Embed Size (px)
Citation preview

J Plant Physiol. Vol. 141. pp. 2-6 {1992}
Introduction
Comparison of Acid Soluble Protein Synthesis in Roots of Endomycorrhizal Wild Type Pisum sativum and Corresponding Isogenic Mutants
LUISA SCHELLENBAUM*, SILVIO GIANINAZZI\ and VIVIENNE GIANINAZZI-PEARSON
Laboratoire de Phytoparasitologie, INRA-CNRS, Station de Genetique et d' Amelioration des Plantes, INRA, BV 1540, 21304 Dijon cedex, France
* Department of Horticulture, The Pennsylvania State University, University Park, PA 16802, USA 1 To whom correspondance should be addressed.
Received April 3, 1992 . Accepted July 20, 1992
Summary
The aim of this study was to compare profiles of acid soluble proteins in roots of Pisum sativum L. cv. Frisson and non-mycorrhizal mutants (myc-) when infected by vesicular arbuscular mycorrhiza funghi. Quantitative and qualitative modifications were detected by native or SDS PAGE in the protein composition of acidic extracts of endomycorrhizal roots of myc+,nod+ wild type P. sativum and of a myc+,nodmutant, as compared to uninoculated control plants or a myc-,nod - mutant, whether this was inoculated or not with Glomus mosseae. In native PAGE two proteins, already present in extracts of non-mycorrhizal pea roots, were enhanced in G. mosseae-infected myc+ plants (nod+ or nod-). One protein which appeared after inoculation with G. mosseae or G. /asciculatum in the roots of all pea plants, including the myc-, nod- mutants, may be associated with appressorium formation. Four new proteins were detected in root extracts of G. mosseae-infected myc+ peas (nod+ or nod-), as compared to the myc-, nod- mutant or uninoculated controls. One of these was probably of fungal origin, since it was also found in G. mosseae-infected tobacco and leek roots. These plants showed protein modifications different to those in endomycorrhizal peas, suggesting host-specific responses. SDS PAGE revealed the presence of two polypeptides of about 25 and 35 kDa in endomycorrhizal roots of myc+ peas. None of the endomycorrhizarelated proteins detected in roots corresponded to those extracted from spores of different fungi. This study provides new evidence that different molecular modifications occur during successful (myc+ pea, leek, tobacco) and unsuccessful (myc- pea) endomycorrhiza infections, some of which are host-determined and others related to the expression of the fungal genome.
Key words: Pisum sativum, Glomus species, Acaulospora laevis, endomycorrhiza, myc-nod- mutants, soluble proteins.
Abbreviations: PAGE = Poly Acrylamide Gel Electrophoresis; PVPP = PolyVinylPolyPyrrolydone; SDS = Sodium Dodecyl Sulphate.
The molecular basis of fungus-plant interactions in mycorrhizal associations with regards to changes in symbiont genome expression during establishment of the symbioses is poorly understood (Gianinazzi, 1991; Bonfante-Fasolo and
Perotto, 1992). Higher transcriptional activity is indicated by an enhanced activity of the host nucleus following mycorrhizal infection (Berta et aI., 1990), and new mycorrhizaspecific polypeptides or proteins have recently been shown to be produced in ectomycorrhiza (Hilbert et aI., 1988, 1991) or in vesicular arbuscular endomycorrhiza (Dumas et al.,
© 1992 by Gustav Fischer Verlag, Stuttgart

1989, 1990; Pacovsky, 1989). Some endomycorrhiza-specific polypeptides have also been reported to immunologically cross-react with antibodies against nodule-specific proteins (nodulins) (Wyss et ai., 1990). However, nothing is known about the function of these mycorrhiza-specific polypeptides and proteins, or whether they playa role in interactions between the plant and fungal symbionts.
Recently, chemically induced isogenic mutants of Pisum sa· tivum have been discovered (Duc et ai., 1989, GianinazziPearson et ai., 1991) that are either resistant to both endomycorrhizal fungi and Rhizobium (myc- ,nod-), the fungal symbiont only reaching the morphological stage of appressorium development, or establish an endomycorrhizal association but do not nodulate (myc+,nod-).
