Embed Size (px)
Citation preview

Proc. Nati. Acad. Sci. USAVol. 86, pp. 5227-5231, July 1989Biochemistry
Identification of a monocyte specific pre-interleukin 1i8convertase activityMATTHEW J. KOSTURA*, MICHAEL J. Tocci, GUADALUPE LIMJuco, JAYNE CHIN, PATRICIA CAMERON,ANDREW G. HILLMAN, NICOLE A. CHARTRAIN, AND JOHN A. SCHMIDTDepartment of Biochemical and Molecular Pathology, Merck Sharp & Dohme Research Laboratories, P.O. Box 2000, Rahway, NJ 07065
Communicated by Edward M. Scolnick, March 13, 1989
ABSTRACT Interleukin 1 (IL-1) is a lymphokine secretedby monocytes in response to a variety of inflammatory stimuli.IL-1fB the predominant form of IL-1 produced by humanmonocytes, is synthesized as an inactive precursor of 31 kDaand is cleaved at Asp"'6-Ala"17 to yield a 17.5-kDa extracellularform. The exact cellular site of cleavage and mechanism ofsecretion is at present unknown. We have prepared cell-freepostnuclear extracts from freshly isolated human monocytes aswell as THP.1 cells, a human monocyte-like cell line, andvarious blood lymphocytes and fibroblast cell lines. Usingpre-IL-18 synthesized by in vitro transcription and translation,we have shown that only extracts derived from human mono-cytes and THP.1 cells were capable of cleaving precursor IL-1i8to authentic mature IL-1g.. Subcellular fractionation of theextracts suggested that the processing activity is found in thecytosol of monocytes or monocyte-like cell lines. The cleavageproduct of this protease is identical to authentic IL-1 as shownby mobility on SDS/PAGE and amino acid sequence analysisof the [3H]leucine-labeled product. The cleavage product is alsocapable of binding to the IL-1 receptor found on fibroblastmembranes. Finally, mutation of Asp'"' - Ala" rendered theIL-1,8 precursor resistant to cleavage by the processing activ-ity. We conclude that a protease activity found only in mono-cytes will specifically process IL-1p to an active form.
Interleukin 1,8 (IL-1,8) is a 17.5-kDa polypeptide hormonesynthesized and secreted in large quantities by stimulatedmonocytes. A number of studies have detailed the synthesisand secretion of IL-1 from activated monocytes. Giri et al. (1)provided the first substantive evidence that, in a mousemonocyte cell line, IL-1 was synthesized as a cell-associatedprecursor that could be chased into an extracellular 17-kDaform. Subsequently, reports emerged which suggested that a31-kDa form of IL-1i3 was associated with human monocytes(2-5) and that this material was cleaved to release the matureform (2, 4, 5). These studies were corroborated by cDNAsequence data from a number of species which indicated thatIL-1 mRNA encodes a larger protein than that identified asmature secreted IL-1 (6-10). As precursor IL-183 (pre-IL-1,j)is unable to bind to IL-1 receptors and is biologically inactive(11), some form of proteolytic processing is apparently re-quired to release active IL-1p8.While the kinetics of IL-1 synthesis and secretion has been
analyzed in some detail, little has been uncovered about themechanism by which IL-1 is synthesized, processed, andsecreted. Analysis ofthe predicted amino acid sequence frompre-IL-1,3 cDNA has not revealed the presence of a uniquehydrophobic signal sequence domain, common to most se-creted proteins (6-10, 12). The N-terminal amino acid ofmature monocyte IL-1p from humans has been sequenced bya number of investigators as Ala"17 (6, 13), suggesting that acleavage site exists between Asp'6 and Ala"7. While the first
116 residues may be considered a signal sequence of sorts, itis not recognized as such by otherwise competent endoplas-mic reticulum membranes (G.L., unpublished observation).Young et al. (14) showed that mature pre-IL-1P was notsecreted from hamster fibroblasts that were stably trans-formed with pre-IL-1,i cDNA. Instead, large amounts of theprecursor accumulated in the cytoplasm of the cell (14).Lomedico et al. (12) demonstrated that murine recombinantpre-IL-la does not appear to enter the lumen of caninepancreatic microsomes in vitro. Similarly, immunolocaliza-tion studies have demonstrated that pre-IL-1,8 accumulates inthe cytoplasm of lipopolysaccharide-stimulated humanmonocytes and was not found to be associated with the Golgiapparatus or the plasma membrane (3, 15). While not con-clusive, these data suggest that IL-1,8 is not synthesized andsecreted cotranslationally as is typical of most secretedproteins in mammalian cells (reviewed in ref. 16).The processing of IL-1f3 has recently been investigated by
using purified recombinant precursor as a substrate (5, 17).Hazuda et al. (5) showed that pre-IL-1f3, when added to intacthuman blood monocytes, was not cleaved or processed inany fashion, arguing against an extracellular site of process-ing. In another report, a potential pre-IL-1,8 cleavage activitywas identified in a pelletable compartment of KG-1 cells, aneutrophil-like cell line. This enzymatic activity was able togenerate IL-1 activity of similar size to authentic IL-1 froma partially purified pre-IL-1f3 substrate (17). However, theproducts were not sequenced and the site ofcleavage was notidentified.
