Embed Size (px)
Citation preview

Colworth
MedalLecture
Colworth Medal LectureThe twin-arginine transport system: movingfolded proteins across membranesF. Sargent1
Centre for Metalloprotein Spectroscopy and Biology, School of Biological Sciences, University of East Anglia, Norwich NR4 7TJ, U.K.
Colworth Medal Lecture
Delivered at the SECC, Glasgow,
on 12 July 2007
Frank Sargent
AbstractThe Tat (twin-arginine transport) pathway is a protein-targetingsystem dedicated to the transmembrane translocation of fullyfolded proteins. This system is highly prevalent in thecytoplasmic membranes of bacteria and archaea, and is alsofound in the thylakoid membranes of plant chloroplasts andpossibly also in the inner membrane of plant mitochondria.Proteins are targeted to a membrane-embedded Tat trans-locase by specialized N-terminal twin-arginine signal peptidesbearing an SRRXFLK amino acid motif. The genes encodingcomponents of the Tat translocase were discovered approx.10 years ago, and, since then, research in this area hasexpanded on a global scale. In this review, the key discoveriesin this field are summarized, and recent studies of bacterialtwin-arginine signal-peptide-binding proteins are discussed.
IntroductionThe efficient targeting of proteins to their sites of physio-logical function is an essential feature of all biologicalsystems. The Tat (twin-arginine transport) system is aprotein-targeting pathway found in all kingdoms of life, andthe transport mechanism has been explored at the molecularlevel most extensively in bacteria and the chloroplastsof higher plants. The Tat system is also active in somearchaea, and genetic analysis suggests that it is present in
Key words: membrane protein, membrane transport, molecular chaperone, protein–protein
interaction, protein targeting, twin-arginine translocase.
Abbreviations used: GFP, green fluorescent protein; HiPIP, high potential iron protein; Tat,
twin-arginine transport; TMAO, trimethylamine N-oxide; TPP, thylakoidal-processing protease.1Present address: Division of Molecular and Environmental Microbiology, College of Life
Sciences, University of Dundee DD1 5EH, Scotland, U. K. (email [email protected]).
plant mitochondria. Moreover, although the Tat system isapparently not a feature of human physiology, recent genomicanalysis points to the presence of this protein-targetingsystem in the mitochondria of at least one animal, albeit asponge [1]. Approx. 10 years ago, the discovery of the genesencoding this novel transport system [2,3], which was quicklyfollowed by the verification, characterization, and naming ofthe ‘Tat pathway’ [4,5], generated much excitement because,unlike any other general protein transport pathway, thissystem has evolved for the transmembrane translocation offully folded proteins across energy-transducing membranes.
In bacteria, the Tat pathway is central to a range of import-ant cellular functions, since proteins targeted by thissystem have been shown to be involved in respiratory andphotosynthetic electron-transfer chains, cell division and bio-genesis of the cell envelope, quorum sensing and motility,symbiosis and pathogenesis [6,7]. In plant chloroplasts, theTat system is equally important and is principally involvedin the assembly of the oxygen-evolving complex and thecytochrome b6f complex within the thylakoid membrane[8,9]. The Tat pathway is clearly of prokaryotic origin, andrecent breakthroughs have come from microbial researchfocused on the Gram-negative bacterium Escherichia coli, theGram-positive bacterium Bacillus subtilis and members ofthe industrially and medically important actinomycetes. Thisreview will attempt to integrate the key discoveries unearthedusing these micro-organisms with the invaluable biochemicalinsights that are still being gained by studying Tat transportin plants, the original studies of which laid the foundationsfor what has become a huge global research effort.
The Tat protein transport systemIn principle, all protein-targeting systems must rely onan amino acid ‘signal’ displayed by the passenger protein.Proteins exported by the Tat pathway are synthesized asprecursors with N-terminal signal peptides bearing conservedSRRXFLK ‘twin-arginine’ amino acid sequence motifs [10].Some examples of twin-arginine signal peptides are given inFigure 1. All twin-arginine signal peptides have a commontripartite structure that includes a polar N-terminal (n-) re-gion of variable length, a moderately hydrophobic (h-) region
C©The Authors Journal compilation C©2007 Biochemical Society 835Bio
chem
ical
So
ciet
y T
ran
sact
ion
s
ww
w.b
ioch
emso
ctra
ns.
