Embed Size (px)
Citation preview

Behavioral Neuroscience1984, Vol. 98, No 2,171-188
Copyright 1984 by theAmerican Psychological Association, Inc
Physiological Plasticity of Single Neuronsin Auditory Cortex of the Cat During Acquisition
of the Pupillary Conditioned Response:I. Primary Field (AI)
Norman M. Weinberger, William Hopkins, and David M. DiamondCenter for the Neurobiology of Learning and Memory
and Department of PsychobiologyUniversity of California, Irvine
The effects of conditioning on the discharges of single neurons in primaryauditory cortex (AI) were determined during acquisition of the pupillaryconditioned response in chronically prepared cats. Acoustic stimuli (1-s whitenoise or tone) were presented with electrodermal stimulation unpaired duringa sensitization control phase followed by pairing during a subsequent condi-tioning phase. Stimulus constancy at the periphery was ensured by the use ofneuromuscular blockade. Discharge plasticity developed rapidly for bothevoked and background activity, the former attaining criterion faster than thelatter. The pupillary dilation conditioned response was acquired at the samerate as were changes in evoked activity (i.e., 10-15 trials) and faster thanbackground activity (i.e., 20-25 trials). Increases in background activity werecorrelated with increasing level of tonic arousal, as indexed by pretrial size ofthe pupil.
Behavioral adaptation requires accurateinformation about the environment. Thatsensory systems provide such informationhas never been in serious question. How-ever, the mechanisms involved have not yetbeen elucidated fully. This problem appearsto be further complicated by the fact thatelectrophysiological responses to environ-mental stimuli within sensory systems aremodified by learning. Numerous studies ofclassical and instrumental conditioning inanimals have demonstrated that responsesto conditioned and discriminative stimuliin sensory systems are altered systemati-cally by associative processes (for reviews,
This research was funded by a grant from theMonsanto Company, and by National Institute ofNeurological and Communicative Disorders andStroke Grant NS16108 and National Science Foun-dation Grant BNS76-81924 to N. M. Weinberger.
The authors gratefully acknowledge Cathy Bennettand Thomas McKenna for helpful discussion duringthe preparation of the manuscript, Herman Birch forwriting the computer programs, and Lisa Weinbergerfor secretarial assistance.
Requests for reprints should be sent to Norman M.Weinberger, Center for the Neurobiology of Learningand Memory, University of California, Irvine, Califor-nia 92717.
see John, 1961; Sokolov, 1977; Thompson,Patterson, & Teyler, 1972). Such responseplasticity is particularly evident in sensorycortex; it has been documented most exten-sively in auditory cortex (e.g., Buchwald,Halas, & Schramm, 1966; Cassady, Cole,Thompson, & Weinberger, 1973; Galam-bos, Sheatz, & Vernier, 1955; Oleson, Ashe,& Weinberger, 1975) and has been reportedas well in olfactory (Freeman, 1980), so-matosensory (e.g., Voronin, Gerstein, Ku-dryashov, & Ioffe, 1975), and visual (e.g.,Shinkman, Bruce, & Pfingst, 1974) cor-tices. Thus, sensory responses are affectedby two types of variables: (a) the physicalparameters of stimuli and (b) the meaningor cue value of stimuli. This situation sug-gests a paradox because the requirementfor veridical responses to the physical pa-rameters of stimuli appears to be compro-mised by the effects of stimulus meaning.
In recent years, this issue has been in-vestigated most thoroughly within the au-ditory system, and there now appears to bea partial resolution of the paradox. Ana-tomical and physiological studies have re-vealed that the auditory system containsboth lemniscal and non-lemniscal or "lem-
171

172 N. WEINBERGER, W. HOPKINS, AND D. DIAMOND
niscal adjunct" ascending pathways (Gray-biel, 1972; Herkenham, 1980; Ryugo & Kil-lackey, 1974; J. Winer, Diamond, & Ra-czkowski, 1977). At the level of the tha-lamic auditory system, these subsystemsengage different subdivisions of the medialgeniculate body (Morest, 1964, 1965). Thelemniscal line projects to the ventral medialgeniculate nucleus (MGv), the neurons ofwhich are tonotopically organized, withnarrow tuning functions (Aitkin & Webs-ter, 1972), whereas the nonlemniscal lineprojects to the magnocellular medial genic-ulate nucleus (MGm), the neurons of whichare not tonotopically organized and havevery broad tuning functions (Aitkin, 1973).Analysis of the physical parameters ofsound and the meaning of sound is alsocompartmentalized at this level of the au-ditory system. Neurons in the lemniscalMGv respond to the physical parameters ofthe acoustic environment, but these re-sponses are unaffected by learning. In con-trast, neurons in the nonlemniscal MGmare not particularly sensitive to changes inthe physical parameters of sound, but theydevelop discharge plasticity rapidly duringlearning. These findings have been ob-tained in cat during classical defensive con-ditioning (Ryugo & Weinberger, 1976,1978), rabbit during instrumental avoid-ance conditioning (Gabriel, Miller, & Sal-twick, 1976), and rat during hybrid classi-cal-instrumental appetitive conditioning(Birt, Nienhuis, & Olds, 1979; Birt & Olds,1981).
At the level of auditory cortex, the situ-ation has not yet been clarified, but it isprobably more complex than at the thala-mus. Auditory cortex consists of severalfields, which have either lemniscal or non-lemniscal characteristics. The primary au-ditory cortex (AI) and the anterior (AAF)and posterior (P) fields are tonotopicallyorganized; in contrast, the secondary audi-tory cortex (AH) and insular (I) and tem-poral (T) fields are not so organized andthus seem to be nonlemniscal in nature(e.g., Reale & Imig, 1980). Almost all pre-vious studies of learning in the auditorycortex have investigated AI, and, as pointedout above, there is abundant documenta-tion that evoked potentials and multiple-
unit activity in this cortical field are mod-ified by associative processes. Insofar asboth the ventral and the magnocellular me-dial geniculate nuclei project upon primaryauditory cortex, we have suggested that theformer, being nonplastic, is not a source ofassociative effects on AI whereas the lattermight be involved in learning-induced cor-tical discharge plasticity (Weinberger, inpress). Additional analysis of this and manyrelated issues requires investigation of theeffects of learning on the discharge prop-erties of single neurons in auditory cortex,because of the inherent limitations ofevoked potentials and multiple-unit data.Single-unit studies during the acquisitionof behavioral conditioned responses aretechnically difficult, requiring that ade-quate recordings be obtained continuouslyduring a prolonged period of training, andthey yield data from only one cell per train-ing session. Nonetheless, such studies areimportant because they provide a link be-tween the fields of single-unit auditoryphysiology and learning and they can revealthe changes underlying neurophysiologicalplasticity as indexed by evoked potentialsand particularly by multiple-unit record-ings (Kraus & Disterhoft, 1982).
The present experiment is concernedwith the discharges of single cells in pri-mary auditory cortex (AI) during the ac-quisition of a behavioral conditioned re-sponse. A companion report is concernedwith the effects of classical conditioning onthe secondary auditory cortical field (All)which is prototypical of nonlemnical audi-tory cortex (D. Diamond & Weinberger,1984). Such comparative information maybe important for understanding the func-tional role of multiple sensory corticalfields (I. Diamond, 1979; Merzenich &Kass, 1980) as well as bearing more directlyon issues regarding the neuronal bases oflearning. Further, it is hoped that the dataso obtained will promote the rapproche-ment of the fields of sensory neurobiologyand learning, as both areas 'are criticallyconcerned with how the brain acquires in-formation about the environment.
A preliminary report of some of theseresults has been presented (Hopkins &Weinberger, 1980).