In this report, we describe an in vitro processing system inwhich mature 17.5-kDa IL-1,8 can be generated by incubationof pre-IL-1,8 with partially fractionated extracts from humanmonocytes or monocyte-like cell lines. We show that IL-1processing enzymes are found in a soluble cytosolic com-partment of monocytes in addition to particulate fractions.However, we find that an activity that partitions into thecytosolic compartment is able to cleave pre-IL-1f3 to theauthentic mature form. The product is similar to authenticmature IL-1f3 in its ability to bind to IL-1 receptors andamino-terminal amino acid sequence. Finally, mutation oftheIL-18 precursor at a highly conserved aspartic acid residue atposition 116 yielded substrate that is incapable of beingcleaved by the monocyte extracts. The data suggest that asingle endoproteolytic cleavage is responsible for generationof mature IL-1,8.
MATERIALS AND METHODSCells and Tissue Culture. Human peripheral blood mono-
cytes were obtained from healthy donors by leukophoresis
Abbreviations: IL, interleukin; pre-IL-1,f, precursor IL-1,8; PMN,polymorphonuclear leukocyte.*To whom reprint requests should be addressed at: Merck Sharp andDohme Research Laboratories, P.O. Box 2000, R80N-A26, Rah-way, NJ 07065.
5227
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Dow
nloa
ded
by g
uest
on
Sep
tem
ber
7, 2
020

Proc. Natl. Acad. Sci. USA 86 (1989)
and purified by sedimentation through lymphocyte separa-tion media (Organon Teknika) followed by elutriation on aBeckman counterflow centrifuge as described (18). Mono-cytes were identified by labeling with anti-MACl antibodyfollowed by fluorescence-activated cell sorting (FACS) anal-ysis. Human THP.1 cells (19, 20) (American Type CultureCollection) were grown in suspension at 370C in Dulbecco'smodified minimal essential medium (Hazelton ResearchProducts, Reston, VA) with 10%o fetal calf serum (HyClone;defined sera with no detectable endotoxin).
Preparation of Radiolabeled Precursor IL-1p8. A 1.5-kilobase (kb) cDNA clone containing the entire coding se-quence of pre-IL-13 was inserted into EcoRI/Pst I-cleavedpGEM-3 plasmid DNA (Promega Biotec) and propagated inEscherichia coli according to standard methods (21). Purifiedplasmid was linearized with Pst I and then transcribed byusing a T7 RNA polymerase in vitro transcription system(Promega Biotec) and then the mRNA was processed ac-cording to the manufacturer's instructions. Translations wereperformed by programing micrococcal nuclease-treated rab-bit reticulocyte extracts (Promega Biotec) with the in vitrosynthesized mRNA in the presence of 25 uCi of [35S]methi-onine (1 Ci = 37 GBq; Amersham) according to the manu-facturer's instructions. The material so synthesized migratesas a doublet on SDS/PAGE with an apparent molecular massof 34 and 31 kDa. Both bands can be immunoprecipitatedwith antisera directed to the carboxyl terminus of IL-13 (4).
Preparation of Human Pre-Llfl Processing Site Mutant.Construction of a mutant pre-IL-1p precursor was performedby site-directed oligonucleotide mutagenesis. A syntheticdouble-stranded 27-nucleotide (27-mer) oligodeoxyribonu-cleotide (corresponding to amino acids 115-126) with ApaII/Hpa II ends was synthesized on an Applied BiosystemsDNA 380A synthesizer according to established protocols.The 27-mer encodes an Asp116 -* Ala'16 amino acid substi-tution at the -1 position adjacent to the proposed processingsite of pre-IL-1,B. The oligonucleotide was ligated to EcoRI/Apa II and Hpa II/Pst I fragments obtained from cleavage ofa full-length IL-1/3 cDNA. The ligated fragments were addedto a ligation reaction mixture containing EcoRI/Pst I-cleavedpGEM-3. Clones containing the pGEM/IL-1P mutant wereidentified by hybridization with the mutant oligonucleotidesequence. Clones were mapped by restriction endonucleasecleavage and the DNA was sequenced to verify authenticityof the mutation. Transcription of the vector bearing themutant or native constructs produced a 1.5-kb mRNA andtranslation resulted in a doublet of 34- and 31-kDa proteins.