org

836 Biochemical Society Transactions (2007) Volume 35, part 5
Figure 1 Twin-arginine signal peptides
(A) A schematic representation of a twin-arginine signal peptide. The n- h- and c-regions are shown, with the position
of signal peptidase cleavage site indicated by an arrow. The + symbol indicates the position of a positive charge. A basic
residue is often found also in the c-region, where it serves a Sec-avoidance function [164]. (B–F) Amino acid sequences of
a representative selection of twin-arginine signal peptides across the kingdoms: E. coli TorA (B), E. coli NapA (C), wheat
OEC23 (after removal of the N-terminal chloroplast transit peptide) (D), Streptomyces lividans xylanase-C (XlnC) (E), and
halocyanin-1 from Haloferax volcanii [165] (F).
of 12–20 amino acids and a C-terminal (c-) region that oftencontains basic residues [10,11] (Figure 1A). The consensusmotif is always located at the junction between the n- andh-regions [10]. Although there are a handful of exceptions(see, e.g., [9,12,13]), the twin-arginine dipeptide is extremelywell conserved and is central to the efficient operation ofTat signal peptides. This sequence conservation, when put incontext with the more general tripartite structure of the signalpeptide, has enabled bioinformaticians to develop useful toolsto help identify twin-arginine signal peptides from genomicdata [14,15]. Of course, in silico predictions should always becoupled with a reliable in vivo transport-activity assay, suchas those engineered by Tullman-Ercek et al. [16] or Widdicket al. [17]. The E. coli genome, for example, is predicted toencode 29 proteins bearing twin-arginine signal peptides, and,of these, 27 are genuine Tat-dependent export signals [16].
All precursors synthesized with bona fide twin-argininesignal peptides are ultimately transported across a membraneby the twin-arginine translocation apparatus: a membrane-embedded ‘translocase’ that catalyses protein transport.Although the crystal structure of even a fragment of thislarge transport machinery is not yet forthcoming, 10 years ofresearch has provided a wealth of biochemical, biophysicaland genetic data. In general, three types of integral membraneprotein have been identified as components of the Tattranslocase (Figure 2): TatA (also called Tha4 and Tha9 in theplant chloroplast system), TatB (Hcf106 in plants), and TatC(for a more detailed discussion of Tat components, see otherrecent reviews, e.g. [18–20]). TatA- and TatB-type proteinsare sequence-related and are predicted to share a commonstructure comprising an N-terminal transmembrane α-helix,followed by an adjacent (basic) amphipathic helix and anunstructured C-terminal region of variable length [21,22].The TatC protein is the largest, most hydrophobic and mosthighly sequence-conserved Tat component. Its most commonmanifestation is as a polytopic protein with six membrane-
spanning domains and both N- and C-termini on the cis sideof the membrane [5,23,24]. However, gene fusions have beenobserved in some archaea that give rise to TatC proteins with14 transmembrane helices (see Figure 2C), which reinforcesgenetic and biochemical evidence that the minimum TatCunit present in membranes is a homodimer [25,26]. Of theseknown Tat components, the TatC-type protein is ubiquitousin all prokaryotes and eukaryotes known or predicted toutilize the Tat transport system. In all biological systemsproven experimentally to undergo Tat transport, a TatA-typeprotein is also essential for protein transport, and mutations inthe genes encoding either TatC or TatA will impair transportof all associated traffic on the Tat pathway [4,5,27,28].
So how and what do each of these types of protein con-tribute to the transport process? The key biochemical studieshave been carried out in bacteria (primarily in E. coli) andin chloroplasts. Two distinct high-molecular-mass complexeshave been identified: a ‘transport channel module’ containingmultiple TatA protomers [29,30] and a ‘signal-recognitionmodule’ comprising a complex of TatB and TatC [31–33].