PLASTICITY OF SINGLE AI NEURONS DURING LEARNING 173
Method
Surgical Preparation
The subjects were 8 adult cats, 3.2-5.0 kg, in goodhealth. The animals were anesthetized with sodiumpentobarbital (Nembutal, 40 mg/kg, ip) and placed ina stereotaxic frame, with care taken to preserve theintegrity of the external auditory meatus and tym-panic membrane. The scalp was incised and reflected,and the calvarium was cleared of connective tissue. Apedestal containing metal fixtures was built with den-tal acrylic and secured to the skull with stainless steelscrews. The pedestal allowed for immobilization of thehead during subsequent training sessions without di-rect pressure on the animal. Body temperature wasmaintained by a thermostatically controlled warmwater pad during the surgery and recording sessions.Ophthalmic ointment (Terramycin) was applied toprevent corneal drying. Antibiotics (Panalog and De-lagon) were applied locally to exposed skin surfaces,and Bicillin (300,000 U im) was administered for 2-3days following the surgery. Training began after arecovery period of 1-2 weeks.
Experimental Design and Procedures
At the beginning of a training session, the animalsunderwent neuromuscular blockade induced by gal-lamine triethiodide (Flaxedil, 10 mg/kg, ip). The tra-chea was intubated with a pediatnc catheter, coatedwith a local anesthetic (Xylocaine), under laryngos-copic control, and the animal was artificially respiredwith a Harvard respirator. Following immobilization,the animal was positioned in a modified stereotaxicframe fitted with bars which attached to the pedestal.All procedures were carried out with the animals en-closed in an acoustically isolated chamber (IAC 1202).Neuromuscular blockade was maintained with intra-venous infusion of Flaxedil (20 mg/hr, iv). ExpiredCO2 levels were not monitored because we had foundin previous experiments that pupillary size and motil-ity in response to sensory stimulation are more sen-sitive indexes of the animal's condition. At the end ofa training session, the animals were recovered withthe assistance of Tensilon (0.6 ml im).
Acoustic stimulation was delivered to the ear con-tralateral to the recording electrode by a Beyer ear-phone which was attached to plastic tubing that fittedinto a mold of the external auditory meatus. The moldwas sealed with plasticine to provide a closed acousticdelivery system. Acoustic conditioned stimuli (CS)were either 1-3 kHz tones (n = 4) produced by aWavetek oscillator (70-85 dB) or white noise (n = 17)produced by a Grason-Stadler generator (bandwidth0.2-20 kHz, 80 dB) and were 1 s in duration. Stimulusintensities are expressed as decibels above a referenceof 20 jjN/m2 and were controlled by Hewlett-Packardattenuators. Sound intensity level was measured witha Bruel and Kjaer sound lever meter, condenser mi-crophone, and probe tube inserted into the externalauditory meatus through a sealable port in the sounddelivery tube. The unconditioned stimulus (US) was
electrodermal stimulation (EDS) consisting of a 375-ms train of 50-Hz pulses (5.0 ms) produced by a GrassS-44 constant-current stimulator via an optical isola-tion transformer. Stimulus intensity (2-9 mA) was setat the beginning of a session to produce a brief (2-5s) pupillary dilation. The US was delivered to thesubcutaneous tissue of the forepaw contralateral tothe recording site via a pair of fine wire electrodes.
Pupillary size was monitored by an infrared pupil-lometer (Cassady, Farley, Weinberger, & Kitzes, 1982)positioned in front of the one eye. Care was taken toavoid possible discomfort due to drying of the corneasby covering them with ophthalmic ointment. In somesessions, pupillary stability was enhanced by a lowlevel of illumination presented to the eye contralateralto the pupillometer, but there were no differences inbehavioral or neuronal results related to the use ofsuch illumination. The output of the pupillometer wasamplified by a dc amplifier and written out on a GrassModel 7 polygraph. The level of drift of the pupillo-metric recording system was negligible.
The skin area surrounding the pedestal was locallyanesthetized with Marcaine (Breon Labs, Inc.) andthen gently retracted to expose the lateral surface ofthe skull. A small burr hole was drilled and a slit wasmade in the dura. These procedures were not stressfulto the animal as indicated by the lack of pupil dilation.
Single-unit discharges were recorded with tungstenelectrodes (1-ftm diameter tips) insulated with Epox-ylite (2-4 Mfi impedance at 1 kHz). Electrodes wereattached to a Narashige stepping microdrive con-trolled from outside the acoustic chamber and ad-vanced through the dural slit until the discharges ofsingle neurons were encountered (see below). Dis-charges were amplified with a conventional differen-tial preamplifier (bandwidth 80 Hz-10 kHz) and ledto an active high-pass filter (0.4-6 kHz), the output ofwhich was displayed on a storage oscilloscope andrecorded on a direct channel of a Hewlett-Packard3964A tape recorder. A single unit was identified byvisual inspection of the spike waveform. Neuronaldata were considered acceptable if the record consistedof a clearly distinguishable unit whose waveform ex-hibited no notches or other signs of injury, with asignal-to-noise ratio of at least 3:1. Data reported hereare only for units meeting these criteria.
A training session consisted of two parts, a sensi-tization phase and a conditioning phase. During sen-sitization, 15 CSs and USs each were presented in anunpaired fashion, at pseudorandom intervals at anaverage density of two per minute with the restrictionthat the stimuli not occur within 10 s of each other.During conditioning, the CS and US were alwayspaired, the US being presented at CS offset. Stimulusdensity was maintained at two per minute, the averagemtertrial interval was 60 s (range, 30-90 s). Thesensitization phase served as a control for nonassocia-tive factors. This phase used unpaired rather thanrandomized presentation of the CS and US becauseconditioned inhibition does not develop during thepresentation of small numbers of unpaired stimuli(Furedy, 1971; Furedy, Poulos, & Schiffman, 1975).Backward pairing was not used because it can produceconditioned inhibition to the CS due to its cue valueas a "safety" signal of the termination of the US (e.g.,

174 N. WEINBERGER, W. HOPKINS, AND D. DIAMOND
Segundo, Galeano, Sommer-Smith, & Roig, 1961).Conditioning trials were initiated without break afterthe last sensitization trial and were continued for upto 60 trials or until acceptable recordings from a singlecell could not be obtained. Animals received only onetraining session on a single day, and at least 7 daysintervened between successive training sessions.
Data Analysis
The effects of training on pupillary behavior wereassessed as described previously (Ashe, Cassady, &Weinberger, 1976; Oleson et al., 1975;, Oleson, Vo-dodnick, & Weinberger, 1973; Oleson, Westenberg, &Weinberger, 1972; Ryugo & Weinberger, 1978).Briefly, the pupillometer write-out was measured im-mediately preceding the presentation of the CSthroughout training and preceding the US for sensi-tization trials on which EDS was given alone. Thepeak amplitude of dilations to these stimuli was mea-sured and the pretnal level was subtracted, whichyielded a difference score hereafter referred to as apupillary response. The scores and baseline levels weredetermined for every trial throughout training. Theywere normalized for comparison across animals byexpressing each value as a percentage change from theaverage of the last five-trial block of the sensitizationphase. The number of trials to criterion for pupillaryconditioning was defined as five consecutive trialsduring the conditioning phase all of which had re-sponses greater than the average of the last five trialsof the sensitization phase. The probability of thisoccurring by chance is .03 (Feller, 1968).
Neuronal discharges were analyzed with the assist-ance of an LSI/11 computer. Single-unit dischargeswere passed through a voltage detector or window thatproduced a single pulse for each discharge detected.The output of the trigger source was led to a pulseinput in the computer which recorded the occurrenceof spikes during 0.5-4 s immediately preceding a trialand during the trial. Spike counts were stored inconsecutive bins of 2 or 3 ms The average number ofcounts per bin was determined for every pretrial periodand every dunng-tnal period. The pretrial average wassubstracted from the during-tnal average for eachtrial; this yielded a difference score. The pretrial scoresare hereafter referred to as background activity, andthe difference scores as evoked activity. Each back-ground and each evoked score were normalized byexpressing it as a percentage change from the averagepretrial and evoked scores, respectively, for the lastfive-trial block of the sensitization period. The numberof trials to criterion of discharge plasticity was definedas five consecutive trials during the conditioning phaseall of which had greater or smaller values than theaverage of the last five trials during the sensitizationphase (p = .03). All data were evaluated by parametric(B Winer, 1971) or nonparametric (Siegel, 1956) sta-tistics.
Histology
Following a training session, a small electrolyticlesion was produced by passing anodal current through
the recording electrode. After the final session, theanimal was given an overdose of Nembutal and per-fused through the carotid arteries with 0.9% salinefollowed by 10% formalin; the brain was removed andstored in formalin. Frozen Berial sections (50 /urn) weretaken throughout auditory cortex and stained withcresyl violet. Recording sites were reconstructed ac-cording to the cytoarchitectural distinctions of thevarious subfields of auditory cortex described by Rose(1949) and Sousa-Pinto (1973).
Results
Pupillary Behavior and Conditioning
Pupillary dilation responses were re-corded for all experimental sessions. At thebeginning of sensitization, the acousticstimulus generally elicited brief, low-ampli-tude dilations. By the end of the sensitiza-tion phase, dilation responses were reducedin both amplitude and duration. The EDS(US) produced consistently large dilationresponses throughout the experimental ses-sion. During conditioning, acousticallyevoked dilations increased rapidly overtrials, as has been reported previously(Ashe et al., 1976; Oleson et al., 1972, 1973,1975; Ryugo & Weinberger, 1978; Weinber-ger, Oleson, & Haste, 1973). The averageconditioned dilations typically exceededthe largest acoustically elicited responsesof the preceding sensitization period duringTrials 6-10, attained asymptote by Trials21-25, and maintained high values there-after (Figure 1). The pupillary dilation con-ditioned response attained criterion in 16of 21 sessions. For these sessions, the meantrials to criterion was 11.81 (SD = 5.76,range, 6-27 trials).
Of the 8 animals, 7 received more thanone, and some as many as four, trainingsessions at weekly intervals. In order toassess the cumulative effect of earlier train-ing sessions, each animal was assigned asavings score based on a sequential com-parison of values for the trials-to-criterionmeasure from session to session. Thus, asubject would receive a net positive savingsscore if the number of all possible sessioncomparisons for which there was a reduc-tion in the trials-to-criterion measure weregreater than the number of comparisonsthat yielded an increase in this measure. Inthis manner it was determined that 4 of 7