Preparation of Cell Extracts. Cell-free extracts were pre-pared from THP.1 cells or human peripheral blood mono-cytes by nitrogen cavitation or by hypotonic lysis. Briefly,cells were harvested by centrifugation at 1000 x g and thenwashed three times with Dulbecco's phosphate-buffered sa-line without MgCl2 and CaCl2 (GIBCO). Cell pellets werewashed once with 10 vol of hypotonic buffer (10 mM KCI/20mM Hepes, pH 7.4/1.5 mM MgCl2/0.1 mM EDTA) and werepelleted at 1000 x g for 10 min. The resulting cell pellets werethen resuspended in 3 vol of hypotonic buffer and placed onice for 20 min. Lysis of the cell suspension was accomplishedwith 20 strokes in a Dounce homogenizer. For gentler lysis,the cell pellets were resuspended in hypotonic buffer andthen placed in a stainless steel nitrogen pressure cell. The cellwas pressurized to 400 psi (1 psi = 6.9 kPa) with nitrogen andheld at this pressure for 30 min at 4°C with agitation. The cellswere lysed by simultaneously releasing the pressure andevacuating the cells from the container. Cellular homoge-nates were clarified by centrifugation at 1000 x g for 20 min,and the supernatant (S-1) was saved and stored at -80°C. Insome cases, further fractionation of the cell extracts wasperformed by high-speed centrifugation. Freshly preparedS-1 supernatant was further centrifuged at 30,000 x g (15,000
rpm in a Sorvall SS-34 rotor) for 10 min followed by centrif-ugation of the S-30 at 50,000 rpm (300,000 x g) for 1.5 hr at40C in a Beckman type 50.2 rotor. The pellets from each stepwere resuspended in the starting volume of hypotonic bufferand then frozen. The final supernatant (S-300) was retainedand stored at -800C.
Percoll Gradient Fractionation of THP.1 Extracts. THP.1S-1.0 extracts were prepared by nitrogen cavitation as de-scribed above. The lysate was layered over preformed 0-100lo Percoll gradients (Pharmacia) made with 0.25 M su-crose/10 mM Hepes, pH 7.4/10 mM KCl/1.5 mM MgCl2/0.1mM EDTA. The gradients were then spun at 48,000 X g for25 min in a Sorvall SS-34 rotor. Gradients were collectedfrom the top and 1.0-ml fractions were collected and assayedfor processing activity. Enzymatic activities associated withthe cytosol (lactate dehydrogenase; ref. 22), lysosomes (N-acetyl f-D-glucosaminidase; ref. 23), plasma membrane (5'-nucleotidase; ref. 24), and microsomes (sulfatase C; ref. 25)were assayed to identify subcellular compartments and theirlocation in the gradient.
In Vitro Pre-EL-113 Cleavage Assay. Cleavage of pre-IL-1,3was performed by incubating 1 Al of rabbit reticulocyteextract containing radiolabeled precursor with 10-20 Al ofcrude cell-free extracts, which had been brought to 0.5%Nonidet P-40, at 30°C-37°C for 1 hr. Detergent is not requiredfor THP.1 cytosolic cleavage activity but was used to solu-bilize particulate material and prevent settling during theincubation. The reaction conditions were determined to beoptimal at apH between 7.0 and 8.0 and at salt concentrations<100 mM KCI (data not shown). The extent of cleavage wasassessed by SDS/PAGE according to Laemmli (26) followedby fluorography.
Sequencing of Cleavage Product. For sequencing of cleav-age product, pre-IL-1f3 was synthesized as described abovewith the exception that the material was labeled with L-[3H]leucine (New England Nuclear). Cleavage reactionswere performed using THP.1 S-300 prepared as describedabove. Mature radiolabeled IL-1,B cleavage product waspurified from the processing reaction by SDS/PAGE fol-lowed by transfer to polyvinylidine difluoride membrane.N-terminal sequencing of the radiolabeled cleavage productwas by automated Edman degradation on an Applied Bio-systems model 470A gas-phase sequencer (27).