The aim of the present study was to compare the soluble protein profiles of roots of endomycorrhizal and non-mycorrhizal wild type (myc+ ,nod +) P. sativum with those of the different mutants (mye,nod- and myc+,nod-), in order to
detect possible molecular changes specific to the symbiotic state. Root soluble proteins have also been analyzed in P. sa· tivum infected with different endomycorrhizal fungi, and in different plant genera infected with the same fungus as P. sa· tivum. Since vesicular arbuscular fungi cannot be grown in pure culture, spore soluble proteins were extracted to see whether any new endomycorrhiza-related proteins were already present in spores and therefore likely to be of fungal OrIgm.
Materials and Methods
Plant material and growth conditions
Seeds of Pisum sativum L. cv. Frisson (myc+ ,nod+) and of the corresponding P56 (myc+,nod-) and P2 (myc- ,nod-) mutants were surface sterilized in 7 % calcium hypochlorite for 10 min, pregerminated in vermiculite in the dark at 20 0 e and planted into pots containing 400g of a disinfected clayloam soil (pH 7.6): perlite (3: 1) mix. Plants were either inoculated with Glomus mosseae (Nicol & Gerd) Gerd and Trappe (isolate LPA 5) using a root inoculum from a stock culture of Allium porrum L. cv. Verina, or grown in disin· fected soil as non-mycorrhizal control plants. All plants were inocu· lated with Rhizobium leguminosarum bv. viciae, strain 1007 (from N. Amarger, INRA, Dijon). Additional plants of wild type P. sati· vum (myc+,nod+) were inoculated with G.fasciculatum (LPA 7) and Acaulospora laevis Gerd and Trappe (LPA 1), using a silty acid soil pH 5.0) for the latter. Nicotiana tabacum L. cv. Xanthi and A. por· rum cv. Verina seeds were sterilized and inoculated with G. mosseae as described above. Plants were maintained in a growth chamber (20 0 e, 80% rh, 16h day, 300llmols- 1 m- 2
) and watered daily with deionized water.
Extraction of soluble proteins from roots and spores
Roots from control and inoculated plants were harvested three weeks after transplanting (except for tobacco and leek which were harvested at 6 weeks), carefully washed in cold water and root pieces with visible nodules were removed. Samples were ground at 4 °e in 0.06 M phosphate-citrate buffer (1 g mL -1) at pH 2.8, containing 5 mM ascorbic acid, 1 % PVPP and 15 mM mercaptoethanol. The samples were centrifuged (60 min 10000 x g) and the supernatant collected. Spores of G. mosseae (400), G. fasciculatum (400) and A. laevis (200) were isolated from soil by wet sieving. Soluble spore
Protein in endomycorrhizal myc+ and myc- plants 3
proteins were extracted in 100 ilL of buffer at pH 7.6 according to Sanders et al. (1992), and at pH 2.8 using the same buffer as that used for roots. The protein contents of the extracts were estimated by the method of Bradford (1976), using bovine serum albumin as a standard.
Electrophoretic separation of proteins
Soluble proteins were separated at 4 °e using discontinuous native (Davis, 1964) and SDS (Laemmli, 1970) PAGE on mini vertical uniform polyacrylamide slab gels (Imm thick, 8x7cm) at 10% (25 rnA h -1) and 15 % (30 rnA h -1), respectively. Polypeptide mobilities were compared with those of a range of molecular weight markers (Sigma kit). The same volume of extract was used for all samples (10 ilL for roots, 15 ilL for spores). Proteins were revealed by silver nitrate staining (Blum et aI., 1987).
Determination of the endomycorrhizal infection
The amount of infection developing in roots was estimated according to Trouvelot et al. (1986), after digestion of roots in 10% KOH and staining with cotton blue (0.1 %) in lactic acid. Parameters are expressed as frequency of infected root samples (F%) and percent colonised root cortical length (M%) within the endomycorrhizal root system.
