Embed Size (px)
Citation preview

TECHNOLOGY REPORT
Conditional LoxP-Flanked Glucosylceramide SynthaseAllele Controlling Glycosphingolipid SynthesisTadashi Yamashita,1 Maria Laura Allende,1 Danielle N. Kalkofen,1 Norbert Werth,2
Konrad Sandhoff,2 and Richard L. Proia1*1Genetics of Development and Disease Branch, National Institute of Diabetes and Digestive and Kidney Diseases,National Institutes of Health, Bethesda, Maryland2Kekule-Institut fur Organische Chemie und Biochemie, Universitat Bonn, Bonn, Germany
Received 20 June 2005; Accepted 15 September 2005
Summary: Glycosphingolipids are organizational build-ing blocks of plasma membranes that participate in keycellular functions, such as signaling and cell-to-cellinteractions. Glucosylceramide synthase—encoded bythe Ugcg gene—controls the first committed step in themajor pathway of glycosphingolipid synthesis. Global dis-ruption of the Ugcg gene in mice is lethal during gastrula-tion. We have now established a Ugcg allele flanked byloxP sites (floxed). When cre recombinase was expressedin the nervous system under control of the nestin pro-moter, the floxed gene underwent recombination, result-ing in a substantial reduction of Ugcg expression and ofglycosphingolipid ganglio-series levels. The mice defi-cient in Ugcg expression in the nervous system show astriking loss of Purkinje cells and abnormal neurologicbehavior. The floxed Ugcg allele will facilitate analysis ofthe function of glycosphingolipids in development, phys-iology, and in diseases such as diabetes and cancer.genesis 43:175–180, 2005. Published 2005 Wiley-Liss, Inc.
Key words: mouse; Ugcg; cre recombinase; glucosylcera-mide; sphingolipid
Glycosphingolipids are ubiquitous molecules found onthe outer leaflet of mammalian plasma membranes(Allende and Proia, 2002; Degroote et al., 2004; Kolteret al., 2002) where they function as structural compo-nents for the organization of lipid microdomains, as wellas mediators of cell-to-cell interactions and modifiers ofsignaling pathways. The first committed step in the syn-thesis of the majority of glycosphingolipids is directed bythe enzyme glucosylceramide synthase, which is encodedby the Ugcg gene (Ichikawa et al., 1996; Yamashita et al.,2002). The enzyme catalyzes the transfer of a glucose moi-ety from UDP-glucose to ceramide to form glucosylcera-mide, the precursor to most cellular glycosphingolipids.
Glycosphingolipids have been shown, largely in stud-ies using knockout mice, to be critical in developmentand physiology. Mice with a global disruption in Ugcg(UgcgKO) die during gastrulation, demonstrating an es-sential role for this gene in early development (Yamashita
et al., 1999). Mice without neuronal gangliosides (sialy-lated glycosphingolipids that are abundant in the nerv-ous system) show neurologic impairment, including neu-rodegeneration (Chiavegatto et al., 2000; Inoue et al.,2002; Kawai et al., 2001; Sheikh et al., 1999; Takamiyaet al., 1996; Yamashita et al., 2005). Glycosphingolipidsof the isoglobo-series are required for the developmentof natural killer T-cells, which are key regulators ofimmunity (Zhou et al., 2004). Fucosylated glycosphingo-lipids are essential for spermatogenesis (Sandhoff et al.,2005). The ganglioside GM3 has been shown to be anegative regulator of insulin receptor signaling, and itsoverexpression may be related to insulin resistance intype 2 diabetes (Tagami et al., 2002; Yamashita et al.,2003). Modulation of ganglioside expression has alsobeen found to be associated with human colon cancer(Kakugawa et al., 2002). In order to study the functionalaspects of glycosphingolipid synthesis in the adult, weestablished mice with a floxed Ugcg allele that can bedeleted in a conditional manner.
