Embed Size (px)
Citation preview

ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS 167, 165-175 (1975)
Comparative Studies on P-Glucan Hydrolases
Isolation and Characterization of an Exo(l-+3)-/3-Glucanase from the Snail,
Helix pomatia ‘* ’
J. J. MARSHALL3 AND R. J. A. GRAND’
Department of Chemistry, Royal Holloway College (University of London), Englefield Green, Surrey TW20 OEX, England., Laboratories for Biochemical Research, Howard Hughes Medical Institute, and Department of
Biochemistry, University of Miami School of Medicine, Miami, Florida 33152
Received September 13, 1974
An exo-fi-glucan hydrolase, present in the digestive juice of the snail, Helix pomatia, has been purified to homogeneity by chromatography on Bio-Gel P-60, Sephadex G-ZOO, DEAE-cellulose, and DEAE-Sephadex. The enzyme degrades p-(1 -3)-linked oligosaccha- rides and polysaccharides, rapidly and to completion, or near completion, yielding glucose as the major product of enzyme action. Mixed linkage (1+3; 1+4)-j3-glucans are also ex- tensively degraded and j3-(l-6)- and &(l-4)-linked glucose polymers are slowly degraded by the enzyme. This enzyme differs from other exo-@-glucanases, reported previously, in the broadness of its substrate specificity. The K, values for action on laminarin and li- chenin are respectively 1.22 and 2.22 mg/ml; the maximum velocity of action on laminarin is approximately twice that on lichenin. The enzyme has a molecular weight of 82,000 as determined by polyacrylamide gel electrophoresis. Maximum activity is exhibited at pH 4.3 and at temperatures of 50-55°C.
The highly complex nature of the /3- glucanase mixture present in snail diges- tive juice has long been recognized (2, 3). Use has been made of the unpurified mix- ture of glucanases for the formation of yeast protoplasts and related purposes (4), but the nature and role of the enzymes functional in the degradation of microbial cell walls has not been established. In view of the importance of purified &glucanases,
with rigidly established specificities, for the examination of the molecular architec- ture of microbial cell walls, and the mecha- nism of their enzymolysis, as well as for the examination of the molecular struc- tures of isolated polysaccharides (5, 6), we have undertaken the separation and purifi- cation of the constituent ,f3-glucanases of snail juice. We now report on the purifica- tion and properties of a new type of exo-
r Supported m part by a grant from the Science (l&3)-P-giucanase [( l-3)+glucan gluco-
Research Council. hydrolase, EC 3.2.1.581 from this source.
’ This paper is the third in the series “Comparative Studies on B-Glucan Hydrolases.” Paper II appeared MATERIALS AND METHODS
in Comp. Biochem. Ph&iol. B., in press (1). - - Glusulase, a crude mixture of snail digestive en- 3 Investigator of the Howard Hughes Medical Insti- zymes, was purchased from Endo Laboratories, Gar-
tute. Address enquiries and reprint requests to J. J. den City, NY. Marshall, Department of Biochemistry, University of Substrates were prepared or obtained as reported Miami School of Medicine, P.O. Box 520875, Bis- in earlier publications (1, 7-10). Laminarin polyal- cayne Annex, Miami, FL 33152. cohol was prepared as by Nelson et al. (11); nonreduc-
’ Present address: Department of Biochemistry, ing chain ends were regenerated by treatment of this University of Birmingham, Birmingham, Warwicks, modified polysaccharide with 0.01 N sulfuric acid for England. 15 h at room temperature.
165
Copyright 0 1975 by Academic Press, Inc. All rights of reproduction in any form reserved

166 MARSHALL AND GRAND
Glucose oxidase (type II), peroxidase (type l), o-dianisidine dihydrochloride, human serum albumin (Cohn fraction V), cysteine hydrochloride, oxidized glutathione, N-bromosuccinimide, gluconic acid b- lactone, p-chloromercuribenzoic acid, and 2-hydroxy 5nitrobenzylbromide were purchased from the Sigma Chemical Company.
Microgranular DEAE-cellulose (DE-X?) was a product of Whatman Biochemicals; DEAE-Sephadex A-50 and Sephadex G-200 were from Pharmacia Fine Chemicals; Bio-Gel P-60 was from Bio-Rad Laborato- ries.
Analytical methods. Protein concentrations of en- zyme solutions were determined by the method of Lowry et al. (12). using bovine serum albumin as standard. In column fractions, protein was detected by measurement of absorbance at 280 nm. Reducing sugars released by enzyme action were measured by reduction of an alkaline copper reagent (13). Glucose was determined specifically by using glucose oxidase (14). Total carbohydrate was determined by the phenol-sulfuric acid method (15), calibrated against glucose.
Enzyme assays. During purification of the enzyme, (1 + 3)-@-glucanase activity was determined by mea- surement of the reducing sugars produced in 1.0 ml digests containing substrate (laminarin, 2.5 mg), acetate buffer (100 mM, pH 5.3, 0.25 ml), calcium chloride (1.25 mg), and a suitable amount of enzyme, incubated at 37°C. One unit of enzyme activity is the amount which releases 1 pmole of glucose equivalents/ min under these conditions. Specific activities are the number of units of activity/mg of protein.
