity was assayed by incubating these membranes for 5 min with 0.25 p~ [y- 32P]GTP in the absence (control) or presence of 1 FM Met-Leu-Phe. Results are mean & S.E. of three observations. The increments in GTPase activity by the formvl-peotide are shown in parentheses.
Treatment GTPase
Control Met-Leu-Phe pmot/mg/min
None NAD
30.8 + 0.8 40.3 t 0.3 (9.5)
IAP 30.4 f 0.6 42.2 2 1.1 (11.8)
IAP + NAD 28.3 f 0.6 40.5 & 0.2 (12.2) 19.1 -C 0.4” 25.0 t 0.5” (5.9)
A-protomer 31.0 f 0.5 40.9 & 0.8 (9.9) A-protomer + NAD 15.7 2 0.3” 17.3 & 0.3” (1.6) Difference from none was significant; p < 0.01.
Signal Transduction by IAP 6763
the enzyme activity in the absence of Met-Leu-Phe) was 20- 30% less than that of control membranes. A much more pronounced effect of IAP was observed when the enzyme activity was assayed in the presence of Met-Leu-Phe; only a 10-20% increase in the activity was caused during the initial 2 min by the addition of the formyl-peptide (Fig. 1).
Direct Action of ZAP or Its A-protomer on GTPase activity of Membrane Preparations from Neutrophils-Membrane GTPase activity was not affected by the direct addition of IAP alone to the membrane (Table I). The inclusion of NAD, the substrate for the ADP-ribosylation reaction, in the IAP containing medium was effective in inhibiting both basal and Met-Leu-Phe-stimulated GTPase activity, despite the failure of NAD by itself to exert any significant effect. A similar, and much greater, effect was obtained when the A-protomer, an active peptide of IAP, was used in place of the native IAP (Table I). The requirement for NAD suggests that ADP- ribosylation of the membrane protein is involved in the IAP (or the A-protomer)-induced inhibition of GTPase activity of neutrophil membranes. NAD was included in the medium when membranes were directly treated with the A-protomer of IAP in the following experiments.
Characterization of fMet-Leu-Phe-induced and ZAP-sensi- tive GTPase of Neutrophil Membranes-The rate of GTP hydrolysis was plotted against various concentrations of GTP in Fig. 2 A . No saturation kinetics was observed for the basal GTPase activity, probably due to the high activity of low- affinity and nonspecific nucleoside triphosphatase as reported by Cassel and Selinger (15). In contrast, met-Leu-Phe-stim- ulated GTPase activity reached a plateau at concentrations of GTP around 1 ~ L M in the control membranes not treated with IAP (Fig. 2C). Lineweaver-Burk plots of GTPase at lower concentrations of GTP (Fig. 2B) showed that met- Leu-Phe increased V,,,,,, without changing K,, of the high- affinity GTPase activity; V,,, (pmol/mg/min) and K, (PM) of the enzyme activity was 84.1 k 5.5 and 0.24 ? 0.02 without Met-Leu-Phe and 116.7 ? 9.5 and 0.22 f 0.02 with the formyl-peptide, respectively.
The GTPase activity of membranes which were treated with the A-protomer of IAP was less than that of control membranes at the lower concentrations of GTP regardless of whether Met-Leu-Phe was present or not; however, the ac- tivities gradually reached the same level as that of control membranes as the concentration of GTP increased (Fig. 2A) . In this case, the Net-Leu-Phe-stimulated GTPase activity also reached a plateau at the concentration of GTP around 1 PM, though the activity was much less than that of control membranes (Fig. 2C). V,,, (pmol/mg/min) and K, ( p ~ ) of the high-affinity enzyme activity were calculated from Line- weaver-Burk plots (Fig. 2B) as 35.7 ? 1.5 and 0.21 & 0.02 without Met-Leu-Phe and 44.9 ? 1.9 and 0.24 f 0.02 with the formyl-peptide, respectively. Thus, the treatment of mem- branes with the A-protomer of IAP suppressed both basal and Met-Leu-Phe-stimulated high-affinity GTPase activity by lowering V,,,.,, without any change in K,,,, of the enzyme.