PLASTICITY OF SINGLE AI NEURONS DURING LEARNING 175
1 2 3StNSITIZAIlON
5 6CONDITIONING
BLOCKS OF FIVE TRIALS
Figure 1. Group pupillary learning curve for the 17cases in which conditioned pupillary dilation re-sponses attained the criterion of learning. (Each pointis the mean percentage change in maximal pupillarydilation during presentation of the conditioned stim-ulus for blocks of five trials relative to the last five-trial block during the sensitization period In somecases, recording was terminated after the fourth blockof conditioning [Trial 20] due to deterioration of iso-lation of discharges from single units; the numbers ofsubjects for Blocks 5-10 were 12,11, 11, 10, 10, and 9,respectively. Vertical bars denote ±1 SE.)
animals had net positive savings scores, 1had a zero savings score, and 2 had negativesavings scores.
Location of Neurons ,
Data were obtained from 21 neurons withrecording sites verified histologically in pri-mary auditory cortex. The laminar sites ofrecording could be determined in 15 cases.Thirteen sites were in infragranular layersV and VI, and two sites were in layer IV.This distribution is insufficient for corre-lating anatomical lamina with neurophysi-ological data. The results presented hereshould be regarded as representing mainlythe deep lamina of primary auditory cortex.
tioned response. However, this index ofbehavioral learning is useful as a sign thatthe subject is "adequate," in order to permitunambiguous interpretation of "negative"neuronal findings. The failure of a neuronto develop a systematic change in its dis-charges during conditioning might be dueto its presumptive membership in a groupof "nonplastic" neurons, at least nonplasticfor the circumstances of a given experi-ment. Although this is the usual interpre-tation of negative findings, an alternativeexplanation is that the subject was "inade-quate," that is, a "substandard" prepara-tion. Thus, a neuron in question might havethe capacity to develop plasticity but beunable to express it because of the inade-quacy of the subject. In short, if the subjectis unable to acquire a conditioned responseand a neuron in that animal also fails toexpress discharge plasticity, no conclusionscan be drawn about the functional plastic-ity of the cell. Accordingly, the data fromsuch neurons should be set aside. In con-trast, a cell that does not develop dischargeplasticity in an animal that does develop abehavioral conditioned response can be saidto be functionally nonplastic for that situ-ation because the alternative explanationof an inadequate preparation can be re-jected.
As noted above, data were obtained from21 neurons in primary auditory cortex. Thedata from 2 neurons were eliminated fromthe total sample because the animals failedto acquire the pupillary dilation condi-tioned response and the neurons developedneither background nor evoked dischargeplasticity. In all other cases in which dis-charge plasticity did not develop, there wasindependent evidence of the adequacy ofthe preparation.
Neuronal Data—Interpretationof Negative Findings
The establishment of behavioral condi-tioned responses provides a frameworkwithin which to interpret neuronal activityduring conditioning procedures. As ex-plained previously (Weinberger, 1982a,1982b), we do not seek the neural circuitunderlying the pupillary dilation condi-
Evoked Activity
For the 19 cells in the analysis set, 14attained the criterion of discharge plastic-ity. However, we excluded any neuronsthat, although meeting this criterion, weresimply continuing a trend existent duringsensitization. This was accomplished bycomputing the slopes of functions for sen-sitization and conditioning, on a trial-by-

176 N. WEINBERGER, W. HOPKINS, AND D. DIAMOND
Table 1Effects of Conditioning on Evoked Discharges
Cell no.
IB5B6C7E8A8D
nMSD
Increase
Pupil
6
177
11
410.253.20
Evoked
2597
12117
611.836.77
Cell no.
3B5D5E7C10A10B
nMSD
Decrease
Pupil
166
116
2738
617.3312.80
Evoked
66
326
2710
614.5011.83
Cell no.
1C1D23E*6A16B7B*8B8C9A
nMSD
No change
Pupil Evoked
12 —8 —
(-) (-)18 —_—(-) (-)16 —7 —
10 —
6 711.83 —4.40 —
* Deleted from analysis because neither pupillary learning nor neuronal plasticity developed.
trial basis. Cells that had the same slopeduring conditioning as during sensitizationwere considered not to be plastic. Two neu-rons failed the slope test and are hereafterclassed as nonplastic with respect to evokedactivity. Thus, 12 cells (63%) were classi-fied as plastic.
Changes in evoked discharges developedrapidly and were evident during the firstblock of 5 trials. Statistical criterion wasattained on the average in 13.17 trials. Sixneurons developed increased responses (M= 11.83), and six developed decreased re-sponses (M = 14.50; Table 1). Functionsfor evoked activity are presented in Figure2. These functions are significantly differ-ent from each other (Mann-Whitney Utests: increases vs. decreases, p < .001; in-creases vs. no change, p < .001; decreasesvs. no change, p < .001). Examples of in-creases in evoked discharges are given inFigures 3B and 4 (cell 18D) and decreasesin Figures 3A and 4 (cell 6C).
Kraus and Disterhoft (1982) reportedthat some portions of the evoked dischargeof neurons in auditory association cortex ofthe rabbit are affected differently than oth-ers, during conditioning of the nictitatingmembrane response. Therefore, we ana-lyzed separately various portions of evokedactivity, in particular discharges having alatency less than 50 ms and longer latency
250 -
a. -150
-200
-250
•300
-350 t
•—•INCREASE n-6o- -o DECREASE n= 7A — A NO CHANGE n=7
1 2 3 1 2 3 4 5 6 7 8 9 10SENSITIZATION CONDITIONING
BLOCKS OF FIVE TRIALS
Figure 2 Group functions for changes in evokeddischarges, sorted according to whether the criterionof discharge plasticity attained was for increases ordecreases, or failure to meet the criterion ("nochange"). (Each point is the mean percentage changein evoked discharges for blocks of five trials relativeto the last five-trial block during the sensitizationperiod. Number of cells for each block during condi-tioning: increases, 6,6,6,5,4,3,3,3,3; decreases,7,7,7,7,7,7,7,6,6,4; no change, 7,7,7,7,6,5,5,4,4,4. Notethat the development of increased and decreasedevoked discharges is evident during the first block ofconditioning [Trials 1-5]. Vertical bars denote ±1SE.)

PLASTICITY OF SINGLE AI NEURONS DURING LEARNING 177
A SENSITIZATION
48-
36-1-3E TR 11-15
1000
1000
1000
1000
msec
B SENSITIZATION12-
9-
6-
3 -
0
1-610 ms bin
TR 6-15
• I I . I A I I . J I , ! ! . . , . . . . f ..l.r0 500 1000
12-
9-
6-
3-
0
CONDITIONINGTR 1-10
-fcj^ ..AI L I . .,.. .. . . . f l y , \',' ,500 1000
2-9-6-
3-
o-... In .A .a,,LTR 11-20
\ ' , 1,—i ' > " M ».
msec
Figure 3. Representative poststinuulus histograms for two neurons that developed discharge plastic-ity during conditioning. (In this, and Figures 4 and 7, the horizontal bars below the abscissa of eachgraph denote presentation of the acoustic conditioned stimulus. A: Cell I-3E—each histogram is thesum of discharges for 5 consecutive trials, as designated. Background discharges were unaffected byconditioning, whereas discharges evoked by the conditioned stimulus decreased. B: Cell 1-6—eachhistogram is the sum of 10 consecutive trials, as indicated. Background discharges decreased duringconditioning, whereas evoked discharges did not develop a statistically significant change relative tobackground activity.)
activity. We were unable to find consistenteffects of conditioning on various portionsof the evoked response.
The probability of developing dischargeplasticity was unrelated to whether the cellwas recorded during the first training ses-sion for an animal or a later session, x2(l>N = 18) = 0.078, p > .05.
Comparisons of the pupillary and neu-ronal evoked data indicated that there wasno significant difference in trials to crite-rion for the entire group (pupil M = 11.81;neuronal M = 13.17), £(24) = 0.89, p > .05.Within subjects, the pupil reached criterion(if at all) more rapidly than did changes inneural activity (if at all) in 9/19 cases, moreslowly in 8/19, and at the same time in 3/
19 cases. There was no significant differ-ence between the rates of change for thisgroup (pupil M = 12.25; evoked M = 15.75,Mann-Whitney U test, p < .05). For those12 cells attaining criterion, the pupil alsoattained criterion in 8 cases. There was nosignificant difference in trials to criterionfor this group (pupil = 12.25; evoked =15.75, Mann-Whitney Utest,p> .05). Also,within subjects, the pupil attained criterionmore rapidly than evoked activity in 4/12,more slowly in 7/12, and at the same timein 1/12 cases. Overall, then, there was nostatistically significant difference betweenthe rate of pupillary learning and the rateat which evoked discharge plasticity devel-oped.