Since the purified enzyme was found to exhibit maximum activity at about pH 4.5, studies on the properties of the enzyme were carried out at this lower pH value, unless otherwise indicated.
Gel filtration. Gel filtration was performed at 2°C on a column (100 x 2.5 cm) of Bio-Gel P-60 (100-200 mesh), eluted with 25 mM citrate-phosphate buffer pH 8.0, containing 1% sodium chloride, 5.0-ml frac- tions being collected automatically. Chromatography on Sephadex G-200 was performed using a column of dimensions 90 x 1.6 cm eluted with the same buffer, 2.6-ml fractions being collected. Both types of gel filtration media were prepared as recommended by their respective manufacturers (16-17).
Ion-exchange chromatography. Chromatography on microgranular DEAE-cellulose, pretreated as rec- ommended (la), and equilibrated with 25 rnru ci- trate-phosphate buffer pH 8.0, was performed at 2°C in a column of dimensions 10 x 3.0 cm. Protein was eluted with a pH gradient formed by running 250 ml 25 mM citric acid into 250 ml 25 mM citrate-phosphate buffer pH 8.0, 10 ml fractions being collected.
DEAE-Sephadex was prepared for use by the recommended method (19), and equilibrated with 25 mM citrate-phosphate buffer pH 8.0. Chromatogra-
phy on this medium was carried out at 2°C in a 5 x 1.0 cm column, the protein being eluted with a linear gradient (0 - 1.0 M) of sodium chloride in the same buffer. Fractions of volume 10 ml were collected.
Polyacrylamide gel electrophoresis. Electrophore- sis in polyacrylamide gels was performed according to the method of Ornstein and Davis (20, 21). Molecular weights were determined by gel electrophoresis in the presence of sodium dodecyl sulfate and 2-mercapto- ethanol (22).
Ultrafiltration. Protein solutions were concen- trated by ultrafiltration at 2°C in a Diaflo cell (Amicon Corp., Lexington, MA) fitted with a UM 10 membrane.
Polarimetry. Optical rotations were measured using a 1-dm quartz cell, thermostatted at 43”C, in a Perkin-Elmer 141 polarimeter.
Paper chromatography. Decending paper chro- matograms were developed in the solvent system ethyl acetate:pyridine:water (10:4:3, by volume). Sugar spots were vizualized using an alkaline silver nitrate reagent (23).
RESULTS
Enzyme Purification
Step 1: Chromatography on Bio-Gel P-60. Snail digestive juice (2.5 ml) was chromatographed on Bio-Gel P-60 and the column fractions were assayed for activity towards laminarin. Two peaks of (l--+3)- @-glucanase activity were found, the first eluted at the void volume of the column, and a second at a much higher elution volume (Fig. 1). Chromatographic examination of the products of action of fractions in the first peak on laminarin showed glucose to be the main product. The fractions in the second peak produced, during action on the same substrate, glu- cose and a series of oligosaccharides with the chromatographic mobility of fi-( l&3)- linked oligosaccharides. These findings indicated that the first peak contained a p-glucosidase or exo-fi-glucanase, while the second was endo-( 1+3)-P-glucanase. Frac- tions of the first peak emerging from the column (loo-160 ml) were combined.
Step 2: Chromatography on Sephadex G-200. The enzyme preparation from Bio- Gel P-60 was concentrated, dialyzed and chromatographed on Sephadex G-200. Assay of the fractions for (1 + 3)+gluca- nase activity showed that this procedure separated the activity into two fractions

SNAIL EXO-B-GLUCANASE 167
(Fig. 2). The fractions comprizing the first peak of activity (material eluting at 44-75 ml) were combined.
Step 3: Chromatography on DEAE- Cellulose. The combined, concentrated and dialyzed enzyme preparation from the Sephadex G-200 column was chromato- graphed on DEAE-cellulose, using a pH gradient for elution. The majority of the (l-3)-@-glucanase activity was eluted at a late stage in the gradient, between pH
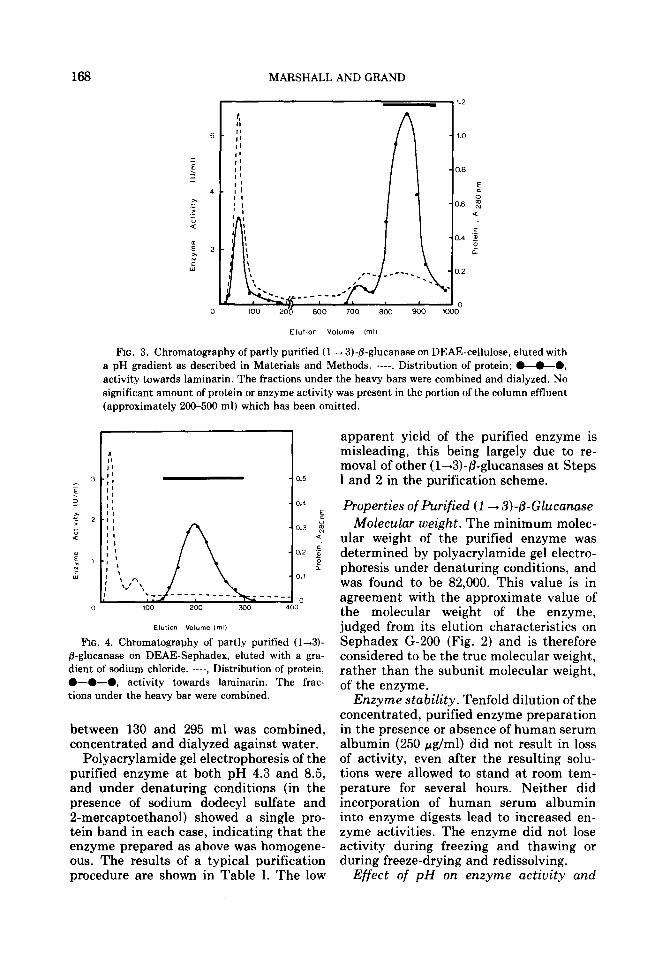
4.5-2.8 (Fig. 3). The material eluted be- tween 790 and 950 ml was combined.