The dose-response effect of Net-Leu-Phe on the high- affinity GTPase is shown in Fig. 3, where the action of cholera toxin, which is known to inhibit the high-affinity GTPase activity induced by various hormones to stimulate adenylate cyclase (17, la), is also shown for comparison. The treatment of membranes with the A-protomer of IAP inhibited M e t - Leu-Phe-stimulated enzyme activity at all the concentrations of the formyl-peptide employed in agreement with the results in Table I and Fig. 2. In sharp contrast, cholera toxin exerted no significant effect on Met-Leu-Phe-induced GTPase activ- ity (Fig. 3). The neutrophil membranes were susceptible to

6764 Uncoupling of Formyl-peptide Signal Transduction by IAP
‘i 0 25 u, 0 /*-*! O 0.01 0.1 1 10 100
GTP QMl FIG. 2. GTPase activity at various GTP concentrations of
neutrophil membranes treated or not treated with the A- protomer of IAP. In A , neutrophil membranes treated (A and A) or not treated (0 and 0) with the A-protomer of IAP were assayed for GTPase activity at the indicated concentration of GTP in the presence (0 and A) or absence (0 and A) of 1 FM met-Leu-Phe as described under “Experimental Procedures.” Incubation time was 2 min. The mean f S.E. of three separate experiments are plotted as a function of GTP concentrations; the scale for GTPase activity esti- mated with GTP below 1 PM is shown on the left-hand while that above 1 FM is on the right-hand ordinate. Inset B shows the Line- weaver-Burk plots of the mean values of the enzyme activity at the concentrations from 0.05 to 1 PM GTP. The data are from A. In C, Net-Leu-Phe-stimulated enzyme activity (i.e. the increment of the activity induced by the formyl-peptide) of membranes treated (0) or not treated (0) with the A-protomer of IAP is plotted against GTP concentrations. The data were from A . The values below 8 PM GTP are shown in C, since there were big variations in the values at the higher concentrations at which the formyl-peptide-stimulated enzyme activities were less than 10% of the total activities.
cholera toxin, as revealed by high adenylate cyclase activity after the toxin treatment; the adenylate cyclase activity (pmol/mg/min) in the presence of 10 ~ L M GTP was 70 k 6 for nontreated membranes and 185 f 9 for the toxin-treated membranes, respectively (number of observations was three).
Formyl-Met-Leu-PHIPhe Binding to Neutrophil Mem- branes Treated or Not Treated with IAP (A-protomer) in the Presence or Absence of Gpp(NH)p-Specific Met-Leu-Phe- [“HJPhe binding to neutrophil membranes was plotted against concentrations of the labeled peptide in Fig. 4A. The binding did not saturate at concentrations of the ligand employed in the present study. To estimate the maximal binding, Scat- chard analysis was achieved. The Scatchard plots were cur- vilinear (Fig. 5B), indicating the existence of multiple inde- pendent sites for the peptide binding. The maximal binding (I?,,,, pmol/mg of protein) in the absence or presence of Gpp(NH)p, a nonhydrolyzable GTP analogue, was estimated from Scatchard plots of the binding at the higher concentra- tion of the ligand (Fig. 4C) as 4.49 k 0.54 or 4.31 f 0.64 for
Met-Leu-Phe (-log MI FIG. 3. Dose-response curves for Met-Leu-Phe-induced
GTPase activity in neutrophil membranes. Neutrophil mem- branes, treated with the A-protomer of IAP (O), treated with preac- tivated cholera toxin (A), or not treated (O), were assayed for GTPase in the presence of the indicated concentration of met-Leu-Phe. The incubation time was 5 min. GTPase activity stimulated by met-Leu- Phe is shown on the ordinate. Results are the mean f S.E. of three separate experiments.
control membranes not treated with the toxin and 4.48 f 0.50 or 4.35 k 0.66 for the membranes treated with the A-protomer of IAP, respectively. These values were slightly less than those estimated from computer analysis (see Table 11).
Although treatment of membranes with the A-protomer thus did not alter B,,,, the binding at lower concentrations of the ligand was significantly less in the toxin-treated mem- branes than that in control membranes unless Gpp(NH)p was added to the assay medium; apparent dissociation constant (i.e. the concentration of the ligand for the half-maximal binding (nM) was 16.0 f 2.6 for control membranes and 89.8 f 24.0 for the toxin-treated membranes, respectively (Fig. 4A). When Gpp(NH)p was present during binding assay, the formyl-peptide binding curve was shifted to the right 20-fold in the control membranes and 4.5-fold in the toxin-treated membranes, respectively. As a result, there was no essential difference in the apparent affinity for fMet-Le~-[~H]Phe binding to its receptors between control membranes and the toxin-treated membranes; apparent dissociation constant (nM) was 335 f 31 for control membranes and 401 f 67 for the toxin-treated membranes, provided the incubation me- dium was supplemented with Gpp(NH)p (Fig. 4A).
