Embed Size (px)
Citation preview

NeuroImage 62 (2012) 1520–1528
Contents lists available at SciVerse ScienceDirect
NeuroImage
j ourna l homepage: www.e lsev ie r .com/ locate /yn img
Common substrate for mental arithmetic and finger representation in theparietal cortex
Michael Andres a,b,c,⁎, Nicolas Michaux a,b, Mauro Pesenti a,b
a Institut de Recherche en Sciences Psychologiques, Université catholique de Louvain, Place Cardinal Mercier 10, 1348 Louvain-la-Neuve, Belgiumb Institute of Neuroscience, Université catholique de Louvain, Avenue Hippocrate 10, 1200 Brussels, Belgiumc Department of Experimental Psychology, Ghent University, Henri Dunantlaan 2, 9000 Gent, Belgium
⁎ Corresponding author at: Department of ExperimentHenri Dunantlaan 2, 9000 Gent, Belgium.
E-mail address: [email protected] (M. Andres)
1053-8119/$ – see front matter © 2012 Elsevier Inc. Alldoi:10.1016/j.neuroimage.2012.05.047
a b s t r a c t
a r t i c l e i n f oArticle history:Accepted 20 May 2012Available online 24 May 2012
Keywords:CountingMemory retrievalCalculationEmbodied cognition
The history of mathematics provides several examples of the use of fingers to count or calculate. These obser-vations converge with developmental data showing that fingers play a critical role in the acquisition of arith-metic knowledge. Further studies evidenced specific interference of finger movements with arithmeticproblem solving in adults, raising the question of whether or not finger and number manipulations rely oncommon brain areas. In the present study, functional magnetic resonance imaging (fMRI) was used to inves-tigate the possible overlap between the brain areas involved in mental arithmetic and those involved in fin-ger discrimination. Solving subtraction and multiplication problems was found to increase cerebral activationbilaterally in the horizontal part of the intraparietal sulcus (hIPS) and in the posterior part of the superiorparietal lobule (PSPL). Finger discrimination was associated with increased activity in a bilateral occipito-parieto-precentral network extending from the extrastriate body area to the primary somatosensory andmotor cortices. A conjunction analysis showed common areas for mental arithmetic and finger representa-tion in the hIPS and PSPL bilaterally. Voxelwise correlations further showed that finger discrimination andmental arithmetic induced a similar pattern of activity within the parietal areas only. Pattern similaritywas more important for the left than for the right hIPS and for subtraction than for multiplication. These find-ings provide the first evidence that the brain circuits involved in finger representation also underlie arithmeticoperations in adults.
© 2012 Elsevier Inc. All rights reserved.
Introduction
The use of fingers to count objects or calculate goes back to theAncient Times (e.g., Cicero, Epistole ad Atticum, V, 21, 13, 106–43 BCN)and is further illustrated in famous counting systems (e.g., BedaVenerabilis,De temporum ratione, 672–735ACN) commonly used acrossEurope in the Middle Ages (Butterworth, 1999; Williams andWilliams,1995). The success of these counting systems across times and culturesis likely to result from their capacity to pass the bottleneck of pre-existing representations in the brain of each individual (De Cruz,2006). In this view, fingers constitute a useful means for acquiringand communicating arithmetic knowledge because they provide a phys-ical counterpart formental operations and they rely on pre-existing rep-resentations in the sensorimotor system. Several developmental studieshave confirmed the intimate relationship between numbers andfingers.Children's score in finger discrimination tests is the best predictor oftheir later arithmetical skills (Noël, 2005) and finger counting provedto be very important while learning to add or subtract numbers (Costa
al Psychology, Ghent University,
.
rights reserved.
et al., 2011; Domahs et al., 2008). In adults, behavioural studies reportedfaster numerical judgements when target numbers were primed byfinger configurations congruent with the finger-counting habits of theparticipants (Badets et al., 2010; Di Luca & Pesenti, 2008; Di Luca et al.,2006, 2010). A trace of finger-counting habits was also found in electro-physiological studies showing increased corticospinal excitability (CSE)in hand muscles during various numerical tasks (Andres et al., 2007;Sato et al., 2007).
There is now a large agreement that the parietal cortex plays animportant role in mental calculation, as shown by numerous neuro-psychological (e.g., Takayama et al., 1994) and brain imaging studies(for a recent meta-analysis, see Arsalidou and Taylor, 2011). Severalstudies showed that arithmetic factors, such as problem size (DeSmedt et al., 2011; Stanescu-Cosson et al., 2000), number of operands(Menon et al., 2000) and strategy (Delazer et al., 2003, 2005; Grabneret al., 2009), influence the blood-oxygen level-dependent (BOLD) sig-nal in the horizontal part of the intraparietal sulcus (hIPS) and in theposterior part of superior parietal lobule (PSPL), a region extendingfrom the posterior segment of the IPS to the precuneus. In contrastto subtraction and addition that may require calculation procedures,the multiplication of single digits generally leads to direct retrievalof the problem and its answer from long-term memory (Campbell,

1521M. Andres et al. / NeuroImage 62 (2012) 1520–1528
1987; Galfano et al., 2003; LeFevre et al., 1996), with the contributionof the middle and superior temporal gyrii (MTG and STG; Lampl et al.,1994; Prado et al., 2011; Sandrini et al., 2003; Zhou et al., 2007) and/or the angular gyrus (ANG; Delazer et al., 2003; Grabner et al., 2009,2011). Recent evidence from fMRI-guided TMS studies showed thatthe integrity of the hIPS is required to perform both subtraction andmultiplication problems, challenging the view that the brain net-works underlying these operations are entirely separated (Andreset al., 2011; Salillas et al., 2012). Finally, mental arithmetic has beenshown to recruit several areas in the frontal lobe (Arsalidou andTaylor, 2011), among which areas in the ventral and dorsal parts ofthe premotor cortex (Pesenti et al., 2000; Piazza et al., 2006). Interest-ingly, the parietal and frontal areas activated during mental arithmeticare very close to those activated by executed or imagined finger move-ments, suggesting that number processing is anchored in the processesunderlying finger representation (Haaland et al., 2004; Harringtonet al., 2000; Pelgrims et al., 2009, 2011). However, so far, this apparentanatomical similarity only stems from indirect comparisons of resultscoming from different studies using different methods.
Moreover, the functional mechanism that accounts for the poten-tial overlap between mental arithmetic and finger representation inthe human brain remains to be specified. First, it is unclear whetherthe processes and/or representations related to finger discriminationform an intrinsic component of number processing, or whether theycomplement the core number system by providing helpful tools tomanipulate numbers, such as other slave systems involved in visuo-spatial or linguistic aspects of number processing (Dehaene et al.,2003). Second, it is essential to determine how much the potentialoverlap between mental arithmetic and finger representation is relat-ed to the somatic representation of the body in order to specifythe embodied nature of their interactions. In line with this, previousfMRI studies showed that activations in the primary motor (M1)and somatosensory cortex (S1) were sensitive to the hand used toperform finger movements, whereas activations of the hIPS andPSPL occurred irrespective of hand laterality (Haaland et al., 2004;Harrington et al., 2000). A third question concerns the influence ofrepresentational and procedural aspects of finger counting on the im-plementation of arithmetic operations in finger circuits. Representa-tional aspects were evidenced in behavioural studies showing thatnumbers or results of arithmetic operations implicitly activate a men-tal configuration of fingers (Badets et al., 2010; Di Luca & Pesenti,2008; Di Luca et al., 2006, 2010). Procedural aspects refer to the roleof fingers in instantiating the one-to-one correspondence betweencounted items and any ordered series (Andres et al., 2007). Whereasrepresentational aspects do not necessarily imply a different imple-mentation of each operation in finger circuits, procedural aspectspredict that the involvement of finger circuits will differ across op-erations as a function of the level of practice with finger counting.In children, single-digit multiplication problems are mostly solved bymemory retrieval from the fourth grade (Cooney et al., 1988), whereassubtraction often involves computational strategies with a great em-phasis on finger-based calculation procedures in the early stages of ac-quisition (Fuson, 1988; pp. 278–279). Adults report almost exclusivereliance on retrieval for multiplication (i.e., 98% and 95% for smalland large problems respectively), whereas this strategy is much lessused for subtraction (i.e., 73% and 42% for small and large problems re-spectively; Campbell and Xue, 2001). The status of addition is less clearwith a rate of memory retrieval in-betweenmultiplication and subtrac-tion (i.e., 88% and 64% for small and large addition problems respec-tively; Campbell and Xue, 2001). A dual-task experiment showed thatconcurrent finger movements slow down addition and subtraction,whereas multiplication remains unaffected, even after matching prob-lems for response speed and accuracy (Michaux et al., submitted forpublication). This difference was attributed to the fact that multiplica-tion is less sensitive to finger interference because answers can be re-trieved from long-term memory without computation. It is unclear,
however, whether the predicted overlap between the brain circuitsunderlying finger discrimination and those involved in mental arith-metic will differ between operations. If the influence of finger use onarithmetic operations is strictly related to procedural aspects of cou-nting in childhood, then finger circuits should overlap exclusivelywith those involved in subtraction. The finding of a similar overlapfor subtraction and multiplication would mean that the influence offinger use on arithmetic operations is not limited to the proceduralaspects of counting but entails a representation of the numbersand/or intermediary results on the fingers regardless of arithmeticoperation.