Results and Discussion
Endomycorrhizal infection and arbuscule formation was already well-established in the three week-old wild type (myc+,nod+) and PS6 mutant (myc+,nod-) pea plants, and values for F% (77 -100) and M% (29 - 55) did not differ significantly for G. mosseae or G. /asciculatum infections in repeated experiments. Significantly lower values (P = 0.05) were, however, obtained for infection of P. sativum (myc+,nod+) inoculated with A. laevis (F% = 42, M% = 4). In the case of G. mosseae-inoculated mye ,nod- mutant, the frequency of root fragments with appressoria was consistently high (average of 78 %).
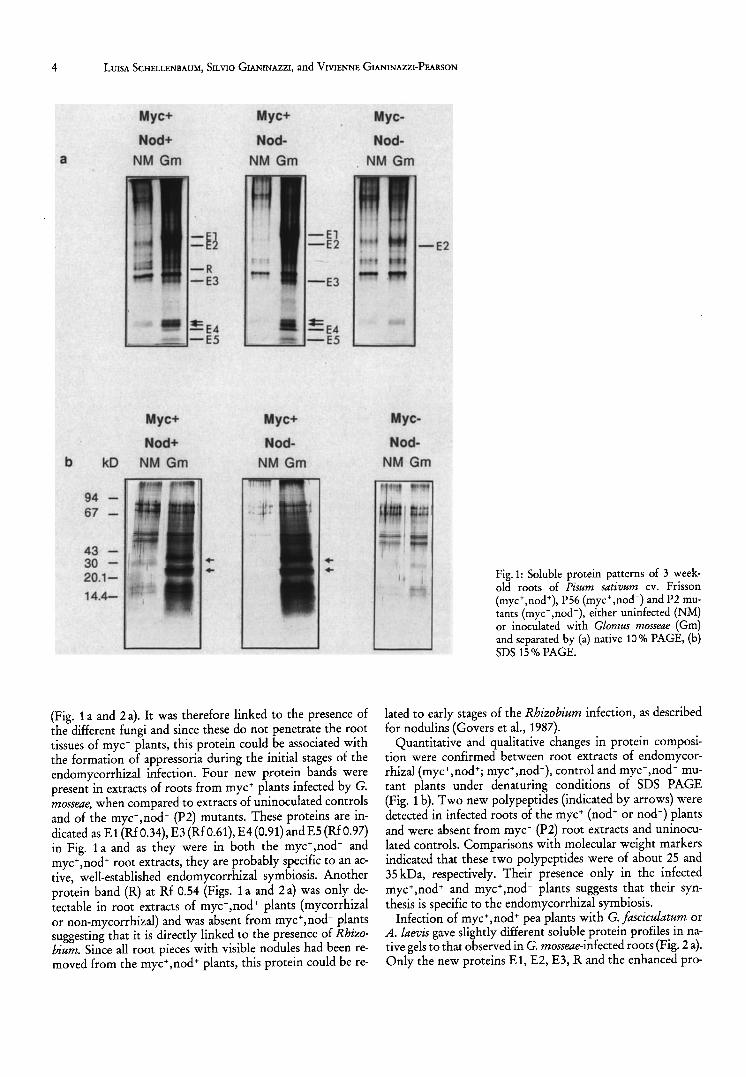
Endomycorrhizal root extracts of P. sativum cv. Frisson contained more soluble proteins than non-mycorrhizal ones, as reported in preliminary analyses (Schellenbaum, 1991) and by Dumas et al. (1989) for endomycorrhizal tobacco roots. In fact, in the present study more protein was detected following PAGE of pH 2.8 extracts of endomycorrhizal roots of wild type P. sativum (myc+, nod+) and of the PS6 mutant (myc+,nod-), as compared to uninoculated controls and to
the P2 mutant (myc-,nod-), whether this was inoculated or not with G. mosseae (Fig. 1 a and b). Protein bands were particularly intense in the upper part of gels of en do mycorrhizal root extracts of myc+ pea plants (wild type and PS6). Both quantitative and qualitative modifications were observed in protein bands for the different extracts and a certain number of these could be clearly distinguished (Figs. 1 a and 2 a).