Step 4: Chromatography on DEAE- Sephadex. The combined, concentrated and dialyzed (1 --+ 3)-/3-glucanase prepara- tion from DEAE-cellulose was applied to DEAE-Sephadex and eluted with a gradi- ent of sodium chloride. A single peak of activity towards laminarin was found, eluted at 0.1-0.35 M sodium chloride con- centration (Fig. 4). The material eluted
ElUtlOn Volume lmll
FIG. 1. Chromatography of crude snail juice on Bio-Gel P-60. --- -, Distribution of protein; C-0, activity towards laminarin. The fractions under the heavy bar were combined and concentrated.
FIG. 2. Chromatography of partly purified (1-3)~@-glucanase on Sephadex G-200. ----, Distribution of protein; CO-O, activity towards laminarin. The fractions under the heavy bar were combined and dialyzed.
168 MARSHALL AND GRAND
ElUtlOn Volume ImU
FIG. 3. Chromatography of partly purified (1 -+ 3)-B-glucanase on DEAE-cellulose, eked with a pH gradient as described in Materials and Methods. ----, Distribution of protein; @-O--O, activity towards laminarin. The fractions under the heavy bars were combined and dialyzed. No significant amount of protein or enzyme activity was present in the portion of the column effluent (approximately 200-500 ml) which has been omitted.
FIG. 4. Chromatography of partly purified (l-3)- P-glucanase on DEAE-Sephadex, eluted with a gra- dient of sodium chloride. ----, Distribution of protein, O-0-0, activity towards laminarin. The frac- tions under the heavy bar were combined.
between 130 and 295 ml was combined, concentrated and dialyzed against water.
Polyacrylamide gel electrophoresis of the purified enzyme at both pH 4.3 and 8.5, and under denaturing conditions (in the presence of sodium dodecyl sulfate and 2-mercaptoethanol) showed a single pro- tein band in each case, indicating that the enzyme prepared as above was homogene- ous. The results of a typical purification procedure are shown in Table I. The low
apparent yield of the purified enzyme is misleading, this being largely due to re- moval of other (l-+3)-@-glucanases at Steps 1 and 2 in the purification scheme.
Properties of Purified (1 -+ 3)-p-Glucanase Molecular weight. The minimum molec-
ular weight of the purified enzyme was determined by polyacrylamide gel electro- phoresis under denaturing conditions, and was found to be 82,000. This value is in agreement with the approximate value of the molecular weight of the enzyme, judged from its elution characteristics on Sephadex G-200 (Fig. 2) and is therefore considered to be the true molecular weight, rather than the subunit molecular weight, of the enzyme.
Enzyme stability. Tenfold dilution of the concentrated, purified enzyme preparation in the presence or absence of human serum albumin (250 pg/ml) did not result in loss of activity, even after the resulting solu- tions were allowed to stand at room tem- perature for several hours. Neither did incorporation of human serum albumin into enzyme digests lead to increased en- zyme activities. The enzyme did not lose activity during freezing and thawing or during freeze-drying and redissolving.
Effect of pH on enzyme activity and

SNAIL EXO-B-GLUCANASE 169
stability. Enzyme activity was determined at different pH values using digests con- taining laminarin (2.5 mg), citrate buffer (25 mM, pH 3.0-6.6) and enzyme solution (10 ~1) in a total volume of 1.0 ml. The pH-activity curve (Fig. 5) which showed maximum activity at pH 4.4-4.5, was the same in the presence and absence of human serum albumin (250 pg/ml) (cf. 10). When the activity was determined after preincubation of the enzyme in the buffers of different pH for 2 h at 37”C, followed by initiation of enzyme action by addition of substrate, a pH-activity curve was ob- tained which was closely similar on the neutral side of the optimum pH, but show- ing decreases in activity at pH 4.0 and 3.5 of 10% and 35%, respectively, suggesting that the purified enzyme is labile under acid conditions.