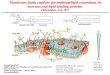
The Ugcg gene in TC1 embryonic stem (ES) cells wastargeted using the ploxPneoUgcg vector (Fig. 1a). TheloxPneo (Yang et al., 1998) cassette was introduced intothe 30 untranslated region, and a loxP site was insertedinto intron 6 of the Ugcg locus (Fig 1a). Completerecombination between the loxP sites would be pre-dicted to delete exons 7, 8, 9, and nearly all of the 30 un-translated region of the Ugcg allele. ES clones were iden-tified with the correct targeting arrangement (Fig. 1b),
Present address for Tadashi Yamashita: Department of Cell Processing, The
Institute of Medical Science, The University of Tokyo, Shirokanedai 4-6-1,
Minato-ku Tokyo, 108-8639 Japan.
* Correspondence to: Dr. Richard L. Proia, Building 10, Room 9N-314,
National Institutes of Health, 10 Center DR MSC 1821, Bethesda, MD 20892-
1821.
E-mail: [email protected]
Contract grant sponsors: Intramural Research Program of the National
Institutes of Health (NIH), National Institute of Diabetes and Digestive and
Kidney Diseases, and the Deutsche Forschungsgemeinshaft (SFB645).Published online in
Wiley InterScience (www.interscience.wiley.com).
DOI: 10.1002/gene.20167
' 2005 Wiley-Liss, Inc. genesis 43:175–180 (2005)

and one was used to establish chimeric mice. A chimericmale transmitted the targeted locus to his offspring,which were used as heterozygous mating pairs. ViableUgcgloxP/UgcgloxP offspring were obtained from the mat-ing pairs with the expected Mendelian frequency. South-ern analysis of genomic DNA from the brain and liver ofUgcgloxP/UgcgloxP mice demonstrated that the Ugcgallele was intact, as indicated by the presence of a 10.5-kb SacI band (Fig. 1c,d). The viability of the UgcgloxP/UgcgloxP mice also indicates that the floxed Ugcg allele
was not functionally disrupted by the targeting proce-dure, since homozygosity of the Ugcg knockout allele(UgcgKO) results in embryonic lethality.
To conditionally disrupt the Ugcg gene, we cross-bredmice carrying a UgcgloxP allele with mice carrying a cretransgene under control of the rat nestin (nes) promoterand enhancer (Tronche et al., 1999). These nes-cre trans-genic mice have been shown to express cre recombi-nase in neuronal and glial cell precursors during embry-onic development. We obtained mice with the genotype
FIG. 1. Generation and characterization of a floxed Ugcg allele. a: Schematic representation of the Ugcg targeting strategy. The structureof the UgcgloxP targeting vector is shown at the top, the mouse wildtype Ugcg allele in the middle, and the predicted structure of the recom-bined UgcgloxP allele on the bottom. UT, untranslated region; Neo, neomycin cassette; TK, thymidine kinase; Probe, location of probe usedto select clones. b: Identification of a targeted ES cell by Southern blotting. A Ugcg wildtype (WT) allele yielded a 5-kb band after PstI diges-tion, and the targeted Ugcg allele (UgcgloxP) yielded a 4.5-kb band. c: Schematic representation of cre-mediated recombination of theUgcgloxP allele. The structure of the UgcgloxP allele is shown at the top. The predicted structure of the allele (UgcgD) after cre-mediatedrecombination is shown on the bottom. d: Identification of cre-mediated recombination of the UgcgloxP allele in brain. The mice used wereUgcgloxP/UgcgloxP without (–) or with (þ) the nestin-cre transgene. Top: Genomic DNA from liver and brain was digested with SacI and sub-jected to Southern analysis with a probe corresponding to Ugcg exon 5. The UgcgloxP allele generates a 10.5-kb fragment and the UgcgD
allele yields a 5.5-kb fragment. Bottom: Genomic DNA from liver and brain was subjected to PCR using primers 1 and 2 to detect theUgcgloxP allele and primers 1 and 3 to detected theUgcgD allele. Location of primers for the PCR reactions is shown by the blue arrows in a and c.