In order to address these issues, we used fMRI to measure theBOLD signal in the brain of healthy adults who had to solve subtrac-tion and multiplication problems or to discriminate the position ofthe fingers on the left and right hand. The similarity between the pat-tern of activity observed during finger discrimination and that ob-served during each arithmetic operation was further explored bycomputing voxelwise correlations between tasks.
Material and methods
Participants
Eighteen French-speaking males (mean±S.D.: 21.3±2.5 years)gave their informed consent to participate to this study. All partici-pants were right-handed according to the Edinburgh HandednessInventory (mean EHI score ([right− left]/[right+left])±S.D.: 0.81±0.16; Oldfield, 1971). They had no history of neurological or psychiatricdisorders, had normal or corrected-to-normal vision, and they were un-aware of the purpose of the study. The experiment was non-invasiveand was performed in accordance with the ethical standards laid downin the 1964 Helsinki Declaration. The experimental protocol was ap-proved by the Biomedical Ethical Committee of the Université catholiquede Louvain.
Tasks and stimuli
The three experimental tasks were matched with specific controltasks in terms of visual display and response requirements (seeFig. 1). In the finger discrimination task (adapted from KinsbourneandWarrington, 1962), the participants held a wooden block of irreg-ular shape in each hand, with half of the fingers flexed in the holesand the other half extended over the bumps of the blocks; thethumb was positioned on the lateral face of the block to allow a stablegrip (Fig. 1A). In each trial, the palm view of a left or right hand wasdisplayed on the screen in black on a white background. During theexperimental blocks, one finger was red and the participants wereinstructed to answer aloud “yes” if their corresponding finger wasflexed into a hole of the wooden block and “no” if it was extendedover a bump, without moving or looking at their fingers (Fig. 1B). Allfingers but the thumb were tested in each experimental block, usinga pseudo-random order, so that the same finger was not coloured inred in two consecutive trials. In the control task, all fingers on the draw-ing had the same colour, either black or red, and the participants had todecide whether it was red by answering aloud “yes” or “no” (Fig. 1C).Different wooden blocks were placed in the left and right hands atthe beginning of each run in order to prevent the participants from re-lying on learned associations between finger names and expected an-swers. The left and right hands were tested in separate series of trialsto avoid confusion between finger discrimination and left–right orien-tation during the task. During the arithmetic tasks, one Arabic digitranging from 3 to 9 was displayed on the screen and the participantshad to subtract it from 11 or 13, or to multiply it by 3 or 4, dependingon the run (Fig. 1C); the control task required reading single uppercaseletters (C, D, F, G, H, J). The wooden blocks were removed from thehands of the participants during the arithmetic tasks. In total, the

Fig. 1. A. Photograph of the wooden blocks used in the finger discrimination task. During the task, the participants held a different block in each hand, palms up, with the thumb in alateral position. B. Illustration of the participant's grip on the wooden block, with half of the fingers placed over the bumps (upper position) and the others in the holes of the block(down position). C. Time course of each experimental task and its reference. In arithmetic tasks, they had to multiply the Arabic digit by 3 or 4, or to subtract it from 11 or 13. In thefinger discrimination task, the participants were instructed to decide whether the red finger was in a down position by saying “yes” or “no”. Control tasks consisted in letter readingor colour judgements (see Procedure for further details).
1522 M. Andres et al. / NeuroImage 62 (2012) 1520–1528
participants performed 15 trials for each finger of the two hands, 12 tri-als for each digit (or letter) in each arithmetic (or reading) task, exceptfor 3 and 4 (or C and D) that were presented 6 times in each arithmetic(or reading) task.
Procedure
The participants practiced all tasks outside the magnet room inorder to get familiar with the instructions and response requirements.In particular, they were trained to produce audible responses whilekeeping bucco-laryngo-facial movements to a minimum. In themagnetroom, the participants were lying in the scanner with both arms restingalong the body, palms up, and viewed the stimuli projected on a screen,in the rear of the scanner, via a tilted mirror mounted on the head coil.Each experimental task and its control were tested twice in 6 runscounterbalanced across participants. We used a block-design paradigmwith short series of 17,500 ms, interleaved with 10,000 ms fixation pe-riods, to optimize the signal-to-noise ratio while controlling for speech-related head motion artefacts (Birn et al., 2004). Each run consisted of12 series alternating between an experimental task and its control.Each series involved 5 trials where hand drawings or digits/letterswere displayed for 150 mswith a 3500-ms intertrial interval. The partic-ipants were reminded of the instructions at the beginning of each run.Stimulus display was controlled by E-prime 2.0 (Psychology SoftwareTools, Pittsburgh, USA) and the verbal responses were recorded by adigital recorder (for more details about this procedure, see Andreset al., 2011).
Imaging protocol
For each participant, a high-resolution anatomical image wasfirst acquired with a 3.0 Tesla magnetic resonance imager and an8-channel phased array head coil (Achieva, Philips Medical Systems,Andover, MA, USA) using a T1-weighted 3D turbo fast field-echo se-quence with an inversion recovery prepulse (TE=4.6 ms, TR=9.1 ms,
flip angle=8°, field of view=220×197 mm, 150 contiguous axialslices of 1 mm, voxel size=0.81×0.95×1 mm, SENSE factor=1.4).Functional images were then acquired as series of blood-oxygen-sensitive T2*-weighted echo-planar image volumes (GRE-EPI). Eachrun consisted of 132 volumes and was preceded by 4 dummy scansto allow for magnetic saturation effects. Acquisition parameters were:TE=50 ms, TR=2500 ms, flip angle=90°, FOV=220×220 mm, 36slices acquired in an ascending interleaved sequence, slice thickness=3.5 mmwith no interslice gap, SENSE factor (parallel imaging)=2.5.
Data analysis
A first analysis of variance (ANOVA) was performed on error ratesand median response latencies (RLs) of correct trials with TASK (sub-traction, multiplication vs. finger) as a within-subject factor. A secondANOVA was performed on the median RLs in the finger discrimina-tion task with HAND SIDE (left vs. right) and FINGERS (index, middle,ring vs. pinkie) as within-subject factors. This ANOVA was not con-ducted on error rates due to empty cells. Paired-sample t-tests wereused for post-hoc comparisons (pb .05, Bonferroni adjustment formultiple comparisons). Average RLs are reported with standard er-rors (S.E.) corrected for within-subject designs (Loftus and Masson,1994).