SENSITIZATION12
</> 9UJ* 6-a.<" 3-
I-6B10 ms bin
TR 1-15
500 1000
SENSITIZftTION
TR 1-15
1000
SENSITIZATIONTR 1-15
1000
12-
<n 9-UJ* 6-O_<" 3-
oilI2i
<nUJ
5£ 6-
<" 3 -
0
CONDITIONING
JJUJJUL
LJILJLI
12
oo 9UJ
¥ 68>3
TR 1-15
500 1000
TR 16-30
500 1000
TR 31-45
500 1000
12-
in 9UJ
* 6CL
u> 3
CONDITIONING
TR 1-15
1000
TR 16-30
500 1000
CONDITIONING
TR 1-15
jOOO
1000
1000
Figure 4 Representative poststimulus histograms for three neurons that developed discharge plasticity during conditioning. (Each histogram is thesum of 15 consecutive trials, as indicated. Left: Cell I-8B—background discharges increased over the course of conditioning, whereas evoked activitywas not significantly altered. Middle: Cell I-6C—background activity was not altered by conditioned, whereas evoked discharges decreased. Right: CellI-18D—background discharges were not changed, whereas evoked activity increased during conditioning.)
i
pD
PLASTICITY OF SINGLE AI NEURONS DURING LEARNING 179
Background Activity
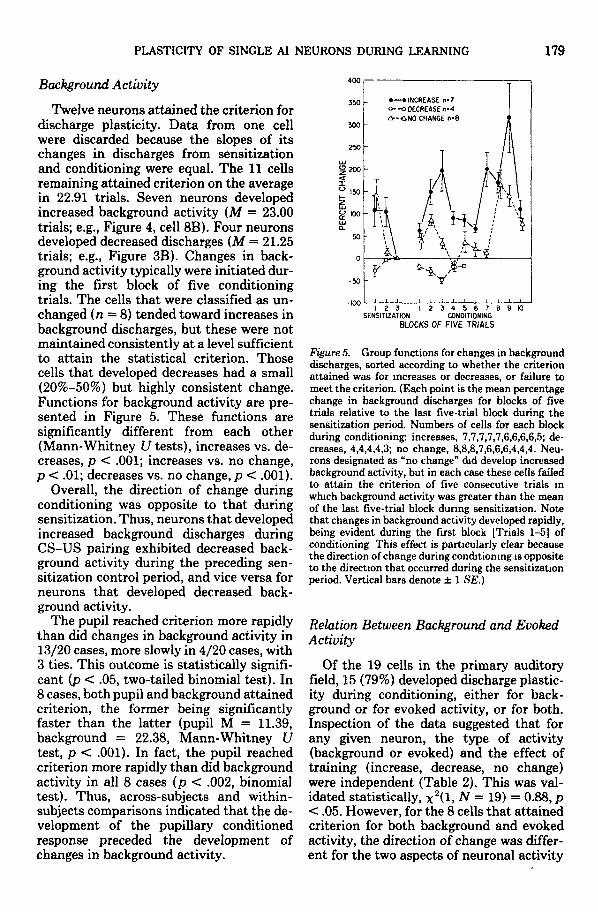
Twelve neurons attained the criterion fordischarge plasticity. Data from one cellwere discarded because the slopes of itschanges in discharges from sensitizationand conditioning were equal. The 11 cellsremaining attained criterion on the averagein 22.91 trials. Seven neurons developedincreased background activity (M = 23.00trials; e.g., Figure 4, cell 8B). Four neuronsdeveloped decreased discharges (M = 21.25trials; e.g., Figure 3B). Changes in back-ground activity typically were initiated dur-ing the first block of five conditioningtrials. The cells that were classified as un-changed (n = 8) tended toward increases inbackground discharges, but these were notmaintained consistently at a level sufficientto attain the statistical criterion. Thosecells that developed decreases had a small(20%-50%) but highly consistent change.Functions for background activity are pre-sented in Figure 5. These functions aresignificantly different from each other(Mann-Whitney U tests), increases vs. de-creases, p < .001; increases vs. no change,p < .01; decreases vs. no change, p < .001).
Overall, the direction of change duringconditioning was opposite to that duringsensitization. Thus, neurons that developedincreased background discharges duringCS-US pairing exhibited decreased back-ground activity during the preceding sen-sitization control period, and vice versa forneurons that developed decreased back-ground activity.
The pupil reached criterion more rapidlythan did changes in background activity in13/20 cases, more slowly in 4/20 cases, with3 ties. This outcome is statistically signifi-cant (p < .05, two-tailed binomial test). In8 cases, both pupil and background attainedcriterion, the former being significantlyfaster than the latter (pupil M = 11.39,background = 22.38, Mann-Whitney Utest, p < .001). In fact, the pupil reachedcriterion more rapidly than did backgroundactivity in all 8 cases (p < .002, binomialtest). Thus, across-subjects and within-subjects comparisons indicated that the de-velopment of the pupillary conditionedresponse preceded the development ofchanges in background activity.
1 2 3 1 2 3 4 5 6 7 8 9 10SENSITIZATION CONDITIONING
BLOCKS OF FIVE TRIALS
Figure 5. Group functions for changes in backgrounddischarges, sorted according to whether the criterionattained was for increases or decreases, or failure tomeet the criterion. (Each point is the mean percentagechange in background discharges for blocks of fivetrials relative to the last five-trial block during thesensitization period. Numbers of cells for each blockduring conditioning: increases, 7,7,7,7,7,6,6,6,6,5; de-creases, 4,4,4,4,3; no change, 8,8,8,7,6,6,6,4,4,4. Neu-rons designated as "no change" did develop increasedbackground activity, but in each case these cells failedto attain the criterion of five consecutive trials inwhich background activity was greater than the meanof the last five-trial block during sensitization. Notethat changes in background activity developed rapidly,being evident during the first block [Trials 1-5] ofconditioning This effect is particularly clear becausethe direction of change during conditioning is oppositeto the direction that occurred during the sensitizationperiod. Vertical bars denote ± 1 SE.)
Relation Between Background and EvokedActivity
Of the 19 cells in the primary auditoryfield, 15 (79%) developed discharge plastic-ity during conditioning, either for back-ground or for evoked activity, or for both.Inspection of the data suggested that forany given neuron, the type of activity(background or evoked) and the effect oftraining (increase, decrease, no change)were independent (Table 2). This was val-idated statistically, x
2 ( l , N = 19) = 0.88, p< .05. However, for the 8 cells that attainedcriterion for both background and evokedactivity, the direction of change was differ-ent for the two aspects of neuronal activity

180 N. WEINBERGER, W. HOPKINS, AND D. DIAMOND
Table 2Effect of Conditioning on Background Activity
Cell no.
1D23B5D5E6B9A10B
nMSD
Increase
Pupil
8166
11
1011
610.333.39
Background
32321632
71428
723.0010.44
Cell no.
5B6C7E8A
nMSD
Decrease
Pupil
177
11
311.675.03
Background
10214317
421.2519.60
Cell no.
IB1C3Ea
6A17B*7C8B8C8D10A
nMSD
No change
Pupil Background
6 —12 —(-) (-)18 -(-) (-)6 —
16 —7 —
27 —
7 813.14 —7.80 —
* Deleted from analysis because neither pupillary learning nor neuronal plasticity developed.
in all cases (Table 2), which is statisticallysignificant x
2 ( l , N = 8) = 4.50, p < .05.The rates of change for evoked and back-
ground activity were found to be signifi-cantly different: Evoked activity attainedcriterion in fewer trials than did back-ground activity, £(21) = 2.27, p < .05.
Relation Between Arousal Level andNeuronal Discharge Plasticity
The measurement of pupillary diameterduring conditioning provides an opportu-nity to investigate the relation betweenarousal level and neuronal discharge plas-ticity. The relation between pupillary di-ameter and arousal level is well established:increased diameter, increased arousal level,and vice versa (e.g., Nunnally, Knott,Duchnowski, & Parker, 1967). It is usefulto distinguish between transient or phasicarousal and enduring or tonic arousal (So-kolov, 1963). In the present study and itscompanion study on secondary auditorycortex (D. Diamond & Weinberger, 1984),unconditioned phasic arousal is operation-ally defined as the dilation that is evokedby the unconditioned stimulus and attainspeak within 1.5 s of its onset. Tonic arousalis defined as the baseline (pretrial) level ofthe pupil.
To assess the effects of phasic arousal on
neurons in AI, we compared cellular dis-charges for the 1.5 s immediately precedingthe EDS with discharges for the 1.5 s im-mediately following this stimulus for everyUS trial during the sensitization phase.This yielded 15 pairs of values which werethen evaluated by the Mann-Whitney Utest for each cell separately. EDS producedpupillary dilation indicative of phasicarousal on every trial. Only 6 of 19 cellswere responsive to EDS (Mann-Whitney Utests, p < .05 or less); 5 cells responded withincreased firing, and 1 cell with decreaseddischarges. There were no significant rela-tions between the effects of phasic arousaland the effects of training, for either back-ground or evoked activity.
Tonic arousal was indexed by the size ofthe pupil immediately preceding the onsetof each acoustic stimulus, hereafter referredto as pupillary baseline. These data wereaveraged for consecutive blocks of fivetrials and are expressed as percentagechange from the mean value of the last fiveCS trials of the sensitization period for eachcase. In order to determine the relation ofpupillary baseline to background or evokedactivity, the data were assigned to groupson the basis of the effects of training onbackground and evoked activity, and aver-aged. Thus, the average pupillary baselinedata were computed for six groups: in-

