Embed Size (px)
Citation preview

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, May 1994, p. 1538-1546 Vol. 60, No. 50099-2240/94/$04.00+0
Compositional Changes in Compost during Composting andGrowth of Agaricus bisporus
KENJI IIYAMA,'* BRUCE A. STONE,1 AND BARRY J. MACAULEY2Departments of Biochemistry' and Microbiology,2La Trobe University, Bundoora, Victoria 3083, Australia
Received 12 November 1993/Accepted 2 March 1994
Samples from conventional compost taken at various stages of composting and mushroom (Agaricusbisporus) growth were analyzed for changes in 80% ethanol and water extracts, monosaccharides in acidhydrolysates of polysaccharides, lignin concentration, and lignin structural features. Variable amounts ofextraneous inorganic solids in the form of fine sandy particles were removed by sedimentation of the samplesin a carbon tetrachloride-dibromomethane mixture. During composting, about two-thirds of the initial wallpolysaccharides were consumed by compost microorganisms, and only 17% of the total polysaccharides wereused during mushroom production. The relative lignin content of composts as measured by the acetyl bromideprocedure increased, both during composting and mushroom growth, and the chemical structure of lignin wasaltered by condensation and oxidation reactions.
Conventional mushroom compost is produced from a mix-ture of wheat straw (40 to 45% total dry weight), stablebedding (20 to 25%), poultry manure (10 to 15%) and gypsum(5 to 10%) with varying amounts (10 to 15%) of optionalingredients, such as brewer's grain, seed meals, cotton hulls,and other agricultural wastes. After thorough mixing, theingredients are generally prewetted in low piles for 3 to 10 daysand periodically mixed. The next stage (phase I) is the forma-tion of this mix, generally in the open, into long rectangularstacks, which are turned and reformed, with wetting whennecessary, at regular intervals over 7 to 14 days. The finalprocessing stage (phase II) is conducted in an indoor chamberor room in which the compost is "pasteurized" (at about 60°C)and conditioned (45 to 50°C) over a 6- to 9-day period (10, 46).During composting, a succession of fungi and bacteria,
including actinomycetes, chemically alter the compost constit-uents, resulting in a material that provides a nutrient sourcewhich promotes rapid growth of the mushroom mycelium. Inaddition, the microbial population appears to confer selectiveproperties on the compost by inhibiting the development ofcompeting fungi and bacteria that could seriously reduce themushroom crop (10).
Since the chemical composition of compost is directly re-lated to mushroom growth, and hence mushroom production,it is important to determine accurately the compositionalchanges during composting and mushroom growth. Because ofthe diversity in the composition of its ingredients, compost ischemically quite complicated. In addition, various amounts ofsoil, sand, and stone are found in commercial compost mix-tures and are partly responsible for the high content of ash.Variation in the amounts of these adventitious inorganic solidsduring composting can give misleading compositional valuesbased on dry weight.
There is general agreement that both cellulose and noncel-lulosic polysaccharides of straw are substrates for mushroomgrowth (9, 13, 14, 29, 58-60). The situation is not so clear forlignin, the second most abundant organic compost component.Falck (9) reported an enrichment of lignin during mushroom
* Corresponding author. Mailing address: Department of Biochem-istry, La Trobe University, Bundoora, Vic. 3083, Australia. Phone:61-3-479-2159. Fax: 61-3-479-2467.
growth, but Waksman and his coworkers (58-60), Iwade (29),Gerrits and coworkers (13, 14), Muller (47), and Wood andcoworkers (7, 61, 62) all produced evidence interpreted asindicating a loss of lignin during mushroom growth on com-post. Consequently, it is important to decide unequivocallywhether lignin is effectively utilized as a nutrient by Agaricusbisporus.
In recent years, there have been many improvements inprocedures for the analysis of plant components (3, 5, 22, 26,39, 51) and for a greater understanding of the organization ofthe structural polymers in wheat straw cell walls (39, 51), whichare quantitatively the most important plant constituents incompost mixtures. We have used these improvements toestablish analytical procedures to reinvestigate changes incompost constituents during composting and cropping. Aprocedure for removing adventitious sand and stone fromcompost has been devised, and a fractionation scheme for thevarious major classes of organic components in composts hasbeen developed. These procedures have been applied tomeasure changes in the organic components in conventionalcomposts.
MATERIALS AND METHODS
Compost. Two conventional compost samples and theirinitial ingredients were obtained from commercial farms inVictoria, Australia. One, from Melbourne Mushrooms, wassampled on one day during phase I, at the end of phase II ofcomposting, and at the end of cropping (mushroom yield, 22.8kg/m2 [259 kg/metric ton of wet compost]) (Fig. 1). A secondsequence of 22 samples was obtained during conventionalcomposting and mushroom cropping at Dandy Mushrooms inthe spring of 1991 (see the horizontal scale of Fig. 4) (mush-room yield, 23.5 kg/m2 [263 kg/metric ton of wet compost]).There were no substantial differences in the composition ofcompost ingredients and composting schedule for productionof compost between Melbourne Mushrooms and Dandy Mush-rooms. Samples (about 1 kg [wet weight]) were immediatelyfrozen and later freeze-dried. These samples (about 100 g)were milled in a Wiley mill to pass through a 420-,um sieve.
Fractionation of samples. The freeze-dried, milled sample(about 50 g [oven-dry weight]) was successively extracted withboiling diethyl ether (1 h; three times), with boiling 80%
1538
on January 4, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from

COMPOSITIONAL CHANGES IN COMPOST FOR AGARICUS BISPORUS 1539
Filling of trays
Pasteurising
Conditioning
Cooldown
Spa
Ca
Sanpling(Raw materials)
Phase Icomposting(- 2 weeks)
Sampling(Phase I compost)
Phase II
composting(- 1 week)
Samplingif
(Phase I1 compost)Iwning Mycelium
developnentasing (- 4 weeks)
IPinning
Picking
Mushroomproduction(- 6 weeks)
Cookout
Spent Sampling(Spent compost
FIG. 1. Sampling schedule for conventional stack composting andmushroom growth.
(vol/vol) aqueous ethanol (1 h; three times), and then withwater at 40°C, overnight, with shaking in the presence oftoluene. After each extraction, the sample was recovered byfiltration. Both the pooled 80% aqueous ethanol and waterextracts were concentrated by vacuum evaporation, freeze-dried, and weighed. The dried material was suspended byvigorous stirring in a mixture of carbon tetrachloride (d25 =
1.59) and dibromomethane (d25 = 2.50) (1:1, vol/vol; specificgravity, about 2.0), allowing inorganic solids (sand and frag-ments of stones, bricks, concrete, and metal) to settle out,before being separated from the floating organic fraction bydecantation. The organic fraction was dried in a vacuum oven
at 40°C before chemical analysis. The fractionation scheme isshown in Fig. 2.Another part of the original sample (about 25 g) was
extracted by a neutral detergent extraction procedure (57).The extracted sample was washed with ethanol and dried in a
vacuum oven at 40°C.Chemical analyses. The neutral sugar composition of acid
hydrolysates was determined by an alditol acetate procedure(5). Lignin content was determined by an acetyl bromideprocedure (26), with the spectrum of each digested samplerecorded between 250 to 400 nm. In addition, a Klason sulfuricacid digestion procedure, following the standardized proce-dure for determining lignin content in wood and pulps, was
used (2). In this procedure, the dry sample (0.5 g) was treatedwith 72% sulfuric acid (10 ml) for 4 h at room temperaturewith occasional stirring; the mixture was diluted with water(372 ml) to make a 3% sulfuric acid solution and refluxed for2 h. After cooling, the residue was filtered and washed with hot
Sample (stored at -30°C)
1. Freeze dry2.VViey rrill (<420Qjm)3. Extract, 3x1.5h boiling diethyl ether
Ether extract Residue
Extract, 3X1h boiling 80% ethanoi
80% ethanol Residueextract Extract, water 24h, 40°C
Water extract ResidueWash with ethanolDry at 400C in vacuoSeparate withCCI4-CH2Br2(1:1 v/v, s.g.2.0)
Sediment Floatingmaterial
Dry at 400C in vacuo
Extractedsample
Chemical analysis(for ash, nitrogen, silicalignin, polysaccharides)
FIG. 2. Compost fractionation scheme.