176 YAMASHITA ET AL.

UgcgloxP/UgcgloxP carrying the nes-cre transgene. Analy-sis of SacI-digested genomic DNA from these micerevealed that the Ugcg locus was intact in liver, as evi-denced by the presence of the 10.5-kb band after South-ern analysis. However, in brain DNA of UgcgloxP/Ugc-gloxP mice carrying the nes-cre transgene the 10.5-kbband was substantially reduced, and a 5.5-kb band wasdetected that corresponded to the predicted cre-recom-
bined allele (UgcgD) (Fig. 1c,d). The tissue specificity ofrecombination was confirmed by polymerase chain reac-tion (PCR) of genomic DNA using primer sets (Fig. 1a,c)that detected either the intact UgcgloxP allele (primers 1and 2) or the cre-recombined UgcgD allele (primers 1and 3) (Fig. 1d, bottom panel).
We next used quantitative reverse transcriptase (RT)-PCR to determine the relative Ugcg mRNA levels in the
FIG. 2. Ugcg expression in the brain of NesUgcgKO mice. a: Real-time PCR was used to determine relative Ugcg mRNA expression inbrains from WT, floxed (UgcgloxP/UgcgloxP), and NesUgcgKO (UgcgloxP/UgcgloxP nes-cre) mice. The data represent the mean 6 standarddeviation, n ¼ 3. Statistical significance was calculated by two-tailed Student’s t-test (*P < 0.05, NesUgcgKO vs. Floxed; NesUgcgKO vs.WT). b: Biosynthetic pathway for brain gangliosides. The first glycosylation step is catalyzed by glucosylceramide synthase. The major braingangliosides are boxed. Cer, ceramide; GlcCer, glucosylceramide; LacCer, lactosylceramide; GalNAc, N-acetylgalactosamine; NeuAc, N-acetylneuraminic acid. c: The acidic lipid fraction was isolated from the brains of NesUgcgKO and age-matched control mice without brainspecific disruption of Ugcg. The major brain gangliosides (GM1, GD1a, GD1b, GT1b), visualized after thin-layer chromatography, are boxed.The ages of the mice were 2 weeks (2W) and 2 months (2M) old.
177CONDITIONAL LOXP-FLANKED GLUCOSYLCERAMIDE

brains of UgcgloxP/UgcgloxP mice carrying the nes-cretransgene and controls (Fig. 2a). The levels of Ugcg RNAfrom brain were not significantly different between thewildtype and UgcgloxP/UgcgloxP mice. Thus, as was sug-gested by the viability of the UgcgloxP/UgcgloxP mice, thetargeting of the Ugcg loci did not substantially alter UgcgRNA expression. However, in the brains of UgcgloxP/Ugc-gloxP mice with the nes-cre transgene present, the levelof Ugcg mRNA was reduced by greater than 70% (P <0.05). Similar results were obtained with mice with thegenotype UgcgloxP/UgcgKO with nes-cre (not shown);hereafter, we collectively refer to mice carrying the nes-cre transgene with the genotype UgcgloxP/UgcgloxP orUgcgloxP/UgcgKO as NesUgcgKO mice.
The major class of glycosphingolipids in brain—theganglio-series gangliosides—are dependent on theexpression of Ugcg (Fig. 2b) (Kolter et al., 2002). Wetherefore determined the level of gangliosides in brainsof NesUgcgKO mice of different ages compared withage-matched control mice. We found that a qualitativelysimilar pattern of ganglioside bands corresponding toGM1, GD1a, GD1b, GT1b was present at all ages tested(Fig. 2b,c; data not shown). However, the total amountof these gangliosides in the brains of the NesUgcgKOmice decreased substantially, similar to the reduction ofUgcg mRNA expression. The decrease in gangliosidecontent was relatively constant among the mice of differ-ent ages: 67% decrease at 2 weeks, 76% decrease at2 months, and 75% decrease at 4 months.
NesUgcgKO mice appeared normal and healthy atbirth and immediately thereafter. However, by 2–3 monthsof age they exhibited an abnormal clasping behaviorwhen suspended by their tail (Fig. 3a). This response hasbeen described in several kinds of neurologicallyimpaired mutant mice (Auerbach et al., 2001; Guptaet al., 2001; Miklyaeva et al., 2004). As the mice aged,they also displayed a labored gait. Histologic evaluationof the brains from NesUgcgKO mice revealed a strikingloss of Purkinje cells (Fig. 3b) that increased in severityas the mice aged.
