Embed Size (px)
Citation preview

INFECTION AND IMMUNITY, Aug. 1978, p. 605-6120019-9567/78/0021-0605$02.00/0Copyright © 1978 American Society for Microbiology
Vol. 21, No. 2
Printed in U.S.A.
Comparative Immunological Analysis of Host Plasma ProteinsBound to Bloodstream Forms of Trypanosoma
brucei SubspeciesPETER DIFFLEY
Department ofZoology, University ofMassachusetts, Amherst, Massachusetts 01003
Received for publication 5 June 1978
The presence, location, host specificity, identity, and quantity of rat plasmaproteins bound to bloodstream forms of Trypanosoma brucei subsp. brucei, T.brucei subsp. rhodesiense, and T. brucei subsp. gambiense were determined bya quantitative indirect fluorescent-antibody method and gel immunoassays. Flu-orescence differences between trypanosomes obtained from rats and mice andtreated with antiserum to normal rat plasma indicated that most, if not all, of thebound plasma proteins were host specific. Removal of plasma proteins by tryp-sinization of parasites provided evidence for their attachment to the surface ofthe parasite. The accreted proteins were found to be host albumin, immunoglobu-lin G (IgG), and complement (C3). The same quantities of these three plasmaproteins were present on T. brucei subspecies collected from normal rats at 2days postinfection, during low or peak parasitemias, or from cortisone-treatedrats. IgM could only be detected on parasites collected from normal rats at peakparasitemia. With the aid of rocket immunoelectrophoresis, host albumin andIgG were found to account for 0.2 and 0.05%, respectively, of the total solubleproteins of the bloodstream forms. It was concluded from this study that: (i) hostplasma proteins were bound to parasites early in the infection, suggesting amechanism of adaptation to the mammalian host; (ii) the surface-bound IgG wasnot the result of a specific immune response against the parasites but might bethe cause of C3 attachment; (iii) among the bloodstream forms of the three T.brucei subspecies, there were no differences in amounts ofsurface-bound albumin,IgG, or C3. A comparison between the present data dealing with T. bruceisubspecies, on the one hand, and the previously published results concerning T.congolense, on the other, revealed significant differences in the amounts of thehost plasma proteins attached to these hemoflagellates.
Of the three morphologically indistinguisha-ble trypanosomatid subspecies, Trypanosomabrucei subsp. brucei, T. brucei subsp. rhode-siense, and T. brucei subsp. gambiense, the for-mer infects livestock whereas the latter two areetiological agents of human trypanosomiasis.Host plasma proteins have been found in asso-ciation with bloodstream forms (BSF) of T. bru-cei subsp. gambiense (2, 25) and T. brucei subsp.brucei (20, 30). The types and quantities of theseproteins have not been fully characterized foreither organism. Heretofore, no reports havebeen published on the host plasma proteinsbound to T. brucei subsp. rhodesiense.
Further analysis of trypanosome-host plasmainteractions might provide clues as to mecha-nisms of infectivity, pathogenesis, and/or eva-sion of the host immune response by the para-sites. The present study was undertaken to iden-tify and quantitate host-specific plasma proteins
bound to the surface of BSF of T. brucei subsp.rhodesiense and to compare the host plasmaproteins attached to BSF of T. brucei subsp.rhodesiense to those found on the remainingtwo T. brucei subspecies and T. congolense.
MATERIALS AND METHODSParasites. The following strains of the three T.
brucei subspecies were employed: a T. brucei subsp.rhodesiense line, TRUM 125 (Trypanosome ResearchUniversity of Massachusetts), initiated from EATRO(East African Trypanosomiasis Research Organiza-tion) 1895; a T. brucei subsp. gambiense line, TRUM126, initiated from TREU (Trypanosome ResearchUniversity of Edinburgh) 1309, kindly provided by A.R. Gray, Centre for Tropical Diseases, University ofEdinburgh; and a line of T. brucei subsp. brucei,TRUM 106. The histories of isolation of the parentstrains EATRO 1895, TREU 1309, and TRUM 106,their rodent passages, and cultivation were previouslydescribed (14, 22, and H. A. Stanley, B. M. Honigberg,
605
on Novem
ber 19, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

606 DIFFLEY
and I. Cunningham, J. Protozool., in press, respec-tively).The T. brucei subsp. brucei and T. brucei subsp.
rhodesiense BSF were pleomorphic isolates whichproduced several peak parasitemias before killing theirrodent hosts within 30 days. The monomorphic strainof T. brucei subsp. gambiense caused low blood par-asitemias in rats which did not succumb to the infec-tion within 30 days postinfection.When needed for immunological tests, BSF were
inoculated into rodents via the intraperitoneal route.The parasites were collected in heparinized blood (10U/ml of blood) and separated from host cells on adiethylaminoethyl-cellulose (DE-52, Whatman Co.)column using phosphate-buffered saline (PBS)-glu-cose (6:4, vol/vol; pH 8) as the suspending solution(18). The parasites were then washed in five changesof cold PBS-glucose by centrifugation (2,000 x g, 15min) at 4VC before being processed for quantitativeindirect fluorescent-antibody (QIFA) analysis or as asoluble antigen preparation used in gel immunoassays.