water through a porous crucible (G4), dried to constant weightat 105°C, and weighed as acid-insoluble residue. The residuewas combusted for 3 h at 7000C to determine the amount ofacid-insoluble ash. The filtrate and washings were adjusted to500 ml, and the UV spectrum was recorded by using a similarlyprepared sulfuric acid blank as a reference. Acid-soluble lignincontent was determined by using the value of 110 g-- litercm- for the specific A205 coefficient (48-50, 52), at which theabsorption by furan aldehydes originating from carbohydrateshas only a small effect.The nitrogen content of extracted samples was measured by
microscale elemental analysis using a combustion procedure.Ash content was determined by combustion at 700°C for 3 h.Silica content was determined gravimetrically after boiling theash in 2 M HCl for 1 h. The silica remained as an insolubleresidue. Structural changes in lignin were monitored by mea-
suring the products formed by alkaline nitrobenzene oxidationas trimethylsilyl derivatives by gas chromatography (22). All ofthe determinations described above were made on triplicatesamples.
Consistency of sampling. An experiment was conducted toensure that each sample was truly representative of a particularstage of the composting process and to determine whetherconsistent samples could be collected from stacks. Sampleswere obtained from two stacks (approximately 1.8 by 1.8 by 100m) at the same stage of processing at the compost facility ofMelbourne Mushrooms at Nagambie, Victoria, Australia. Twoprotocols were followed.
(i) Procedure A. Ten samples, each 6 to 7 kg, were taken atregular distances along one stack by digging in from the top ofthe stack to a depth of about 0.5 m. These samples were bulked(total weight, about 64 kg), mixed well with a front-end loader,and then divided into four approximately equal parts. One-kilogram subsamples (subgroup I) were taken from three ofthe segments, while the fourth segment (approximately 16 kg)
Compost ingredient
Premix ingredients
Stacking of heaps
Turning of heaps
VOL. 60, 1994
I
on January 4, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from
APPL. ENVIRON. MICROBIOL.
TABLE 1. Changes in solvent extracts and inorganic materials during composting and mushroom growth
% Oven-dry matter
Extraction solvent Inorganic materialSample type_______________________________
Neutral Diethyl 80% Ethanol H20 Sand and Ashdetergent ether stone
Compost ingredientsWheat straw 26.2 0.2 5.9 0.9 3.2 3.2Stable bedding 31.8 1.4 11.3 6.5 7.2 5.1Poultry manure 48.5 3.3 10.9 5.7 7.0 7.1Rice hulls 47.2 3.0 12.2 8.5 14.3 10.2Cotton hulls 25.0 3.3 17.9 3.3 15.6 12.4Cotton seed 74.5 3.2 12.1 3.7 14.0 12.9
Conventional compostPhase I compost 33.6 0.5 6.3 3.8 5.1 5.9Phase II compost 46.4 0.5 5.4 5.0 19.0 9.6Spent compost 48.2 0.3 7.7 6.2 18.0 9.6
was mixed again. This 16-kg sample was divided into foursegments, and the sampling procedure was repeated by taking1 kg from each of three segments (subgroup II), the fourthsegment being remixed and divided to give 4 1-kg samples(subgroup III), giving a total of 10 samples.
(ii) Procedure B. With a front-end loader, the end was
knocked off a second stack and a segment of the total profilewas taken and mixed. A composite sample of about 64 kg was
isolated, and subsamples were collected as described in proce-dure A.
Five analytical parameters were measured in the 20 sub-samples to allow an estimation of variance between samples.These were 80% ethanol extracts, H20 extracts, inorganicsolids, ash content (after removal of inorganic solids), andlignin content.
RESULTSAnalytical procedures were first evaluated with compost
from Melbourne Mushrooms, and selected methods were thenapplied to 22 sequential samples from Dandy Mushrooms.
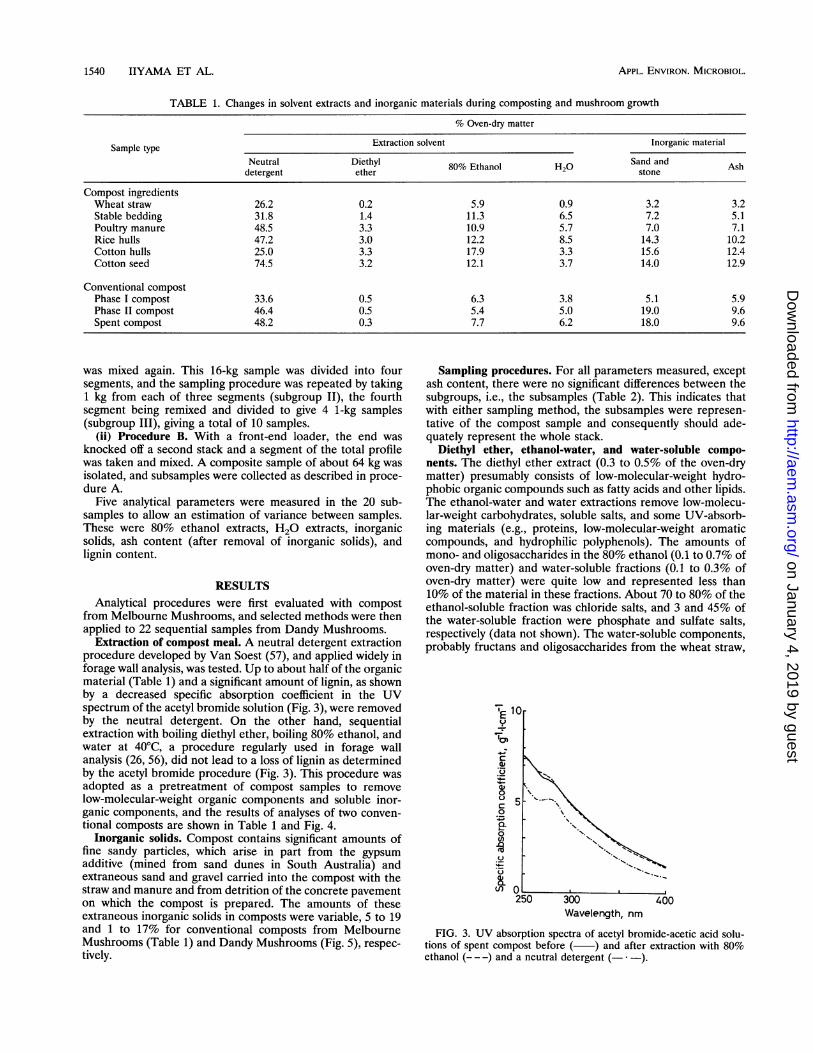
Extraction of compost meal. A neutral detergent extractionprocedure developed by Van Soest (57), and applied widely inforage wall analysis, was tested. Up to about half of the organicmaterial (Table 1) and a significant amount of lignin, as shownby a decreased specific absorption coefficient in the UVspectrum of the acetyl bromide solution (Fig. 3), were removedby the neutral detergent. On the other hand, sequentialextraction with boiling diethyl ether, boiling 80% ethanol, andwater at 40°C, a procedure regularly used in forage wallanalysis (26, 56), did not lead to a loss of lignin as determinedby the acetyl bromide procedure (Fig. 3). This procedure was
adopted as a pretreatment of compost samples to remove
low-molecular-weight organic components and soluble inor-ganic components, and the results of analyses of two conven-
tional composts are shown in Table 1 and Fig. 4.Inorganic solids. Compost contains significant amounts of
fine sandy particles, which arise in part from the gypsum
additive (mined from sand dunes in South Australia) andextraneous sand and gravel carried into the compost with thestraw and manure and from detrition of the concrete pavementon which the compost is prepared. The amounts of theseextraneous inorganic solids in composts were variable, 5 to 19and 1 to 17% for conventional composts from MelbourneMushrooms (Table 1) and Dandy Mushrooms (Fig. 5), respec-
tively.