PLASTICITY OF SINGLE AI NEURONS DURING LEARNING 181
creases, decreases, and no change for back-ground discharges and for evoked dis-charges, respectively. These findings arepresented in Figure 6.
Two sets of findings emerged from thisanalysis. First, with respect to background
60 -
_ i i i i
1 2 3 1 2 3 4 5 6 7 8 9SENSITIZATION CONDITIONING
BLOCKS OF FIVE TRIALS
Figure 6. The level of tonic arousal for the variousoutcomes of training upon background (A) and evoked(B) discharges (Tonic arousal is defined as pupillarybaseline, i.e., the level of dilation during intertrialintervals, preceding presentation of the conditionedstimulus. Each point is the mean percentage changeof the average level of the pupillary baseline for blocksof five trials, relative to the last five-trial block duringsensitization. Each function is the change in pupillarybaseline for those neurons that developed increases[solid circles], decreases [open circles], or no signifi-cant change [open triangles] in background andevoked discharges during conditioning. A: Backgrounddischarges—neurons that developed increased back-ground activity occurred in subjects whose level oftonic arousal increased throughout CS-US pairing.Neurons that either developed decreased or no changein background activity were obtained in subjects whoselevel of tonic arousal did not increase. B: Evokeddischarges—cells whose evoked activity failed tochange during conditioning were recorded in animalswhose level of tonic arousal increased during condi-tioning. Such increased tonic arousal was not associ-ated with neurons that developed evoked dischargeplasticity during conditioning. Vertical bars denote ±1SE.)
discharges, those neurons that developedincreased background activity during CS-US pairing were in subjects that developedincreased tonic arousal during pairing(Mann-Whitney U test, p < .001). Thosecells that developed decreased backgroundactivity or failed to change in backgroundactivity were in subjects that tended towardlower, but not significant, levels of tonicarousal (Figure 6A).
Second, with respect to evoked activity,those neurons that failed to develop dis-charge plasticity were in subjects in whichtonic arousal increased during CS-US pair-ing (Mann-Whitney Utest,p < .02). Therewas no statistically significant relation be-tween pupillary baseline and neurons thatdeveloped either increases or decreases inevoked discharges (Mann-Whitney U test,p > .05; Figure 6B).
Thus, increasing tonic arousal duringconditioning was compatible with the de-velopment of increased background dis-charges but was incompatible with the de-velopment of plasticity of evoked dis-charges.
Single-Unit Data and Multiple-Unit Data
One reason that the present data forsingle neurons were obtained is that mul-tiple-unit data, that is, unsorted dischargesrecorded simultaneously from more thanone neuron by a single electrode, may beinsensitive to different types of dischargeplasticity; for example, such data may maskdivergent changes in various neurons. Al-though this experiment did not involve therecording of multiple-unit data, it was fea-sible to construct composite "multiple-unit" histograms by combining the recordsof all individual neurons. Because this hadto be done laboriously by hand on a bin-by-bin basis, we undertook this processonly for selected blocks of trials, specificallyfor the last 5 CS trials of sensitization andTrials 16-20 during conditioning. The lat-ter were selected because most dischargeplasticity was present during this part oftraining. These compilations yielded the"multiple-unit" histograms shown in Fig-ure 7. They reveal an effect of conditioning.

182 N. WEINBERGER, W. HOPKINS, AND D. DIAMOND
Note that there is an increase in evokeddischarges following onset of the CS duringTrials 16-20 of conditioning relative to thelast 5 trials of sensitization. Decreases thatare evident in single unit data are masked,as are the changes in background activityof single neurons.
Discussion
Acoustically evoked pupillary responsesdeveloped progressive increases during theconditioning phase of training. Suchchanges never occurred during sensitiza-tion during which the CS and US were notpaired. Further, increments in the pupillaryresponse developed rapidly, reaching crite-
1000
1000
Figure 7. "Multiple-unit" poststimulus histogramsconstructed by summing the number of discharges ofall 21 neurons from which data were recorded duringtraining. (The numbers of spikes were determined for500 ms immediately preceding presentation of theconditioned stimulus and for the 1,000 ms duringpresentation of the conditioned stimulus. Histogramsdepicted are for the last five trials of the sensitizationperiod [SENS.] and for a representative five-trialblock [COND.] during conditioning [Trials 16-20].Notice the increase in the evoked discharge followingonset of the conditioned stimulus during conditioningrelative to sensitization.)
rion in an average of 12 trials. In the pres-ent case, a discrimination paradigm, todemonstrate stimulus-specific pupillaryconditioning, was not employed because itwould have prolonged the duration of train-ing and thus greatly reduced the probabilityof obtaining continuous recordings fromsingle cells during the development of con-ditioned responses. We have reported thatpupillary conditioning exhibits all of themajor characteristics of Pavlovian condi-tioned responses: systematic increase inmagnitude due to stimulus pairing, discrim-ination both within and between modali-ties, discrimination reversal, conditionedinhibition, and inhibition of delay (Olesonet al., 1972, 1973, 1975; Ryugo & Weinber-ger, 1978; Weinberger et al., 1973). There-fore, the pupillary dilation conditioned re-sponse can serve as a framework for theanalysis of the effects of associative proc-esses on the discharges of neurons in audi-tory cortex. Further, as discussed in Re-sults, it provides a basis for the interpre-tation of "negative" neuronal data, that is,instances in which discharge plasticity forbackground or evoked activity did not de-velop during the pairing of the conditionedand unconditioned stimuli.
A major finding is that discharge plastic-ity is prevalent in Al neurons during clas-sical conditioning. Of 19 cells, 15 developedstatistically significant changes in eitherevoked or background activity, or both.These effects are attributable to associativeprocesses, for several reasons. The effectsof CS-US pairing were assessed relative toa sensitization control period. Stimulusconstancy was assured by neuromuscularblockade, which eliminates movement ofthe head or pinna with respect to the soundsource (Marsh, Worden, & Hicks, 1962;Wiener, Pfeiffer, & Backus, 1966), contrac-tion of the middle ear muscles (Starr, 1964),and movement-induced masking noise(Imig & Weinberger, 1970). Finally, neu-ronal changes due to alleged sensory feed-back from conditioned responses, that is,the pupillary dilation conditioned response,can be ruled out because the pupillary mus-culature does not contain proprioceptors(Lowenstein & Loewenfeld, 1969).
Although these effects are associative,