The functional data were processed and analyzed using StatisticalParametric Mapping (SPM5, Welcome Department of Cognitive Neu-rology, London, UK, http://www.fil.ion.ucl.ac.uk/spm). The functionalimages were (1) corrected for slice acquisition delays, (2) re-alignedto the first scan of the first run (closest to the anatomical scan) to cor-rect for within- and between-run motion, (3) coregistered with theanatomical image, (4) normalized to the MNI template using an affinefourth degree ß-spline interpolation transformation and a voxel sizeof 2×2×2 mm3 after the skull and bones had been removed with amask based on the individual anatomical images, and (5) spatiallysmoothed using an 8-mm FWHM Gaussian kernel. Condition-relatedBOLD signal changes were estimated for each participant by a general

1523M. Andres et al. / NeuroImage 62 (2012) 1520–1528
linear model in which the responses evoked by each condition weremodelled by a standard hemodynamic response function. The con-trasts of interest were computed at the individual level to identifythe cerebral areas significantly activated by subtraction or multiplica-tion. The significant cerebral activations for the critical contrasts (fin-ger discrimination minus colour judgement; subtraction minus letterreading; multiplication minus letter reading; multiplication minussubtraction; subtraction minus multiplication) were examined at thegroup level in random-effect analyses with the statistical thresholdset at pb0.05 corrected for the false discovery rate (pFDR). In order toreveal the brain areas commonly activated in finger and arithmetictasks, the contrasts computed at the group level were entered in aconjunction analysis using the minimum statistic (MS) compared tothe conjunction null (CN; Nichols et al., 2005).
Voxelwise correlations were then computed between task-relatedchanges of the BOLD signal in the areas revealed by the conjunctionanalysis. If two tasks recruit the same neuronal populations, a positivecorrelation is expected because the pattern of voxels showing lowand high activity increases should be similar for both tasks; dissimilarpatterns of peak voxels between tasks should be reflected by a nullor negative correlation (Dormal et al., 2010; Downing et al., 2007;Peelen and Downing, 2007; Peelen et al., 2006, 2007). The clusters re-vealed by the conjunction analysis were intersected with a 5-mm-radius sphere centred on peak voxels, using the MarsBAR toolbox(http://marsbar.sourceforge.net; Brett et al., 2002), and t-valueswere extracted for each contrast and each participant from a set ofnormalized but unsmoothed data. First, we measured (1) the correla-tion between the t-values of the contrast [finger discrimination–colourjudgement] and the t-values of the contrast [subtraction–letter reading]and (2) the correlation between the t-values of the contrast [fingerdiscrimination–colour judgement] and the t-values of the contrast[multiplication–letter reading] across the voxels of each cluster. Pearsoncoefficients were computed at the individual level and the mean coeffi-cient of each correlation was tested against 0 using a one-sample t-testwith SUBJECT as a random factor (for statistical procedures, see Lorchand Myers, 1990). Because the finger task was correlated both withthe subtraction task andwith themultiplication task, p-values were ad-justed to control for Type 1 error with α=0.05/2. Second, as a control,we measured voxelwise correlations between tasks in 5-mm spherescentred on the peak voxel in the left and right extrastriate body areas(EBA), which were activated during finger discrimination but not dur-ing mental arithmetic. The between-task correlations measured foreach area revealed by the conjunction analysis were directly comparedto the between-task correlations measured for the EBA of the samehemisphere using paired-sample t-tests with SUBJECT as a random factor.A Bonferroni correction was applied to take into account that each EBAwas compared to multiple areas (2 in the left hemisphere, i.e. the hIPSand PSPL, and 3 in the right hemisphere, i.e. the hIPS, PSPL and IFG).Third, the coefficients obtained for homologue parietal areas wereentered in a repeated-measure ANOVA with CORRELATION (finger-subtraction vs. finger-multiplication), SITE (hIPS vs. PSPL) and HEMISPHERE
(left vs. right) as within-subject factors, and SUBJECT as a random factor,in order to investigate the effect of these factors on the size of the coef-ficients. A Bonferroni correction was applied where needed in post-hoccomparisons.
Results
Behavioural results
The error rates did not differ between subtraction (mean±S.E.:3.5±1.1%), multiplication (3.9±0.5%) and finger discrimination(3.3±0.6%; Fb1). A significant effect of TASK was found on RLs(F(2,34)=14.84, pb .001), showing that performance was slower inthe subtraction task (905±23 ms) than in the multiplication (792±24 ms, t(17)=3.20, pb .016) and finger discrimination tasks (695±
20 ms, t(17)=5.02, pb .001; Fig. 2A). In the finger discriminationtask, a main effect of FINGER (F(3,51)=7.55, pb .001) was found, withslower responses for themiddle finger (746±13 ms)when comparedto the index (683±11 ms, t(17)=3.77, pb .01) and the pinkie (661±9 ms, t(17)=4.29, pb .01), and for the ring finger (721±14 ms)whencompared to the pinkie (t(17)=3.69, pb .01). Performance was asfast and accurate for the left (701±8.3 ms and 2.1±0.6%) and righthands (705±8.3 ms and 4.6±1.2%; Fb1).
fMRI results
The contrast betweenmental arithmetic and letter reading showedbilateral activations in the hIPS, in the PSPL, and right-sided activa-tions in the orbital part of the inferior frontal gyrus (IFG; Fig. 2B andTable 1). The direct contrast between multiplication and subtractionrevealed further activations in the left and right superior temporalgyrii (STG). No area was significantly more activated during subtrac-tion than multiplication. The contrast between finger discriminationand colour judgement showed a bilateral occipito-parieto-precentralnetwork (Fig. 2C and Table 1). Increased activity was observed bilater-ally in the EBA, the hIPS, the PSPL and the frontal eye fields (FEF). Clus-ters of activation were also found in the left inferior precentral sulcus(PrCS), the right IFG and the right middle frontal gyrus (MFG). The di-rect contrasts between the left and right hands, and vice-versa, re-vealed additional activations in the contralateral M1 and S1.
The conjunction analysis showed common activations for fingerdiscrimination, subtraction and multiplication in the hIPS and PSPL,bilaterally, and in the right IFG (Fig. 2D). In order to investigate furtherthe functional overlap between finger discrimination and each arith-metic task, we measured voxelwise correlations in each area identifiedin the conjunction analysis. Table 2 provides the Pearson coefficientsfor the correlation between the t-values of the [finger discrimination–colour judgement] contrast and the t-values of the [subtraction–letterreading] contrast, and for the correlation between the t-values of the[finger discrimination–colour judgement] contrast and the t-values ofthe [multiplication–letter reading] contrast for the four parietal clusters.In the hIPS and PSPL, positive correlations were found between the ac-tivation patterns elicited by finger discrimination and each arithmetictask (all r ranging between .3 and .6; all pb .004). Voxelwise correlationsbetween task-related patterns in the EBA, chosen as a control site be-cause of its specific contribution to finger discrimination, showed onlyweak and nonsignificant correlations (all r between −.08 and .2, allp>.1, corrected for multiple comparisons). In the left hemisphere,correlations between finger- and arithmetic-related patterns were sig-nificantly larger in the hIPS and PSPL than in the EBA (all pb .004,corrected for multiple comparisons; Table 2). In the right hemisphere,a significant difference was observed between the PSPL and the EBA(all pb .021, corrected for multiple comparisons) but not between thehIPS and the EBA (all p>.1). The ANOVA with HEMISPHERE (left vs. right),AREA (hIPS vs. PSPL) and CORRELATION (finger-subtraction vs. finger-multiplication) as within-subject factors revealed a main effect of AREA
(F(1,17)=8.34, pb .01), with larger correlations in the PSPL than in thehIPS, and an interaction between AREA and HEMISPHERE (F(1,17)=5.49,pb .032), indicating that the difference between the two areas mainlyconcerned the right hemisphere (right PSPL vs. right hIPS: t(17)=3.3,pb .05). No significant difference was found between activation patternsin the PSPL and the hIPS of the left hemisphere (t(17)=1.56, p>.1). Re-sults also showed a main effect of CORRELATION (F(1,17)=8.23, pb .001),indicating that the activation pattern observed during finger dis-criminationwasmore similar to the one observed during subtractionthan during multiplication across all parietal areas. Finally, correla-tions between activation patterns in the right IFG (finger-subtraction:r=.21±.07; finger-multiplication: r=.22±.08) failed to be significantand did not differ from the correlations measured for the right EBA, cho-sen as a control site (all p>.1).