Sampling procedures. For all parameters measured, exceptash content, there were no significant differences between thesubgroups, i.e., the subsamples (Table 2). This indicates thatwith either sampling method, the subsamples were represen-tative of the compost sample and consequently should ade-quately represent the whole stack.
Diethyl ether, ethanol-water, and water-soluble compo-nents. The diethyl ether extract (0.3 to 0.5% of the oven-drymatter) presumably consists of low-molecular-weight hydro-phobic organic compounds such as fatty acids and other lipids.The ethanol-water and water extractions remove low-molecu-lar-weight carbohydrates, soluble salts, and some UV-absorb-ing materials (e.g., proteins, low-molecular-weight aromaticcompounds, and hydrophilic polyphenols). The amounts ofmono- and oligosaccharides in the 80% ethanol (0.1 to 0.7% ofoven-dry matter) and water-soluble fractions (0.1 to 0.3% ofoven-dry matter) were quite low and represented less than10% of the material in these fractions. About 70 to 80% of theethanol-soluble fraction was chloride salts, and 3 and 45% ofthe water-soluble fraction were phosphate and sulfate salts,respectively (data not shown). The water-soluble components,probably fructans and oligosaccharides from the wheat straw,
E 10U
'c
0)C
1=u 5 X0
0;a \
0.250 300 400
Wavelength, nm
FIG. 3. UV absorption spectra of acetyl bromide-acetic acid solu-tions of spent compost before ( ) and after extraction with 80%ethanol (--- ) and a neutral detergent (- ).
1540 IIYAMA ET AL.
on January 4, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from
COMPOSITIONAL CHANGES IN COMPOST FOR AGARICUS BISPORUS
cm
E C CM1 wUv-c U) 0v
1620Aug
IL I i, Yi0 l 0
ei c'0IM Il I
0o
30 10 20 30 10 20Sep Oct
1991
30 10Nov
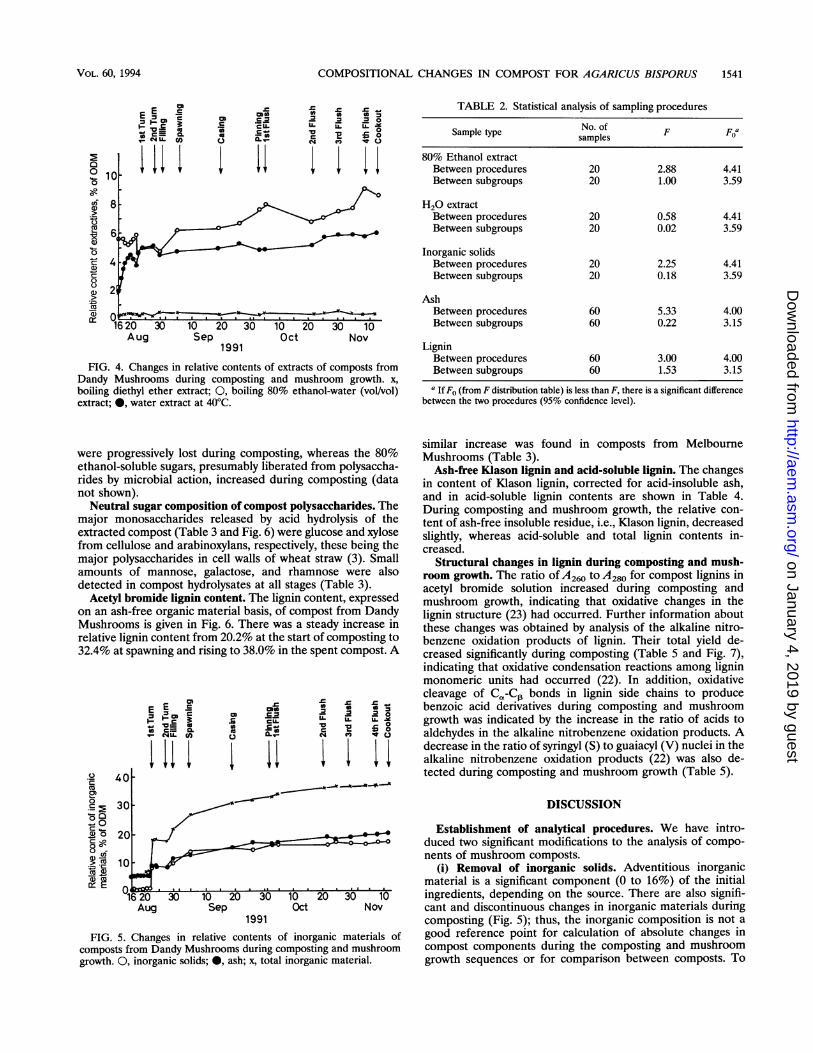
FIG. 4. Changes in relative contents of extracts of composts fromDandy Mushrooms during composting and mushroom growth. x,boiling diethyl ether extract; 0, boiling 80% ethanol-water (vol/vol)extract; 0, water extract at 40°C.
were progressively lost during composting, whereas the 80%ethanol-soluble sugars, presumably liberated from polysaccha-rides by microbial action, increased during composting (datanot shown).