To obtain the more accurate parameters for the formyl- peptide binding to neutrophil membranes, computer analysis of these data was achieved by fitting to a two-site model in Table 11. Neither the total receptor number nor the K d values of a high-affinity receptor (RH) and a low-affinity receptor ( R L ) were significantly changed by a prior treatment of mem- branes with the A-protomer of IAP and/or an addition of Gpp(NH)p during binding assay (Table 11). Incubation with Gpp(NH)p, however, resulted in a decrease in the number of RH to 40%, associated with an increase in the number of RL to 230%, of its absence in the control membranes not treated

Uncoupling of Formyl-peptide Signal Transduction by IAP 6765
5 .̂
Bound [pmol/mgl FIG. 4. Formyl-Met-Le~-[~H]Phe binding to neutrophil
membranes. Neutrophil membranes treated (A and A) or not treated (0 and 0) with the A-protomer of IAP were incubated with the indicated concentration of fMet -Le~- [~H]Phe in the presence (0 and A) or absence (0 and A) of 100 p~ Gpp(NH)p. The specific binding for the labeled ligand to membranes, which was measured as described under “Experimental Procedures,” was plotted as a function of the concentration of the peptide in A. Scatchard plots were shown in B and C; the ratio of bound peptide to free peptide (bound/free, pmol/ mg, nM ) was plotted as a function of bound peptide (bound, pmol/ mg). In C, only the values calculated from the data at the concentra- tions of the ligand above 500 nM were plotted. The results shown are representative of four separate experiments.
with the A-protomer of IAP (Table 11). Thus, the ratio of the number of RH to RL was decreased from 1.96 to 0.32 by the addition of Gpp(NH)p. These data are consistent with a notion that Gpp(NH)p induces interconversion of RH to RL in neutrophil membranes.
When membranes treated with the A-protomer of IAP were used, the number of RH was decreased to 61% of that of nontreated membranes, which was associated with a 2.2-fold increment in the number of RL, in the absence of Gpp(NH)p during binding assay (Table 11). In the presence of Gpp(NH)p, however, no change was provoked in numbers of RH or RL by the treatment of membranes with the A-protomer of IAP (Table 11). Thus, IAP treatment of neutrophil membranes
- 20-
15 -
10 -
5 -
0 - 75 -
50-
25 -
0 L
FIG. 5. The effects of Ni reconstituted to membranes on ADP-ribosylation and Met-Leu-Phe-stimulated GTPase ac- tivity. Membranes, which were prepared from neutrophils treated (0) or not treated (0) with IAP, were reconstituted with the indicated concentration of Ni as described under “Experimental Procedures.” In A, the reconstituted membranes with Ni were incubated with 10 p~ [a-3’P]NAD and 20 pg/ml of the A-protomer of IAP and then submitted to polyacrylamide gel electrophoresis as described under “Experimental Procedures.” The ADP-ribose moiety of [ B - ~ ~ P ] N A D incorporated into the M, = 41,000 protein (pmol/mg of membrane proteins) was calculated from the 32P content of the protein and plotted against the concentration of Ni present during reconstitution. In B, the reconstituted membranes were assayed for GTPase in the presence or absence of 1 p~ Met-Leu-Phe as described under “Ex- perimental Procedures.” The assay was conducted in the presence of 0.5 pM [T-~’P]GTP for 2 min. The increment in GTPase by m e t - Leu-Phe was shown on the ordinate. The basal GTPase activity (pmol/mg/ml) of membranes reconstituted with 0, 2.4, 7.3, or 14.6 pg/ml of Ni was 37 f 1, 51 f 2, 82 + 1, or 142 f 2 for IAP-treated membranes and 49 f 2, 69 + 2, 91 f 2, or 140 f 7 for nontreated membranes, respectively. Results are the mean f S.E. of three obser- vations.
lowered the relative population of RH to RL in the absence, but did not in the presence, of Gpp(NH)p.
Reversal by Reconstitution with Ni of the ZAP-induced in- hibition of fMet-Leu-Phe-stimulated GTPase Activity and the Formyl-peptide Binding-Foregoing results suggest that the IAP substrate (i.e. a membrane protein to be ADP-ribosylated by IAP) is involved in the regulation of Net-Leu-Phe-stim- ulated GTPase activity and also in the formyl-peptide binding to its receptors. Since the IAP substrate is now known to be Ni in various cells (l), we next examined the direct action of Ni purified from rat brain on the GTPase activity and the formyl-peptide receptor binding.
6766 Uncoupling of Formyl-peptide Signal Transduction by IAP TABLE I1
Parameters for fMet-Leu-PHIPhe binding to IAP (A-protomerj-treated or nontreated neutrophil membranes in the presence or absence of Gpp(NH)p
Data were from Fig. 4. Values were derived from nonlinear least squares fit to a two-site binding equation as described under “Experimental Procedures.” RH or RL represents a high- or low-affinity receptor, respectively. See “Experimental Procedures” for other abbreviations. Results are the mean f S.E. of four separate experiments.