RESULTSPre-IL-1f Cleavage Activity Is Found in Monocyte Cell
Extracts. S-1 cell-free extracts were prepared from a humanmonocyte-like cell line (THP.1 cells, which are known tosecrete IL-1f3 having the same amino terminus as monocyte-derived material; ref. 28), human embryonic lung fibroblasts(MRC-5), and Chinese hamster ovary cells (CHO) and testedfor their ability to cleave in vitro synthesized and radiolabeledpre-IL-1l3 in the presence of 0.5% Nonidet P-40 (Fig. 1).Visualization of the reaction products by SDS/PAGE andfluorography clearly shows that pre-IL-1,8 is cleaved inextracts from THP.1 cells (Fig. 1). THP.1 cell extractsproduce a 17.5-kDa fragment that migrates at the samemolecular mass as recombinant mature 1251-labeled IL-113(Fig. 1). A slightly larger fragment, but one oflow abundance,is also produced that migrates at 19.0 kDa. The activity thatwill produce a fragment this size varies in amount betweenS-1 extracts but is dissimilar to the one that produces the17.5-kDa form (see below). In contrast, no proteolysis of anysort was observed in reaction mixtures containing MRC-5 orCHO cell extract. Mixing of the MRC-5 and THP.1 extractsin a 1:1 ratio did not prevent detection of convertase activity(data not shown).
Similarly, we searched for pre-IL-1i8 cleavage activities inhuman blood cells separated by elutriation into fractions
5228 Biochemistry: Kostura et al.
Dow
nloa
ded
by g
uest
on
Sep
tem
ber
7, 2
020

Proc. Natl. Acad. Sci. USA 86 (1989) 5229
N~Q0, -00l
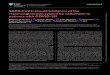
FIG. 1. THP.1 cell-free extracts process pre-IL-1.8 to a 17.5-kDa
molecule. Extracts were prepared from THP.1 cells, MRC.5 fibro-
blast cells, and Chinese hamster ovary cells. Twenty microliters of
extract was used for processing 1 j.d of pre-IL-lp synthesized in vitro
in rabbit reticulocyte lysates. Electrophoresis of radioiodinated
recombinant IL-113 is shown on the right as a molecular weightstandard.
enriched for platelets, lymphocytes, or monocytes (Fig. 2).Extracts were prepared from each fraction and tested for
their ability to cleave IL-113 precursor. Each cleavage reac-
tion was run in the presence or absence of a specific poly-morphonuclear leukocyte (PMN) elastase inhibitor, one of a
variety of substituted cephalosporins known to inhibit human
leukocyte elastase 1-[3-[(acetyloxy)methyl]-7-methoxy-8-oxo-5-thia-1-azabicyclo[4.2.0]oct-2-en-2-yl~carbonyI morpholine,S,S-dioxide, (6R-cis) (Merck compound number L-659,166;ref. 29) to eliminate proteolysis of the substrate by PMN
elastase during incubation. A PMN elastase-like activity, as
shown by cleavage of the elastase substrate methoxysuccinyl-Ala-Ala-Pro-Val-paranitroanflide, is found in monocytes and
PMN elastase is known to cleave pre-IL-1p generating a
19-kDa as well as a smaller 16.5- to 17.5-kDa product (ref. 17;M.J.K., unpublished observation). Our results clearly indicate
that pre-IL-10 was not cleaved in extracts derived from
platelets or lymphocytes but was cleaved in extracts derived
from cell populations substantially enriched or homogenousfor human blood monocytes (Fig. 2, lanes 4 and 5). The effects
of PMN elastase inhibition by L-659,116 are clearly demon-
strated in the starting fraction and in fraction 5. Without the
inhibitor, substantial degradation of the 17.5-kDa product and
precursor occurs, whereas the material is protected in its
presence. However, the activity that generates the 17.5-kDa
form is not affected by the inhibitor.
Subcellular Fractionation of Precursor Cleavage Activity.Cytological evidence including immunolocalization by lightand electron microscopy have indicated that pre-IL-1f3 is
found predominantly in the cytoplasmic ground substance (3,
15). Little to no mature IL-1f3 has been detected in freshlylysed monocytes by immunoblot analysis (3, 4). These data
suggest that IL-1 is synthesized as a precursor that is cleaved
to yield mature product and is secreted very shortly there-
after. Thus, the identification of the cellular compartment in
which potential cleavage activities may reside will be impor-tant in understanding the entire secretion pathway. Fraction-
ation of cellular organelles was performed by Percoll gradientfractionation of nitrogen cavitated THP.1 cells. The fractions
1 2 3 4 6 8 10 12 14
1251Start 1 2 3 4 5 IL-13-+ -+ -+ -+ -+ -+ L659
FIG. 2. IL-1P convertase activity is associated with monocytes.Leukophoresed cells from a single healthy donor were separated firstby separation on lymphocyte separation medium (LSM) followed bycounterflow centrifugal elutriation. The starting population and fivesequentially eluted fractions were collected in equal volumes,scanned by a Coulter particle counter and examined by fluorescence-activated cell sorting analysis. The starting fraction is the pool ofcellsresulting from LSM separation. Fraction 1 contains platelets with aminor contaminant of lymphocytes. Fraction 2 contains T and Blymphocytes but no monocytes. Fractions 3, 4, and 5 contain amixture ofT and B lymphocytes and monocytes. The three fractionscontain approximately 10o, 50%1, and 95% monocytes, respectively.Cell-free extracts were made from each of these fractions and thenassessed for levels of convertase activity. All S-1.0 extracts wereprepared by Dounce homogenization. Where indicated, the Merckelastase inhibitor L-659,166 (100 uM) was added to prevent degra-dation by monocyte-associated elastase. "2I-labeled recombinanthuman IL-1,B is shown at right.