Fig. 2. A. Mean RLs and S.E. as a function of task (S = subtraction, M = multiplication, F = finger discrimination) and finger. B. Brain areas showing increased activity in mentalarithmetic tasks compared to letter reading (pFDRb .05). C. Brain regions showing increased activity in finger discrimination compared to colour judgements. D. Overlapping acti-vations in finger discrimination and arithmetic tasks. Diagrams represent the percent of signal change in peak voxels (MNI coordinates) for each experimental task (black) andits reference (grey).
1524 M. Andres et al. / NeuroImage 62 (2012) 1520–1528
Discussion
The present study tested the hypothesis that the cerebral networkinvolved in mental arithmetic overlaps with the one involved in fingerdiscrimination. To do so, whole brain activity was measured while par-ticipants solved subtraction and multiplication problems or performed
a finger discrimination task. The behavioural results showed that sub-traction was performed more slowly than multiplication or fingerdiscrimination, whereas the error rate was equal in the three tasks.Discriminating the position of the middle and ring fingers took longerthan discriminating the position of the index and pinkie, which isreminiscent of the greater difficulties of brain-damaged patients
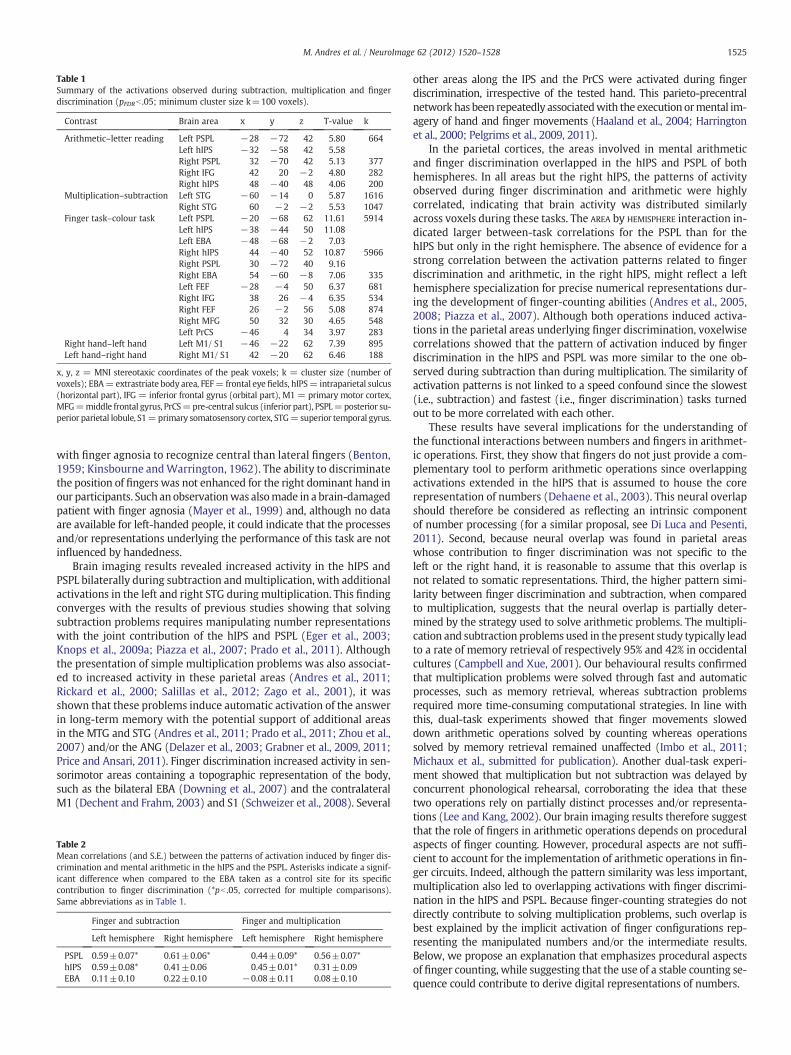
Table 1Summary of the activations observed during subtraction, multiplication and fingerdiscrimination (pFDRb .05; minimum cluster size k=100 voxels).
Contrast Brain area x y z T-value k
Arithmetic–letter reading Left PSPL −28 −72 42 5.80 664Left hIPS −32 −58 42 5.58Right PSPL 32 −70 42 5.13 377Right IFG 42 20 −2 4.80 282Right hIPS 48 −40 48 4.06 200
Multiplication–subtraction Left STG −60 −14 0 5.87 1616Right STG 60 −2 −2 5.53 1047
Finger task–colour task Left PSPL −20 −68 62 11.61 5914Left hIPS −38 −44 50 11.08Left EBA −48 −68 −2 7.03Right hIPS 44 −40 52 10.87 5966Right PSPL 30 −72 40 9.16Right EBA 54 −60 −8 7.06 335Left FEF −28 −4 50 6.37 681Right IFG 38 26 −4 6.35 534Right FEF 26 −2 56 5.08 874Right MFG 50 32 30 4.65 548Left PrCS −46 4 34 3.97 283
Right hand–left hand Left M1/ S1 −46 −22 62 7.39 895Left hand–right hand Right M1/ S1 42 −20 62 6.46 188
x, y, z = MNI stereotaxic coordinates of the peak voxels; k = cluster size (number ofvoxels); EBA=extrastriate body area, FEF= frontal eye fields, hIPS= intraparietal sulcus(horizontal part), IFG = inferior frontal gyrus (orbital part), M1 = primary motor cortex,MFG=middle frontal gyrus, PrCS=pre-central sulcus (inferior part), PSPL=posterior su-perior parietal lobule, S1=primary somatosensory cortex, STG=superior temporal gyrus.
1525M. Andres et al. / NeuroImage 62 (2012) 1520–1528
with finger agnosia to recognize central than lateral fingers (Benton,1959; Kinsbourne andWarrington, 1962). The ability to discriminatethe position of fingers was not enhanced for the right dominant hand inour participants. Such anobservationwas alsomade in a brain-damagedpatient with finger agnosia (Mayer et al., 1999) and, although no dataare available for left-handed people, it could indicate that the processesand/or representations underlying the performance of this task are notinfluenced by handedness.
Brain imaging results revealed increased activity in the hIPS andPSPL bilaterally during subtraction andmultiplication, with additionalactivations in the left and right STG during multiplication. This findingconverges with the results of previous studies showing that solvingsubtraction problems requires manipulating number representationswith the joint contribution of the hIPS and PSPL (Eger et al., 2003;Knops et al., 2009a; Piazza et al., 2007; Prado et al., 2011). Althoughthe presentation of simple multiplication problems was also associat-ed to increased activity in these parietal areas (Andres et al., 2011;Rickard et al., 2000; Salillas et al., 2012; Zago et al., 2001), it wasshown that these problems induce automatic activation of the answerin long-term memory with the potential support of additional areasin the MTG and STG (Andres et al., 2011; Prado et al., 2011; Zhou et al.,2007) and/or the ANG (Delazer et al., 2003; Grabner et al., 2009, 2011;Price and Ansari, 2011). Finger discrimination increased activity in sen-sorimotor areas containing a topographic representation of the body,such as the bilateral EBA (Downing et al., 2007) and the contralateralM1 (Dechent and Frahm, 2003) and S1 (Schweizer et al., 2008). Several
Table 2Mean correlations (and S.E.) between the patterns of activation induced by finger dis-crimination and mental arithmetic in the hIPS and the PSPL. Asterisks indicate a signif-icant difference when compared to the EBA taken as a control site for its specificcontribution to finger discrimination (*pb .05, corrected for multiple comparisons).Same abbreviations as in Table 1.