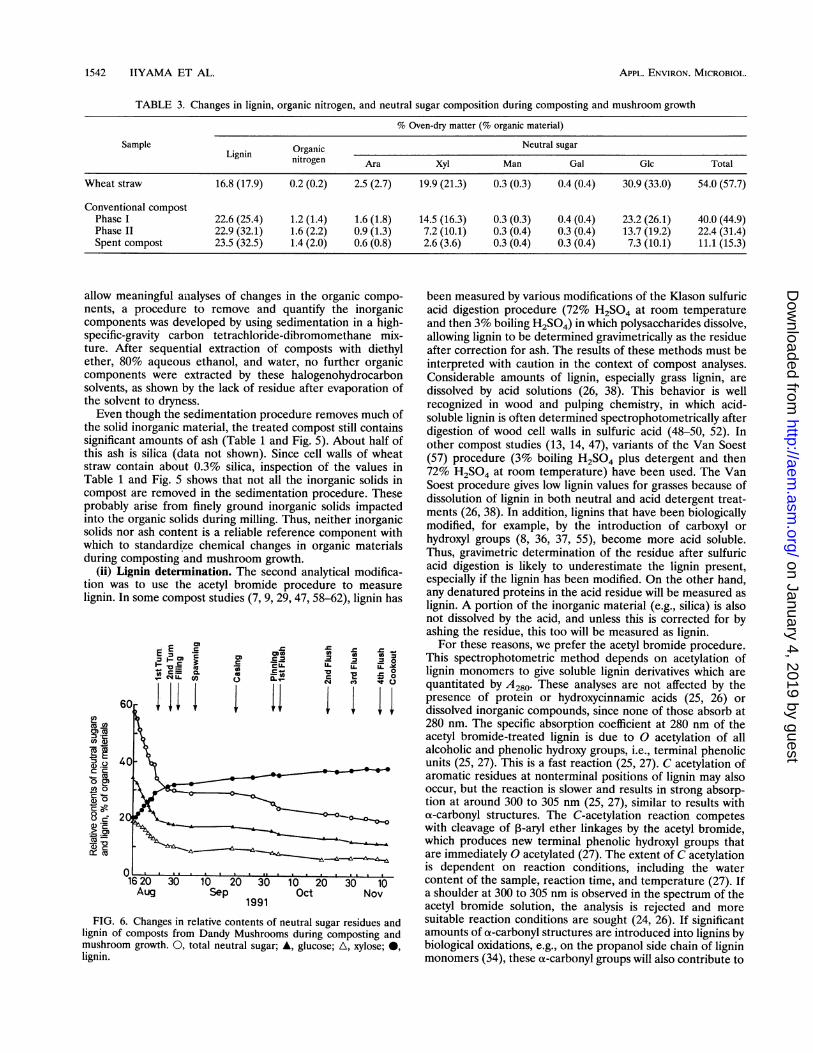
Neutral sugar composition of compost polysaccharides. Themajor monosaccharides released by acid hydrolysis of theextracted compost (Table 3 and Fig. 6) were glucose and xylosefrom cellulose and arabinoxylans, respectively, these being themajor polysaccharides in cell walls of wheat straw (3). Smallamounts of mannose, galactose, and rhamnose were alsodetected in compost hydrolysates at all stages (Table 3).
Acetyl bromide lignin content. The lignin content, expressedon an ash-free organic material basis, of compost from DandyMushrooms is given in Fig. 6. There was a steady increase inrelative lignin content from 20.2% at the start of composting to32.4% at spawning and rising to 38.0% in the spent compost. A
40
30 -
20 - = t
10 ;0s 2 . . . .. ..s L, 4^ ^ ^^ 4 ^^t~% 4^
EEE 3 o
I.F-r 3: CU.
C = .'tVI' cm U. CD 0 z
1111 1 1
16 20 3 1U ZU 30Aug Sep
1991
".
0
C 2E -5
C.,-e
0.
1i 20 1UOct Nov
FIG. 5. Changes in relative contents of inorganic materials ofcomposts from Dandy Mushrooms during composting and mushroomgrowth. 0, inorganic solids; 0, ash; x, total inorganic material.
TABLE 2. Statistical analysis of sampling procedures
No.ofaSample type samples F Fo
80% Ethanol extractBetween procedures 20 2.88 4.41Between subgroups 20 1.00 3.59
H20 extractBetween procedures 20 0.58 4.41Between subgroups 20 0.02 3.59
Inorganic solidsBetween procedures 20 2.25 4.41Between subgroups 20 0.18 3.59
AshBetween procedures 60 5.33 4.00Between subgroups 60 0.22 3.15
LigninBetween procedures 60 3.00 4.00Between subgroups 60 1.53 3.15
a If Fo (from F distribution table) is less than F, there is a significant differencebetween the two procedures (95% confidence level).
similar increase was found in composts from MelbourneMushrooms (Table 3).
Ash-free Klason lignin and acid-soluble lignin. The changesin content of Klason lignin, corrected for acid-insoluble ash,and in acid-soluble lignin contents are shown in Table 4.During composting and mushroom growth, the relative con-tent of ash-free insoluble residue, i.e., Klason lignin, decreasedslightly, whereas acid-soluble and total lignin contents in-creased.
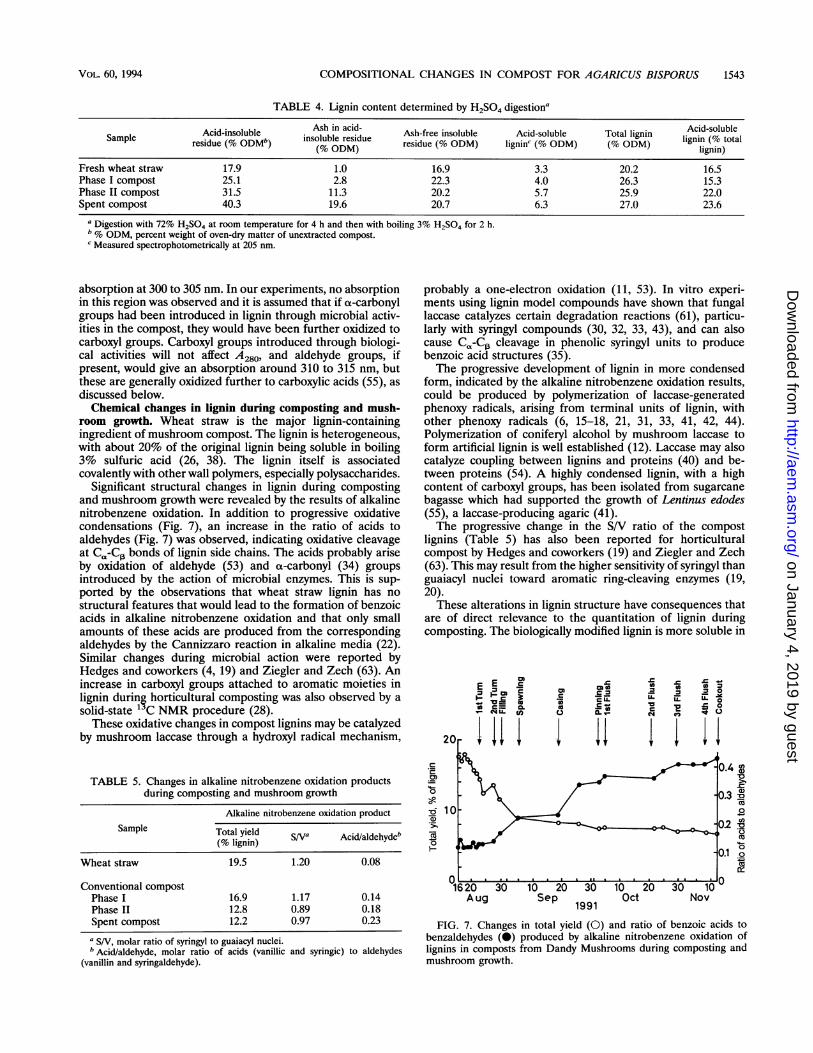
Structural changes in lignin during composting and mush-room growth. The ratio ofA260 to A280 for compost lignins inacetyl bromide solution increased during composting andmushroom growth, indicating that oxidative changes in thelignin structure (23) had occurred. Further information aboutthese changes was obtained by analysis of the alkaline nitro-benzene oxidation products of lignin. Their total yield de-creased significantly during composting (Table 5 and Fig. 7),indicating that oxidative condensation reactions among ligninmonomeric units had occurred (22). In addition, oxidativecleavage of Ca-C13 bonds in lignin side chains to producebenzoic acid derivatives during composting and mushroomgrowth was indicated by the increase in the ratio of acids toaldehydes in the alkaline nitrobenzene oxidation products. Adecrease in the ratio of syringyl (S) to guaiacyl (V) nuclei in thealkaline nitrobenzene oxidation products (22) was also de-tected during composting and mushroom growth (Table 5).
DISCUSSION
Establishment of analytical procedures. We have intro-duced two significant modifications to the analysis of compo-nents of mushroom composts.