Hosts. Outbred adult male rats (Holtzman Co.,Madison, Wis.) were used for T. brucei subsp. bruceiand T. brucei subsp. rhodesiense infections, whereas14- to 20-day-old outbred male rats (Rat Labs, Belch-ertown, Mass.) served as hosts for T. brucei subsp.gambiense. For some experiments, parasites were col-lected from rats that had been immunosuppressed byintramuscular injections of hydrocortisone acetate(400 mg/kg of animal weight) by previously describedmethods (P. Diffley and B. M. Honigberg, J. Parasitol.,in press). The mice used in all experiments were adultfemales (CF-1, Carworth Farms, Wilmington, Mass).Serum sources. All prepared and purchased anti-
sera were the same as used to characterize rat plasmaprotein accretion on T. congolense (8; Diffley andHonigberg, in press). The procedure for absorption ofthe antiserum to normal rat plasma (NRP) with NRPhave also been described (8). Normal human serum(NHS), used in neutralization tests, was drawn fromthe author on the days of the tests.
Neutralization tests. Although not an infalliblecriterion, NHS neutralization tests have been used todifferentiate T. brucei subsp. brucei from T. bruceisubsp. rhodesiense. A method based on that describedby Hawking (16) was employed to determine if the T.brucei subsp. brucei and T. brucei subsp. rhodesiense,examined in the present study, were susceptible toneutralization by NHS. Trypanosomes were collectedin citrated blood (0.6%/ml of blood) from mice at 2 to3 days postinfection. Aliquots (0.35 ml) of the parasi-tized blood were mixed with 0.7 ml of NHS or withnormal rat serum. The parasite suspensions were in-cubated for 30 min at room temperature before beingadministered intraperitoneally (0.2 ml) to 9 mice persuspension. Infections were monitored by microscopicexamination of tail blood for 30 days postinfection.QIFA method. All rat infections were initiated by
injections of BSF obtained from mice. BSF of T.brucei subsp. rhodesiense were collected from rats atpeak parasitemia on two occasions (postinfection days7 and 12) and from an early infection (postinfectionday 2). Parasites were also obtained from hydrocorti-sone-treated rats during a fulminating parasitemia (5-day infection). Rat-derived BSF of T. brucei subsp.brucei and T. brucei subsp. gambiense were collected
at low parasitemias, the former at postinfection day 2and the latter at 14 days postinfection.
Control preparations used in QIFA analysis were T.brucei subsp. rhodesiense BSF derived from 3- to 5-day infections in mice, completely transformed cultureprocyclics, and rat-derived T. brucei subsp. rhode-siense trypsinized by the method recommended byStanley et al. (in press). Briefly, trypsinization pro-ceeded as follows. Trypanosomes were collected fromrats at 7 days postinfection and separated from bloodcomponents. The cells were suspended in 1 ml ofPucks solution containing 0.55 mg of trypsin and in-cubated at 37°C for 30 min. After incubation, thetrypsinized organisms were washed three times in coldPucks solution and processed for QIFA analysis.The procedures for antigen fixation and fluorescent-
antibody staining have been described previously (8).Briefly, trypanosomes suspended in PBS-glucose werefixed with 2.5% Formalin and affixed to slides. Trypan-osome smears were covered first with unconjugatedrabbit anti-NRP serum or normal rat serum dilutedwith 0.01 M PBS (pH 7.4). After a 2-h incubation, theunbound antibodies were removed by washing in PBS;the smears were then overlaid with 1:40 PBS dilutionsof the fluorescein isothiocyanate-conjugated goat anti-rabbit immunoglobulin G IgG) serum. At the end ofthe 2-h incubation, the preparations were washed andstored at 4°C.The ultramicrofluorometer, filters, and ratio record-