Optimum temperature and heat stability. Enzyme activity was determined at different temperatures in digests of com- position: laminarin (2.5 mg), acetate buffer (pH 4.5, final concentration 25 mM) and enzyme solution (10 ~1) in a total volume of 1.0 ml, incubated at the chosen tempera- ture for 30 min. Under these conditions, the activity of the enzyme was maximal at 53°C (Fig. 6).
The thermal stability of the enzyme was determined by preincubation of all constit- uents of the above digests, except sub- strate, at the chosen temperature for 30 min, followed by addition of substrate and measurement of the activity remaining by incubation at 37°C. The enzyme was found to be stable up to 50°C and rapidly inacti- vated at higher temperatures (Fig. 7).
Substrate specificity of purified (l-3)-
TABLE I
PURIFICATION SCHEME FOR Helix pomatia Exo-(1 4 3b@-GLUCANASE
Step Stage Protein (14 3)-P- Specific Yield” (mg) Glucanase activity (o/o)
activity (U) Wmd
Crude snail juice 370 436 1.2 100 1 Bio-Gel P-60 chromatography 261 108 0.4 24.6 2 Sephadex G-100 chromatography 78 44 0.6 10.1 3 DEAE-cellulose chromatography 7.2 33.3 4.6 7.6 4 DEAE-Sephadex chromatography 1.0 24.2 24.2 5.4
’ The low overall yield is misleading, being partly due to the removal of other (1 + 3)-B-glucanases at Steps 1 and 2. The presence of a multiplicity of (1 + 3)-@-glucanases in the starting material makes it difficult to calculate accurately the overall extent of purification of the exo-(I + 3)-B-glucanase.
01
3.0 3.5 4.0 4.5 5.0 5.5 6.0 6.5
PH
FIG. 5. Dependence of activity of Helix pomatia (1 + 3)-P-glucanase on pH. For conditions see the text.

170 MARSHALL AND GRAND
Temperature (“Cl
FIG. 6. Dependence of Helix pomatia (l-3)-,%glu- canase activity on temperature.
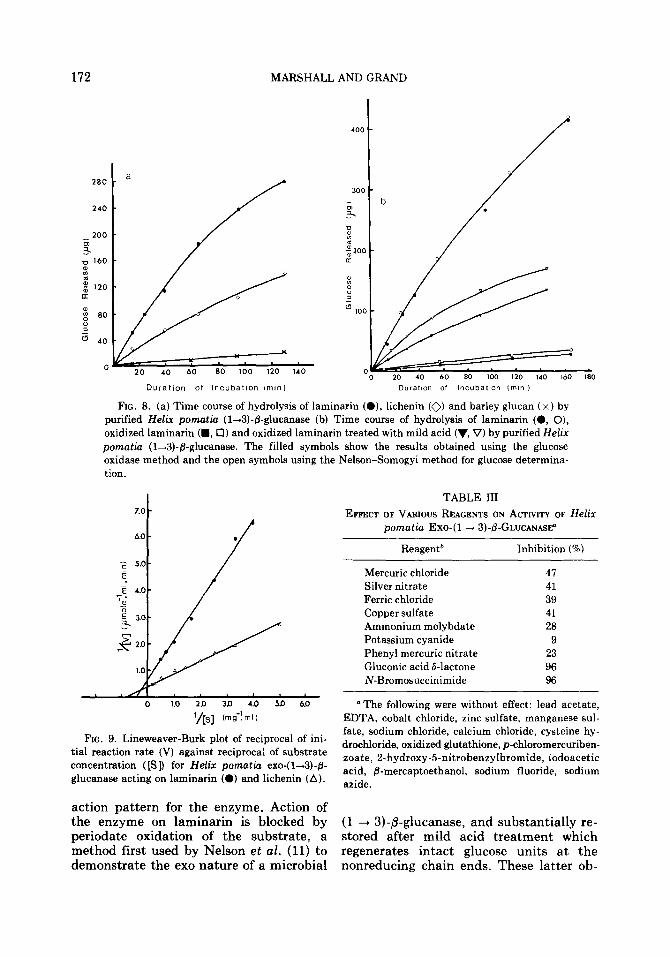
fi-glucanase. To determine the relative rates of action of the purified enzyme on oligo- and polysaccharides, digests of vol- ume 7.0 ml containing substrate (approxi- mate concentration 0.5 mM), acetate buffer (pH 4.5, final concentration 25 mM) and enzyme (20 ~1) were incubated at 37°C. Samples were removed at intervals and analyzed for reducing sugars. In the case of laminarin, laminarin polyalcohol and acid- hydrolyzed laminarin polyalcohol, liber- ated glucose was also determined specifi- cally by using glucose oxidase. The total extent of hydrolysis achieved in 24 h (and in the case of laminarin and lichenin, also 70 h) was determined. The nature of the products produced by action of the enzyme on the various substrates was ascertained by chromatography of samples removed from the digests after incubation for 24 h. The results are shown in Table II and Fig. 8 (a and b).