(i) Removal of inorganic solids. Adventitious inorganicmaterial is a significant component (0 to 16%) of the initialingredients, depending on the source. There are also signifi-cant and discontinuous changes in inorganic materials duringcomposting (Fig. 5); thus, the inorganic composition is not agood reference point for calculation of absolute changes incompost components during the composting and mushroomgrowth sequences or for comparison between composts. To
10000
of
4-)0
cni
C
a)C4._0V
TI)
8
6
4
21
.0co
0,=0)o- O0
C4-028a cn
a) X
--Q -.-.- . . . . .
VOL. 60, 1994 1541
on January 4, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from
1542 IIYAMA ET AL.
TABLE 3. Changes in lignin, organic nitrogen, and neutral sugar composition during composting and mushroom growth
% Oven-dry matter (% organic material)
Sample Organic Neutral sugarLignin nitrogen Ara Xyl Man Gal Glc Total
Wheat straw 16.8 (17.9) 0.2 (0.2) 2.5 (2.7) 19.9 (21.3) 0.3 (0.3) 0.4 (0.4) 30.9 (33.0) 54.0 (57.7)
Conventional compostPhase I 22.6 (25.4) 1.2 (1.4) 1.6 (1.8) 14.5 (16.3) 0.3 (0.3) 0.4 (0.4) 23.2 (26.1) 40.0 (44.9)Phase II 22.9 (32.1) 1.6 (2.2) 0.9 (1.3) 7.2 (10.1) 0.3 (0.4) 0.3 (0.4) 13.7 (19.2) 22.4 (31.4)Spent compost 23.5 (32.5) 1.4 (2.0) 0.6 (0.8) 2.6 (3.6) 0.3 (0.4) 0.3 (0.4) 7.3 (10.1) 11.1 (15.3)
allow meaningful analyses of changes innents, a procedure to remove and qu
components was developed by using sedispecific-gravity carbon tetrachloride-ditture. After sequential extraction of co
ether, 80% aqueous ethanol, and watercomponents were extracted by these hsolvents, as shown by the lack of residuethe solvent to dryness.Even though the sedimentation procedi
the solid inorganic material, the treated c
significant amounts of ash (Table 1 and Ithis ash is silica (data not shown). Sincestraw contain about 0.3% silica, inspectTable 1 and Fig. 5 shows that not all thcompost are removed in the sedimentatiprobably arise from finely ground inorginto the organic solids during milling. Thsolids nor ash content is a reliable referewhich to standardize chemical changesduring composting and mushroom growt
(ii) Lignin determination. The secondtion was to use the acetyl bromide pr
lignin. In some compost studies (7, 9, 29,
cn X
:3 .C
g-Caa)._ C.)
a)E
0cm
00)0
IL)0
C
e0'Dc
E E ,
1 1 1
Aug Sep1991
FIG. 6. Changes in relative contents of net
lignin of composts from Dandy Mushroomsmushroom growth. 0, total neutral sugar; A,
lignin.
the organic compo- been measured by various modifications of the Klason sulfuricantify the inorganic acid digestion procedure (72% H2SO4 at room temperatureimentation in a high- and then 3% boiling H2SO4) in which polysaccharides dissolve,)romomethane mix- allowing lignin to be determined gravimetrically as the residuemposts with diethyl after correction for ash. The results of these methods must be, no further organic interpreted with caution in the context of compost analyses.ialogenohydrocarbon Considerable amounts of lignin, especially grass lignin, areafter evaporation of dissolved by acid solutions (26, 38). This behavior is well
recognized in wood and pulping chemistry, in which acid-ure removes much of soluble lignin is often determined spectrophotometrically afterompost still contains digestion of wood cell walls in sulfuric acid (48-50, 52). InFig. 5). About half of other compost studies (13, 14, 47), variants of the Van Soesta cell walls of wheat (57) procedure (3% boiling H2SO4 plus detergent and thention of the values in 72% H2SO4 at room temperature) have been used. The Vane inorganic solids in Soest procedure gives low lignin values for grasses because ofon procedure. These dissolution of lignin in both neutral and acid detergent treat-,anic solids impacted ments (26, 38). In addition, lignins that have been biologically
Ius, neither inorganic modified, for example, by the introduction of carboxyl or3nce component with hydroxyl groups (8, 36, 37, 55), become more acid soluble.in organic materials Thus, gravimetric determination of the residue after sulfuric
* y. acid digestion is likely to underestimate the lignin present,oanalytcal modefica especially if the lignin has been modified. On the other hand,
58-62), lignin has any denatured proteins in the acid residue will be measured as47'5862lignnhas lignin. A portion of the inorganic material (e.g., silica) is also
not dissolved by the acid, and unless this is corrected for byashing the residue, this too will be measured as lignin.
- =- For these reasons, we prefer the acetyl bromide procedure.tntn"This spectrophotometric method depends on acetylation of
E lignin monomers to give soluble lignin derivatives which are
"̂ "quantitated by A280. These analyses are not affected by thepresence of protein or hydroxycinnamic acids (25, 26) or
Ir n , , fl , dissolved inorganic compounds, since none of those absorb at280 nm. The specific absorption coefficient at 280 nm of theacetyl bromide-treated lignin is due to 0 acetylation of allalcoholic and phenolic hydroxy groups, i.e., terminal phenolicunits (25, 27). This is a fast reaction (25, 27). C acetylation of
________________ aromatic residues at nonterminal positions of lignin may alsooccur, but the reaction is slower and results in strong absorp-tion at around 300 to 305 nm (25, 27), similar to results withoL-carbonyl structures. The C-acetylation reaction competeswith cleavage of 3-aryl ether linkages by the acetyl bromide,which produces new terminal phenolic hydroxyl groups thatare immediately 0 acetylated (27). The extent of C acetylationis dependent on reaction conditions, including the water
0030510 content of the sample, reaction time, and temperature (27). IfOct Nov a shoulder at 300 to 305 nm is observed in the spectrum of the
acetyl bromide solution, the analysis is rejected and moreutral sugar residues and suitable reaction conditions are sought (24, 26). If significantduring composting and amounts of ox-carbonyl structures are introduced into lignins byglucose; A, xylose; *, biological oxidations, e.g., on the propanol side chain of lignin
monomers (34), these ot-carbonyl groups will also contribute to
APPL. ENVIRON. MICROBIOL.
on January 4, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from
COMPOSITIONAL CHANGES IN COMPOST FOR AGARICUS BISPORUS 1543
TABLE 4. Lignin content determined by H2SO4 digestiona
Acid-insoluble Ash in acid- Ash-free insoluble Acid-soluble Total lignin Acid-solubleSample residue (%ODM()insoluble residue residue (% ODM) lignin' (% ODM) (% ODM) lignin )
~ (% DM) ignin)Fresh wheat straw 17.9 1.0 16.9 3.3 20.2 16.5Phase I compost 25.1 2.8 22.3 4.0 26.3 15.3Phase II compost 31.5 11.3 20.2 5.7 25.9 22.0Spent compost 40.3 19.6 20.7 6.3 27.0 23.6
a Digestion with 72% H2SO4 at room temperature for 4 h and then with boiling 3% H2SO4 for 2 h.b % ODM, percent weight of oven-dry matter of unextracted compost.Measured spectrophotometrically at 205 nm.
absorption at 300 to 305 nm. In our experiments, no absorptionin this region was observed and it is assumed that if oa-carbonylgroups had been introduced in lignin through microbial activ-ities in the compost, they would have been further oxidized tocarboxyl groups. Carboxyl groups introduced through biologi-cal activities will not affect A280, and aldehyde groups, ifpresent, would give an absorption around 310 to 315 nm, butthese are generally oxidized further to carboxylic acids (55), asdiscussed below.Chemical changes in lignin during composting and mush-
room growth. Wheat straw is the major lignin-containingingredient of mushroom compost. The lignin is heterogeneous,with about 20% of the original lignin being soluble in boiling3% sulfuric acid (26, 38). The lignin itself is associatedcovalently with other wall polymers, especially polysaccharides.