ing equipment used for QIFA measurements weredescribed in previous reports (8, 10). All fluorescencemeans were based on 42 determinations of individualorganisms made in two preparations, 21 per smear.Since the amount of surface area of the cell influencessuch readings, only short, stumpy forms of T. bruceisubsp. rhodesiense and T. brucei subsp. brucei, withapproximately the same size, were used. The surfaceareas of the T. brucei subsp. gambiense long, slenderforms and T. brucei subsp. rhodesiense culture formswere measured by planimetry, and their fluorescencereadings were adjusted to those that would be re-corded from short, stumpy forms. Fluorescence meanswere compared statistically by the paired t test or,when appropriate, by analysis of variance (at the 1%level of significance). Whenever significant F varianceratios were obtained, the means were ranked by Tu-key's test (5% level).Gel immunoassays. A soluble antigen extract was
derived from T. brucei subsp. rhodesiense BSF col-lected from 10 rats at peak parasitemia (5 to 8 dayspostinfection) as previously described (Diffley andHonigberg, in press). Briefly, trypanosomes, sus-pended in blood-free PBS-glucose with 0.8 mM phen-ylmethylsulfonylfluoride (protease inhibitor) were dis-rupted by freeze-thawing in a dry ice-alcohol bath.Insoluble material was removed from the lysed-cellsuspension by centrifugation (12,000 x g, 30 min). Theparticle-free supernatant fluid was dialyzed againstPBS at 4°C and lyophilized. The antigen preparationwas dissolved in sterile distilled water before use.Concentrations of proteins in all antigen preparationswere estimated by the Folin phenol method (21).As described in detail elsewhere (8), immunoelec-
trophoresis (IEP) was performed in 0.5% agarose at4°C for 3 h using a constant amperage of 10 mA perplate. A 0.05 M Veronal buffer (pH 8.4) was used. Gel
INFECT. IMMUN.
on Novem
ber 19, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
HOST PLASMA PROTEINS ON T. BRUCEI SUBSPECIES
diffusion and quantitative IEP methods have also beendescribed previously (Diffley and Honigberg, in press).Diffusion reactions were carried out at 40C in 1%agarose dissolved in distilled water, and rocket IEPwas at 40C using 4 V/cm for 18 h. Rocket IEP gelsconsisted of 1% agarose mixed with monospecific an-tiserum at the final dilution of 1%. A 0.087 M Veronolbuffer (pH 8.7) was used. For determinations of theIgG content in the parasite extract, both standardsand parasite antigens were carbamylated for 4 h at370C by the method of Weeke (32).
RESULTS
Neutralization tests. Parasites were ob-served in all of the 18 mice infected with T.brucei subsp. rhodesiense that had been incu-bated in either NHS or rat serum; the averageprepatent period equaling 3.4 + 0.8 days. TheBSF of T. brucei subsp. brucei, incubated in ratserum, also proved to be infective for all ninemice, with an average prepatent period of 2 ±0.5 days. Parasites were never observed in thenine animals injected with the T. brucei subsp.brucei that had been incubated in NHS. Evi-dently, the T. brucei subsp. brucei and T. bruceisubsp. rhodesiense isolates used in this studycould be differentiated by the NHS neutraliza-tion test.Presence of surface-bound plasma pro-
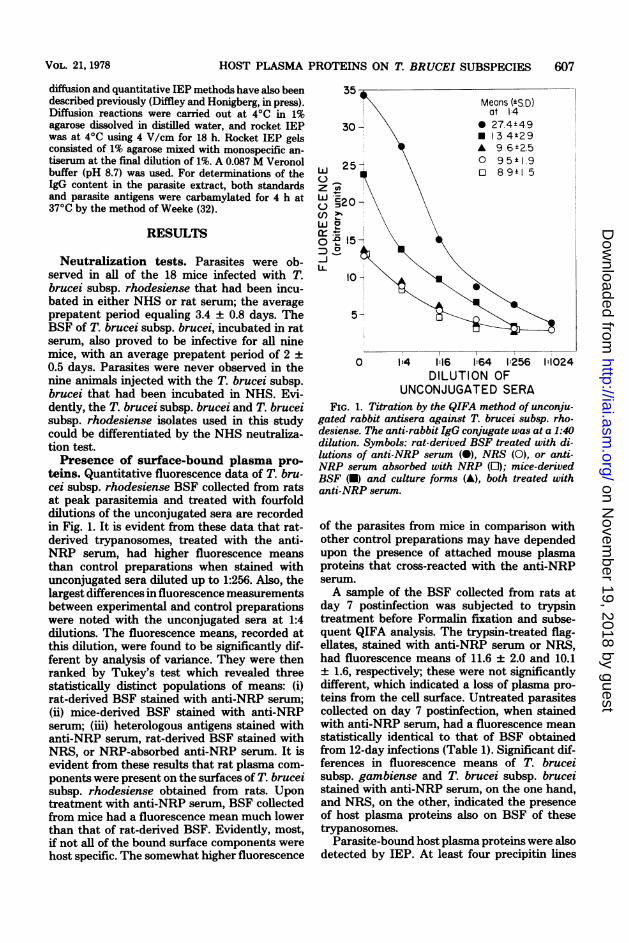