PLASTICITY OF SINGLE AI NEURONS DURING LEARNING 183
Kitzes, Farley, and Starr (1978) argued thatsuch discharge plasticity in sensory systemsmay be unrelated to the cue value of theconditioned stimulus. These experimenterspresented brief tones continuouslythroughout defensive Pavlovian training inwhich the CS was white noise. They re-ported changes in AI single-unit activity tothe tones and periods of silence presentduring CS-US intervals relative to inter-trial intervals. They concluded that train-ing causes merely a general change in cor-tical excitability so that evoked dischargeplasticity is not directly related to the sig-nificance of the conditioned stimulus. How-ever, this conclusion rests on the assump-tion that the tone pips were devoid of sig-nificance during the CS-US interval, butKitzes et al. failed to provide independentevidence to support this assumption. Quitethe contrary, these stimuli may have hadCS properties because of the particulartraining regimen employed. The subjectswere trained initially with a 5-s white noiseCS, followed by electrodermal stimulation.After establishment of the pupillary con-ditioned response, they were shifted from adelay to a trace paradigm, which resultedin a 0.5-s CS (white noise) followed by 4.5s of silence during which "neutral" tonepips could be presented. But the originaltraining caused conditioning to 5 s of acous-tic stimulation, and the pips filling the CS-US interval could have acquired CS valuedue to within-modality stimulus generali-zation, that is, the effective CS was actuallywhite noise followed by tone pips. A moresevere problem is that the control groupreceived only tones, not tones unpairedwith EDS, hence the effects were not de-monstrably associative. Finally, the rele-vance of these findings to the acquisitionof response plasticity, as in the presentcase, is unknown. Resolution of this partic-ular issue must await more definitive ex-periments.
Relation Between Pupillary and NeuralConditioned Responses
The present study replicates and extendsto single neurons two findings reported pre-viously (Oleson et al., 1975): (a) The rates
of acquisition of pupillary conditioned re-sponses (CRs) and evoked discharge plas-ticity are rapid and do not differ from eachother and (b) development of pupillary CRsand evoked discharge plasticity were notclosely related, for example, conditionedresponses developed during some sessionsin which neuronal plasticity did not appear,and vice versa. These findings suggest thatthe neuronal changes are not causal to thepupillary CRs. That both develop at thesame rapid rate during associative learningsuggests that they are related to a commonprocess which has not yet been delineated.
Although neuronal changes did not pre-cede acquisition of the pupillary dilationconditioned reflex, it is likely that theyprecede the acquisition of several otherconditioned responses because the rate ofpupillary conditioning is among the fastestof any response system (Weinberger, 1982a,1982b). During defensive conditioning, re-sponses that are not specific to the natureof the unconditioned stimulus develop con-ditioned responses more rapidly than dothose systems in which the conditioned re-sponse is determined by the specific char-acteristics of the unconditioned response(Schneiderman, 1972). Among the formerare pupil, cardiovascular, respiration, andgeneral skeletal movement. Responses thatare specific to the nature of the US includelimb flexion, eye blink, and extension ofthe nictitating membrane. The presentfindings indicate that discharge plasticityfor single neurons in primary auditory cor-tex develops as rapidly as does pupillaryconditioning. Such rapidly developing be-havioral and neuronal plasticity may com-prise part of the first stage of a two-stage(Konorski, 1967) or three-stage (Thompsonet al., in press) conditioning process inwhich the second stage is the elaborationof a somatic conditioned response which isspecific to the nature of the unconditionedstimulus (Weinberger, 1982a, 1983).
The functional role of associatively in-duced discharge plasticity in primary au-ditory cortex, and for that matter in othersensory systems, is unknown. Evoked plas-ticity may reflect changes in the thresholdfunctions or in the receptive field propertiesof sensory neurons, but appropriate exper-

184 N. WEINBERGER, W. HOPKINS, AND D. DIAMOND
iments have not yet been reported It isknown that ablations of auditory cortexproduce deficits in the discrimination ofcomplex acoustic stimuli (Neff, Diamond,& Cassady, 1975), and evidence suggeststhat auditory cortex is essential for theappreciation of stimulus constancy orequivalence (Whitfield, 1979). The factthat discharge plasticity develops rapidly inauditory cortex during Pavlovian condi-tioning suggests that even apparently sim-ple learning situations may be involved insuch complex processes.
Relation to Previous Studies
The present report appears to be the firstin which the discharges of single neuronswere recorded in the primary auditory cor-tex during the acquisition of a behavioralconditioned response. Issues relating to au-ditory cortical unit discharges during theperformance of previously acquired re-sponses are considered in a companion ar-ticle on auditory cortical field All (D. Dia-mond & Weinberger). However, there areseveral previous reports in which multiple-unit activity was recorded during condi-tioning.
With respect to evoked discharges, thereis consistent evidence that responses inauditory cortex to an acoustic conditionedstimulus are augmented during condition-ing. This has been found in the cat duringPavlovian defensive conditioning both infreely moving and in muscle-blocked sub-jects (Buchwald et al., 1966; Halas et al.,1970; Oleson et al., 1975) and during in-strumental conditioning (Halas et al.,1970), in the rat during a hybrid classi-cal-instrumental appetitive task (Dister-hoft & Olds, 1972; Disterhoft & Stuart,1976; Olds, Disterhoff, Segal, Kornblith, &Hirsh, 1972) and in the rabbit during in-strumental avoidance learning (Foster,Orona, Lambert, & Gabriel, 1980). Thisconcordance is noteworthy, given the vari-ations in subjects, tasks, and conditions oftraining, plus the fact that the data re-ported for the rat include activity fromcortical sites beyond the limits of auditorycortex. The present results for single neu-rons reveal that decreases in evoked dis-
charges also develop and that they do so atthe same rapid rate as do increases inevoked discharges. Thus it appears likelythat multiple-unit recordings mask de-creases in discharge plasticity during learn-ing. This conclusion is underscored by theanalysis in which we constructed "multiple-unit" histograms by combining the datafrom single neurons (Figure 7).
In regard to background multiple-unitactivity, there is less agreement. Disterhoftand co-workers reported the developmentof decreases in background activity (Dister-hoft & Olds, 1972; Disterhoft & Stuart,1976), whereas we found a pronounced in-crease in background discharges during dif-ferential conditioning and reversal of dif-ferential conditioning (Oleson et al., 1975).A further difference is that those experi-menters reported that background activitychanged before evoked activity whereas wefound the opposite relation. The presentstudy extends this finding to single units:Evoked activity attained criterion signifi-cantly earlier than did background dis-charges. It is difficult to reconcile the con-trary findings, particularly because the con-cordance regarding evoked activity in stud-ies of multiple-unit activity suggests thatdifferences in subjects and training condi-tions may still yield similar findings forevoked activity. It may be that changes inbackground activity are more tightly linkedto the cytoarchitectonically delimited re-gion of primary auditory cortex than arechanges in evoked activity. In any event, itis now evident that previous studies havereported either increases or decreases inbackground activity whereas the presentstudy of single neurons has revealed bothincreases and decreases under the sameconditions of training and acquisition of abehavioral conditioned response. Studies,such as the current investigation, that un-dertake finer grain analyses than previousexperiments are likely to find complexitiesheretofore not discovered. Determinationof the functional role indexed by such find-ings remains a challenge.
Single-Unit and Multiple-Unit DataKraus and Disterhoft (1982) argued for
the importance of obtaining discharge data

PLASTICITY OF SINGLE AI NEURONS DURING LEARNING 185
from single neurons because of the limita-tions of multiple-unit recordings. We arein complete agreement with this view, andthe present findings offer empirical supportfor the position that multiple-unit recordsare not adequate representations of thechanges that develop for single cells; forexample, as pointed out above, multiple-unit histograms indicative of response plas-ticity indicate an increase in evoked dis-charges during learning. In this study, weconstructed "multiple-unit" histograms byadding the histograms of single neurons.These histograms are not identical to stan-dard multiple-unit records because theywere obtained from separate experimentalsessions and various loci and their compo-sition is known. Nonetheless, they also pro-vided a picture of an increase in evokedresponse to the CS during conditioning(Figure 7). Yet the histograms of the indi-vidual neurons are heterogeneous, includ-ing decreases and no changes as well asincreases during CS-US pairing. Thus,multiple-unit records are not necessarilycomprised of the discharges of a homoge-neous population of neurons, all or most ofwhich develop an increase in response.Also, the "masking" of unit heterogeneityholds for background as well as evokedactivity (Figure 7). Therefore, it would beincorrect to conclude that the general ex-citability of sensory cortex simply increasesduring conditioning.
Previously, we reported similar resultsfor discharge plasticity in the magnocellu-lar medial geniculate nucleus (MGm).Poststimulus histograms of multiple-unitactivity exhibit the same general pattern ofactivity and the same sort of increase inevoked activity during conditioning as re-ported for the primary auditory cortex(Ryugo & Weinberger, 1976, 1978). How-ever, changes in the discharges of individ-ual neurons in the MGm also are hetero-geneous for both background and evokedactivity (Weinberger, 1982a). Thus, multi-ple-unit activity overshadows certain as-pects of single-unit discharge plasticity inthe thalamus as well as in the cortex. Thereis no reason to doubt that the same problemmay exist for multiple-unit recordings fromother parts of the brain, as well. Therefore,
appropriate caution should be exercised inthe interpretation of multiple-unit records.
Discharge Plasticity, Conditioning, andTonic Level of Arousal
Plasticity of background and evoked dis-charges was a common event in this study.Given the caveat that any microelectrodestudy may be limited by sampling bias andthat classes of neurons not yet describedmay exist in primary auditory cortex, thesefindings are in essential agreement with theobservations of Kraus and Disterhoft(1982), who emphasized the heterogeneityof discharge plasticity in the auditory as-sociation cortex of the rabbit. This can beseen in the highly variable shapes of post-stimulus histograms and in the fact thatconditioning is accompanied by both in-creases and decreases in discharges as wellas lack of change.
However, one facet of this heterogeneitymay be explicable. With the pretrial size ofthe pupil used as an index of the level oftonic arousal, it was found that increasedtonic arousal was accompanied by the fail-ure of neurons to develop discharge plastic-ity in evoked activity. Although no causalroles can be assigned yet, it does seem thatincreased tonic arousal level is not condu-cive to evoked discharge plasticity. Thisseems to be a neurophysiological expressionof the well-known Yerkes-Dodson law thathigh levels of arousal impair learning andperformance (e.g., for review, see Eysenck,1982).
Possible Mechanisms of DischargePlasticity in Primary Auditory Cortex
The discharge properties of neurons inAI during conditioning may be, in part, afunction of the combined influences of theirthalamic sources of input, that is, the ven-tral and magnocellular medial geniculatenuclei. As the ventral medial geniculatedoes not develop discharge plasticity duringconditioning whereas the magnocellulardoes (see introduction), the plastic proper-ties of AI neurons might relate to inputfrom the latter region. In addition, theremay be mechanisms of plasticity intrinsic