Two proteins already present in extracts of non-mycorrhizal pea roots, and indicated by arrows at Rfs 0.86 and 0.88 in the lower part of gels, were enhanced in G. mosseae-infected myc+ plants, whether these were nod + or nod - (Fig. 1 a). One band at Rf 0.40 (E2) was present in the roots of all endomycorrhiza-inoculated pea plants, including the mye,nodmutant, but it was absent from the uninoculated controls
4 LUISA SCHELLENBAUM, SILVIO GIANINAZZI, and VIVIENNE GIANINAZZI-PEARSON
a
b
94 67
kD
43 -30 -20.1-
14.4-
Myc+
Nod+
NM Gm
Myc+
Nod+
NM Gm
Myc+
Nod
NM Gm
Myc+
Nod
NM Gm
(Fig. 1 a and 2 a). It was therefore linked to the presence of the different fungi and since these do not penetrate the root tissues of myc- plants, this protein could be associated with the formation of appressoria during the initial stages of the endomycorrhizal infection. Four new protein bands were present in extracts of roots from myc+ plants infected by G. mosseae, when compared to extracts of uninoculated controls and of the myc-,nod- (P2) mutants. These proteins are indicated as E1 (RfO.34), E3 (RfO.61), E4(0.91) andES (RfO.97) in Fig. 1 a and as they were in both the myc+ ,nod- and myc+,nod+ root extracts, they are probably specific to an active, well-established endomycorrhizal symbiosis. Another protein band (R) at Rf 0.54 (Figs. 1 a and 2 a) was only detectable in root extracts of myc+,nod+ plants (mycorrhizal or non-mycorrhizal) and was absent from myc+,nod- plants suggesting that it is directly linked to the presence of Rhizobium. Since all root pieces with visible nodules had been removed from the myc+,nod+ plants, this protein could be re-
Myc
NOd
NM Gm
Myc
Nod
NM Gm
Fig. 1: Soluble protein patterns of 3 weekold roots of Pisum sativum cv. Frisson (myc+,nod+), P56 (myc+,nod-) and P2 mutants (myc- ,nod -), either uninfected (NM) or inoculated with Glomus mosseae (Gm) and separated by (a) native 10% PAGE, (b) SDS 15 % PAGE.
lated to early stages of the Rhizobium infection, as described for nodulins (Govers et al., 1987).
Quantitative and qualitative changes in protein composition were confirmed between root extracts of endomycorrhizal (myc+,nod+; myc+,nod-), control and myc,nod- mutant plants under denaturing conditions of SDS PAGE (Fig. 1 b). Two new polypeptides (indicated by arrows) were detected in infected roots of the myc+ (nod+ or nod-) plants and were absent from myc- (P2) root extracts and uninoculated controls. Comparisons with molecular weight markers indicated that these two polypeptides were of about 25 and 35 kDa, respectively. Their presence only in the infected myc+,nod+ and myC+,nod- plants suggests that their synthesis is specific to the endomycorrhizal symbiosis.
Infection of myc+ ,nod+ pea plants with G. /asciculatum or A. laevis gave slightly different soluble protein profiles in native gels to that observed in G. mosseae-infected roots (Fig. 2 a). Only the new proteins E1, E2, E3, R and the enhanced pro-

a
b
Gft
P. sativum
Gm AI
A. porrum
NMGm
_ El
- E2
- R
Rf
0.48 0.52 0.56
E4
E3
N. tabacum
NMGm
Fig. 2: Native soluble proteins from (a) 3 week-old Pisum sativum cv. Frisson roots inoculated with Glomus Jasciculatum (Gft), G. mosseae (Gm) or Acaulospora laevis (AI) and from (b) 6 week-old Nicotiana tabacum and Allium pm'rum roots, either uninfected or inoculated with G. mosseae.
tein at Rf 0.88 were detectable in the presence of G. Jasciculatum, whilst protein modifications were even less striking in the A. laevis infections as compared to the other host-fungus combinations, possibly due to the low level of infection developed by A. laevis in P. sativum cv. Frisson. A new band at the same Rf as E4 was found in root extracts of six week-old G. mosseae-infected tobacco (F% = 95, M% = 20) and leek (F% = 79, M% = 49). Since this protein band was also present in myc+ pea roots infected by the same fungus, it seems highly probable that this protein is of fungal origin.