Significant structural changes in lignin during compostingand mushroom growth were revealed by the results of alkalinenitrobenzene oxidation. In addition to progressive oxidativecondensations (Fig. 7), an increase in the ratio of acids toaldehydes (Fig. 7) was observed, indicating oxidative cleavageat C,-C,3 bonds of lignin side chains. The acids probably ariseby oxidation of aldehyde (53) and ct-carbonyl (34) groupsintroduced by the action of microbial enzymes. This is sup-ported by the observations that wheat straw lignin has nostructural features that would lead to the formation of benzoicacids in alkaline nitrobenzene oxidation and that only smallamounts of these acids are produced from the correspondingaldehydes by the Cannizzaro reaction in alkaline media (22).Similar changes during microbial action were reported byHedges and coworkers (4, 19) and Ziegler and Zech (63). Anincrease in carboxyl groups attached to aromatic moieties inlignin during horticultural composting was also observed by asolid-state ' C NMR procedure (28).These oxidative changes in compost lignins may be catalyzed
by mushroom laccase through a hydroxyl radical mechanism,
TABLE 5. Changes in alkaline nitrobenzene oxidation productsduring composting and mushroom growth
Alkaline nitrobenzene oxidation productSample Total yield sNa Acid/aldehydeb
(% lignin)
Wheat straw 19.5 1.20 0.08
Conventional compostPhase I 16.9 1.17 0.14Phase II 12.8 0.89 0.18Spent compost 12.2 0.97 0.23
a S/V, molar ratio of syringyl to guaiacyl nuclei.bAcid/aldehyde, molar ratio of acids (vanillic and syringic) to aldehydes
(vanillin and syringaldehyde).
probably a one-electron oxidation (11, 53). In vitro experi-ments using lignin model compounds have shown that fungallaccase catalyzes certain degradation reactions (61), particu-larly with syringyl compounds (30, 32, 33, 43), and can alsocause C.-C,3 cleavage in phenolic syringyl units to producebenzoic acid structures (35).The progressive development of lignin in more condensed
form, indicated by the alkaline nitrobenzene oxidation results,could be produced by polymerization of laccase-generatedphenoxy radicals, arising from terminal units of lignin, withother phenoxy radicals (6, 15-18, 21, 31, 33, 41, 42, 44).Polymerization of coniferyl alcohol by mushroom laccase toform artificial lignin is well established (12). Laccase may alsocatalyze coupling between lignins and proteins (40) and be-tween proteins (54). A highly condensed lignin, with a highcontent of carboxyl groups, has been isolated from sugarcanebagasse which had supported the growth of Lentinus edodes(55), a laccase-producing agaric (41).The progressive change in the S/V ratio of the compost
lignins (Table 5) has also been reported for horticulturalcompost by Hedges and coworkers (19) and Ziegler and Zech(63). This may result from the higher sensitivity of syringyl thanguaiacyl nuclei toward aromatic ring-cleaving enzymes (19,20).These alterations in lignin structure have consequences that
are of direct relevance to the quantitation of lignin duringcomposting. The biologically modified lignin is more soluble in
Ecm
mM,cm*cCMF = co) U) C.~U ,.
20 F1 1 1
C
0)
.5.~~~~~~~~~~~~~~~..
02I-.~~~ ~ ~ ~ ~ ~~~~~nI0
Aug Sep Oct1991
Nov
FIG. 7. Changes in total yield (0) and ratio of benzoic acids tobenzaldehydes (0) produced by alkaline nitrobenzene oxidation oflignins in composts from Dandy Mushrooms during composting andmushroom growth.
VOL. 60, 1994
on January 4, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from

1544 IIYAMA ET AL.
TABLE 6. Consumption of xylose and glucose derived fromcompost polysaccharides during composting and mushroom growth
Consumption of neutral sugar (% ofConventional original sugar)
compost sampleXylose Glucose Total
Phase I 34.5 29.0 37.6Phase II 67.9 58.8 65.6Spent compost 88.5 78.4 83.3
acid reagents than native lignin (8, 45) (Table 4) because of theintroduction of carboxyl and hydroxyl groups on lignin mono-mers. This acid-soluble lignin fraction is not measured in lignindeterminations using sulfuric acid digestion. On the otherhand, the modified acetyl bromide procedure measures bothforms of lignin. This point is illustrated by comparing theresults shown in Tables 3 and 4. The proportion of acid-solublelignin to total lignin increases from 15% for phase I to 24% forspent composts, a result similar to that of Effland (8) forbiologically modified wood lignin. The slightly higher totallignin values obtained from a summation of acid-soluble andash-free insoluble (Klason) lignin compared with those fromacetyl bromide lignin (Table 3) are probably due to proteininsolubilization in the Klason procedure. The decrease inrelative amounts of ash-free lignin during mushroom growthhas been interpreted in other studies as indicating utilization oflignin by the mushroom. However, our results show that totallignin (acid-soluble and ash-free insoluble [Klason] lignin)increases slightly during mushroom growth. These consider-ations would explain the decreases in relative amounts ofsulfuric acid digestion residues during mushroom growth ob-served by other workers (7, 13, 60).Our present conclusion is that during processing, lignin
monomers are chemically modified and lignins are to someextent polymerized by condensation reactions. Some cleavageof intermonomer linkages may also occur, but an oxidativedepolymerization such as is achieved by certain lignin-degrad-ing fungi does not take place, as suggested by the increase inrelative content of lignin in samples during composting and
E cm
CI- ~ LL.I.- 'a = XA *- $0D_ c
0.0t
i1
..
LL. - EL
X 'E -* 0l l l
Jv - v * v v t v v v v v
30 = =
*0 X
20 S
Irr . ... . .. . . .16 2u 30 10 20 30
Aug Sep1991
10 20 30 10
Oct Nov
FIG. 8. Consumption of xylose (-) and glucose (0) derived frompolysaccharides in composts from Dandy Mushrooms during compost-ing and mushroom growth (as a percentage of total content of eachmonosaccharide).
mushroom growth. This is in agreement with the conclusions ofInbar and coworkers (28), whose results, based on solid-state13C NMR and FTIR analyses of horticultural compost, sug-gested that lignin is not decomposed by compost microorgan-isms but that oxidative changes do occur.There are, however, other experiments which have a bearing
on the ability of a mushroom to utilize lignin carbons. Inscreening 74 basidiomycetes for their ligninolytic capacitywhen cultured on ['4C]lignin-labelled lignocellulose and "4C-whole-labelled wheat straw, Agosin and coworkers (1) showedthat only 15 of the isolates had relatively high ligninolyticactivity. Two varieties of A. bisporus liberated 8 to 14% of thetotal label as '4Co2, after 10 weeks, compared with 52% by theactive species. Since phenolic acids represented 17.6% of thetotal 14C and 10% of the phenylalanine-14C was in protein, thisis not substantial evidence that A. bisporus can "degrade"lignin under the conditions used. Using the same lignin-'4C-lignocellulose added to a solid-state fermentation, it was found(7) that up to 45% of the 14C label was lost as 14C02 in 80 days.[1 C]lignin (artificially prepared by dehydrogenative polymer-ization of coniferyl alcohol by peroxidase) was also decom-posed to 14Co2 to various extents: 10% (62) and 60% (7). Theapparent quantitative difference between the labelling experi-ments of Durrant et al. (7) and the direct determination oflignin in our experiments remains to be explained.