The kinetic constants for the (1 --t 3)-p- glucanase acting on laminarin and lichenin were determined by measuring initial reac- tion rates in digests (1.0 ml) containing substrate (0.1-5.0 mg/ml), acetate buffer (pH 4.5, final concentration 25 mM) and enzyme solution (10 ~1) incubated at 37°C. Values for K, from Lineweaver-Burk dou- ble reciprocal plots (Fig. 9), were 1.22 mg/ml and 2.22 mg/ml for laminarin and lichenin respectively. The maximum veloc- ities of enzyme action on laminarin and lichenin were 2.70 and 1.49 pmoles glu- cose/ml enzymelmin, respectively.
A control experiment in which the en-
zyme was incubated with glucose (5%) for 24 h showed the absence of any detectable oligosaccharide formation indicating that the enzyme does not catalyze reversion reaction. Neither was any evidence found from paper chromatographic examination of the products of enzyme action on its various substrates, to suggest that the enzyme possesses transglycosylase activ- ity.
Anomeric form of liberated glucose. To determine whether action, of the enzyme proceeded with retention or inversion of anomeric configuration, the hydrolysis of laminarin in a digest containing substrate (5.0 mg), acetate buffer (pH 5.6, final concentration 5 mM) and enzyme solution (400 ~1) was followed polarimetrically. An increase in optical rotation during incuba- tion, decreasing to the equilibrium value of a mutarotated glucose solution on addition of a drop of ammonium hydroxide solution indicated that the enzyme released glucose with the cu-configuration.
Inhibition of purified (1+3)-/3-glucanase. The effect of a number of metal ions and compounds known to inhibit other fi- glucanases was determined by measuring the activity of the enzyme in digests of composition: laminarin (2.5 mg), acetate buffer (final concentration 100 mM) and enzyme (10 ~1) in a total volume of 1.0 ml,
1
80
-@l--l
s - 60 >I
z
; 4 40
f zL-!d 10 20 30 40 50 60 70
Temperature of Prelncubatlon (“0
FIG. 7. Heat stability of Helix pomatia (14)-j3- glucanase. Enzyme activities were determined at 37°C after heat treatment at the temperatures indi- cated for 30 min, and are expressed relative to the activity in a control digest incubated at 37°C.

SNAIL EXO-P-GLUCANASE 171
TABLE II SUBSTRATE SPECIFICITY OF Helix pomatia Exo-(1 + 3)-8-GLUCANASE'
Substrate Predominant linkages present
(all 8-W=)
Initial rate of hydrolysis (pg glucose/min)
Average degree of polymerization of
products after 24 h
Product(s)
Laminaribiose l-3 465 1.0 Glucose Laminaritriose l-3 471 1.1 Glucose Laminaritetraose l-3 502 1.0 Glucose Laminaripentaose l-3 485 1.0 Glucose Laminarin l-3 320 1.8 (1.1 after 70 h) Glucose, gentiobiose
and traces of higher oligosaccharides
Pachyman l-3 very low N.D.b Glucose (trace) Yeast glucan l-3 very low N.D. Glucose Lichenin l-3; l-4 163 4.2 (1.7 after 70 h) Glucose Barley glucan 1+3;1-4 15 6.7 Glucose Oat glucan l-3;1+4 N.D. 4.0 Glucose Gentiobiose l-6 N.D. N.D. Glucose Luteose 1-6 N.D. N.D. Glucose CM-Cellulose 144 very low 44 Glucose (trace) Cellodextrin 144 N.D. N.D. Glucose
0 The following were not attacked: (1 + 4)-&mannan, (1 --t 4)-b- and (1 + 3)-b-xylans, cellobiose. b N.D. indicates not determined.
enzyme action being initiated by addition of substrate, after preincubation of all other constituents of the digests for 15 min. Appropriate control digests were included to correct for any effect of the species being tested on the reducing power measure- ments. The results are shown in Table III.
DISCUSSION
Snail digestive juice provides an alterna- tive to microbial enzyme systems for use in studying the mechanism of enzymic degra- dation of cellulose, and the enzymic lysis of the cell walls of living yeasts. Crude snail juice contains an extremely complex mix- ture of glycosidases (2, 3) and it has there- fore been necessary to develop methods for separation of these closely related en- zymes, prior to examination of their prop- erties. This work has been simplified by the development of affinity binding techniques for the purification of ,&glucanases (7, 8, 10, 24, 25), but the enzyme which is the subject of the present paper could not be purified by such methods. However a com- bination of conventional ion-exchange and molecular-sieve chromatography tech- niques, summarized in Table I, yielded a
homogeneous preparation of the enzyme. It has not been possible to calculate an over- all purification factor for the enzyme, be- cause of the multiplicity of (1 -+ 3)+gluca- nases present in the starting material. Removal of the other (1 --+ 3)-/3-glucanases is a factor contributing to the apparently low yield of the enzyme; it is estimated that the recovery corresponds to approxi- mately 50% of the amount of the enzyme in the starting material.