were assayed for a variety of enzymatic markers as well aspre-IL-1,8 cleavage activities. Membranous compartmentssuch as lysosomes and microsomes remained essentiallyintact, based on the distribution of enzymatic markers asso-ciated with them. Within the gradients, pre-IL-113 cleavageactivities were detected in two regions ofdensity (Fig. 3). Thefirst is a rapidly sedimenting component that generates 19-and 16.5-kDa cleavage products (Fig. 3, fractions 14-22).This activity is most likely granule associated, since itsediments at a density slightly greater than that of thelysosomal marker N-acetyl f-D-glucosaminidase. The activ-ity also proved to be latent, or encapsulated in a membranouscompartment, since no cleavage was observed in the absenceof detergent. In addition, the enzyme was inhibited by serineprotease inhibitors including the specific PMN elastase in-hibitor L-659,166 (unpublished data). The second activityobserved, capable of generating the 17.5-kDa product, ismost likely cytosolic in origin, since it cosediments with thecytosolic marker LDH (fractions 1-4). Demonstration of thisactivity did not require detergent, showing that the enzymeis not encapsulated in a vesicle. These data suggest that atleast two distinct enzymes capable of cleaving pre-IL-1, arefound in THP.1 cells and human blood monocytes (data notshown). The activities are separable based on velocity sed-imentation and sensitivity to protease inhibitors, and theyyield unique and distinct cleavage products. Only the cyto-solic activity appears capable of generating a product similarin size to mature IL-1,B. Therefore, in the experimentspresented below, S-300 cell-free extracts were used to elim-inate the formation of products generated by PMN elastase.
16 on 18 20 22 24 26 28 30 32 39
19.0 kD17.5 -16.5 Aftk*
FIG. 3. Percoll gradient fractionation of THP.1 cell-free extract. THP.1 S-1.0 cell-free extract was prepared and layered onto a 0-100%6gradient of Percoll (Pharmacia) and then centrifuged for 20 min at 48,000 x g. The gradient was fractionated and aliquots of each fraction wereassayed for the indicated enzymatic activities (see Materials and Methods) as well as pre-IL-1,8 convertase activities. Note the differentialpartitioning of a cytosolic activity (fractions 1-4) capable of generating a 17.5-kDa product from a particulate activity (fractions 14-22), whichgenerates 19- and 16.5-kDa forms. The standard lane (STD.) represents radioiodinated recombinant mature IL-1,8.
Biochemistry: Kostura et al.
9qI
931fI'IV
lmwq.IV lw PrW:- .4"Wqp MP'
do
Dow
nloa
ded
by g
uest
on
Sep
tem
ber
7, 2
020

Proc. Natl. Acad. Sci. USA 86 (1989)
020.
A
Amino Acid
FIG. 4. Sequence analysis of 17.5-kDa cleavage product.[3H]Leucine-labeled pre-IL-1P was cleaved with THP.1 S-300 ex-tract. The 17.5-kDa cleavage product was isolated and sequenced.Fractions from the sequence run were counted for associated radio-activity. Peaks of radioactivity were found in cycles 6 and 10corresponding to the known amino acid sequence of mature IL-li,which is plotted as the ordinate axis. Amino acids are designated bythe single-letter code.
Protein Sequence Analysis of 17.5-kDa Cleavage Product.To identify the amino terminus of the 17.5-kDa cleavageproduct [3H]leucine-labeled pre-IL-1(8 was cleaved by THP.1S-300 extract, electrophoresed, and transferred to polyvinyl-idine difluoride membrane, and subjected to automated Ed-man degradation sequencing (27). Peaks of radiolabeledleucine were detected at cycles 6 and 10 (Fig. 4). This resultis consistent with the cleavage product containing an aminoterminus congruent with the known sequence of matureIL-1p (Fig. 4) and clearly indicates that the extract is able toprocess synthetic pre-IL-1j3 accurately. Nevertheless, thereappeared to be a greater than expected amount of carryoverbetween fractions eluted during the sequencing. Correctingfor lag, nearly 70%o of the sequenced product contains thealanine amino terminus with the remainder having asparticacid in the + 1 position. Whether this microheterogeneity is
A IL-1 Beta sequences Contain a Conserved Aspartic Acid at the -1Position in the Cleavage Site.