Sequential changes in carbohydrates during compostingand mushroom growth. A rapid decrease in the relativecontent of neutral sugars from wall polysaccharides was ob-served during composting, with a more gradual decreaseduring mushroom growth. These changes have relevance to theincrease in relative content of lignin (Fig. 6). If we accept thehypothesis that lignin is not decomposed during compostingand mushroom growth as discussed earlier, then we maycalculate the polysaccharide consumption by setting the ligninvalue constant. Calculations from data in Table 3 and Fig. 6show that about 65 to 67% of compost polysaccharides wereconsumed during composting, at both Melbourne Mushrooms(Table 6) and Dandy Mushrooms (Fig. 8), presumably bymicrobial action. During mushroom growth (spawning tofourth flush) on conventional compost, a further 16 to 18% ofthe compost polysaccharides were consumed and 17% of theoriginal polysaccharides remained unutilized. Whether thesepolysaccharides were inaccessible to the mushroom enzymes isunknown.
ACKNOVVLEDGMENTS
This project was supported by the Australian Mushroom GrowersAssociation and the Horticulture Research & Development Corpora-tion.We thank R. Staudte (Department of Statistics, La Trobe Univer-
sity) for suggesting statistical procedures and P. S. Perrin and E. R.Harper (Department of Microbiology, La Trobe University) for col-lecting compost samples. We are also grateful to F. C. Miller of SylvanFoods Inc., Worthington, Pa., for his helpful comments on themanuscript.
REFERENCES1. Agosin, E., J. J. Daudin, and E. Odier. 1985. Screening of white-rot
fungi on ('4C)lignin-labelled and (14C)whole-labelled wheat straw.Appl. Microbiol. Biotechnol. 22:132-138.
2. Australian and New Zealand Pulp and Paper Industry TechnicalAssociation. 1978. Standard Plls-78. Klason lignin in wood andpulp. Australian and New Zealand Pulp and Paper IndustryTechnical Association, Melbourne, Australia.
3. Bacic, A., P. J. Harris, and B. A. Stone. 1988. Structure andfunction of plant cell walls, p. 297-371. In J. Preiss (ed.), Thebiochemistry of plants, vol. 14. Academic Press, New York.
-c1
0-
cn
LD-,
cn 8
a) oC =0 S 4
o .
O: aO .
APPL. ENVIRON. MICROBIOL.
)A-
on January 4, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from

COMPOSITIONAL CHANGES IN COMPOST FOR AGARICUS BISPORUS 1545
4. Benner, R., K. Weliky, and J. I. Hedges. 1990. Early diagenesis ofmangrove leaves in a tropical estuary: molecular-level analyses ofneutral sugars and lignin-derived phenols. Geochim. Cosmochim.Acta 54:1991-2001.
5. Blakeney, A. B., P. J. Harris, R. J. Henry, and B. A. Stone. 1983.A simple and rapid preparation of alditol acetates for monosac-charide analysis. Carbohydr. Res. 113:291-299.
6. Bollag, J. M., R. D. Sjoblad, and S. Y. Liu. 1979. Characterizationof an enzyme from Rhizoctonia praticola which polymerizes phenolcompounds. Can. J. Microbiol. 25:229-233.
7. Durrant, A. J., D. A. Wood, and R. B. Cain. 1991. Lignocellulosebiodegradation byAganicus bisporus during solid substrate fermen-tation. J. Gen. Microbiol. 137:751-755.
8. Eflland, M. J. 1977. Modified procedure to determine acid-solublelignin in wood and pulp. TAPPI 60(10):143-144.
9. Falck, R. 1928. Two natural bacterial processes of cellulose andlignin decomposition in lignified membranes. Cellulosechimie9:1-6.
10. Fermor, T. R., P. Randle, and J. F. Smith. 1985. Compost as asubstrate and its preparation, p. 81-109. In P. B. Flegg, D. M.Spencer, and D. A. Wood (ed.), The biology and technology of thecultivated mushroom. John Wiley & Sons, Chichester, UnitedKingdom.
11. Forney, L. J., C. A. Reddy, M. Tien, and S. D. Aust. 1982. Theinvolvement of hydroxyl radicals derived from hydrogen peroxidein lignin degradation by the white rot fungus Phanerochaetechrysosporium. J. Biol. Chem. 257:11455-11462.
12. Freudenberg, K. 1965. Lignin: its constitution and formation fromp-hydroxycinnamyl alcohols. Science 148:595-600.
13. Gerrits, J. P. G. 1968. Organic compost constituents and waterutilized by the cultivated mushroom during spawn run and crop-ping. Mushroom Sci. 7:111-126.
14. Gerrits, J. P. G., H. C. Bels Koning, and F. D. Muller. 1967.Changes in compost constituents during composting, pasteuriza-tion and cropping. Mushroom Sci. 6:225-243.
15. Gierer, J., and A. E. Opara. 1973. Studies on the enzymaticdegradation of lignin. The action of peroxidase and laccase onmonomeric and dimeric model compounds. Acta Chem. Scand.27:2909-2922.
16. Haars, A., and A. Huttermann. 1980. Function of laccase in thewhite-rot fungus Fomes annosus. Arch. Microbiol. 125:233-237.
17. Haars, A., and A. Huttermann. 1983. Macromolecular mechanismof lignin degradation by Fomes annosus. Naturwissenschaften67:39-40.
18. Haars, A., and A. Huttermann. 1983. Laccase induction in thewhite-rot fungus Heterobasidion annosum (Fr.) Bref. (Fomes an-tIosus Fr. Cooke). Arch. Microbiol. 134:309-313.
19. Hedges, J. I., R. A. Blanchette, K. Weliky, and A. H. Devol. 1988.Effects of fungal degradation on the CuO oxidation products oflignin: a controlled laboratory study. Geochim. Cosmochim. Acta52:2717-2726.
20. Higuchi, T. 1987. Catabolic pathways and role of ligninase for thedegradation of lignin substructure model compounds by white-rotfungi. Wood Res. 73:58-81.
21. Huttermann, A., C. Herche, and A. Haars. 1980. Polymerisation ofwater-insoluble lignins by Fomes annosus. Holzforschung 34:64-66.
22. Iiyama, K., and T. B. T. Lam. 1990. Lignin in wheat internodes. I.The reactivities of lignin units during alkaline nitrobenzene oxi-dation. J. Sci. Food Agric. 51:481-491.
23. Iiyama, K., J. Nakano, and N. Migita. 1967. Studies on colour oflignin. V. Effects of cooking and preparative conditions on thecolour of thiolignin. Kamipa Gikyoshi 21:755-760.
24. Iiyama, K., and A. F. A. Wallis. 1988. An improved acetyl bromideprocedure for determining lignin in woods and wood pulps. WoodSci. Technol. 22:271-280.
25. Iiyama, K., and A. F. A. Wallis. 1989. Effect of acetyl bromidetreatment on the ultraviolet spectra of lignin model compounds.Holzforschung 43:309-316.
26. Iiyama, K., and A. F. A. Wallis. 1990. Determination of lignin inherbaceous plants by an improved acetyl bromide procedure. J.Sci. Food Agric. 51:145-161.
27. Iiyama, K., and A. F. A. Wallis. 1990. Dissolution of wood withacetyl bromide solutions-reactions of lignin model compounds. J.
Wood Chem. Technol. 10:39-58.28. Inbar, Y., Y. Chen, and Y. Hadar. 1989. Solid state carbon-13
nuclear magnetic resonance and infra-red spectroscopy of com-posted organic matter. Soil Sci. Soc. Am. J. 53:1695-1701.