teins. Quantitative fluorescence data of T. bru-cei subsp. rhodesiense BSF collected from ratsat peak parasitemia and treated with fourfolddilutions of the unconjugated sera are recordedin Fig. 1. It is evident from these data that rat-derived trypanosomes, treated with the anti-NRP serum, had higher fluorescence meansthan control preparations when stained withunconjugated sera diluted up to 1:256. Also, thelargest differences in fluorescence measurementsbetween experimental and control preparationswere noted with the unconjugated sera at 1:4dilutions. The fluorescence means, recorded atthis dilution, were found to be significantly dif-ferent by analysis of variance. They were thenranked by Tukey's test which revealed threestatistically distinct populations of means: (i)rat-derived BSF stained with anti-NRP serum;(ii) mice-derived BSF stained with anti-NRPserum; (iii) heterologous antigens stained withanti-NRP serum, rat-derived BSF stained withNRS, or NRP-absorbed anti-NRP serum. It isevident from these results that rat plasma com-ponents were present on the surfaces of T. bruceisubsp. rhodesiense obtained from rats. Upontreatment with anti-NRP serum, BSF collectedfrom mice had a fluorescence mean much lowerthan that of rat-derived BSF. Evidently, most,if not all of the bound surface components werehost specific. The somewhat higher fluorescence
Z UnLLJ *2
Oh._O) =
-
LL
0 1:4 1:16 1:64 1:256 1:1024DILUTION OF
UNCONJUGATED SERAFIG. 1. Titration by the QIFA method of unconju-
gated rabbit antisera against T. brucei subsp. rho-desiense. The anti-rabbit IgG conjugate was at a 1:40dilution. Symbols: rat-derived BSF treated with di-lutions of anti-NRP serum (0), NRS (0), or anti-NRP serum absorbed with NRP (E); mice-derivedBSF (-) and culture forms (A), both treated withanti-NRP serum.
of the parasites from mice in comparison withother control preparations may have dependedupon the presence of attached mouse plasmaproteins that cross-reacted with the anti-NRPserum.A sample of the BSF collected from rats at
day 7 postinfection was subjected to trypsintreatment before Formalin fixation and subse-quent QIFA analysis. The trypsin-treated flag-ellates, stained with anti-NRP serum or NRS,had fluorescence means of 11.6 ± 2.0 and 10.1+ 1.6, respectively; these were not significantlydifferent, which indicated a loss of plasma pro-teins from the cell surface. Untreated parasitescollected on day 7 postinfection, when stainedwith anti-NRP serum, had a fluorescence meanstatistically identical to that of BSF obtainedfrom 12-day infections (Table 1). Significant dif-ferences in fluorescence means of T. bruceisubsp. gambiense and T. brucei subsp. bruceistained with anti-NRP serum, on the one hand,and NRS, on the other, indicated the presenceof host plasma proteins also on BSF of thesetrypanosomes.
Parasite-bound host plasma proteins were alsodetected by IEP. At least four precipitin lines
VOL. 21, 1978 607
on Novem
ber 19, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

608 DIFFLEY
formed between the well containing the solubleantigen extract of T. brucei subsp. rhodesienseand that with anti-NRP serum (Fig. 2).Identification of surface-bound plasma
proteins. The QIFA measurements of trypan-osomes stained with 1:4 dilutions of rabbit anti-sera to rat proteins are presented in Table 1.Statistically significant differences were ob-
served between the fluorescence means of con-trol preparations and those of the BSF stainedwith each of the following: antisera to rat albu-min, gamma globulin, IgG, and complement(C3). Upon treatment with these antisera, nosignificant differences were found among thefluorescence means of T. brucei subsp. rhode-siense, T. brucei subsp. brucei, and T. brucei
TABLE 1. Fluorescence means of T. brucei subspecies treated with antisera to ratplasma proteinsFluorescence mean (+SD) in arbitrary units' of cells after treatment with:
Days Antisera to ratTrypanosoma species Host postin-
fection GammaPlasma Albumin C3 globm IgG IgM
T. brucei subsp. rho- NMC 3-5 13.4 ± 2.9 _d 10.0 ± 1.9desiense NR 7f 11.6 ± 2.0 - - - 10.1 ± 1.6'
27.5 ± 4.9 _ - - - 16.3 ± 2.3 9.5 ± 1.912' 28.2 ± 5.0 19.1 + 2.1 19.1 ± 2.5 21.6 + 3.6 14.8 + 2.7 20.7 ± 3.2 10.2 ± 2.02 24.8 ± 3.6 20.0 ± 2.6 19.9 ± 2.1 20.7 + 3.3 14.0 + 2.0 11.0 + 2.0 9.9 + 1.7