Membrane GPP(NH)P RH RL treatment addltlon
Total
& ( I ) &(a) B-(2) number receptor
nM Prnllmg nM pmollmg -
None 8.15 f 0.91 3.55 f 0.38 1624 f 729 1.81 k 0.51 5.36 k 0.77 5.24 f 0.98 1.32 f 0.11“ 1619 f 270 4.08 k 0.63* 5.40 & 0.71 +
A-protomer - 7.23 f 1.84 2.16 f 0.31‘ 2056 f 493 3.99 f 0.73‘ 6.10 f 0.82
5.98 f 1.36 1.21 f 0.25b 1393 f 382 3.67 +. 0.46 4.88 f 0.57 + a Effect of Gpp(NH)p addition is significant; p < 0.01. ‘Effect of Gpp(NH)p addition is significant; p < 0.05. Effect of A-protomer treatment is significant; p < 0.05.
The content of the IAP substrate estimated from IAP (A- protomer)-catalyzed ADP-ribosylation of a M, = 41,000 mem- brane protein was about 70% less in the membranes from neutrophils previously treated with IAP than in the mem- branes from nontreated cells (Fig. 5A), reflecting the prior ADP-ribosylation of the protein during exposure of cells to IAP as shown in the previous publication (2). These mem- branes were reconstituted with the increasing concentration of Ni, an oligomeric peptide consisting of an a (Mr = 41,000), a (Mr = 35,000), and a y (Mr = 10,000) subunit (Fig. 5 ) . The amount of Ni inserted into membranes was estimated based on the incorporation of 32P from [cY-~’P]NAD to the M, = 41,000 membrane protein, the a subunit of Ni, as induced by incubation of the membranes with the A-protomer of IAP. The incorporation was increased progressively in both mem- brane preparations, as the amount of Ni used for the recon- stitution was increased (Fig. 5A). The 32P incorporation was slightly lower at all concentrations of Ni employed in the membranes from IAP-treated cells than in membranes from nontreated cells, probably reflecting the lower content of the IAP substrate in the former before reconstitution (Fig. 5A).
These Ni-reconstituted membranes were assayed for GTPase activity (Fig. 5B). Formyl-Met-Leu-Phe-stimulated GTPase activity of control cell membranes did not show a significant increase when Ni was present at concentrations below 7.4 pg/ml, while the enzyme activity of membranes prepared from IAP-treated neutrophils was significantly in- creased by the addition of the lowest concentration (2.4 pg/ ml) of Ni. The GTPase activity of the treated cell membranes further increased as the concentration of Ni was increased until it reached to the activity of the nonreconstituted mem- branes of control cells at the Ni concentration of 14.6 pg/ml, though the GTPase activity of the control cell membranes was also further increased upon addition of this highest con- centration of Ni.
Reconstitution of Ni into the membranes prepared from IAP-treated neutrophils was also effective in increasing the specific binding of fMet-Le~-[~H]Phe to these membranes (Table 111). This effect was observed when the binding assay was achieved with a low concentration (50 nM), but was not observed with a high (2 p ~ ) concentration, reflecting the increase in binding affinity rather than binding capacity (see Fig. 4). The effect was not observed in the presence of Gpp(NH)p either, even with a low concentration of the labeled peptide (Table 111). Thus, the action of IAP to lower the affinity of met-Leu-Phe binding to membrane receptors in the absence of Gpp(NH)p was reversed by reconstitution of Ni to the membrane, confirming the notion that IAP inter-
TABLE I11 Effect of reconstitution of N, into IAP-treated neutrophil membranes
on fMet-Leu-PHIPhe binding Membranes from IAP-treated neutrophils were reconstituted with
(Nil or without (control) 14.6 pg/ml of Ni purified from rat brain as described under “Experimental Procedures.” The fhIet-Le~-[~H]Phe binding experiments were conducted in the presence of 50 nM or 2 pM ligand with or without (none) 100 p~ Gpp(NH)p. See “Experi- mental Procedures” for details. Results are mean f S.E. of three observations.
None GPP(NH)P
pmollmg 50 nM ligand
Control 1.07 f 0.08 0.82 -+ 0.14 Ni 3.23 f 0.17” 1.09 f 0.12
Control 5.65 f 0.57 6.44 f 0.72 2 p~ ligand
Ni 5.74 f 0.93 6.11 0.46 a Difference from control is significant; p < 0.01.