BpL precursor
.1 .25 .5 .75 1.0 -THP.1
.. . ., .V
aud. AWL'm 4wI
pL precursor
.1 .25 .5 .75 1.0 -THP.1
?-3W.w d
FIG. 6. Cleavage of wild-type or mutant pre-IL-1.8 by THP.1S-300. Increasing volumes of either wild type (A) or mutant (B)precursor were added to the same volume (10 p.1) of THP.1 S-300extract, incubated, and analyzed by SDS/PAGE. Lanes labeled-THP.1 indicate incubation of 1 A.l of precursor without THP.1cell-free extract.
representative of the action of a single or multiple enzymesis at present unclear.
Production of Mutant Pre-EL-ifi and Characterization ofCleavage Products in Vitro. Sequence analysis of pre-IL-1(3cDNA from four species (6-9) reveals that the presumptivecleavage site contains an aspartic acid residue in the -1position (Fig. 5A). Because of the striking nature of thisconservation as well as the fact that there are no character-ized eukaryotic proteases that will cleave at an Asp-Alalinkage, we decided to mutate this amino acid to determinethe effects, if any, on cleavage activity (Fig. SB). Oligonu-cleotide site-directed mutagenesis was performed as de-scribed to convert Asp6 --) Ala116. Site-directed mutagen-esis of aspartyl-116 did not affect in vitro transcription ortranslation of the pre-IL-13 mRNA (data not shown). Whenadded to THP.1 S-300 extract, little or no cleavage of themutant precursor was observed (Fig. 6). Titration of thesubstrate revealed that some cleavage would occur at ele-vated concentrations of substrate. However, cleavage of themutant did not produce the appropriately sized fragments. Itis quite apparent then that the aspartyl residue plays animportant role in maintaining pre-IL-1,8 in a cleavage com-petent form either directly through interactions with theconvertase or indirectly through maintenance of secondarystructure.
HumanMouseBovineRabbit
ALA TYR VAL HIS ASP ALA PRO VAL ARG SERLEU LEU VAL CYS ASP VAL PRO ILE ARG GLUGLU PHE LEU CYS ASP ALA PRO VAL GLN SERSER LEU GLU CYS ASP ALA --- VAL ARG SER LEU
B Apa Ll Hpal
TG CAC GCT GCA CCT GTA CGA TOG CTG AAC TGC ACG CTC
G CGA CGT GGA CAT GCT AGC GAC TTG ACG TGC GAG GCHIS ALA ALA PRO VAL ARG SER LEU ASN CYS THR LEU
EcoRI Pst II Is\\ \
Precursor Domain
I"\ \ \\M \ \ \a-D-
Mature Domain
FIG. 5. Construction of mutant pre-IL-1, by oligonucleotidesite-directed mutagenesis. (A) Comparison of amino acid sequencesadjacent to the presumptive cleavage site in pre-IL-1,8 from fourdifferent species. (B) Nucleotide sequence of complimentary oligo-nucleotides used for construction of Asp"16 -* Ala"6 mutant by
site-directed mutagenesis.
DISCUSSIONWe have identified a protease activity present in human bloodmonocytes or THP.1 cells but not found in a variety oflymphoid, fibroblast, or epithelial cell lines, which is capableof specifically cleaving synthetic pre-IL-1(3. The fact that thisactivity is only found in human blood monocytes or mono-cyte-like cell lines that are capable of secreting IL-1 is animportant indication of a monocyte-specific processing andsecretion pathway. The cleavage product of the wild-typeprecursor is identical in size and has the same amino-terminalsequence as mature IL-1,3 secreted by monocytes. In addi-tion, we have also shown that the cleavage product specifi-cally binds IL-1 receptors found on MRC-5 fibroblast mem-branes (data not shown).The soluble processing activity we have described is ca-
pable of cleaving in vitro synthesized pre-IL-1, in a site-specific manner between amino acid residues Asp116 andAla'17. We have confirmed the importance of this site byoligonucleotide site-directed mutagenesis of the highly con-served aspartic acid residue at position 116. Conversion ofthis residue to an alanine results in a substrate that is nowunrecognized by convertase. A residual amount of cleavageto a 29-kDa product occurs but this is also observed in thewild-type form (Fig. 6). Whether this represents a processingintermediate or selection of an alternative cleavage site isunclear. The data from this single mutant are quite clear inthat the aspartic acid residue is involved in substrate selec-