IR' 5 24.2 ± 3.8 19.1 + 2.9 19.0 ± 2.8 - 14.6 ± 1.9 10.7 ± 1.8 10.1 ± 1.8T. brucei subsp. brucei NR 2h 25.3 ± 3.7 20.1 + 2.7 19.4 ± 2.6 20.0 ± 2.4 14.8 + 2.3 12.2 ± 1.8 10.8 + 2.2T. brucei subsp. gam- NR 14" 23.8 ± 3.2 19.5 ± 2.8 19.8 ± 3.4 - 14.9 + 2.6 11.7 ± 2.3 11.6 ± 2.2
bienseT. brucei subsp. rho- Culturei 9.6 ± 2.5 10.1 + 1.6 9.5 ± 1.5 9.6 ± 1.9 9.4 ± 1.6 9.7 + 1.8 11.0 ± 2.0
desienseI IIIIIII" Means of 42 individual determinations (21 per smear). The unconjugated rabbit antisera to rat plasma proteins were diluted
1:4 in PBS; the anti-rabbit IgG conjugate was used at 1:40 PBS-dilutions. SD, Standard deviation.Normal rabbit serum (control preparation) at a 1:4 dilution.
'Normal mice."-, Not tested.'Normal rats.' Parasites collected at peak parasitemia.X'Fluorescence means of trypsinized bloodstream forms.h Parasites collected at low parasitemias.'Immunosuppressed rats. The parasites were collected during a fulminating parasitemnia.i Culture forms; the heterologous antigen (control) preparation.
-
FIG. 2. Immunoelectrophoretic analysis of reactions between trypanosome antigens and NRP with anti-NRP serum. (Upper well) twenty micrograms of protein of normal rat plasma (N); (lower well) 190 pg ofprotein of the soluble antigen extract of T. brucei subsp. rhodesiense BSF (T).
INFECT. IMMUN.
ML2
on Novem
ber 19, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

HOST PLASMA PROTEINS ON T. BRUCEI SUBSPECIES 609
subsp. gambiense. From these results it wasconcluded that the same quantities of the hostplasma proteins were present on BSF of thethree subspecies. Further, the concentrations ofthese proteins were not altered on T. bruceisubsp. rhodesiense obtained from immunosup-pressed hosts, indicating that the surface-boundIgG was not the result of a specific immuneresponse against the parasites. Significant differ-ences in the IgM levels were found by QIFAanalysis among the various populations of T.brucei subsp. rhodesiense. The parasites ob-tained at peak parasitemia had appreciableamounts of this immunoglobulin bound to theirsurfaces (Table 1). On the other hand, T. bruceisubsp. rhodesiense collected at day 2 postinfec-tion, those derived from immunosuppressedhosts, as well as the T. brucei subsp. brucei andT. brucei subsp. gambiense preparations treatedwith anti-IgM serum, had fluorescence readingssimilar to those of controls. Evidently, there wasno surface-bound IgM on the BSF collected fromnormal rats during low parasitemias or from
immunosuppressed rats. African pathogenic try-panosomes produce several peaks of parasitemiain their mammalian hosts, and the decline inparasite populations has been attributed to avariant antigen-specific IgM response. There-fore, the IgM found on the parasites collected atpeak parasitemia probably represented a specificimmune response directed against the parasite.The presence of rat albumin, IgG, and C3 was
established also by gel diffusion reactions (Fig.3). The anti-IgG and anti-gamma globulin seraformed the same number of precipitin bands inreactions with the trypanosome antigens orNRP. Thus, the differences in fluorescencemeans of BSF treated with these two antisera(Table 1) in all probability reflected differencesin antibody titers.Quantitation of surface-bound albumin
and IgG. Rocket IEP was used to estimate theamounts of rat IgG and albumin present on theT. brucei subsp. rhodesiense BSF obtained fromrats. Averages of replicate tests indicated that 1mg of protein on the parasite soluble antigen
L~ ~'6a ;sS;AGSJ.. S.
~ ~ ~ rARSCt
4~~~~~~~~~~~~~~~~~~~~~~~~~~~~~S
FIG. 3. Gel diffusion tests involving a soluble antigen extract of T. brucei subsp. rhodesiense (PTr.) (0.9 mgofprotein) and antisera to rat: plasma (ARS), albumin (AAS), C3 (ACS), IgG (AGS), and gamma globulin(A -y S).