As in endomycorrhizal pea roots, soluble protein synthesis was enhanced both quantitatively and qualitatively in endomycorrhizal leek and tobacco roots (Fig. 2 b). At least three new protein bands appeared in leek and two in tobacco roots infected by G. mosseae, in addition to the band correspond-
Protein in endomycorrhizal myc+ and myc" plants 5
a
b
pH 2.8
AI Gft Gm
pH 7.6
AI Gft Gm
Fig. 3: Native soluble proteins extracted from Acaulospora laevis (AI), Glomus Jasciculatum (Gft) or G. mosseae (Gm) spores at pH 2.8 (a) and 7.6 (b).
ing to E4. Two bands showing the same mobilities in extracts of both plants (Rfs 0.48 and 0.56) were not present in mycorrhizal pea roots, whilst one additional band (Rf 0.52) was specific to infected leek roots and it is therefore probable that some of these modifications represent host-specific responses.
Only a few proteins were detected after electrophoretic separation of A. laevis spore proteins soluble at pH 2.8 and none could be shown for C. Jasciculatum or G. mosseae (Fig. 3 a), even when a large number of spores were extracted (900 spores 100 IlL"1, results not shown). Although more proteins, including some of C. mosseae, could be detected by extracting at pH 7.6 (Fig. 3 b), none corresponded to those appearing in root extracts after endomycorrhizal infection. These observations suggest that the newly detected proteins reported in the present work in endomycorrhizal myc+ pea, leek or tobacco roots are not present in spores. Proteins which are thought to be of fungal origin may be specifically

6 LUISA SCHELLENBAUM, SILVIO GIANINAZZI, and VIVIENNE GIANINAZZI-PEARSON
induced, or enhanced, during the symbiotic phase of fungal development within the host tissues.
The results presented here show that a vesicular arbuscular endomycorrhizal infection results in the synthesis of new soluble proteins in different host-fungus combinations and enhances production of some already present in uninfected roots. This is in agreement with previous reports of modifications in protein metabolism in tobacco and soybean (Dumas et aI., 1989, 1990; Pacovsky, 1989; Wyss et aI., 1991). Moreover, this study comparing molecular modifications occurring in pea mutants that are resistant to endomycorrhizal fungi and! or Rhizobium provides further evidence that specific proteins are produced only when roots successfully interact with vesicular arbuscular fungi, some of which are probably due to modifications in host protein synthesis and others to changes in expression of the fungal genome.
Acknowledgement
This work was partly supported by a BRIDGE (EEe) program grant (L.S.).
References
BERTA, G., S. SGORBATI, V. SOLER, A. FUSCONI, A. TROTTA, A. CITTERIO, M. G. BOTTONE, E. SPARVOLI, and S. SCANNERINI: Variations in chromatin structure in host nuclei of a vesicular-arbuscular mycorrhiza. New Phytol. 114, 199-205 (1990).
BLUM, H., H. BEIER, and H. J. GROSS: Improved silver staining of plant proteins, RNA and DNA in polyacrylamide gels. Electrophoresis 8, 93 - 99 (1987).
BONFANTE-FASOLO, P. and S. PEROTTO: Plants and endomycorrhizal fungi: the cellular and molecular basis of their interaction. In: VERMA, D. P. S. (ed.): Molecular Signals in Plant Microbe Communications, pp. 445-470. CRC Press Inc. (1992).
BRADFORD, M. M.: A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Ana!. Biochem. 72,248-254 (1976).
DAVIS, B. J.: Disc electrophoresis II. Method and application to human serum proteins. Ann. N.Y. Acad. Sci. 121, 404-427 (1964).
Duc, G., A. TROUVELOT, V. GIANINAZZI-PEARSON, and S. GIANINAZZI: First report of non-mycorrhizal plant mutants (Myc-) obtained in pea (Pisum sativum L.) and fababean (Vicia faba L.). Plant Science 60, 215-222 (1989).