29. Iwade, I. 1958. Cultivation of mushrooms, p. 358. Chikyu-sha,Tokyo.
30. Kamaya, Y., F. Nakatsubo, and T. Higuchi. 1983. Degradation oftrimeric lignin model compounds, arylglycerol-,-syringaresinolethers, by Fusarium solani M-13-1. Agric. Biol. Chem. 47:299-308.
31. Kaplan, D. L. 1979. Reactivity of different oxidases with ligninsand lignin model compounds. Phytochemistry 18:1917-1919.
32. Katayama, T., F. Nakatsubo, and T. Higuchi. 1986. Degradation ofa phenylcoumaran, a lignin substructure model, by Fusarium solaniM-13-1. Mokuzai Gakkaishi. 32:535-544.
33. Kawai, S., H. Ohashi, T. Hirai, H. Okuyama, and T. Higuchi. 1993.Degradation of syringyl lignin model polymer by laccase ofCoriolus versicolor. Mokuzai Gakkaishi 39:98-102.
34. Kirk, T. K., and H. M. Chang. 1975. Decomposition of lignin bywhite-rot fungi. II. Characterization of heavily degraded ligninsfrom decayed spruce. Holzforschung 29:56-64.
35. Kirk, T. K., and R. L. Farrell. 1987. Enzymatic "combustion": themicrobial degradation of lignin. Annu. Rev. Microbiol. 41:465-505.
36. Kofujita, H., K. Nabeta, H. Okuyama, and M. Miyake. 1989.Biodegradation of milled wood lignin on cellulose particle byLentinus edodes. Mokuzai Gakkaishi 35:268-274.
37. Kofujita, H., T. Sato, K. Nabeta, H. Okuyama, and M. Miyake.1989. Components in mizunara (Quercus mongolica var. grosser-rata) bed-logs decayed by shiitake (Lentinus edodes). MokuzaiGakkaishi 35:543-548.
38. Kondo, T., K. Mizuno, and T. Kato. 1987. Variation in solubilitiesof lignin in acid detergent and in alkali. Nippon Sochi Gakkaishi33:296-299.
39. Lam, T. B. T., K. Iiyama, and B. A. Stone. 1990. Primary andsecondary walls of grasses and other forage plants: taxonomic andstructural considerations, p. 43-69. In D. E. Akin, M. G. Ljundahl,R. J. Wilson, and P. J. Harris (ed.), Microbial and plant opportu-nities to improve lignocellulose utilization by ruminants. Elsevier,New York.
40. Leatham, G. F., V. King, and M. A. Stahmann. 1980. In vitroprotein polymerisation by quinones or free radicals generated byplant or fungal oxidative enzymes. Phytopathology 70:1134-1140.
41. Leatham, G. F., and M. A. Stahmann. 1981. Studies on the laccaseof Lentinus edodes: specificity, localization and association with thedevelopment of fruiting bodies. J. Gen. Microbiol. 125:147-157.
42. Leonowicz, A., G. Szklarz, and M. Wojtas-Wasilewska. 1985. Theeffect of fungal laccase on fractionated lignosulphonates (PeritanNa). Phytochemistry 24:393-396.
43. Liu, S. Y., R. D. Minard, and J. M. Bollag. 1981. Oligomerizationof syringic acid, a lignin derivative, by a phenoloxidase. Soil Sci.Soc. Am. J. 45:1100-1105.
44. Lundquist, K., and P. Kristersson. 1985. Exhaustive laccase-catalysed oxidation of a lignin model compound (vanillyl glycol)produces methanol and polymeric quinoid products. Biochem. J.229:277-279.
45. McCarthy, A. J., A. Paterson, and P. Broda. 1986. Lignin solu-bilisation by Themiomonospora mesophila. Appl. Microbiol. Bio-technol. 24:347-352.
46. Miller, F. C., E. R. Harper, and B. J. Macauley. 1989. Fieldexamination of temperature and oxygen relationships in mush-room composting stacks-consideration of stack oxygenationbased on utilisation and supply. Aust. J. Exp. Agric. 29:741-750.
47. Muller, F. M. 1967. Some thoughts about composting. MushroomSci. 6:213-223.
48. Musha, Y., and D. A. I. Goring. 1974. Klason and acid-solublelignin content of hardwoods. Wood Sci. 7:133-134.
49. Pearl, I. A., and L. R. Busche. 1960. Studies on the chemistry ofaspenwood. XI. The Klason lignin determination as applied toaspenwood with special reference to acid-soluble lignin. TAPPI43:961-970.
50. Richtzenhain, H., and E. Dryselius. 1953. Zur Frage der Existanzvon saureloslichem Lignin. Svensk Papperstidn. 56:324-327.
51. Scalbert, A., B. Monties, J. L. Lallemond, E. Guittet, and C.Rolando. 1985. Ether linkage between phenolic acids and lignin
V()L. 60, 1994
on January 4, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from

1546 IIYAMA ET AL. APPL. ENVIRON. MICROBIOL.
fractions from wheat straw. Phytochemistry 24:1359-1362.52. Schoning, A. G., and G. Johansson. 1965. Absorptiometric deter-
mination of acid-soluble lignin in semichemical bisulfite pulp andin some woods and plants. Svensk Papperstidn. 68:607-613.
53. Shimada, M., T. Habe, T. Higuchi, T. Okamoto, and B. Penijpan.1987. Biomimetic approach to lignin degradation. II. The mecha-nism of oxidative C-C bond cleavage reactions of lignin modelcompounds with natural iron(III) porphyrin chloride as a heme-enzyme model system. Holzforschung 41:277-285.
54. Stahmann, M. A., A. K. Spencer, and G. R. Honold. 1977. Crosslinking of proteins in vitro by peroxidase. Biopolymers 16:1307-1318.
55. Suzuki, H., K. Iiyama, 0. Yoshida, S. Yamazaki, N. Yamamoto,and S. Toda. 1990. Structural characterization of the immunoac-tive and antiviral water solubilized lignin in an extract of theculture medium of Lentinus edodes mycelia (LEM). Agric. Biol.Chem. 54:479-487.
56. Theander, O., P. Uden, and P. Aman. 1981. Acetyl and phenolicacid substituents in timothy of different maturity and after diges-tion with rumen microorganisms or a commercial cellulase. Agric.Environ. 6:127-133.
57. Van Soest, P. J. 1963. Use of detergents in analysis of fibrous feeds.II. A rapid method for the determination of fiber and lignin. J.Assoc. Off. Agric. Chem. 46:829-835.
58. Waksman, S. A., and J. M. McGrath. 1931. Preliminary study ofchemical processes involved in the decomposition of manure byAgaricus campestris. Am. J. Bot. 18:573-581.
59. Waksman, S. A., and W. Nissen. 1931. Lignin as a nutrient for thecultivated mushroom, Agaricus campestris. Science 74:271-272.
60. Waksman, S. A., and W. Nissen. 1932. On the nutrition of thecultivated mushroom Agaricus campestris, and the chemicalchanges brought about by this organism in the manure compost.Am. J. Bot. 19:514-537.
61. Wood, D. A., and T. R. Fermor. 1981. Nutrition of Agaricusbisporus in compost. Mushroom Sci. 11:63-71.
62. Wood, D. A., and G. F. Leatham. 1983. Lignocellulose degradationduring the life cycle of Agaricus bisporus. FEMS Microbiol. Lett.20:421-424.
63. Ziegler, F., and W. Zech. 1991. Chemical changes in beech litterand barley straw during decomposition under laboratory condi-tions. Z. Pflanzenemaehr. Bodenkd. 154:377-385.
on January 4, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from





