5230 Biochemistry: Kostura et al.
---
Dow
nloa
ded
by g
uest
on
Sep
tem
ber
7, 2
020

Proc. Natl. Acad. Sci. USA 86 (1989) 5231
tion by what appears to be a highly specific processingactivity.The protease activity we have described is qualitatively
different from one described by Black et al. (17) in KG-1cells, a neutrophil-like cell line. In this case, the processingactivity was reported to be located in a 30,000 x g sediment-able compartment and was extractable and active in 1 MNaCl. The activity generated a 19-kDa and what appears tobe a 17.5-kDa fragment when partially purified pre-IL-1(3 was
used as a substrate. Amino-terminal sequence data were notprovided for the 17.5-kDa product, although it was able toreact with a monoclonal antibody directed toward the car-
boxyl terminus of mature IL-1f (17). The cleavage patternreported by Black et al. is reminiscent of an activity we havedescribed from THP.1 cells, which generates products of 19.0and 16.5 kDa (Fig. 3). This activity sediments as a large densegranule in Percoll gradients (Fig. 3) and is also pelletable bysedimentation at 30,000 x g (data not shown). Our datasuggest that the cytosolic and sedimentable protease activi-ties found in THP.1 cells are distinct and not due to differ-ential partitioning of the same enzyme during cell lysis.Differences in the sizes of the cleavage products formed,latency, salt tolerance, and sensitivity to elastase inhibitorsdistinguish the two activities.
In the report of Hazuda et al. (5), human monocytes were
lysed in the presence of exogenous E. coli-derived humanpre-IL-18 and fractionated by differential centrifugation.Their results showed that a 17.5-kDa cleavage product was
found in the cytosol. While the production of the 17.5-kDaprotein by human monocytes was confirmed, we believethese data do not completely address the cellular compart-ment in which the convertase resides. Rather the data simplyreflect the fractionation of the cleavage products after lysis.No attempt was made to further characterize the convertaseactivity or the 17.5-kDa product.Of importance in the identification of any putative pro-
cessing activity is the requirement that the substrate and theprotease exist in the same cellular compartment for at leastsome portion of the secretory process. In the case of IL-13,the evidence suggests that the precursor is a cytosolic pro-
tein. Evidence based on immunoblotting and immunolocal-ization suggests that only pre-IL-1j3, and not mature IL-1f3,is associated with actively secreting monocytes and that thismaterial is located in the cytosol (3, 4, 15). The convertaseactivity also partitions into the cytosol. However, we cannotrule out entirely the possibility that the activity is releasedfrom a loose association with the plasma membrane or fromthe luminal compartment of the endoplasmic reticulum.The available evidence suggests that mature IL-1,8 is
secreted posttranslationally in monocytes and that process-
ing closely precedes or is coupled to secretion. In contrast,fibroblasts transfected with pre-IL-1f3 cDNA appear incapa-ble of secreting mature IL-1/3 even though a substantialquantity of precursor can be synthesized and localized in thecytosol (14). Since fibroblasts appear not to contain a specificpre-IL-1l3 cleavage enzyme, it is possible that the convertasewe have described above is required for secretion of matureIL-1,f.
The authors would like to thank Dr. Paul Finke for the sample ofelastase inhibitor and Dr. Mervyn Turner for his critical reading ofthis manuscript. Special thanks to Dr. Phil Davies for criticallyreading this manuscript as well as for the enthusiasm and support thathe has shown this project.
1. Gin, J. G., Lomedico, P. T. & Mizel, S. B. (1985) J. Immunol.134, 43-349.
2. Auron, P. E., Warner, S. J. C., Webb, A. C., Cannon, J. G.,Bernheim, H. A., McAdam, K. J. P. W., Rosenwasser, L. J.,LoPreste, G., Mucci, S. F. & Dinarello, C. A. (1987) J. Im-munol. 138, 1447-1456.
3. Bayne, E. K., Rupp, E. A., Limjuco, G., Chin, J. & Schmidt,J. A. (1986) J. Exp. Med. 163, 1267-1280.
4. Limjuco, G., Galuska, S., Chin, J., Cameron, P., Boger, J. &Schmidt, J. A. (1986) Proc. Natl. Acad. Sci. USA 83, 3972-3976.