VOL. 21, 1978
on Novem
ber 19, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

610 DIFFLEY
extract contained 1.4 tig of albumin and 0.51 Mgof IgG. Apparently, these two proteins togetheraccounted for about 0.2% of the total solubleproteins of the BSF. It has been determined byothers (1, 4) that the surface coat of T. bruceisubsp. brucei accounts for about 10% of the totalprotein of the parasite. Thus, rat albumin andIgG together would represent less than 2% of thesurface coat protein of T. brucei BSF.
DISCUSSION
The presence of host-specific plasma proteinson the surface of T. brucei subsp. rhodesienseBSF was established. Surface-bound plasmaproteins were also detected by the QIFA methodon T. brucei subsp. brucei and T. brucei subsp.gambiense, supporting the conclusions based onother immunological tests reported previously(2, 20, 25, 30). Parasite-bound plasma proteinshave also been found on T. lewisi (6, 11), T.vivax (17), and T. congolense (8, 9). Negativeresults have also been published. Fruit et al.(12), using a fluorescent-antibody method toanalyze glutaraldehyde-fixed T. brucei subsp.brucei, were unable to detect host plasma pro-teins on the parasites. This failure may havebeen due to the method of fixation, which hasbeen considered unsuitable for use with fluores-cent-antibody methods in determinations of IgGon the surfaces of lymphocytes (28) and of allaccreted plasma proteins on T. congolense (un-published results).The plasma proteins identified on BSF of the
three T. brucei subspecies were albumin, IgG,and complement (C3). The relatively rapid ap-pearance, within 2 days postinfection, of theseproteins suggests that the accretion might rep-resent a mechanism of adaptation of the parasiteto its mammalian host. It has been proposedthat host plasma protein attachment is neces-sary for the adaptation of T. vivax to rodenthosts (7). The accreted proteins could be in-volved in membrane stabilization as has beenreported for other cells (11, 23) or might serve ametabolic need. With regard to metabolic func-tion, the albumin requirement in defined culturemedia for trypanosomatids (5, 28) should benoted. Further, D-mannose, D-galactose, and D-glucosamine, all components of the T. bruceisubsp. brucei variant antigen (1) are also foundin the Fc region of the IgG molecule.No evidence was found in this study, however,
for a positive relationship between parasite pop-ulation growth in the rodent host and theamount of accreted host plasma proteins. Allthree T. brucei subspecies have identicalamounts of IgG, albumin, and C3 on their sur-faces. The T. brucei subsp. gambiense isolate
INFECT. IMMUN.
produced in rodents a chronic infection charac-terized by fluctuating but generally low parasi-temias. On the other hand, inoculation of the T.brucei subsp. brucei and T. brucei subsp. rho-desiense isolates resulted in several high peakparasitemias, the last of which overwhelmed thehosts.The IgM, detected only on T. brucei subsp.
rhodesiense collected from rats at peak parasit-emia, was probably the result of a specific im-mune response. The IgG, however, found on themembers of all BSF populations tested, includ-ing those obtained from immunosuppressedhosts, appeared to be nonspecific. This result isin agreement with those of Bogucki and Seed (2)who reported the attachment of heterospecificIgG on the parasite surface of a T. brucei subsp.gambiense strain capable of causing an acutedisease in rodents.As measured by rocket IEP, 1 mg of protein
of the parasite extract contained 1.4 Mg of albu-min and 0.51 Mig of IgG. Using the same methodsfor parasite antigen extraction and the sameantisera for rocket IEP, Diffley and Honigberg(in press) reported that T. congolense had 4.3Mug of albumin and 0.9 Mg of IgG per mg of proteinof the parasite extract. Thus, it appears that T.congolense has three times more albumin andabout twice as much IgG as the T. brucei sub-species. A comparison between the QIFA read-ings of BSF, stained with the same dilutions ofanti-albumin or anti-IgG sera, indicates that, perunit surface area, T. congolense has 2.5 timesthe amount of albumin and 1.7 times the amountof IgG than the T. brucei subspecies. The twoquantitative immunological methods seem toyield closely comparable results.A comparison was also made between QIFA
measurements of the amounts of IgG and C3bound to T. brucei subsp. rhodesiense (presentstudy) and T. congolense, reported elsewhere(9). The latter organism has, per unit surfacearea, 1.7 times more IgG and 1.8 times more C3.These closely comparable ratios suggest a pos-sible correlation. The presence of nonspecificIgG on the parasites might cause subsequentcomplement fixation and C3 attachment. Withregard to this, immune complexes were detectedin the circulation of T. brucei subsp. brucei-infected rodents at 2 days postinfection (12).The complexes consisted of the variant antigen,the major component of the surface coat of theparasite, and immunoglobulins capable of fixingcomplement. At this early phase of infection, anonspecific attachment of IgG seems more likelythan binding by parasite-elicited antibodies. Im-mune complexes may have serious implicationswith regard to pathogenesis in African trypano-somiasis. Nagle et al. (23) attributed glomerulo-
on Novem
ber 19, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

HOST PLASMA PROTEINS ON T. BRUCEI SUBSPECIES 611
nephritis in T. brucei subsp. rhodesiense-in-fected rhesus monkeys to immune complex ac-tivation of the alternate complement pathway.If parasite-mediated complement depletion oc-curs during the course of African trypanosomi-asis, as has been suggested by low C3 levels ininfected humans (15), monkeys (23), and cattle(18), the hosts might be less prepared to combatthe trypanosome infection (26) and secondarymicrobial invasions.