Action of the enzyme on all substrates results in production of glucose as the sole or major chromatographically-mobile product (Table II). Measurement of the degree of polymerization of the products after action on laminarisaccharide sub- strates shows that complete degradation into glucose takes place (Table II). The degree of polymerization of the products resulting from action on laminarin also approaches unity after extended incuba- tion times. Analysis of the products during the hydrolysis of laminarin, by using both a copper reducing method and the glucose oxidase method (Fig. 8b), showed glucose to be the primary product of enzyme ac- tion. These observations suggest a exo
172 MARSHALL AND GRAND
20 40 60 so 100 120 Id0
Dllrat10n of lncubatlon ,m,n, D”rarlOn Of lncubatlon (mln )
FIG. 8. (a) Time course of hydrolysis of laminarin (O), lichenin (0) and barley glucan (x) by purified Helix pomatia (1+3)-&glucanase (b) Time course of hydrolysis of laminarin (0, O), oxidized laminarin (m, 0) and oxidized laminarin treated with mild acid (‘I, V) by purified Helix pomatia (l-3)-@-glucanase. The filled symbols show the results obtained using the glucose oxidase method and the open symbols using the Nelson-Somogyi method for glucose determina- tion.
‘/[s] Cm 0-Y m I )
FIG. 9. Lineweaver-Burk plot of reciprocal of ini- tial reaction rate (V) against reciprocal of substrate concentration ([Sl) for Helix pomatia exo-( l&+3)-& glucanase acting on laminarin (0) and lichenin (A).
TABLE III
EFFECT OF VARIOUS REAGENTS ON Acrrvrr~ OF Helix pomatia Exo-(1 + 3)-fl-GLUCANAsF
Reagent* Inhibition (%)
Mercuric chloride 47 Silver nitrate 41 Ferric chloride 39 Copper sulfate 41 Ammonium molybdate 28 Potassium cyanide 9 Phenyl mercuric nitrate 23 Gluconic acid d-lactone 96 N-Bromosuccinimide 96
“The following were without effect: lead acetate, EDTA, cobalt chloride, zinc sulfate, manganese sul- fate, sodium chloride, calcium chloride, cysteine hy- drochloride, oxidized glutathione, p-chloromercuriben- zoate, 2-hydroxy-5-nitrobenzylbromide, iodoacetic acid, &mercaptoethanol, sodium fluoride, sodium azide.
action pattern for the enzyme. Action of the enzyme on laminarin is blocked by periodate oxidation of the substrate, a method first used by Nelson et al. (11) to demonstrate the exo nature of a microbial nonreducing chain ends. These latter ob-
(1 -+ 3)-/3-glucanase, and substantially re- stored after mild acid treatment which regenerates intact glucose units at the

SNAIL EXO-B-GLUCANASE 173
servations confirm that the enzyme we have purified is an exo-( l&3)-/3-glucanase. The traces of oligosaccharides produced during action on laminarin are presumed to arise from reducing, or mannitol-termi- nated, chain. ends.
Helix pomatia exo-( 1 + 3)-/3-glucanase is rather unspecific, having the ability to cleave (1 -+ 4)-p- and (1 + 6)-&glucosidic linkages in addition to (1 -+ 3)-P-linkages. Thus the mixed linkage (l-3; l&4)-/3- glucan, lichenin, is degraded at half the rate of laminarin, and to an extent ap- proaching completion. Comparison of the kinetic constants of the purified exo- (1+3)-/3-glucanase for laminarin and li- chenin (K, values 1.22 mg/ml and 2.22 mg/ml; V values 2.70 pmoles glucose re- leased/ml enzyme/min and 1.49 pmoles glucose released/ml enzyme/min, for lami- narin and lichenin, respectively) shows the mixed-linkage glucan to be only a slightly poorer substrate for the enzyme than is laminarin. Barley and oat glucans, al- though somewhat more slowly hydrolyzed than laminarin and lichenin, are neverthe- less also extensively degraded. The lower rate of action on the cereal glucans than on lichenin is difficult to explain in view of the indication that they contain a rather smaller proportion (cu. 50%) of (l-4)- linkages than does lichenin (cu. 70%), al- though it is possible that this may be explicable on the basis of differences in the arrangement of the various types of gluco- sidic linkages in these polysaccharides (5). The less than complete conversion of the “mixed-linkage” substrates into glucose, as apparent from the degrees of polymeri- zation of the products of action on these substrates, indicates the presence in these substrates of some structural feature which resists enzyrne action. As yet, however, we do not have any information to suggest what the nature of such a structural feature might be.