acted with Ni in the membranes. Concentration-dependent Inhibition by IAP of fMet-Leu-
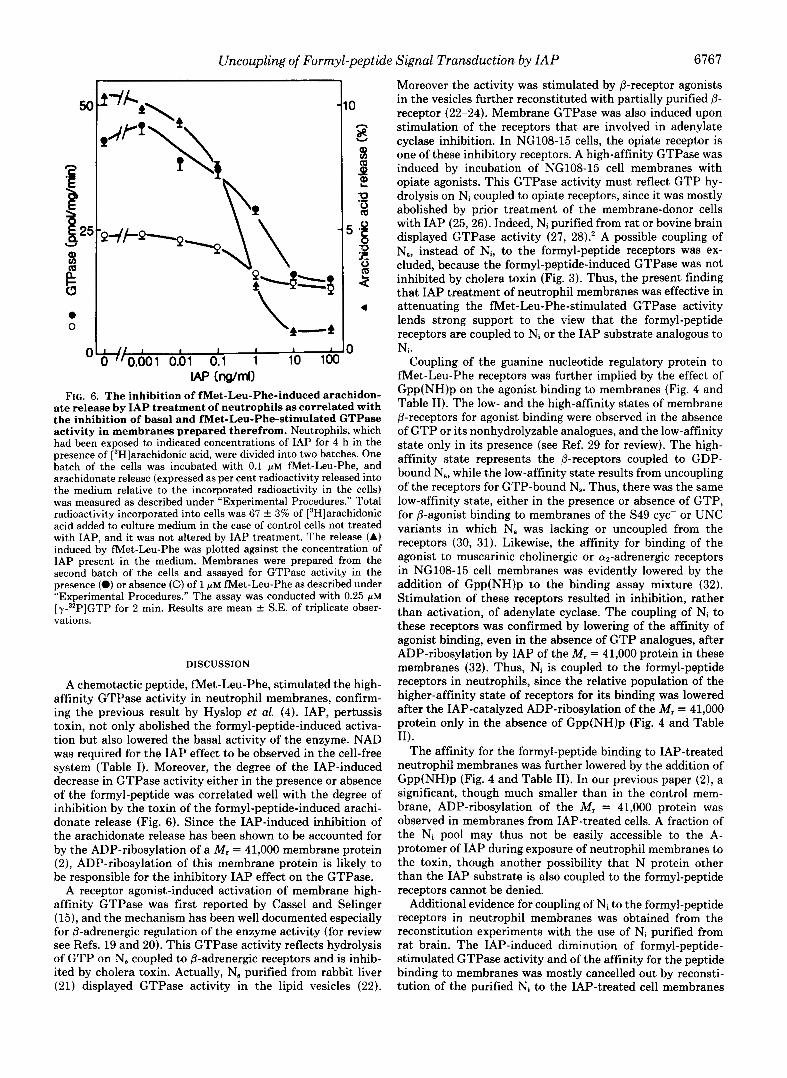
Phe-induced Arachidonate Release from Neutrophils and Its Correlation with the Inhibition of Basal and Formyl-peptide- stimulated GTPase Activity of Membranes Prepared There- from-The basal and Met-Leu-Phe-stimulated GTPase ac- tivity of membranes prepared from 4-h cultured neutrophils were inhibited progressively as the concentration of IAP was increased from 0.01 to 100 ng/ml in the culture medium (Fig. 6). The concentration of IAP required for the half-maximal inhibition was 0.5 ng/ml either for the basal or for the formyl- peptide-stimulated GTPase activity. We showed in the pre- vious paper (2) that the formyl-peptide-induced arachidonate release from [3H]arachidonate-labeled cells was also inhibited by exposure of cells to the increasing concentration of IAP. The arachidonate releasing response to met-Leu-Phe de- picted in Fig. 6 was achieved by using the same batch of neutrophil from which membranes were prepared for the assay of GTPase activity. The half-maximal concentration of IAP to inhibit the arachidonate releasing response was roughly 0.4 ng/ml, the value comparable to its half-maximal concentra- tion to inhibit the basal and the formyl-peptide-stimulated GTPase activity of membranes prepared therefrom (Fig. 6). Since our previous report (2) also showed that IAP inhibition of arachidonate releasing response was correlated well with the IAP-catalyzed ADP-ribosylation of the M, = 41,000 pro- tein, the results in Fig. 6 suggest that IAP inhibition of membrane GTPase activity is also correlated with ADP- ribosylation of the membrane protein.
Uncoupling of Formyl-peptide Signal Transduction by IAP 6767
FIG. 6. The inhibition of met-Leu-Phe-induced arachidon- ate release by IAP treatment of neutrophils as correlated with the inhibition of basal and met-Leu-Phe-stimulated GTPase activity in membranes prepared therefrom. Neutrophils, which had been exposed to indicated concentrations of IAP for 4 h in the presence of [3H]arachidonic acid, were divided into two hatches. One batch of the cells was incubated with 0.1 PM Net-Leu-Phe, and arachidonate release (expressed as per cent radioactivity released into the medium relative to the incorporated radioactivity in the cells) was measured as described under “Experimental Procedures.” Total radioactivity incorporated into cells was 67 & 3% of [3H]arachidonic acid added to culture medium in the case of control cells not treated with IAP, and it was not altered by IAP treatment. The release (A) induced by Net-Leu-Phe was plotted against the concentration of IAP present in the medium. Membranes were prepared from the second batch of the cells and assayed for GTPase activity in the presence (0) or absence (0) of 1 PM Met-Leu-Phe as described under “Experimental Procedures.” The assay was conducted with 0.25 @M [y-:P]GTP for 2 min. Results are mean & S.E. of triplicate obser- vations.