Finger and subtraction Finger and multiplication
Left hemisphere Right hemisphere Left hemisphere Right hemisphere
PSPL 0.59±0.07* 0.61±0.06* 0.44±0.09* 0.56±0.07*hIPS 0.59±0.08* 0.41±0.06 0.45±0.01* 0.31±0.09EBA 0.11±0.10 0.22±0.10 −0.08±0.11 0.08±0.10
other areas along the IPS and the PrCS were activated during fingerdiscrimination, irrespective of the tested hand. This parieto-precentralnetwork has been repeatedly associatedwith the execution ormental im-agery of hand and finger movements (Haaland et al., 2004; Harringtonet al., 2000; Pelgrims et al., 2009, 2011).
In the parietal cortices, the areas involved in mental arithmeticand finger discrimination overlapped in the hIPS and PSPL of bothhemispheres. In all areas but the right hIPS, the patterns of activityobserved during finger discrimination and arithmetic were highlycorrelated, indicating that brain activity was distributed similarlyacross voxels during these tasks. The AREA by HEMISPHERE interaction in-dicated larger between-task correlations for the PSPL than for thehIPS but only in the right hemisphere. The absence of evidence for astrong correlation between the activation patterns related to fingerdiscrimination and arithmetic, in the right hIPS, might reflect a lefthemisphere specialization for precise numerical representations dur-ing the development of finger-counting abilities (Andres et al., 2005,2008; Piazza et al., 2007). Although both operations induced activa-tions in the parietal areas underlying finger discrimination, voxelwisecorrelations showed that the pattern of activation induced by fingerdiscrimination in the hIPS and PSPL was more similar to the one ob-served during subtraction than during multiplication. The similarity ofactivation patterns is not linked to a speed confound since the slowest(i.e., subtraction) and fastest (i.e., finger discrimination) tasks turnedout to be more correlated with each other.
These results have several implications for the understanding ofthe functional interactions between numbers and fingers in arithmet-ic operations. First, they show that fingers do not just provide a com-plementary tool to perform arithmetic operations since overlappingactivations extended in the hIPS that is assumed to house the corerepresentation of numbers (Dehaene et al., 2003). This neural overlapshould therefore be considered as reflecting an intrinsic componentof number processing (for a similar proposal, see Di Luca and Pesenti,2011). Second, because neural overlap was found in parietal areaswhose contribution to finger discrimination was not specific to theleft or the right hand, it is reasonable to assume that this overlap isnot related to somatic representations. Third, the higher pattern simi-larity between finger discrimination and subtraction, when comparedto multiplication, suggests that the neural overlap is partially deter-mined by the strategy used to solve arithmetic problems. The multipli-cation and subtraction problems used in the present study typically leadto a rate of memory retrieval of respectively 95% and 42% in occidentalcultures (Campbell and Xue, 2001). Our behavioural results confirmedthat multiplication problems were solved through fast and automaticprocesses, such as memory retrieval, whereas subtraction problemsrequired more time-consuming computational strategies. In line withthis, dual-task experiments showed that finger movements sloweddown arithmetic operations solved by counting whereas operationssolved by memory retrieval remained unaffected (Imbo et al., 2011;Michaux et al., submitted for publication). Another dual-task experi-ment showed that multiplication but not subtraction was delayed byconcurrent phonological rehearsal, corroborating the idea that thesetwo operations rely on partially distinct processes and/or representa-tions (Lee and Kang, 2002). Our brain imaging results therefore suggestthat the role of fingers in arithmetic operations depends on proceduralaspects of finger counting. However, procedural aspects are not suffi-cient to account for the implementation of arithmetic operations in fin-ger circuits. Indeed, although the pattern similarity was less important,multiplication also led to overlapping activations with finger discrimi-nation in the hIPS and PSPL. Because finger-counting strategies do notdirectly contribute to solving multiplication problems, such overlap isbest explained by the implicit activation of finger configurations rep-resenting the manipulated numbers and/or the intermediate results.Below, we propose an explanation that emphasizes procedural aspectsof finger counting, while suggesting that the use of a stable counting se-quence could contribute to derive digital representations of numbers.

1526 M. Andres et al. / NeuroImage 62 (2012) 1520–1528
In the finger discrimination task, the absence of visual feedbackput strong demands on the ability to represent the relative position ofthe fingers, as evidenced by increased RLs for the middle fingers thathave more neighbours than the others. The pattern of activation inthis task was analogue to the one observed in the contrast betweencomplex and simple finger movements in other fMRI studies (Haalandet al., 2004; Harrington et al., 2000). These studies showed that activityalong the IPS increased each time a new finger wasmoved, irrespectiveof hand laterality, whereas M1 and S1 were activated in response tocontralateral movements regardless of finger changes. The parietal cor-tex was found to react to finger transitions, whatever the total numberof fingers involved in the sequence, indicating that its contribution wasmainly determined by the need to individuatefingers. Cell recordings inthe monkey homologue of the hIPS showed that neuronal activity inthis area predicts a transition between two different movements, withincremental increases reflecting the number of times a movement isrepeated before a transition (Sawamura et al., 2002). Altogether, theseresults suggest that the parietal network underlying finger discrimina-tion is endowedwith suitable properties for keeping track of incremen-tal changes during arithmetic operations. We therefore propose that,during development, thewiring of number and finger circuits in the pa-rietal cortex enables the implementation of a successor function thatimproves the representation of the number sequence and helps keepingtrack of incremental changes during arithmetic operations. This inter-pretation fits with electrophysiological results showing an increase ofCSE in handmuscles whenever items have to be put in correspondencewith the elements of an ordered series, being the number sequence orthe alphabet (Andres et al., 2007). Importantly, the implementation ofa successor function in the parietal cortex would not only contributeto keep track of incremental changes but it also provides a mediumto represent numbers on fingers. Because finger counting relies on astable sequence (i.e. from the thumb to the pinkie in most occidentalcultures; Lindemann et al., 2011), the relative position of each fingercan be used to infer the cardinality of a number. The digital representa-tions derived from the mapping of the number sequence onto fingerscan in turn support other functions in mental arithmetic, such as thecreation/consolidation of arithmetic facts or the decomposition of aproblem in intermediate steps (Domahs et al., 2008). The latter func-tions could explain the overlap between finger discrimination andmultiplication in the parietal cortex. Along the same lines, the finger in-dividuation process underlying one-to-one correspondence is likely tosupport exact arithmetic operations with small and large numbers,but we do not exclude a more specific contribution to small arithmeticproblems whose answer can be readily represented through configuralrepresentations (Kaufmann et al., 2008; Thompson et al., 2004). In orderto address this issue, future fMRI studies should investigate whetherthe BOLD signal in finger-related areas correlates with problem size inarithmetic tasks.