Early in the transport process, twin-arginine signalpeptides are recognized by the TatBC (Hcf106-TatC inthylakoids) ‘signal-recognition complex’ [31,32]. Cross-linking studies of both E. coli and thylakoidal Tat systemshave shown that the important twin-arginine dipeptide isclose to the TatC protein, while TatB cross-links most readilyto the hydrophobic h-region of the signal peptide [31,32,34].TatBC varies in size between 370 and ∼650 kDa, dependingon the experimental conditions, and comprises equimolaramounts of TatB and TatC [31,33,35–37]. Cysteine-scanningmutagenesis and disulfide cross-linking has shown that,within the TatBC complex, TatB is arranged as a higher-orderhomo-oligomer, probably at least a tetramer [38]. Consistentwith this, a similar study of the E. coli TatC protein has shownthat TatC is organized at least as a homodimer and that eachof the six transmembrane helices make self-contacts [26].
C©The Authors Journal compilation C©2007 Biochemical Society

The twin-arginine transport system: moving folded proteins across membranes 837
Figure 2 Components of the twin-arginine translocase
Schematic representation of Tat system components from a representative selection of experimentally studied systems
across the kingdoms: (A) E. coli K-12, (B) Bacillus subtilis, (C) Halobacterium sp. NRC-1, and (D) Maize thylakoids. For each
panel, TatA family proteins (Tha4 and Tha9 in maize thylakoids) are green, TatB proteins (Hcf106 in thlyakoids) are yellow
and TatC proteins are pink. The relative postion of C-termini are designated by the letter ‘C’.
Taken together, this suggests a tertiary structure in whichTatC is located at the periphery of a central TatB bundle[26,38]. Negative-stain electron microscopy has also beenused to study TatBC complexes from the Gram-negativebacteria E. coli, Salmonella enterica serotype Typhimurium
and Agrobacterium tumefaciens [39]. In each case, oval-shaped particles were observed that were ‘lobed’, and theanalysis of the area covered by each lobe suggested that eachcould represent a bundle of seven or eight transmembranehelices, enough to correspond to a single TatBC unit [39].
C©The Authors Journal compilation C©2007 Biochemical Society

838 Biochemical Society Transactions (2007) Volume 35, part 5
Under resting conditions, the TatA-comprised ‘transportchannel module’ of the Tat translocase exists as a separatecomplex from TatBC. Low-resolution structural analysisutilizing electron microscopy shows toroid-shaped particles(quite distinct from the oval TatBC particles) with thickwalls, channels through the middle and asymmetric ‘lids’ atone of the ends [29]. These isolated TatA complexes were aheterologous mix of sizes. Although the width of the thickwalls and the ‘height’ were essentially identical for each sizeof particle, the key differences lay in the capacity of thecentral cavities. This structural analysis suggested stronglythat different numbers of TatA protomers could combine togenerate the different-sized channels needed to transport adiverse array of folded proteins across a membrane [29].
Clearly, the signal-binding module and the channel modulemust interact efficiently, and probably extensively, during theprotein-export process. Indeed, some preparations of isolatedTatBC do contain traces of TatA, suggesting strongly thatthere is a TatA-binding site within the TatBC complex. Themost compelling current model for a ‘Tat-transport cycle’ isbased on elegant in vitro work using thylakoid membranes[30,31,34,40,41]. Initial signal peptide binding to TatBC,which is dependent on the transmembrane protonmotiveforce [40], induces a conformational change in TatBC thatexposes a TatA-binding site [31,41]. Thus signal peptidebinding, in conjunction with the protonmotive force, inducesassembly of the complete translocase. The binding pocketfor TatA may be on the TatC protein, since pre-incubationof thylakoid membranes with a specific anti-TatC antibodyprevented the formation of the TatABC translocase [30]. Atthis point, the signal peptide is extended so that the c-regionis now exposed on the trans side of the membrane, and someresearchers believe this action is the ‘power-stroke’ thatpropels the passenger domain across through the channel [42],although this would probably limit the size of passenger thatcould be ‘pulled’. The n-region of the signal peptide remainson the cis side of the membrane throughout this process, andindeed can be covalently bonded to TatC before translocationwith no effect on the transport of the substrate [43,32].This ultimate transport step is probably also energized bythe transmembrane proton electrochemical gradient [44,45],although this is controversial. Following transport, theTatABC complex dissociates again, perhaps because the signalpeptide somehow disengages itself from its binding site onTatC.