The early lethality caused by the global disruption ofthe Ugcg gene and concomitant block of glycosphingoli-pid synthesis prompted us to establish mice with a func-tional floxed Ugcg allele. Here we show that the floxedUgcg allele can be conditionally disrupted in the nervoussystem by expressing cre recombinase. NesUgcgKOmice showed substantially reduced levels of brain gan-gliosides and an abnormal neurologic phenotype, to-gether with a profound loss of Purkinje cells. The pheno-type mirrors that of ganglioside-deficient mice, whichshow a profound neurodegenerative phenotype (Yama-shita et al., 2005). The milder phenotype of theNesUgcgKO mice might be the result of residual Ugcgexpression present in the NesUgcgKO brain or perhapsthe transfer of glycosphingolipids shed from cells withnormal Ugcg expression (Olshefski and Ladisch, 1998).These mice carrying floxed Ugcg alleles will be usefulfor delineating the functional roles of glycosphingolipid
FIG. 3. Neurologic phenotype in NesUgcgKO mice. a: A 6-month-old NesUgcgKO mouse showing abnormal clasping behavior when sus-pended by the tail. b: Sections from the cerebellar region of brain stained by H&E (left) and immunostained with calbindin antibody (right)showing a severe loss of Purkinje cells (arrows) in a 9-month-old NesUgcgKO mouse (bottom) compared with an age-matched control (top).Magnification3200.
178 YAMASHITA ET AL.

synthesis in the nervous system and in other physiologicsystems.
MATERIALS AND METHODS
Targeting and Mice
A genomic clone containing the Ugcg locus was isolatedfrom a 129-mouse library (Genome Systems, St. Louis,MO) using a mouse Ugcg cDNA probe. An 8.5-kb BglIIfragment containing exons 6, 7, 8, 9, and the 30 untrans-lated region was used to construct the pUgcgloxPneotargeting vector (Fig. 1a) using the ploxPneo plasmid asa cloning vehicle (Yang et al., 1998). A loxP site wasinserted into intron 6 and loxP sites surrounding the neocassette were placed into the 30 untranslated region. ThepUgcgloxPneo vector was linearized and electroporatedinto TC-1 ES cells as described (Liu et al., 1999) to targetthe Ugcg allele. Clones resistant to ganciclovir and G418were screened by Southern blot analysis with a probecorresponding to exon 5 of Ugcg. The targeting proce-dure gave about a 3% targeting frequency. One of the tar-geted clones was used to establish chimeric males,which were bred to C57BL/6 females. Genotyping wasaccomplished by PCR of tail DNA using primer1 (50-ATGTGCTAGATCAGGCAGGAGGGCTCATAG-30) and pri-mer2 (50-CCAACAGATATTGAATGCGAATGCTCTGCC-30)(45 cycles of 948C for 1 min, 688C for 1 min, and 728Cfor 1 min). The wildtype allele yielded a DNA fragmentof about 200 bp and the targeted allele yielded a frag-ment of �250 bp.
Transgenic mice carrying the nestin-cre transgene(Tronche et al., 1999) (Strain Name: B6.Cg-Tg(Nes-cre)1Kln/J; Stock Number: 003771) were obtained fromThe Jackson Laboratory (Bar Harbor, ME). The globalUgcg KO mice carrying the UgcgKO allele have beendescribed (Yamashita et al., 1999). Cre-mediated recom-bination of the Ugcg allele was identified by Southernblot analysis of SacI-digested genomic DNA with a 50probe external to the sequences contained within thetargeting vector. Recombination was also detected byPCR of genomic DNA using primers 1, 2, and 3 (50-GAGCCAGTCCATTACTCTCGTTGATTGCAT-30) (Fig. 1a,c).Primers 1 and 2 detected the UgcgloxP allele and primers1 and 3 detected the UgcgD allele (30 cycles of 948C for1 min, 688C for 1 min, and 728C for 1 min).