The hIPS activation extends in areas whose role in visuomotor func-tions, such as pointing or grasping, is well established both in humanand non-human primates (Binkofski et al., 1999; Simon et al., 2002).The same areas have also been related to movement preparation andmotor intention (Göbel et al., 2004; Rushworth et al., 2003; Thoenissenet al., 2002). In contrast, the PSPL is known to interact with the FEF tosupport eye movements and attention shifts (for a meta-analysis, seeGrosbras et al., 2005). The correlation between the pattern of activationobserved in the PSPL during finger discrimination and arithmetic couldtherefore indicate that the two tasks share common resources for atten-tion allocation over the mental representation of fingers and numbers(Hubbard et al., 2005). It has been shown that computing the result ofan arithmetic problem is analogue to shifting attention to the left orright side of a mental number line (Knops et al., 2009b; McCrink et al.,2007; Pinhas and Fischer, 2008). Indeed, a common neural code wasfound for leftward saccades and subtraction and for rightward saccadesand addition in the PSPL (Knops et al., 2009a). The involvement of thePSPL in multiplication problems solved by memory retrieval is more
difficult to reconcile with its role in attention orientation. Severalbrain imaging studies corroborated the finding of increased activity inthe hIPS and PSPL during basic multiplication (Andres et al., 2011;Chochon et al., 1999; Dehaene et al., 1996; Kiefer and Dehaene, 1997;Rickard et al., 2000), sometimes with an extension to the parieto-occipital junction (POJ) known for its role in eye–hand coordination(Prado et al., 2005; Zago et al., 2001). The companion role of the PSPLand POJ in solving basic multiplication problems was also evidencedwith fMRI in a patient whose lesions in the left perisylvian areasprevented her to solve this operation normally (Cohen et al., 2000). Afirst explanation could be that the PSPL contributes to narrow downthe range of potential answers tomultiplication problemsby shifting at-tention along a mental number line, as for addition and multiplication(Dehaene and Cohen, 1991). A recent developmental study showedthat 5-to-7-year-old children are indeed able to approximate the an-swer of non-symbolic multiplication problems above chance level, be-fore they receive formal schooling in this operation (McCrink andSpelke, 2010). Alternatively, it has been proposed that the robust parie-tal activations observed during basic multiplication problems reflectsmagnitude-based strategies that can facilitate their solving (Chochonet al., 1999), for example to reverse the order of the operands or to de-compose a multiplication into a series of additions (Campbell, 1994;LeFevre et al., 1996). In this view, the PSPL would contribute to shiftingattention between operands or zooming in and out the different parts ofa decomposed problem. The FEF were not activated during arithmetictasks in our study but previous studies suggest that these areas also con-tribute to represent numbers on a visuospatial medium (Knops et al.,2009a; Rusconi et al., 2011). Further research is required, however, todetermine the exact nature of the processes shared by finger discrimi-nation and mental arithmetic in the hIPS and PSPL.
Finally, a cluster of activationwas found in the inferior part of the leftPrCS during finger discrimination but not during mental arithmetic, incontrastwith several previous results (Arsalidou and Taylor, 2011). Fur-ther studies are required to explain this discrepancy, for example, byusing a larger set of arithmetic problems. It is worth noting that activa-tion in the left PrCS might have been underestimated in the finger dis-crimination task as well, because the colour judgement task, used as areference, also involved finger drawings. A recent study tested the in-volvement of the left and right premotor/motor cortex as a function ofthe preferred counting direction of the participants (Tschentscher etal., 2012). Viewing numbers between 1 and 5, with both hands at rest,led to higher activation in the right premotor/motor cortex of left-starters when compared to right-starters, showing that the preferredstarting hand influenced the activations induced by small numbers inthe contralateral hemisphere. No difference was observed for numbersranging from 6 to 10. These results suggest that future studies shouldtake into account the relationship between hand preference for finger-based calculation and hemispheric lateralization of motor activity inorder to decipher the role of the premotor/motor cortex inmental arith-metic. In the present study, the only common focus of activation in thefrontal lobe was located in the right IFG. It has been argued that thisarea controls the search within working memory contents (Lepsien etal., 2005). A previous study showed that this area was equally activatedwhennumbers, syllables or locationsweremanipulated vs. rehearsed inworking memory (Zago et al., 2008). Our multi-voxel pattern analysissuggests that numbers and fingers do not activate the same parts ofthe IFG, meaning that the recruitment of this area is not completely in-dependent of the contents stored in working memory.
Conclusion
The present study adds further evidence to the view that the brainareas involved in finger representation might offer a support forarithmetic operations. Results showed that finger discrimination andmental arithmetic share common parietal and frontal areas in theadult brain. In contrast with the EBA and S1/M1, these areas do not

1527M. Andres et al. / NeuroImage 62 (2012) 1520–1528
contribute to somatic representations, as evidenced by a similar increaseof activity while representing the left or right hand. Voxelwise correla-tions were used to explore further the neural overlap between fingerdiscrimination and arithmetic tasks, and similar patterns of activitywere found in the hIPS and PSPL but not in the IFG. Correlations werehigher in the left than in the right hIPS and they were also influencedby arithmetic operation. The parietal areas involved in finger discrimina-tion seem more important to index numerical changes during subtrac-tion than during multiplication, presumably because multiplicationproblems aremostly solved bymemory retrieval with the support of ad-ditional areas in the temporal cortex. Future research should clarify thenature of the neural mechanisms shared by finger discrimination andmental arithmetic in the hIPS and PSPL with a special emphasis on therespective contribution of motor and spatial processes.
Acknowledgments
This work was supported by grants from the Fonds National pourla Recherche Scientifique (grant 1.B099.09, FRS-FNRS, Belgium), andthe Fonds Spéciaux de Recherche of the Université catholique deLouvain (grant FSR09-CWS/09.285).M.A.was a post-doctoral researcher,N.M. is a research fellow and M.P. a research associate at the Fonds Na-tional pour la Recherche Scientifique (FRS-FNRS, Belgium).We are grate-ful to Pierre Leclef for his contribution to the acquisition of fMRI data, toCécile Grandin for the medical supervision, to the Radiodiagnosis Unitat the Cliniques Universitaires St. Luc (Brussels) for technical supportduring fMRI testing, and to Manuela Piazza for her suggestions aboutdata analysis.
References
Andres, M., Seron, X., Olivier, E., 2005. Hemispheric lateralization of number comparison.Cogn. Brain Res. 25, 283–290.
Andres, M., Seron, X., Olivier, E., 2007. Contribution of hand motor circuits to counting.J. Cogn. Neurosci. 19, 563–576.
Andres, M., Olivier, E., Badets, A., 2008. Actions, words, and numbers. Curr. Dir. Psychol.Sci. 17 (5), 313.
Andres, M., Pelgrims, B., Michaux, N., Olivier, E., Pesenti, M., 2011. Role of distinct parietalareas in arithmetic: an fMRI-guided TMS study. Neuroimage 54, 3048–3056.
Arsalidou, M., Taylor, M.J., 2011. Is 2+2=4? Meta-analyses of brain areas needed fornumbers and calculations. Neuroimage 54, 2382–2393.
Badets, A., Pesenti, M., Olivier, E., 2010. Response-effect compatibility of finger-numeralconfigurations in arithmetical context. Q. J. Exp. Psychol. 63, 16–22.
Benton, A.L., 1959. Right–Left Discrimination and Finger Localization: Developmentand Pathology. Hoeber-Harper, New York, NY.
Binkofski, F., Buccino, G., Posse, S., Seitz, R.J., Rizzolatti, G., Freund, H., 1999. A fronto-parietal circuit for object manipulation in man: evidence from an fMRI-study.Eur. J. Neurosci. 11 (9), 3276–3286.
Birn, R.M., Cox, R.W., Bandettini, P.A., 2004. Experimental designs and processing strate-gies for fMRI studies involving overt verbal responses. Neuroimage 23, 1046–1058.
Brett, M., Anton, J.-L., Valabregue, B., Poline, J.-B., 2002. Region of interest analysis usingan SPM toolbox [abstract]. Presented at the 8th International Conference on Func-tional Mapping of the Human Brain, June 2–6, 2002, Sendai, Japan: Available onCD-ROM in NeuroImage, vol. 16, No 2.
Butterworth, B., 1999. The Mathematical Brain. MacMillan, London.Campbell, J.I.D., 1987. Network interference and mental multiplication. J. Exp. Psychol.
Learn. Mem. Cogn. 13, 109–123.Campbell, J.I., 1994. Architectures for numerical cognition. Cognition 53 (1), 1–44.Campbell, J.I.D., Xue, Q., 2001. Cognitive arithmetic across cultures. J. Exp. Psychol. Gen.
130 (2), 299–315.Chochon, F., Cohen, L., Moortele, P.F.V.D., Dehaene, S., 1999. Differential contributions
of the left and right inferior parietal lobules to number processing. J. Cogn. Neurosci.11, 617–630.
Cohen, L., Dehaene, S., Chochon, F., Lehericy, S., Naccache, L., 2000. Language and calcu-lation within parietal lobe: a combined cognitive, anatomical and fMRI study.Neuropsychologia 38 (10), 1426–1440.
Cooney, J.B., Swanson, H.L., Ladd, S.F., 1988. Acquisition of mental multiplication skill:evidence for the transition between counting and retrieval strategies. Cogn. Instr.5, 323–345.