DUMAS, E., V. GIANINAZZI-PEARSON, and S. GIANINAZZI: Production of new soluble proteins during VA endomycorrhiza formation. Agric., Ecosyst. and Envir. 29, 111-114 (1989).
DUMAS, E., A. TAHIRI-ALAOUl, S. GIANINAZZI, and V. GIANINAZZIPEARSON: Observations on modifications in gene expression with
VA endomycorrhiza development in tobacco: qualitative and quantitative changes in protein profiles. In: NARDON, P., V. GIANINAZZI-PEARSON, A. M. GRENIER, L. MARGULIS, and D. C. SMITH (eds.): Endocytobiology IV, pp. 153 -157. INRA Editions (1990).
GIANINAZZI, S.: Vesicular-arbuscular (endo-) mycorrhizas: cellular, biochemical and genetic aspects. Agric. Ecosyst. and Envir. 35, 105 -119 (1991).
GIANINAZZI-PEARSON, V., S. GIANINAZZI, J. P. GUILLEMIN, A. TROUVELOT, and G. Duc: Genetic and cellular analysis of resistance to vesicular arbuscular (VA) mycorrhizal fungi in pea plants. In: HENNECKE, H. and D. P. S. VERMA (eds.): Advances in Molecular Genetics of Plant-Microbe Interactions vol. I, pp. 336-342. Kluwer Academic Publishers (1991).
GOVERS, F., J. P. NAP, A. VAN KAMMEN, and T. BISSELING: Nodulins in the developing root nodule. Plant Physiol. Biochem. 25, 309-322 (1987).
HILBERT, J.-L. and F. MARTIN: Regulation of gene expression in ectomycorrhizas. 1. Protein changes and the presences of ectomycorrhiza-related proteins in the Pisolithus-Eucalyptus symbiosis. New Phytol. 110,339-346 (1988).
HILBERT, J.-L., G. COSTA, and F. MARTIN: Ectomycorrhizin synthesis and polypeptide changes during the early stage of eucalypt mycorrhiza development. Plant Physiol. 97, 977 -984 (1991).
LAEMMLI, U. K.: Cleavage of structural proteins during the assembly of the head of the bacteriophage T4. Nature 227, 680-689 (1970).
PACKOVSKY, R. S.: Carbohydrate, protein and amino acid status of Glycine-Glomus-Bradyrhizobium symbioses. Physio!. Plant. 75, 346-354 (1989).
SANDERS, 1. R., F. RAVOLANIRINA, V. GIANINAZZI-PEARSON, and S. GIANINAZZI: Identification and detection of vesicular-arbuscular mycorrhizal fungi using polyclonal antibodies (IgG) to soluble spore fractions of Gigaspora margarita (Becker & Hall) and Acau· lospora laevis (Gerdemann & Trappe). Mycological Research 96, 477 -490 (1992).
SCHELLENBAUM, L.: Plant mutants resistant to VAM infection: an experimental model for studying gene expression during symbiosis. Mini-Symposium on Symbiosis and Crop Growth: Molecular and Organismal Aspects, Grasmere, G.B., 30 April-3 May (1991).
TROUVELOT, A., J. L. KOUGH, and V. GIANINAZZI-PEARSON: Mesure du taux de mycorhization d'un systeme radiculaire. Recherche de methodes d'estimation ayant une signification fonctionnelle. In: GIANINAZZI-PEARSON, V. and S. GIANINAZZI (eds.): Physiological and Genetical Aspects of Mycorrhizae, pp. 217 - 221. INRA (1986).
WySS, P., R. B. MELLOR, and A. WIEMKEN: Vesicular-arbuscular mycorrhizas of wild-type soybean and non-nodulating mutants with Glomus mosseae contain symbiosis-specific polypeptides (mycorrhizins), immunologically cross-reactive with nodulins. Planta 182,22-26 (1990).





