Accretion of host plasma proteins might alsohave important implications for immunologicalaspects of African trypanosomiasis, e.g., carrier-mediated tolerance occurs when haptens arecoupled to albumin and IgM (13) and IgG (3,29). Possibly, the accreted plasma proteins arebound to specific sites on the surface of theparasite and such sites might represent constantand potentially immunogenic determinants. Thehost would be unable to elicit an immune re-sponse against these immunogens because theyare rendered tolerogenic by the attachment ofhost plasma proteins.
ACKNOWLEDGMENTS
I am grateful to B. M. Honigberg for his help and guidancein the course of the study.
This investigation was supported by Public Health Serviceresearch grant AI 00742-22 from the National Institute ofAllergy and Infectious Diseases. P. D. is a predoctoral traineesupported by Public Health Service training grant 1T32AI07109-01 from the National Institute of Allergy and InfectiousDiseases.
LITERATURE CITED
1. Allsopp, B. A., and A. R. Njogu. 1974. Monosaccharidecomposition of the surface glycoprotein antigens of Try-panosoma brucei. Parasitology 69:271-281.
2. Bogucki, M. S., and J. R. Seed. 1978. Parasite-boundheterospecific antibody in experimental trypanosomia-sis. RES J. Reticuloendothel. Soc. 23:89-101.
3. Chiller, J. M., G. S. Habicht, and W. 0. Weigle. 1971.Kinetic differences in unresponsiveness of thymus andbone marrow cells. Science 171:813-814.
4. Cross, G. A. M. 1973. Identification and purification of aclass of soluble surface proteins from Trypanosomabrucei. Trans. Roy. Soc. Trop. Med. Hyg. 67:261.
5. Cross, G. A. M., and J. C. Manning. 1973. Cultivationof Trypanosoma brucei sspp. in semi-defined and de-fined media. Parasitology 67:315-331.
6. D'Alesandro, P. A. 1972. Trypanosoma lewisi: produc-tion of exoantigens during infection in the rat. Exp.Parasitol. 32:149-164.
7. Desowitz, R. S., and H. J. G. Watson. 1953. Studies onTrypanosoma vivax. VI. The occurrence of antibodiesin the sera of infected sheep and white rats and theirinfluence on the course of infection in white rats. Ann.Trop. Med. Parasitol. 47:247-255.
8. Diffley, P., and B. M. Honigberg. 1977. Fluorescentantibody analysis of host plasma components on blood-stream forms of African pathogenic trypanosomes. I.Host specificity and time of accretion in Trypanosomacongolense. J. Parasitol. 63:599-606.
9. Du Buy, H. G., C. L. Greenblatt, J. E. Hayes, and D.R. Lincicome. 1966. Regulation of cell membranepermeability in Trypanosoma lewisi. Exp. Parasitol.
18:231-243.10. Dwyer, D. M. 1972. Analysis of the antigenic relationships
among Trichomonas, Histomonas, Dientamoeba, andEntamoeba. I. Quantitative fluorescent antibody meth-ods. J. Protozool. 19:316-325.
11. Dwyer, D. M. 1976. Immunologic and fine structure evi-dence of avidly bound host serum proteins in the surfacecoat of a bloodstream trypanosome. Proc. Natl. Acad.Sci. U.S.A. 73:1222-1226.
12. Fruit, J., D. Afchain, A. Petitprez, N. Van Meirvenne,D. Le Ray, D. Bout, and A. Capron. 1977. Antigenicanalysis of variant-specific components of Trypano-soma brucei brucei: localization on the surface coatwith labelled specific antibody. Parasitology 74:185-190.