186 N. WEINBERGER, W. HOPKINS, AND D. DIAMOND
to the cortex and other extrinsic sources aswell. However, there is a striking similaritybetween the effects of conditioning onMGm and the effects on AI neurons. Usingan identical preparation and training regi-men, we reported that evoked dischargeplasticity developed at the same rate as forAI cells, that is, 10-20 trials.
Although the present picture is very in-complete, it is now possible to propose apreliminary hypothesis of how associativeevoked discharge plasticity may develop inthe primary auditory cortex. The lemniscal,tonotopic projection from the ventral me-dial geniculate body (MGv) terminatesdensely in the middle layers of AI (I. Dia-mond, 1979; Herkenham, 1980; Jones &Rockel, 1971). However, the discharges ofthe MGv are not altered by learning, sothat the predominant input to the middlelayers of AI probably reflects only the phys-ical parameters of stimuli. It should benoted that the rates of discharges of indi-vidual neurons in MGv can vary widely inresponse to a physically constant stimuluswhen background activity changes in theunanesthetized cat (Humphrey & Orman,1977; Imig & Weinberger, 1973). However,the pattern of response is conserved despitefluctuations in the actual number of dis-charges (Imig & Weinberger, 1973). There-fore, it is not necessary to hold that primaryauditory cortex always receives the samenumber of discharges to the same acousticstimulus via the MGv but only that thereis some constant relation between the phys-ical parameters of an acoustic stimulus andthe input from the MGv to primary audi-tory cortex. In contrast, the magnocellularmedial geniculate body projects to layer Iof AI (Burton & Jones, 1976; Herkenham,1980; Jones & Rockel, 1971; Ryugo & Kil-lackey, 1974). This arrangement suggeststhat the MGm regulates the responses toacoustic information of neurons whosesoma are in lower layers, perhaps by effectson apical dendrites that ascend to the upperlamina.
The failure to develop evoked dischargeplasticity in primary auditory cortex hasbeen linked in the present study to highlevels of tonic arousal, as discussed above.The possible mechanisms cannot be con-
sidered yet because there are essentially nopertinent data on this point. Rather, itwould seem that future inquiry into themechanisms of response plasticity in audi-tory cortex, and perhaps in many otherareas as well, also will have to attempt toaccount for such failures to develop plastic-ity.
Finally, the present findings were ob-tained from single neurons located mainlyin the infragranular lamina (V and VI)which are the major sources of subcorticalprojections from AI (I. Diamond, 1979;Kelly & Wong, 1981). It is therefore note-worthy that these "output" neurons arealtered by associative processes. The con-sequences of associative discharge plastic-ity in these neurons, and the intracorticalmechanisms that may lead to thesechanges, will require extensive detailedstudy, including laminar analyses duringlearning.
References
Aitkin, L. M. (1973). Medial geniculate body of thecat: Responses to tonal stimuli of neurons in medialdivision. Journal of Neurophysiology, 36, 275-283.
Aitkin, L. M., & Webster, W. R. (1972). Medial genic-ulate body of the cat: Organization and responses totonal stimuli of neurons in the ventral division.Journal of Neurophysiology, 35, 365-380.
Ashe, J. H., Cassady, J. M., & Weinberger, N. M.(1976). The relationship of the cochlear micro-phonic potential to the acquisition of a classicallyconditioned pupillary dilation response. BehavioralBiology, 16, 45-62.
Birt, D., Nienhuis, R., & Olds, M. (1979). Separationof associative from non-associative short latencychanges in medial geniculate and inferior colhculusduring differential conditioning and reversal in rats.Brain Research, 167, 129-138.
Birt, D., & Olds, M. (1981). Associate responsechanges in lateral midbrain tegmentum and medialgeniculate during differential appetitive condition-ing. Journal of Neurophysiology, 46, 1039-1055.
Buchwald, J. S., Halas, E. S., & Schramm, S. (1966).Changes in cortical and subcortical unit activityduring behavioral conditioning. Physiology and Be-havior, 1, 11-22.
Burton, H., & Jones, E. G. (1976). The posteriorthalamic region and its cortical projection in NewWorld and Old World monkeys. Journal of Compar-ative Neurology, 168, 249-302.
Cassady, J. M., Cole, M., Thompson, R. F., & Wein-berger, N. M. (1973). Neural correlates of asymp-totic avoidance and classical conditioned leg flexion.Experimental Neurology, 40, 207-215.
Cassady, J. M., Farley, G. R., Weinberger, N. M., &

PLASTICITY OF SINGLE AI NEURONS DURING LEARNING 187
Kitzes, L. M. (1982). Pupillary activity measured byreflected infra-red light. Physiobgy and Behavior,28, 851-854.
Diamond, D. M , & Weinberger, N. M. (1984). Phys-iological plasticity of single neurons in auditorycortex of the cat during acquisition of the pupillaryconditioned response: II. Secondary field (All). Be-havioral Neuroscience, 98, 189-211.
Diamond, I. (1979). The subdivisions of neocortex: Aproposal to revise the traditional view of sensory,motor, and association areas. In J. M. Sprague & A.N. Epstein (Eds.), Progress in psychobiobgy andphysiological psychology (Vol. 8, pp. 1-43). NewYork: Academic Press.
Disterhoft, J., & Olds, J. (1972). Differential develop-ment of conditioned unit changes in thalamus andcortex of rat. Journal of Neurophysiology, 35, 665-679.
Disterhoft, J., & Stuart, D. (1976). Trial sequence ofchanged unit activity in auditory system of alert ratduring conditioned response acquisition and extinc-tion. Journal of Neurophysiology, 39, 266-281.
Eysenck, M. W. (1982). Attention and arousal. NewYork: Springer.
Feller, W. (1968). An introduction to probability theoryand its applications. New York: Wiley.
Foster, K., Orona, E., Lambert, R., & Gabriel, M.(1980). Neuronal activity in the auditory systemduring differential conditioning in rabbits. Societyfor Neuroscience Abstracts, 6, 424.
Freeman, W. J. (1980). Evidence for an olfactorysearch image or representation in the EEG of con-ditioned cats and rabbits. Advancement in Physio-logical Sciences, 16, 421-429.
Furedy, J. J. (1971). Explicitly-unpaired and truly-random CS- controls in human classical differentialautonomic conditioning. Psychophysiology, 8, 497-503.
Furedy, J. J., Poulos, C. X., & Schiffman, K. (1975).Contingency theory and classical autonomic excit-atory and inhibitory conditioning: Some problemsof assessment and interpretation. Psychophysiology,12, 98-105.
Gabriel, M., Miller, J. D., & Saltwick, S. E. (1976).Multiple unit activity of the rabbit medial geniculatenucleus in conditioning, extinction, and reversal.Physiological Psychology, 4, 124-134.
Galambos, R., Sheatz, G. C , & Vernier, B. (1955).Electrophysiological correlates of a conditioned re-sponse in cats. Science, 123, 376-377.
Graybiel, A. (1972). Some fiber pathways related tothe posterior thalamic region in the cat. Brain,Behavior and Evolution, 6, 363-393.
Halas, E. S., Beardsley, J. V., & Sandlie, M. E. (1970).Conditioned neuronal responses at various levels inconditioning paradigms. Electroencephalographyand Clinical Neurophysiology, 28, 468-477.
Herkenham, M. (1980). Laminar organization of tha-lamic projections to the rat neocortex. Science, 207,532-535.
Hopkins, W., & Weinberger, N. M. (1980). Modifica-tion of auditory cortex single unit activity duringpupillary conditioning [Abstract]. Society for neu-roscience: Proceedings of the Tenth Annual Meeting,6, 424.
Humphrey, G. L., & Orman, S. S. (1977). Activity ofthe auditory system related to arousal. ExperimentalNeurology, 55, 520-537.
Imig, T. J., & Weinberger, N. M. (1970). Auditorysystem multi-unit activity and behavior in the rat.Psychonomic Science, 18, 164-165.
Imig, T. J., & Weinberger, N. M. (1973). Relationshipsbetween the rate and pattern of unitary dischargesin the medial geniculate body of the cat in responsesto click and amplitude-modulated white noise stim-ulation. Journal of Neurophysiology, 36, 385-397.
John, E. R. (1961). High nervous functions: Brainfunctions and learning. Annual Review of Physiol-ogy, 23, 451-484.
Jones, E. G., & Rockel, A. J. (1971). The synapticorganization in the medial geniculate body of affer-ent fibres ascending from the inferior colliculus.Zeitschrift fur Zellforschung und MikroskopischeAnatomie, 113, 44-66.
Kelly, J. P., & Wong, D. (1981). Laminar connectionsof the cat's auditory cortex Brain Research, 212, 1-15.
Kitzes, L. M., Farley, G. R., & Starr, A. (1978). Mod-ulation of auditory cortex unit activity during theperformance of a conditioned response. Experimen-tal Neurology, 62, 678-697.
Konorski, J. (1967). Integratiue activity of the brain.Chicago: University of Chicago Press.
Kraus, N., & Disterhoft, J. F. (1982). Response plas-ticity of single neurons in rabbit auditory associa-tion cortex during tone-signalled learning. BrainResearch, 246, 205-215.
Lowenstein, O., & Loewenfeld, I. E. (1969). The Pupil.In H. Davson (Ed.), The eye (pp. 255-337). NewYork: Academic Press.
Marsh, J. T., Worden, F. G., & Hicks, L. (1962). Someeffects of room acoustics on evoked auditory poten-tials. Science, 137, 280-282.
Merzenich, M., & Kaas, J. (1980). Principles of orga-nization of sensory-perceptual systems in mammals.In J. M. Sprague & A. N. Epstein (Eds.), ProgressIn Psychobiology and Physiological Psychology (Vol.9, pp. 1-42). New York: Academic Press.
Morest, D. K. (1964). The neuronal architecture ofthe medial geniculate body of the cat. Journal ofAnatomy, 98, 611-630.
Morest, D. K (1965). The laminar structure of themedial geniculate body of the cat. Journal of Anat-omy, 99, 611-634.
Neff, W. D., Diamond, I. T., & Cassady, J. H. (1975).Behavioral studies of auditory discrimination: cen-tral nervous system. In W. D. Keibel & W. D. Neff(Eds.), Handbook of sensory physiology (Vol. 5, Pt.2, pp. 307-400). New York: Springer Verlag.
Nunnally, J. C , Knott, P. D., Duchnowski, A., &Parker, R. (1967). Pupillary response as a generalmeasure of activation. Perception & Psychophysics,2, 149-155.
Olds, J., Disterhoft, J. F., Segal, M., Kornblith, C , &Hirsh, R. (1972). Learning centers of the brainmapped by measuring latencies of conditioned unitresponses. Journal of Neurophysiology, 35, 202-219.
Oleson, T. D., Ashe, J. H., & Weinberger, N. M. (1975).Modification of auditory and somatosensory systemactivity during pupillary conditioning in the para-