Although the scheme described above fits well with thecurrent data coming from E. coli and thylakoid studies, thereare numerous alternative models that can be constructedbased on other studies of Tat subunit structure and function.The roles of members of the wider TatA and TatB families ishotly debated in the literature. For instance, TatA-type pro-teins are less than 100 amino acids in length, but, despite theirsmall size, they have generated a disproportionate amount ofcontroversy. In E. coli, the TatA protein is fully integrated intothe inner membrane [29,46,47]. However, the model Gram-positive bacterium B. subtilis contains three copies of TatAand studies of one of these suggests a fraction may be soluble
in the cell cytoplasm [48,49]. Similarly the Gram-positiveactinomycete Streptomyces lividans has also been shown tocontain a proportion of water-soluble cytoplasmic TatA andTatB [50]. This has led to the alternative hypothesis, whichresonates with the original ideas of Chanal et al. [21], thatTatA may be a cycling ‘receptor’ subunit. Indeed, peptidearray experiments [48] and surface plasmon resonance data[51] suggest that the TatA has affinity for signal peptide-bearing precursor proteins in these biological systems.
The topological organization of E. coli TatA is also notwithout controversy. A recent study using thiol-specificcross-linking suggested that this protein had a topologywhere the extreme N-terminus was exposed to the cytoplasm[52]. This was a very surprising find, since it contradicts the‘positive-inside rule’ postulated by Sipos and von Heijne [53].The evidence for an N-out topology for membrane-boundTatA proteins was strengthened recently by a study of TatAoperation in the opportunistic bacterial pathogen Providenciastuartii [7]. The TatA protein in P. stuartii is synthesized asan inactive ‘zymogen’ bearing an N-terminal extension ofseven amino acids which is sufficient to completely annulcellular Tat transport activity [7]. Why TatA should beinactivated by this addition remains unclear; however, theextended protein is membrane-bound [7], which suggeststhat impairment of membrane integration is not the reason.Very interestingly, Tat translocation in P. stuartii is actuallyinstigated by proteolytic removal of the seven-amino-acidextension from TatA, which activates the protein [7]. It isknown that the TatA transmembrane helix is responsiblefor oligomerization of TatA in the membrane [47], probablyinitially into bundles of at least three TatA protomers[29,46] and ultimately as large ring-shaped channels [29]. Itis possible that the N-terminally extended inactive TatA isin monomeric form in the membrane and that proteolyticcleavage enables the essential oligomerization of the TatAtransmembrane helices to occur [29,46]. This process isuseful in inferring the topology of TatA, since the proteolyticevent must occur when TatA is already in the membrane. Theprotease responsible has been unequivocally identified as amember of the GlpG family of rhomboid serine proteases[7]. Crystal structures of these integral membrane proteinsare available (see, e.g., [54,55]), and it can be clearly seenthat the active site of GlpG family proteases is located closerto the periplasmic side of the membrane [54,55], suggestingstrongly that TatA must adopt an N-out orientation in orderto be efficiently cleaved and thus activated (Figure 3).
The TatB protein is also interesting. Sequence analysisclearly shows that TatB and TatA are distantly relatedand thus probably arose from a common ancestor [21,22].The Gram-negative E. coli TatB protein and the Hcf106equivalent in plants are essential for Tat transport ofphysiological substrates and clearly have physiological rolesdistinct from their TatA-like cousins [2,56]. However, inmost Gram-positive bacteria, with the notable exception ofthe actinomycetes, the Tat translocation system comprisesonly TatA-like and TatC-like proteins [27,28]. In these cases,it seems most likely that the Gram-positive TatA proteins
C©The Authors Journal compilation C©2007 Biochemical Society

The twin-arginine transport system: moving folded proteins across membranes 839
Figure 3 Proteolytic activation of the TatA protein from Providencia stuartii
The TatA protein is show in green. The seven-amino-acid N-terminal extension that results in inactivation of the Tat system
is shown in purple. Stevenson et al. [7] have demonstrated that proteolytic removal of the N-terminal extension activates
the Tat transport stystem. The protease responsible is called AarA and is a member of the GlpG family of rhomboid proteases
[7]. The crystal structure of E. coli GlpG [55] is shown in blue and the catalytic diad residues Ser201 and His254 close to the
periplasmic side, which are located close to the periplasmic side of the membrane [55], are shown in red. It is possible
that the role of the N-terminal extension may be to suppress oligomerization of TatA, which, when removed, results in TatA
assembling first into bundles and finally into channel-like structures. TatA is shown in the N-out orientation as a monomer
[47], which would enable easy access to the AarA active site, and as hairpin-like structures following oligomerization to form
thick-walled channels [29,166].
retain both biological activities. Indeed, a study using anin vivo genetic screen for successful Tat transport has isolatedvariant bifunctional E. coli TatA proteins that can bypassthe requirement for TatB [57]. This work suggests that mostGram-positive bacteria utlize a Tat system componentmost closely related to the ancient progenitor of thefunctionally distinct TatA-like and TatB-like proteins.