The specificity of the Helix pomatia exo-P-glucanase we have purified appears to be less rigid than that of all the other exo-/3-glucanases which have been reported previously. Thus the exo-( l&3)-ggluca- nase from .Basidiomycetes QM806 (26), and those from various yeasts (27-29) do
not degrade the mixed linkage (l-3; l-4) glucans; that from Euglena grucilis (30) releases only small amounts of glucose from these substrates. These enzymes de- grade (1 + 3; 1 -+ 6)-glucans by bypassing the (1 + 6)-linkages which are released in the form of gentiobiose. The presence of gentiobiose as a product after action of the enzyme on laminarin suggests that the Helix pomatia also acts in this way, al- though subsequent hydrolysis of the genti- obiose may take place. While the Helix pomatia enzyme differs from the yeast exo-P-glucanases in being able to degrade lichenin and cereal glucans, it bears certain resemblances to these enzymes in its abil- ity to degrade ,f3-( 1+6)-linked glucans. There are indications that one of the yeast enzymes, that produced by Hansenula wingei (29), can also degrade b-(1+4)- linked glucose polymers so that the Helix pomatia enzyme bears, qualitatively at least, more similarity to this enzyme than any of the others. It is clear that a number of exo-( 1+3)-P-glucanases exist, each dif- fering in substrate specificity, the Helix pomatia enzyme having the broadest speci- ficity yet found. An analogous situation exists in the case of exo-acting (1 -+ 4)-a- glucanases, enzymes of this type differing in their ability to act on (1 + 3) - and (1 + 6)-cY-glucosidic linkages (31).
In addition to specificity differences, the Helix pomatia enzyme differs in molecular weight from other enzymes of this type for which values have been reported (82,000 as compared with 51,000 for the Basidi- omycetes enzyme (32) and 20,000-40,000 for the yeast enzymes (29)). The molecular weight of Helix pomatia exo-(l-3)-/3- glucanase is considerably higher than that of most endo-( 1+3)-P-glucanases which have been studied, including that from Helix pomatia, which lie in the range IO-30,000. This situation, namely exo- acting enzymes having molecular weights considerably higher than the corresponding exo-acting enzymes, is very common (33, 34). Unlike a number of endo-acting (l&3)- /3-glucanases (10, 35), the Helix porn&u (l&3)-/3-glucanase does not show any tend- ency to lose activity in dilute solution, and is not stabilized by human serum

174 MARSHALL AND GRAND
albumin. The temperature stability (stable up to 50°C) and optimum temperature (55°C) and susceptibility to acid inactiva- tion suggest that the enzyme does not possess any unusual stability properties.
While certain microbial exo-( l-+3)-@- glucanases are reportedly activated by manganese, cobalt and ferric ions (36, 37), the Helix pomatia enzyme is unaffected by manganese or cobalt, and markedly inhibited by ferric ions. A number of other workers have failed to demonstrate d:.o- (l&3)-P-glucanase activation by manga- nese and cobalt ions (32, 3%40), and ob- served Fe3+ inhibition of the enzymes (39). The lack of inhibition of the Helix pomatia enzyme by cysteine, mercaptoethanol, oxi- dized glutathione and p-chloromercuriben- zoate makes it seem unlikely that disulfide or thiol groups are involved in enzyme activity; the effect of phenylmercuric ni- trate may not be meaningful since organic mercurial compounds may inhibit other enzymes which do not depend on thiol groups for activity (41-42). The inhibition by N-bromosuccinimide might indicate the participation of tyrosine, tryptophan or histidine residues in the activity of the enzyme. The involvement of tryptophan is, however, unlikely in view of the ineffective- ness of 2-hydroxy 5-nitrobenzylbromide as an inhibitor. The inhibition by mercuric, cupric, ferric and silver ions may support the involvement of histidine (43). Like many other glycosidases (10, 44-46) the Helix pomatia exo-(1 -+ 3)-P-glucanase is strongly inhibited by ammonium molyb- date but the mechanism of inhibition by this reagent remains to be determined.
The classification of enzymes releasing glucose from oligosaccharide and polysac- charide substrates has been discussed by Reese and co-workers (47). It is not possible to classify the Helix pomatia enzyme unambiguously on the basis of the relative rates of action on (l-3)-P-linked sub- strates of different chain lengths. The in- version of anomeric configuration which takes place during hydrolysis indicates that the enzyme is an exo-P-glucanase. However, the inhibition by low concentra- tions of gluconic acid Slactone, which has been considered an indication of ,&glucosi-
dase activity argues against this conclu- sion. Thus it is not possible to classify the enzyme we have examined unambiguously by using the criteria of Reese and co-work- ers. This is a further difference between the Helix pomatia enzyme and other enzymes of this general type. We have preferred to consider the enzyme as an exo-P-glucanase because of its action on a wide range of P-glucan substrates, compared with its limited action on disaccharide substrates.