DISCUSSION
A chemotactic peptide, fMet-Leu-Phe, stimulated the high- affinity GTPase activity in neutrophil membranes, confirm- ing the previous result by Hyslop et al. (4). IAP, pertussis toxin, not only abolished the formyl-peptide-induced activa- tion but also lowered the basal activity of the enzyme. NAD was required for the IAP effect to be observed in the cell-free system (Table I). Moreover, the degree of the IAP-induced decrease in GTPase activity either in the presence or absence of the formyl-peptide was correlated well with the degree of inhibition by the toxin of the formyl-peptide-induced arachi- donate release (Fig. 6). Since the IAP-induced inhibition of the arachidonate release has been shown to be accounted for by the ADP-ribosylation of a M, = 41,000 membrane protein (2), ADP-ribosylation of this membrane protein is likely to be responsible for the inhibitory IAP effect on the GTPase.
A receptor agonist-induced activation of membrane high- affinity GTPase was first reported by Cassel and Selinger (15), and the mechanism has been well documented especially for @-adrenergic regulation of the enzyme activity (for review see Refs. 19 and 20). This GTPase activity reflects hydrolysis of GTP on N. coupled to @-adrenergic receptors and is inhib- ited by cholera toxin. Actually, N, purified from rabbit liver (21) displayed GTPase activity in the lipid vesicles (22).
Moreover the activity was stimulated by preceptor agonists in the vesicles further reconstituted with partially purified /3- receptor (22-24). Membrane GTPase was also induced upon stimulation of the receptors that are involved in adenylate cyclase inhibition. In NG108-15 cells, the opiate receptor is one of these inhibitory receptors. A high-affinity GTPase was induced by incubation of NG108-15 cell membranes with opiate agonists. This GTPase activity must reflect GTP hy- drolysis on Ni coupled to opiate receptors, since it was mostly abolished by prior treatment of the membrane-donor cells with IAP (25,26). Indeed, Ni purified from rat or bovine brain displayed GTPase activity (27, 28).* A possible coupling of N., instead of Ni, to the formyl-peptide receptors was ex- cluded, because the formyl-peptide-induced GTPase was not inhibited by cholera toxin (Fig. 3). Thus, the present finding that IAP treatment of neutrophil membranes was effective in attenuating the Met-Leu-Phe-stimulated GTPase activity lends strong support to the view that the formyl-peptide receptors are coupled to Ni or the IAP substrate analogous to Ni.
Coupling of the guanine nucleotide regulatory protein to fMet-Leu-Phe receptors was further implied by the effect of Gpp(NH)p on the agonist binding to membranes (Fig. 4 and Table 11). The low- and the high-affinity states of membrane @-receptors for agonist binding were observed in the absence of GTP or its nonhydrolyzable analogues, and the low-affinity state only in its presence (see Ref. 29 for review). The high- affinity state represents the P-receptors coupled to GDP- bound N,, while the low-affinity state results from uncoupling of the receptors for GTP-bound N,. Thus, there was the same low-affinity state, either in the presence or absence of GTP, for @-agonist binding to membranes of the S49 cyc- or UNC variants in which N, was lacking or uncoupled from the receptors (30, 31). Likewise, the affinity for binding of the agonist to muscarinic cholinergic or cY,-adrenergic receptors in NG108-15 cell membranes was evidently lowered by the addition of Gpp(NH)p to the binding assay mixture (32). Stimulation of these receptors resulted in inhibition, rather than activation, of adenylate cyclase. The coupling of Ni to these receptors was confirmed by lowering of the affinity of agonist binding, even in the absence of GTP analogues, after ADP-ribosylation by IAP of the M , = 41,000 protein in these membranes (32). Thus, Ni is coupled to the formyl-peptide receptors in neutrophils, since the relative population of the higher-affinity state of receptors for its binding was lowered after the IAP-catalyzed ADP-ribosylation of the M , = 41,000 protein only in the absence of Gpp(NH)p (Fig. 4 and Table 11).
The affinity for the formyl-peptide binding to IAP-treated neutrophil membranes was further lowered by the addition of Gpp(NH)p (Fig. 4 and Table 11). In our previous paper (2), a significant, though much smaller than in the control mem- brane, ADP-ribosylation of the M, = 41,000 protein was observed in membranes from IAP-treated cells. A fraction of the Ni pool may thus not be easily accessible to the A- protomer of IAP during exposure of neutrophil membranes to the toxin, though another possibility that N protein other than the IAP substrate is also coupled to the formyl-peptide receptors cannot be denied.