188 N. WEINBERGER, W. HOPKINS, AND D. DIAMOND
lyzed cat. Journal of Neurophysiology, 38, 1114-1139.
Oleson, T. D., Vododnick, D. S., & Weinberger, N. M.(1973). Pupillary inhibition of delay during Pavlov-ian conditioning in paralyzed cat. Behavioral Biol-ogy, 8, 337-346.
Oleson, T. D., Westenberg, I. S., & Weinberger, N. M.(1972). Characteristics of the pupillary dilation re-sponse during Pavlovian conditioning in paralyzedcats. Behavioral Biology, 7, 829-840.
Reale, R. A., & Imig, T. J. (1980). Tonotopic organi-zation in auditory cortex of the cat. Journal ofComparative Neurology, 192, 265-294.
Rose, J. E. (1949). The cellular structure of the audi-tory region of the cat. Journal of Comparative Neu-rology, 21, 409-440.
Ryugo, D. K., & Killackey, H. P. (1974). Differentialtelencephalic projections of the medial and ventraldivisions of the medial geniculate body of the rat.Brain Research, 82, 173-177.
Ryugo, D. K., & Weinberger, N. M. (1976). Differen-tial plasticity of morphologically distinct neuronpopulations in the medial geniculate body of the catduring classical conditioning [Abstract]. Society forNeuroscience- Proceedings of the Sixth Annual Meet-ing 2, 435.
Ryugo, D. K., & Weinberger, N. M. (1978). Differen-tial plasticity of morphologically distinct neuronpopulations in the medial geniculate body of the catduring classical conditioning. Behavioral Biobgy,22, 275-301.
Schneiderman, N. (1972). Response system divergen-cies in aversive classical conditioning, In A. H.Black & W. F. Prokasy (Eds.), Classical conditioningII: Current research and theory (pp. 341-376). NewYork: Appleton-Century-Crofts.
Segundo, J. P., Galeano, C, Sommer-Smith, J. A., &Roig, J. A. (1961). Behavioural and EEG effects oftones 'reinforced' by cessation of painful stimuli. InJ. F. Delafresnaye (Ed.), Brain mechanisms andteaming (pp. 265-291). Oxford: Blackwell ScientificPublications.
Shinkman, P. G., Bruce, C. G., & Pfingst, B. E. (1974)Operant conditioning of single-unit response pat-terns in visual cortex. Science, 184, 1194-1196.
Siegel, S. (1956). Nonparametric statistics for the be-havioral sciences. New York: McGraw-Hill.
Sokolov, E. N. (1963). Perception and the conditionedreflex. Oxford: Pergamon Press.
Sokolov, E. N. (1977). Brain function: Neuronal mech-anisms of learning and memory. Annual Review ofPsychology, 28, 85-112.
Sousa-Pinto, A. (1973). The structure of the firstauditory cortex (AI) in the cat. I. Light microscopicobservations on its organization. Archives of ItalianBiology, 111, 112-137.
Starr, A. (1964). Influence of motor activity on click-evoked responses in the auditory pathway of wakingcats. Experimental Neurology, 10, 191-204.
Thompson, R. F., Clark, G. A., Donegan, N. H., La-vond, D. G., Lincoln, J. S., Madden, J., Mamounas,L. A., Mauk, M. D., McCormick, D. A., & Thomp-son, J. K. (in press). Neuronal substrates of learningand memory: A "multiple-trace" view. In G. S.Lynch, J. L. McGaugh, & N. M. Weinberger (Eds.),Neurobiology of learning and memory. New York:Guilford Press.
Thompson, R. F., Patterson, M. M., & Teyler, T. J.(1972). The neurophysiology of learning. AnnualReview of Psychology, 23, 73-104.
Voronin, L. L., Gerstein, G. L., Kudryashov, I. E., &Ioffe, S. V. (1975). Elaboration of a conditionedreflex in a single experiment with simultaneousrecording of neural activity. Brain Research, 92,385-^03.
Weinberger, N. M. (1982a). Effects of conditionedarousal on the auditory system. In A. L. Beckman.(Ed.), The neural basis of behavior (pp. 63-91). NewYork: Spectrum Publications.
Weinberger, N. M. (1982b). Sensory plasticity andlearning: The magnocellular medial geniculate nu-cleus of the auditory system. In C. D. Woody, (Ed.),Conditioning: Representation of involved neuralfunction (pp. 697-710). New York: Plenum Publish-ing.
Weinberger, N. M. (in press). The neurophysiology oflearning: A view from the sensory side. In N. Butters& L. Squire (Eds.), The neuropsychology of memory.New York: Guilford Press.
Weinberger, N. M., Oleson, T. D., & Haste, D. (1973).Inhibitory control of conditional pupillary dilationresponse in the paralyzed cat. Behavioral Biology, 9,307-316.
Whitfield, I. C. (1979). The object of sensory cortex.Brain, Behavior and Evolution, 16, 129-154.
Wiener, J. M., Pfeiffer, R. R., & Backus, A. S. M.(1966). On the sound pressure transformation bythe head and auditory meatus of the cat. Acta Oto-laryngolica, 61, 255-269.
Winer, B. J. (1971). Statistical principles in experimen-tal design (2nd ed.). New York: McGraw-Hill.
Winer, J. A., Diamond, I. T., & Raczkowski, D. (1977).Subdivisions of the auditory cortex of the cat: Re-trograde transport of horseradish peroxidase to themedial geniculate body and posterior thalamic nu-clei. Journal of Comparative Neurology, 176, 387-418.
Received August 17,1983Revision received November 29,1983








![American Psychological Association (APA) [Bibliography Writing]](https://img.dokumen.tips/doc/110x75/58f0f8bc1a28abde038b4575/american-psychological-association-apa-bibliography-writing.jpg)




