Costa, A.J., Silva, J.B.L., Chagas, P.P., Krinzinger, H., Lonneman, J., Willmes, K., Wood, G.,Haase, V.G., 2011. A hand full of numbers: a role for offloading in arithmetics learning?Front. Psychol. 2 (Article 368).
De Cruz, H., 2006. Why are some numerical concepts more successful than others? Anevolutionary perspective on the history of number concepts. Evol. Hum. Behav. 27,306–323.
Dechent, P., Frahm, J., 2003. Functional somatotopy of finger representations in humanprimary motor cortex. Hum. Brain Mapp. 18, 272–283.
Dehaene, S., Cohen, L., 1991. Two mental calculation systems: a case study of severeacalculia with preserved approximation. Neuropsychologia 29, 1045–1074.
Dehaene, S., Tzourio, N., Frak, V., Raynaud, L., Cohen, L., Mehler, J., Mazoyer, B., 1996.Cerebral activations during number multiplication and comparison: a PET study.Neuropsychologia 34, 1097–1106.
Dehaene, S., Piazza, M., Pinel, P., Cohen, L., 2003. Three parietal circuits for number pro-cessing. Cogn. Neuropsychol. 20, 487–506.
Delazer, M., Domahs, F., Bartha, L., Brenneis, C., Lochy, A., Trieb, T., Benke, T., 2003.Learning complex arithmetic—an fMRI study. Cogn. Brain Res. 18 (1), 76–88.
Delazer, M., Ischebeck, A., Domahs, F., Zamarian, L., Koppelstaetter, F., Siedentopf, C.M.,Kaufmann, L., Benke, T., Felber, S., 2005. Learning by strategies and learning bydrill-evidence from an fMRI study. Neuroimage 25 (3), 838–849.
De Smedt, B., Holloway, I.D., Ansari, D., 2011. Effects of problem size and arithmeticoperation on brain activation during calculation in children with varying levelsof arithmetical fluency. Neuroimage 57 (3), 771–781.
Di Luca, S., Pesenti, M., 2008. Masked priming effect with canonical finger numeralconfigurations. Exp. Brain Res. 185 (1), 27–39.
Di Luca, S., Pesenti, M., 2011. Finger numeral representations: more than just anothersymbolic code. Front. Psychol. 2 (Article 272).
Di Luca, S., Lefèvre, N., Pesenti, M., 2010. Place and summation coding for canonical andnon-canonical finger numeral representations. Cognition 117 (1), 95–100.
Di Luca, S., Granà, A., Semenza, C., Seron, X., Pesenti, M., 2006. Finger-digit compatibilityin Arabic numeral processing. Q. J. Exp. Psychol. 59, 1648–1663.
Domahs, F., Krinzinger, H., Willmes, K., 2008. Mind the gap between both hands: evidencefor internal finger-based number representations in children's mental calculation.Cortex 44, 359–367.
Dormal, V., Andres, M., Dormal, G., Pesenti, M., 2010. Mode-dependent and mode-independent representations of numerosity in the right intraparietal sulcus.Neuroimage 52 (4), 1677–1686.
Downing, P.E., Wiggett, A.J., Peelen, M.V., 2007. Functional magnetic resonance imaginginvestigation of overlapping lateral occipitotemporal activations using multi-voxelpattern analysis. J. Neurosci. 27, 226–233.
Eger, E., Sterzer, P., Russ, M., Giraud, A., Kleinschmidt, A., 2003. A supramodal numberrepresentation in human intraparietal cortex. Neuron 37 (4), 719–725.
Fuson, K.C., 1988. Children's Counting and the Concepts of Number. Springer-Verlag,New York.
Galfano, G., Rusconi, E., Umiltà, C., 2003. Automatic activation of multiplication facts:evidence from the nodes adjacent to the product. Q. J. Exp. Psychol. 56, 31–61.
Göbel, S.M., Johansen-Berg, H., Behrens, T., Rushworth, M.F.S., 2004. Response-selection-related parietal activation during number comparison. J. Cogn. Neurosci. 16 (9),1536–1551.
Grabner, R.H., Ansari, D., Koschutnig, K., Reishofer, G., Ebner, F., Neuper, C., 2009. Toretrieve or to calculate? Left angular gyrus mediates the retrieval of arithmeticfacts during problem solving. Neuropsychologia 47 (2), 604–608.
Grabner, R.H., Ansari, D., Koschutnig, K., Reishofer, G., Ebner, F., 2011. The function ofthe left angular gyrus in mental arithmetic: evidence from the associative confu-sion effect. Hum. Brain Mapp. http://dx.doi.org/10.1002/hbm.21489 (Epub aheadof print).
Grosbras, M.-H., Laird, A.R., Paus, T., 2005. Cortical regions involved in eye movements,shifts of attention, and gaze perception. Hum. Brain Mapp. 25, 140–154.
Haaland, K.Y., Elsinger, C.L., Mayer, A.R., Durgerian, S., Rao, S.M., 2004. Motor sequencecomplexity and performing hand produce differential patterns of hemisphericlateralization. J. Cogn. Neurosci. 16, 621–636.
Harrington, D.L., Rao, S.M., Haaland, K.Y., Bobholz, J.A., Mayer, A.R., Binderx, J.R., Cox,R.W., 2000. Specialized neural systems underlying representations of sequentialmovements. J. Cogn. Neurosci. 12, 56–77.
Hubbard, E.M., Piazza, M., Pinel, P., Dehaene, S., 2005. Interactions between numberand space in parietal cortex. Nat. Rev. Neurosci. 6 (6), 435–448.
Imbo, I., Vandierendonck, A., Fias, W., 2011. Passive hand movements disrupt adults'counting strategies. Front. Psychol. 2, 201.
Kaufmann, L., Vogel, S., Wood, G., Kremser, C., Schocke, M., Zimmerhackl, L., Koten, J.,2008. A developmental fMRI study of nonsymbolic numerical and spatial process-ing. Cortex 44 (4), 376–385.
Kiefer, M., Dehaene, S., 1997. The time course of parietal activation in single-digit mul-tiplication: evidence from event-related potentials. Math. Cogn. 3 (1), 1–30.
Kinsbourne, M., Warrington, E.K., 1962. A study of finger agnosia. Brain 85, 47–66.Knops, A., Thirion, B., Hubbard, E.M., Michel, V., Dehaene, S., 2009a. Recruitment of an area
involved in eye movements during mental arithmetic. Science 324, 1583–1585.Knops, A., Viarouge, A., Dehaene, S., 2009b. Dynamic representations underlying sym-
bolic and nonsymbolic calculation: evidence from the operational momentum ef-fect. Atten. Percept. Psychophys. 71 (4), 803–821.
Lampl, Y., Eshel, Y., Gilad, R., Sarova-Pinhas, I., 1994. Selective acalculia with sparing of thesubtraction process in a patient with left parietotemporal hemorrhage. Neurology 44,1759.
Lee, K., Kang, S., 2002. Arithmetic operation and working memory: differential sup-pression in dual tasks. Cognition 83, B63–B68.
LeFevre, J., Bisanz, J., Daley, K.E., Buffone, L., Greenham, S.L., Sadesky, G.S., 1996. Multipleroutes to solution of single-digit multiplication problems. J. Exp. Psychol. Gen. 125,284–306.
Lepsien, J., Griffin, I.C., Devlin, J.T., Nobre, A.C., 2005. Directing spatial attention in mentalrepresentations: interactions between attentional orienting and working-memoryload. Neuroimage 26, 733–743.
Lindemann, O., Alipour, A., Fischer, M.H., 2011. Finger counting habits in Middle-Easternand Western individuals: an online survey. J. Cross Cult. Psychol. 42, 566–578.

1528 M. Andres et al. / NeuroImage 62 (2012) 1520–1528
Loftus, E.J.R., Masson, L., 1994. Using confidence intervals in within-subject designs.Psychon. Bull. Rev. 1 (4), 476–490.