13. Fruit, J., F. Santoro, D. Afchain, G. Duvallet, and A.Capron. 1977. Les immunocomplexes circulents dansla trypanosomaise Africaine humaine et experimentale.Ann. Soc. Belge Med. Trop. 57:257-266.
14. Gray, A. R. 1972. Variable agglutinogenic antigens ofTrypanosoma gambiense and their distribution amongisolates of the trypanosomes collected in different placesin Nigeria. Trans. Roy. Soc. Trop. Med. Hyg. 66:263-284.
15. Greenwood, B. M., and H. C. Whittle. 1976. Comple-ment activation in patients with Gambian SleepingSickness. Clin. Exp. Immunol. 24:133-138.
16. Hawking, F. 1976. Relative resistance to human plasmain Trypanozoon group. Trans. Roy. Soc. Trop. Med.Hyg. 70:504-512.
17. Ketteridge, D. S. 1970. The presence of host serumcomponents on the surface of rodent-adapted Trypa-nosoma vivax. J. Protozool. 17(Suppl):24.
18. Kobayashi, A., and I. R. Tizard. 1976. The response toTrypanosoma congolense infection in calves. Determi-nation of immunoglobulins IgG,, IgG2, IgM, and C3 andthe complement fixing antibody titers during the courseof infection. Tropenmed. Parasitol. 27:411-417.
19. Lanham, S. M., and D. G. Godfrey. 1970. Isolation ofsalivarian trypanosomes from man and other mammalsusing DEAE-cellulose. Exp. Parasitol. 28:521-534.
20. Le Ray, D. 1975. Structures antigeniques de Trypano-soma brucei (Protozoa:Kinetoplastida) analyse immu-noelectrophoretique et etude comparative. Ann. Soc.Belge Med. Trop. 55:134-292.
21. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J.Randall. 1951. Protein measurement with the Folinphenol reagent. J. Biol. Chem. 193:265-275.
22. Mendez, Y., and B. M. Honigberg. 1972. Infectivity ofTrypanosoma brucei subgroup flagellates maintainedin culture. J. Parasitol. 58:1122-1136.
23. Nagle, R. B., P. A. Ward, H. B. Lindsley, E. H. Sadun,A. J. Johnson, R. E. Berkaw, and P. K. Hilde-brandt. 1974. Experimental infections with African try-panosomes. VI. Glomerulonephritis involving the alter-nate pathway of complement activation. Am. J. Trop.Med. Hyg. 23:15-26.
24. Najjar, V. A. 1974. The physiological role of y-globulin,p. 129-178. In A. Meister (ed.), Advances in enzymology,vol. 41. John Wiley & Sons, Inc., New York.
25. Seed, J. R. 1974. Antigens and antigenic variability inAfrican trypanosomes. J. Protozool. 21:639-646.
26. Seed, J. R. 1977. Role of immunoglobulins in Trypano-soma brucei gambiense infections. Int. J. Parasitol.7:55-61.
27. Smit, J. W., C. J. L. M. Meijer, F. Decary, and T. M.Feltkamp-Vroom. 1974. Paraformaldehyde fixation inimmunofluorescence and immunoelectron microscopy.Preservation of tissue and cell surface membrane anti-gens. J. Immunol. Methods 6:93-98.
28. Steiger, R. F., and E. Steiger. 1977. Cultivation ofLeishmania donovani and Leishmania braziliensis indefined media: nutritional requirements. J. Protozool.
VOL. 21, 1978
on Novem
ber 19, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

612 DIFFLEY INFECT. IMMUN.
24:437-441.29. Venkataraman, M., M. Aldo-Bensen, Y. Borel, and
D. W. Scott. 1977. Persistence of antigen-binding cellswith surface tolerogens: isologous vs heterologous im-munoglobulin carriers. J. Immunol. 119:1006-1009.
30. Vickerman, K. 1972. Host-parasite interface of parasiticprotozoa, p. 71-91. In A. E. R. Taylor and R. Muller(ed.), Functional aspects of parasitic surfaces, vol. 10.Symposium of the British Society of Parasitologists.
Blackwell, Oxford.31. Walters, C. S., and H. N. Claman. 1974. Immunologic
reactions to haptens on autologous carriers. II. Induc-tion of hapten-specific tolerance and correlations be-tween antibody response and antigen-driven cell prolif-eration in vitro. J. Immunol. 113:645-653.
32. Weeke, B. 1968. Carbamylated human immunoglobulinstested by electrophoresis in agarose and antibody-con-taining agarose. Scand. J. Clin. Lab. Invest. 21:351-354.
on Novem
ber 19, 2018 by guesthttp://iai.asm
.org/D
ownloaded from





