Additional evidence for coupling of Ni to the formyl-peptide receptors in neutrophil membranes was obtained from the reconstitution experiments with the use of Ni purified from rat brain. The IAP-induced diminution of formyl-peptide- stimulated GTPase activity and of the affinity for the peptide binding to membranes was mostly cancelled out by reconsti- tution of the purified Ni to the IAP-treated cell membranes

6768 Uncoupling of Formyl-peptide Signal Transduction by IAP
(Fig. 5 and Table 111). The endogeneous IAP substrate was equivalent to 1.5 pmol of the ADP-ribose transfer/mg of protein (Fig. 5 ) . About 10-fold as much Ni was needed for the formyl-peptide-induced GTPase to be recovered to the origi- nal value, probably because of inefficient insertion of added Ni into the membrane under the present conditions. The ADP-ribosylated protein increased similarly, upon addition of Ni, in membranes from control and IAP-treated cells; the same amount of Ni was inserted into both membranes.
Thus, the experimental results in the present communica- tion lend support to the notion that Ni is coupled to m e t - Leu-Phe receptors in guinea pig neutrophils. The function of Ni is to mediate receptor-coupled adenylate cyclase inhibition in many other cell types. In the case of guinea pig neutrophils, however, no evidence has been obtained for fMet-Leu-Phe- induced decreases in cellular cAMP (see Ref. 2). Instead, the formyl peptide slightly increased the nucleotide content (2). The mechanism by which the formyl-peptide increases cellu- lar cAMP is not well understood. Such a small, but significant, increase in cAMP content was observed when human neutro- hils were exposed to A23187, a Ca2+ ionophore (33). Further- more, the formyl-peptide failed to activate adenylate cyclase in the neutrophil cell-free system, while &adrenergic agonists or prostaglandins were effective in stimulating the enzyme activity under the same conditions (34). Prostaglandin E*- induced accumulation of cAMP in neutrophils was not inter- fered with by the simultaneous addition of fMet-Leu-Phe, making it unlikely that the chemotactic peptide receptor is coupled to adenylate cyclase in an inhibitory fa~hion .~ Thus , the increase in the cellular cAMP content induced by fMet- Leu-Phe might be secondary to the increase in the intracel- lular concentration of Ca2+, though the mechanism whereby Caz+ increases cAMP content remains unclear.
We previously showed that the M, = 41,000 protein suscep- tible to IAP is involved in Met-Leu-Phe-induced arachidon- ate release (probably reflecting phospholipase Az activation) (2) and also polyphosphoinositide breakdown by the phospho- lipase C type e n ~ y m e . ~ Similar participation of the IAP sub- strate in the arachidonate release or polyphosphoinositide breakdown in response to various receptor agonists was ob- served in mast cells (35, 36) and 3T3 fibroblast (37). The inhibition by IAP of the cellular arachidonate-releasing re- sponse to Wet-Leu-Phe was correlated well with the inhibi- tion of GTPase activity of membranes prepared therefrom (Fig. 6), indicating that the M , = 41,000 protein involved in arachidonate-releasing response or polyphosphoinositide breakdown is an entity identical with the IAP substrate directly coupled to the formyl-peptide receptors. It is thus reasonable to assume that the IAP substrate, probably Ni, exerts its effect, in a guanine nucleotide-dependent manner, on phospholipase Az or C, as does N, or Ni regulate adenylate cyclase activity. Further studies are now in progress to clarify the more detailed mechanism by which the IAP substrate regulates these phospholipase activities.
In fact, met-Leu-Phe enhanced prostaglandin E,-induced cAMP accumulation strikingly in neutrophils; cAMP content (fm01/104 cells) in the absence or presence of 0.1 PM fMet-Leu-Phe was 14.8 f 0.4 or 25.3 + 1.4 without prostaglandin E, and 120.9 f 3.4 or 300.5 + 6.3 with 10 FM prostaglandin E2, respectively (number of observations was three). The mechanism for this peculiar effect is now under study.
H. Ohta, F. Okajima, and M. Ui, manuscript in preparation.
REFERENCES 1. Ui, M. (1984) Trends Pharmacol. Sci. 5 , 277-279 2. Okajima, F., and Ui, M. (1984) J. Biol. Chem. 259, 13863-13871 3. Koo, C., Lef'kowitz, R. J., and Snyderman, R. (1983) J. Clin.
Invest. 7 2 , 748-753 4. Hyslop, P. A., Oades, Z. G., Jesaitis, A. J., Painter, R. G.,
Cochrane, C. G., and Sklar, L. A. (1984) FEBS Lett. 166,165- 169
5. Yajima, M., Hosoda, K., Kanbayashi, Y., Nakamura, T., Nogi- mori, K., Nakase, Y., and Ui, M. (1978) J. Biochem. (Tokyo)
6. Tamura, M., Nogimori, K., Murai, S., Yajima, M., Ito, K., Katada,
7. Washida, N., Sagawa, A., Tamoto, K., and Koyama, J. (1980)
83,295-303
T., Ui, M., and Ishii, S. (1982) Biochemistry 21,5516-5522
Biochim. Biophys. Acta 6 3 1 , 371-379 8. Honma, M., Satoh, T., Takezawa, J., and Ui, M. (1977) Biochem.