Lorch, R.F., Myers, J.L., 1990. Regression analysis of repeated measures data in cognitiveresearch. J. Exp. Psychol. Learn. Mem. Cogn. 16, 149–157.
Mayer, E., Martory, M., Pegna, A.J., Landis, T., Delavelle, J., Annoni, J., 1999. A pure caseof Gerstmann syndrome with a subangular lesion. Brain 122, 1107–1120.
McCrink, K., Spelke, E.S., 2010. Coremultiplication in childhood. Cognition 116 (2), 204–216.McCrink, K., Dehaene, S., Dehaene-Lambertz, 2007. Moving along the number line:
operational momentum in nonsymbolic arithmetic. Percept. Psychophys. 69 (8),1324–1333.
Menon, V., Rivera, S.M., White, C.D., Glover, G.H., Reiss, A.L., 2000. Dissociating prefrontaland parietal cortex activation during arithmetic processing. Neuroimage 12, 357–365.
Michaux, N.,Masson, N., Pesenti,M., Andres, M., submitted for publication. Selective inter-ference of finger movements on basic addition and subtraction problem solving. Exp.Psychol.
Nichols, T., Brett, M., Andersson, J., Wager, T., Poline, J.B., 2005. Valid conjunction infer-ence with the minimum statistic. Neuroimage 25, 653–660.
Noël, M.-P., 2005. Finger gnosia: a predictor of numerical abilities in children? ChildNeuropsychol. 11, 413–430.
Oldfield, R.C., 1971. The assessment and analysis of handedness: the Edinburgh inven-tory. Neuropsychologia 9, 97–113.
Peelen, M.V., Downing, P.E., 2007. Using multi-voxel pattern analysis of fMRI data tointerpret overlapping functional activations. Trends Cogn. Sci. 11 (1), 4–5.
Peelen, M.V., Wiggett, A.J., Downing, P.E., 2006. Patterns of fMRI activity dissociateoverlapping functional brain areas that respond to biological motion. Neuron 49(6), 815–822.
Peelen, M.V., Atkinson, A.P., Andersson, F., Vuilleumier, P., 2007. Emotional modulationof body-selective visual areas. Soc. Cogn. Affect. Neurosci. 2 (4), 274–283.
Pelgrims, B., Andres, M., Olivier, E., 2009. Double dissociation between motor and visualimagery in the posterior parietal cortex. Cereb. Cortex 19, 2298–2307.
Pelgrims, B., Michaux, N., Olivier, E., Andres, M., 2011. Contribution of the primary motorcortex tomotor imagery: a subthreshold TMS study. Hum. BrainMapp. 32, 1471–1482.
Pesenti, M., Thioux, M., Seron, X., De Volder, A.D., 2000. Neuroanatomical substrates ofArabic number processing, numerical comparison, and simple addition: a PETstudy. J. Cogn. Neurosci. 12, 461–479.
Piazza, M., Mechelli, A., Price, C.J., Butterworth, B., 2006. Exact and approximate judge-ments of visual and auditory numerosity: an fMRI study. Brain Res. 1106, 177–188.
Piazza, M., Pinel, P., Le Bihan, D., Dehaene, S., 2007. A magnitude code common tonumerosities and number symbols in human intraparietal cortex. Neuron 53 (2),293–305.
Pinhas, M., Fischer, M., 2008. Mental movements without magnitude? A study of spatialbiases in symbolic arithmetic. Cognition 109 (3), 408–415.
Prado, J., Clavagnier, S., Otzenberger, H., Scheiber, C., Kennedy, H., Perenin, M., 2005.Two cortical systems for reaching in central and peripheral vision. Neuron 48 (5),849–858.
Prado, J., Mutreja, R., Zhang, H., Mehta, R., Desroches, A.S., Minas, J.E., Booth, J.R., 2011.Distinct representations of subtraction and multiplication in the neural systems fornumerosity and language. Hum. Brain Mapp. 32, 1932–1947.
Price, G.R., Ansari, D., 2011. Symbol processing in the left angular gyrus: evidence frompassive perception of digits. Neuroimage 57, 1205–1211.
Rickard, T.C., Romero, S.G., Basso, G., Wharton, C., Flitman, S., Grafman, J., 2000. The cal-culating brain: an fMRI study. Neuropsychologia 38 (3), 325–335.
Rusconi, E., Bueti, D., Walsh, V., Butterworth, B., 2011. Contribution of frontal cortex tothe spatial representation of number. Cortex 47, 2–13.
Rushworth, M.S.F., Johansen-Berg, H., Göbel, S.M., Devlin, J.T., 2003. The left parietaland premotor cortices: motor attention and selection. Neuroimage 20, S89–S100.
Salillas, E., Semenza, C., Basso, D., Vecchi, T., Siegal, M., 2012. Single pulse TMS induceddisruption to right and left parietal cortex on addition and multiplication.Neuroimage 59 (4), 3159–3165.
Sandrini, M., Miozzo, A., Cotelli, M., Cappa, S., 2003. The residual calculation abilities ofa patient with severe aphasia: evidence for a selective deficit of subtraction proce-dures. Cortex 39, 85–96.
Sato, M., Cattaneo, L., Rizzolatti, G., Gallese, V., 2007. Numbers within our hands: modu-lation of corticospinal excitability of hand muscles during numerical judgment.J. Cogn. Neurosci. 19, 684–693.
Sawamura, H., Shima, K., Tanji, J., 2002. Numerical representation for action in theparietal cortex of the monkey. Nature 415, 918–922.
Schweizer, R., Voit, D., Frahm, J., 2008. Finger representations in human primarysomatosensory cortex as revealed by high-resolution functional MRI of tactile stimu-lation. Neuroimage 42, 28–35.
Simon, O., Mangin, J.F., Cohen, L., Le Bihan, D., Dehaene, S., 2002. Topographical layoutof hand, eye, calculation, and language-related areas in the human parietal lobe.Neuron 33 (3), 475–487.
Stanescu-Cosson, R., Pinel, P., van de Moortele, P., Le Bihan, D., Cohen, L., Dehaene, S.,2000. Understanding dissociations in dyscalculia: a brain imaging study of theimpact of number size on the cerebral networks for exact and approximate calcula-tion. Brain 123, 2240–2255.
Takayama, Y., Sugishita, M., Akigushi, I., Kimura, J., 1994. Isolated acalculia due to leftparietal lesion. Arch. Neurol. 21, 286–294.
Thoenissen, D., Zilles, K., Toni, I., 2002. Differential involvement of parietal andprecentral regions in movement preparation and motor intention. J. Neurosci. 22(20), 9024–9034.
Thompson, J., Abbott, D., Wheaton, K., Syngeniotis, A., Puce, A., 2004. Digit representa-tion is more than just hand waving. Cogn. Brain Res. 21 (3), 412–417.
Tschentscher, N., Hauk, O., Fischer, M.H., Pulvermüller, F., 2012. You can count on themotor cortex: finger counting habits modulate motor cortex activation evokedby numbers. Neuroimage 59 (4), 1–10.
Williams, B.P., Williams, R.S., 1995. Finger numbers in the Greco-Roman world and theearly middle ages. Isis 86, 587–608.
Zago, L., Pesenti, M., Mellet, E., Crivello, F., Mazoyer, B., Tzouriomazoyer, N., 2001. Neuralcorrelates of simple and complex mental calculation. Neuroimage 13 (2), 314–327.
Zago, L., Petit, L., Turbelin, M.-R., Andersson, F., Vigneau, M., Tzourio-Mazoyer, N., 2008.How verbal and spatial manipulation networks contribute to calculation: an fMRIstudy. Neuropsychologia 46, 2403–2414.
Zhou, X., Chen, C., Zang, Y., Dong, Q., Chen, C., Qiao, S., Gong, Q., 2007. Dissociated brainorganization for single-digit addition andmultiplication. Neuroimage 35 (2), 871–880.





