5. Hazuda, D. J., Lee, J. C. & Young, P. R. (1988)J. Biol. Chem.263, 8473-8479.
6. March, C. J., Mosley, B., Larsen, A., Cerretti, D. P., Braedt,G., Price, V., Gillis, S., Henney, C. S., Kronheim, S. R.,Grabstein, K., Conlon, P. J., Hopp, T. P. & Cosman, D. (1985)Nature (London) 315, 641-647.
7. Gray, P. W., Glaister, D., Chen, E., Goeddel, D. V. & Pen-nica, D. (1986) J. Immunol. 137, 3644-3648.
8. Maliszewski, C. R., Baker, P. E., Schoenborn, M., Davis,B. S., Cosman, D., Gillis, S. & Cerretti, D. P. (1988) Mol.Immunol. 25, 429-437.
9. Mori, S., Goto, F., Kumiko, G., Ohkawara, S., Maeda, S.,Shimada, K. & Yoshinaga, M. (1988) Biochem. Biophys. Res.Commun. 150, 1237-1243.
10. Auron, P. E., Webb, A. C., Rosenwasser, L. J., Mucci, S. F.,Rich, A., Wolff, S. M. & Dinarello, C. A. (1984) Proc. Natl.Acad. Sci. USA 81, 7907-7911.
11. Mosley, B., Urdal, D. L., Prickett, K. S., Larsen, A., Cosman,D., Conlon, P. J., Gillis, S. & Dower, S. K. (1987) J. Biol.Chem. 262, 2941-2944.
12. Lomedico, P. T., Gubler, U., Hellmann, C. P., Dukovich, M.,Giri, J. G., Pan, Y.-C. E., Collier, K., Semionow, R., Chua,A. 0. & Mizel, S. B. (1984) Nature (London) 312, 458-462.
13. Cameron, P., Limjuco, G., Rodkey, J., Bennett, C. & Schmidt,J. A. (1985) J. Exp. Med. 162, 790-801.
14. Young, P. R., Hazuda, D. H. & Simon, P. (1988) J. Cell Biol.107, 447-456.
15. Singer, I., Scott, S., Hall, G., Limjuco, G., Chin, J. & Schmidt,J. A. (1988) J. Exp. Med. 167, 389-407.
16. Verner, K. & Schatz, G. (1988) Science 241, 1307-1313.17. Black, R., Kronheim, S. R., Cantrell, M., Deeley, M. C.,
March, C. J., Prickett, K. S., Wignall, J., Conlon, P. J., Cos-man, D., Hopp, T. & Mochizuki, D. Y. (1988) J. Biol. Chem.263, 9437-9442.
18. Wicker, L., Boltz, R., Nichols, E. N., Miller, B., Sigal, N. &Peterson, L. B. (1987) Cell. Immunol. 106, 318-329.
19. Tsuchiya, S., Yamabe, M., Yamaguchi, Y., Kobayashi, Y.,Konno, T. & Tada, K. (1980) Int. J. Cancer 26, 171-176.
20. Krakauer, T. & Oppenheim, J. J. (1983) Cell. Immunol. 80,223-229.
21. Maniatis, T., Fritsch, E. F. & Sambrook, J. (1982) MolecularCloning:A Laboratory Manual (Cold Spring Harbor Lab., ColdSpring Harbor, NY).
22. Morgorstern, S., Flor, R., Kessler, G. & Klein, B. (1965) Anal.Biochem. 13, 149-161.
23. Wollen, J. W., Heyworth, R. & Walker, P. G. (1961) Biochem.J. 78, 111-121.
24. Rome, L. H., Garvin, A. J., Allietta, M. M. & Neufeld, E. F.(1979) Cell 17, 143-153.
25. Canonico, P. G., Beaufay, H. & Nyssens-Jadin, M. (1978) J.Reticuloendothel. Soc. 24, 115-135.
26. Laemmli, U. K. (1970) Nature (London) 227, 680-685.27. Matsudaira, P. (1987) J. Biol. Chem. 261, 10035-10038.28. Matsushima, K., Copeland, T. D., Onozaki, K. & Oppenheim,
J. J. (1986) Biochemistry 25, 3424-3429.29. Doherty, J. B., Ashe, B. M., Argenbright, L. W., Barker,
P. L., Bonney, R. J., Chandler, G. O., Dahlgren, M. E., Dorm,C. P., Finke, P. E., Firestone, R. A., Fletcher, D., Hagmann,W. K., Mumford, R., O'Grady, L., Maycock, A. L., Pisano,J. M., Shah, S. K., Thompson, K. R. & Zimmerman, M. (1986)Nature (London) 322, 192-194.
Biochemistry: Kostura et al.
Dow
nloa
ded
by g
uest
on
Sep
tem
ber
7, 2
020