REFERENCES
1. GRAND, R. J. A., AND MARSHALL, J. J. (1975) Comp. Biochem. Physiol. B, in press.
2. HOLDEN, M., AND TRACEY, M. V. (1950) Biochem. J. 47, 407-414.
3. MYERS, F. L., AND NORTHCOTE, D. H. (1958) J. Exp. Biol. 35, 639-648.
4. ANDERSON, F. B., AND MILLBANK, J. W. (1966) Biochem. J. 99, 682-687.
5. MARSHALL, J. J. (1974) Aduan. Carbohyd. Chem. Biochem. 30, 257-370.
6. MARSHALL, J. J. (1975) Advan. Carbohyd. Chem. Biochem. 32, in press.
7. MARSHALL, J. J. (1973) Comp. Biochem. Physiol. 44B, 981-988.
8. MARSHALL, J. J. (1973) Carbohyd. Res. 26, 274-277.
9. MARSHALL, J. J. (1973) Anal. Biochem. 53, 191-198.
10. MARSHALL, J. J. (1974) Carbohyd. Res., 34, 289-305.
11. NELSON, T. E., SCALE~I, J. V., SMITH, F., AND KIRKWOOD, S. (1963) Can. J. Chem. 41, 1671-1678.
12. LOWRY, 0. H., ROSEBROUGH, N. J., FARR, A. L., AND RANDALL, R. J. (1951) J. Biol. Chem. 193, 265-275.
13. ROBYT, J. F., AND WHELAN, W. J. (1972) Anal. Biochem. 45, 510-516.
14. LLOYD, J. B., AND WHELAN, W. J. (1969) Anal. Biochem. 30, 467-470.
15. DUBOIS, M., GILLES, K. A., HAMILTON, J. K., REBERS, P. A., AND SMITH, F. (1956) Anal. Chem. 28, 350-356.
16. “Gel Chromatography,” Bio-Rad Laboratories, Richmond, California (1971).
17. “Sephadex Gel Filtration in Theory and Prac- tice,” Pharmacia Fine Chemicals, Uppsala, Sweden.
18. “Whatman Advanced Ion-Exchange Celluloses. Laboratory Manual,” W. & L. Balston, Maid- stone, Kent, England.
19. “Sephadex Ion Exchangers. An Outstanding Aid in Biochemistry,” Pharmacia Fine Chemicals, Uppsala, Sweden.

SNAIL EXO-,%GLUCANASE 175
20. ORNSTEIN, L. (1964) Ann. N.Y. Acad. Sci. 121, 321-349.
21. DAVIS, B. J. (1964) Ann. N.Y. Acad. Sci. 121, 404-427.
22. WEBER, K., AND OSBORN, M. (1969) J. Biol. Chew 244, 4406-4412.
23. TREVELYAN, W. E., PROCTER, D. P., AND HARRISON, J. S. (1950) Nature (London) 166, 444-445.
24. MARSHALL, J. J. (1973) Biochem. Sot. Z’rans. 1, 143-145.
25. MARSHALL, J. J. (1973) J. Chromatogr. 76, 257-260.
26. NELSON, T. E., JOHNSON, J., JANTZEN, J., AND KIRKWOOD, S. (1969) J. Biol. Chem. 244, 5972-5980.
27. TINGLE, M. A., AND HALVORSON, H. 0. (1971) Biochim. Biophys. Acta 250, 165-171.
28. BROCK, T. D. (1965) Biochem. Biophys. Res. Commun. 19, 623-629.
29. ABD-EL-AL, A. T. H., AND PHAFF, H. J. (1968) Biochem. J. 109, 347-360.
30. BARRAS, D. R., AND STONE, B. A. (1969) Biochim. Biophys. Acta. 191, 342-360.
31. MARSHALL, J. J. (1974) Cereal Sci. Today 19,389. 32. HOUTARI, F. I., NELSON, T. E., SMITH, F., AND
KIRKWOOD, S. (1968) J. Biol. Chem. 243, 952-956.
33. AHLGREN, E., ERIKSSON, K-E., AND VESTERBERG, 0. (1967) Acta Chem. Stand. 21, 937-944.
34. MANNERS, D. J., AND MARSHALL, J. J. (1973) Phytochemistry 12, 547-553.
35. MOORE, A. E., AND STONE, B. A. (1972) Biochim. Biophys. Acta 258, 238-247.
36. FELLIG, J. (1960) Science 131, 832. 37. CHESTERS, C. G. C., AND BULL, A. T. (1963)
Biochem. J. 86, 38-46. 38. VOGEL, K., AND BARBER, A. A. (1968) J. Protozool.
15, 657-662. 39. KAJI, A., OHSAKI, T., AND YOSHIHARA, 0. J. Agr.
Chem. Sot. Jap. 45, 278-283. 40. MOORE, A. E., AND STONE, B. A. (1972) Biochim.
Biophys. Acta 258, 248-264. 41. FISCHER, E. H., AND HASELBACH, C. H. (19.51) Helu.
Chim. Acta 34, 325-334. 42. SOHLFX, M. R., SEIBERT, M. A., KREKE, C. W.,
AND COOK, E. S. (1952) J. Biol. Chem. 198, 281-291.
43. GURD, F. R. N., AND WILCOX, P. E. (1956) Aduan. Protein Chem. 11, 311-427.
44. MACWILLIAM, I. C., AND HARRIS, G. (1959) Arch. Biochem. Biophys. 84, 442-454.
45. MANNERS, D. J., AND SPARRA, K. L. (1966) J. Inst. Brew. 72, 360-365.
46. MANNERS, D. J., AND ROWE, K. L. (1969) Car- bohyd. Res. 9, 107-121.
47. REESE, E. T., MAGUIRE, A. H., AND PARRISH, F. W. (1968) Can. J. Biochem. 46, 25-34.





