Med. 18, 257-273 9. Katada, T., Tamura, M., and Ui, M. (1983) Arch. Biochem.
10. Enomoto, K., and Gill, D. M. (1980) J. Biol. Chem. 255 , 1252-
11. Bokoch, G. M., Katada, T., Northup, J. K., Ui, M., and Gilman,
12. Laemmli, U. K. (1970) Nature (Lond.) 227 , 670-685 13. Katada, T., and Ui, M. (1982) Proc. Natl. Acad. Sci. U. S. A. 7 9 ,
14. Katada, T., and Ui, M. (1982) J. Biol. Chem. 257, 7210-7216 15. Cassel, D., and Selinger, Z. (1976) Biochim. Biophys. Acta 452,
16. Katada, T., Amano, T., and Ui, M. (1982) J. Biol. Chem. 257,
17. Cassel, D., and Selinger, Z. (1977) Proc. Natl. Acad. Sci. U. S. A.
18. Lester, H. A., Steer, M. L., and Levitzki, A. (1982) Proc. Natl.
19. Ross, E. M., and Gilman, A. G. (1980) Annu. Rev. Biochem. 49 ,
20. Limbird, L. E. (1981) Biochem. J. 195 , 1-13 21. NorthuD. J. K.. Smipel. M. D.. and Gilman. A. G. (1982) J. Biol.
Biophys. 224, 290-298
1258
A. G. (1984) J. Biol. Chem. 259, 3560-3567
3129-3133
538-551
3739-3746
74,3307-3311
Acad. Sci. U. S. A. 7 9 , 719-723
533-564
Chem.'257,1141&li423 . ,
22. Brandt. D. R.. Asano. T., Pedersen. S. E.. and Ross. E. M. (1983) Biochemist& 22,4357-4362 '
23. Cerione, R. A., Sibley, D. R., Codina, J., Benovic, J. L., Winslow, J., Neer, E. J., Birnbaumer, L., Caron, M. G., and Lef'kowitz, R. J. (1984) J. Biol. Chem. 259 , 9979-9982
24. Cerione, R. A., Codina, J., Benovic, J. L., Lefkowitz, R. J., Birnbaumer, L., and Caron, M. G. (1984) Biochemistry 23, 4519-4525
25. Burns. D. L.. Hewlett. E. L.. Moss. J.. and Vauehan. M. (1983)
. ,
J. Bioi. Chem. 258,'1435-1438 I , - , . I
26. Ui, M., Katada. T., Murayama, T.,Kurose, H., Yaiima. M., Ta- mura, M., Nakamura, TI, and Nogimori, K. (1984) in Advances in Cyclic Nucleotide and Protein Phosphorylation Research
eds) Vol. 17, pp. 145-151, Raven Press, New York (Greengard, P., Robison, G. A., Paoletti, R., and Nicosia, S.,
27. Sternweis, P. C., and Robishaw, J. D. (1984) J. Biol. Chem. 2 5 9 ,
28. Neer, E. J., Lok, J. M., and Wolf, L. G. (1984) J. Biol. Chem.
29. Stiles, G. L., Caron, M. G., and Lefkowitz, R. J. (1984) Physiol.
30. Haga, T., Ross, E. M., Anderson, H. J., and Gilman, A. G. (1977)
31. Ross, E. M., Maguire, M. E., Sturgill, T. W., Biltonen, R. L., and
32. Kurose, H., Katada, T., Amano, T., and Ui, M. (1983) J. Biol.
33. Smolen, J. E., and Weissmann, G. (1981) Biochim. Biophys. Acta
34. Lad, P. M., Glovsky, M. M., Richards, J. H., Learn, D. B., Reisinger, D. M., and Smiley, P. A. (1984) Mol. Zmmunol. 21 ,
13806-13813
259,14222-14229
Reu. 64,661-743
Proc. Natl. Acad. Sci. U. S. A. 7 4 , 2016-2020
Gilman, A. G. (1977) J. Biol. Chem. 252, 5761-5775
Chem. 258,4870-4875
672,197-206
627-639 35. Nakamura, T., and Ui, M. (1984) FEBS Lett. 173,414-418 36. Nakamura, T., and Ui, M. (1985) J. Biol. Chem. 260,3584-3593 37. Murayama, T., and Ui, M. (1985) J. Biol. Chem., in press





