Ante-transport events: protein folding andtransport quality-control systemsThe evidence that the prokaryotic and thylakoidal Tattranslocase transports folded proteins is extremely strong[58–64]. The central dogma of the Tat research field hastherefore come to be that this system is specifically dedicatedto the transmembrane translocation of fully folded proteins.Does this mean that the Tat transporter is physically unableto transport unfolded polypeptides? DeLisa et al. [64] suggestthat the bacterial Tat translocase may be able to sense regionsof unfolded protein, much like a housekeeping chaperoninsenses the exposed hydrophobic core of a misfolded protein,and thus reject any immature proteins attempting export(designated folding ‘quality control’ [64]). However, it hasbeen shown for the thylakoid Tat system that at least somemalfolded protein can be successfully translocated [65].
A number of reports have indicated that general house-keeping chaperones such as DnaK and GroEL interact withTat-dependent substrates [66–69]. Though chaperonin con-tamination is often seen in recombinant protein and protein–protein interaction studies (see, e.g., [70,71]), Graubner et al.[68] have demonstrated convincingly that DnaK binds to the
CueO full-length precursor and was essential for the exportof CueO by the Tat pathway. The latter may be good evidencethat the mature domain of CueO cannot reach a stable tertiaryfold without the chaperone activity of DnaK and is thusrejected by the Tat translocase quality-control system; moreevidence that the Tat system transports folded proteins. Theformer, however, suggests that either CueO is not completelyfolded prior to translocation, which raises the questions ofwhen is it correctly folded and when is DnaK released, orthat DnaK interacts specifically with the CueO signal peptideand that this interaction is essential for Tat transport. Itis known that general chaperones such as DnaK do bindto isolated twin-arginine signal peptides [68,69,72], mostprobably through interaction with the h-region. This is notaltogether surprising, however, since the physiological role ofthe DnaK chaperonin system is to recognize and bind non-specifically to exposed, unstructured, hydrophobic regions ofpolypeptide [73] and this is exactly what Tat signal peptidespossess [74,75]. It is unlikely, however, that the interactionbetween signal peptide h-regions and DnaK is involveddirectly in the Tat-export pathway, since covalent attachmentof the CueO signal peptide on to the mature domain ofHiPIP (high potential iron protein) did not confer DnaK-dependence on translocation of the CueO::HiPIP fusion [68].However, such signal-swap experiments may also muddy thewaters somewhat, since the nature of the passenger domainhas a direct effect on the function of the Tat signal peptide.For example, one of the most heavily exploited bacterial twin-arginine signal peptides is that of the TMAO (trimethylamineN-oxide) reductase (TorA) from E. coli. The TorA signal
C©The Authors Journal compilation C©2007 Biochemical Society

840 Biochemical Society Transactions (2007) Volume 35, part 5
peptide has been attached to GFP (green fluorescent protein)[62,63,76], colicin V [77], PhoA [64], OEC23 [4], LepB [11],MalE [78], DMSO reductase [79], hydrogenase-2 [80] andGFOR (glucose/fructose oxidoreductase) [81]. Interestingly,the nature of the passenger protein to which the TorA signalis attached seems to have an influence on its operation atthe molecular level. For example, a mutant SRKRFLA TorATat motif was incapable of transporting the native TMAOreductase to the periplasm [82]; however, TorA signalscarrying this very sequence still directed export of both GFP[63] and colicin V [77]. Clearly, understanding and harnessingthis effect is of paramount importance if the Tat pathway isto be used for industrial-scale protein production; however,currently, the molecular basis by which passenger proteinsexert influence on signal peptide activity remains unknown.