Analysis of Ugcg Expression and theResultant Phenotype
For real-time PCR, total RNA was purified using TRIZOL(Invitrogen, Carlsbad, CA). Mouse UgcgmRNAwas quanti-fied using Assays-on-Demand reagents (Applied Biosys-tems, Foster City, CA) with an ABI Prism 7700 SequenceDetection System (Applied Biosystems). Glyceraldehyde 3-phosphate dehydrogenase mRNA was quantified as aninternal standard. Acidic gangliosides from the brain wereisolated and subjected to analysis by thin-layer chromatog-raphy as described (Yamashita et al., 2005). For quan-tification of the lipids, the thin-layer plates were developed
and scanned with a densitometer (CS-9301PC; Shimadzu,Kyoto, Japan). For histology, brains were fixed, processedfor paraffin embedding, and cut into 5-l sections. The sec-tions were stained with hematoxylin-eosin (H&E) asdescribed (Wu and Proia, 2004). To identify calbindin-posi-tive cells, paraffin sections were reacted with rabbit anti-calbindin D-28K affinity purified antibody (1:50, ChemiconInternational, Temecula, CA; AB1778) and then reactedwith a goat antirabbit IgG labeled with AlexaFluor 594(1:1000, Molecular Probes, Eugene, OR; A-11037) asdescribed (Wu and Proia, 2004). The sections weremounted and analyzed using a Leica DMRmicroscope.
LITERATURE CITED
Allende ML, Proia RL. 2002. Lubricating cell signaling pathways withgangliosides. Curr Opin Struct Biol 12:587–592.
Auerbach W, Hurlbert MS, Hilditch-Maguire P, Wadghiri YZ, WheelerVC, Cohen SI, Joyner AL, MacDonald ME, Turnbull DH. 2001. TheHD mutation causes progressive lethal neurological disease inmice expressing reduced levels of huntingtin. Hum Mol Genet10:2515–2523.
Chiavegatto S, Sun J, Nelson RJ, Schnaar RL. 2000. A functional role forcomplex gangliosides: motor deficits in GM2/GD2 synthase knock-out mice. Exp Neurol 166:227–234.
Degroote S, Wolthoorn J, van Meer G. 2004. The cell biology of glyco-sphingolipids. Semin Cell Dev Biol 15:375–387.
Gupta P, Soyombo AA, Atashband A, Wisniewski KE, Shelton JM,Richardson JA, Hammer RE, Hofmann SL. 2001. Disruption ofPPT1 or PPT2 causes neuronal ceroid lipofuscinosis in knockoutmice. Proc Natl Acad Sci U S A 98:13566–13571.
Ichikawa S, Sakiyama H, Suzuki G, Hidari KI, Hirabayashi Y. 1996.Expression cloning of a cDNA for human ceramide glucosyltrans-ferase that catalyzes the first glycosylation step of glycosphingoli-pid synthesis. Proc Natl Acad Sci U S A 93:4638–4643.
Inoue M, Fujii Y, Furukawa K, Okada M, Okumura K, Hayakawa T,Sugiura Y. 2002. Refractory skin injury in the complex knock-outmice expressing only GM3 ganglioside. J Biol Chem 277:29881–29888.
Kakugawa Y, Wada T, Yamaguchi K, Yamanami H, Ouchi K, Sato I,Miyagi T. 2002. Up-regulation of plasma membrane-associated gan-glioside sialidase (Neu3) in human colon cancer and its involve-ment in apoptosis suppression. Proc Natl Acad Sci U S A 99:10718–10723.
Kawai H, Allende ML, Wada R, Kono M, Sango K, Deng C, Miyakawa T,Crawley JN, Werth N, Bierfreund U, Sandhoff K, Proia RL. 2001.Mice expressing only monosialoganglioside GM3 exhibit lethalaudiogenic seizures. J Biol Chem 276:6885–6888.
Kolter T, Proia RL, Sandhoff K. 2002. Combinatorial ganglioside biosyn-thesis. J Biol Chem 277:25859–25862.
Liu Y, Wada R, Kawai H, Sango K, Deng C, Tai T, McDonald MP, AraujoK, Crawley JN, Bierfreund U, et al. 1999. A genetic model of sub-strate deprivation therapy for a glycosphingolipid storage disorder.J Clin Invest 103:497–505.
Miklyaeva EI, Dong W, Bureau A, Fattahie R, Xu Y, Su M, Fick GH,Huang JQ, Igdoura S, Hanai N, Gravel RA. 2004. Late onset Tay-Sachs disease in mice with targeted disruption of the Hexa gene:behavioral changes and pathology of the central nervous system.Brain Res 1001:37–50.
Olshefski R, Ladisch S. 1998. Synthesis, shedding, and intercellular trans-fer of human medulloblastoma gangliosides: abrogation by a newinhibitor of glucosylceramide synthase. J Neurochem 70:467–472.
Sandhoff R, Geyer R, Jennemann R, Paret C, Kiss E, Yamashita T, Gorgas K,Sijmonsma TP, Iwamori M, Finaz C, et al. 2005. Novel class of glyco-sphingolipids involved in male fertility. J Biol Chem 280:27310–27318.
Sheikh KA, Sun J, Liu Y, Kawai H, Crawford TO, Proia RL, Griffin JW,Schnaar RL. 1999. Mice lacking complex gangliosides developWaller-ian degeneration and myelination defects. Proc Natl Acad Sci U S A96:7532–7537.
179CONDITIONAL LOXP-FLANKED GLUCOSYLCERAMIDE

Tagami S, Inokuchi J, Kabayama K, Yoshimura H, Kitamura F, Uemura S,Ogawa C, Ishii A, Saito M, Ohtsuka Y, et al. 2002. GangliosideGM3 participates in the pathological conditions of insulin resist-ance. J Biol Chem 277:3085–3092.
Takamiya K, Yamamoto A, Furukawa K, Yamashiro S, Shin M, Okada M,Fukumoto S, Haraguchi M, Takeda N, Fujimura K, et al. 1996. Micewith disrupted GM2/GD2 synthase gene lack complex ganglio-sides but exhibit only subtle defects in their nervous system. ProcNatl Acad Sci U S A 93:10662–10667.
Tronche F, Kellendonk C, Kretz O, Gass P, Anlag K, Orban PC, Bock R,Klein R, Schutz G. 1999. Disruption of the glucocorticoid receptorgene in the nervous system results in reduced anxiety. Nat Genet23:99–103.
Wu YP, Proia RL. 2004. Deletion of macrophage-inflammatory protein 1alpha retards neurodegeneration in Sandhoff disease mice. ProcNatl Acad Sci U S A 101:8425–8430.
Yamashita T, Wada R, Sasaki T, Deng C, Bierfreund U, Sandhoff K, ProiaRL. 1999. A vital role for glycosphingolipid synthesis during devel-opment and differentiation. Proc Natl Acad Sci U S A 96:9142–9147.
Yamashita T, Wada R, Proia RL. 2002. Early developmental expressionof the gene encoding glucosylceramide synthase, the enzyme con-trolling the first committed step of glycosphingolipid synthesis.Biochim Biophys Acta 1573:236–240.
Yamashita T, Hashiramoto A, Haluzik M, Mizukami H, Beck S, Norton A,Kono M, Tsuji S, Daniotti JL, Werth N, et al. 2003. Enhanced insu-lin sensitivity in mice lacking ganglioside GM3. Proc Natl Acad SciU S A 100:3445–3449.
Yamashita T, Wu YP, Sandhoff R, Werth N, Mizukami H, Ellis JM, DupreeJL, Geyer R, Sandhoff K, Proia RL. 2005. Interruption of ganglio-side synthesis produces central nervous system degeneration andaltered axon-glial interactions. Proc Natl Acad Sci U S A 102:2725–2730.
Yang X, Li C, Xu X, Deng C. 1998. The tumor suppressor SMAD4/DPC4is essential for epiblast proliferation and mesoderm induction inmice. Proc Natl Acad Sci U S A 95:3667–3672.
Zhou D, Mattner J, Cantu C 3rd, Schrantz N, Yin N, Gao Y, Sagiv Y, Hud-speth K, Wu YP, Yamashita T, et al. 2004. Lysosomal glycosphingo-lipid recognition by NKT cells. Science 306:1786–1789.
180 YAMASHITA ET AL.