Ante-transport events: cofactor loadingand proofreading systemsThe E. coli TMAO reductase enzyme is a water-solubleperiplasmic protein that contains a single redox cofactor andis synthesized with a twin-arginine signal peptide. The avail-ability of crystal structures for the TorA homologues [83–86],together with its high abundance under certain growth condi-tions, and its tractability to various in vitro and in-gel activityassays [4,60], has led to this particular periplasmic proteinenjoying the attention of Tat researchers from the outset.E. coli TMAO reductase is encoded by the torA gene withinthe torCAD operon [87] and is a member of a vast familyof molybdenum cofactor-containing redox enzymes [88,89].Correct loading of the molybdenum cofactor into the TorAapoprotein is essential before Tat transport can proceed [60],and this suggests either that the Tat translocase simply cannottransport a non-globular/unfolded substrate or that some-thing is repressing signal peptide activity until the cofactor isloaded [60]. The reality is probably a combination of both.
In the case of TorA, a dedicated signal-peptide-bindingcytoplasmic protein has been positively identified. E. coliTorD is a 199-amino-acid cytoplasmic protein that has beenshown to bind specifically to the TorA apoenzyme andmaintain it in a conformation suitable for cofactor loading[90–95]. Although it seems likely that TorD binds to TorA atmore than one locale [80,92], in vivo two-hybrid analysis [80]and in vitro calorimetry [96] have established unequivocallythat TorD binds directly to the TorA twin-arginine signalpeptide. Optimum binding by TorD appears to involve a 27-residue core region of the TorA signal peptide stretching fromthe twin-arginine motif, through the h-region, to anotherarginine dipeptide at the C-terminus [96]. The twin-argininedipeptide itself is not critical for the binding since a transport-inactive ‘twin-lysine’ variant signal peptide is also recognizedby TorD [96]. In addition, calorimetry clearly showed thatTorD binds to the TorA signal peptide in an energeticallyfavourable reaction with an apparent dissociation constant(Kd) for the 27-residue synthetic peptide of ∼1 µM [96].Similar calorimetry experiments using a fusion of the full-length TorA signal peptide (39 amino acids) to the C-terminus of maltose-binding protein point to an apparent
Kd of ∼60 nM for TorD binding (J. Maillard and F. Sargent,unpublished work). Having established that TorD does bindto the TorA signal, the next pressing questions are why doesTorD attach to the signal peptide and when and how is thesignal peptide ‘handed-over’ to the Tat translocase?
Biophysical studies of the twin-arginine signal peptidesof the Allochromatium vinosum HiPIP [74] and the E. coliSufI protein [75] suggest that Tat signal peptides are largelyunstructured in aqueous solution. Such peptides are un-doubtedly susceptible to opportunistic (or otherwise)binding by housekeeping chaperonins (see previous section),and probably also proteolytic enzymes. Indeed, when eitherthe TorA apoenzyme or a fusion of the TorA signal peptideto GFP was synthesized in vivo, the signal peptide wasrapidly degraded unless excess TorD was provided [94,97]. Itis therefore conceivable that the role of TorD is to ‘protect’the signal peptide from the molecular environment of the cellcytoplasm before transport [94,97]. However, experimentsdesigned to unravel the physiological activity of TorDsuggest that it has other functions in addition to protectingTorA from proteolysis [80].
E. coli hydrogenase-2 is a heterodimer of a catalyticα-subunit (HybC) and an electron-transferring β-subunit(HybO), which also bears the twin-arginine signal peptide[98]. The αβ dimer is transported to the periplasm as afully active folded complex by the Tat translocase [4,99].Because HybC has no signal sequences whatsoever, it isclearly important that assembly of the HybOC dimeris carefully co-ordinated in order to prevent prematureexport of HybO without its HybC partner. The key tothis co-ordination lies in the primary structure of the nativeHybO signal peptide, since swapping it for the TorA signalpeptide impaired assembly of the hydrogenase and resultedin premature targeting of HybO to the membrane withoutits HybC partner [80]. The removal of the native signalpeptide probably also removed the activity of a biosyntheticaccessory protein (HybE) whose job was to assist in theassembly of HybOC [80,100]. Most surprising, however,was that the introduction of the TorA signal peptide tothe hydrogenase system also introduced a requirement forTorD for the correct assembly of this otherwise completelyalien enzyme [80]. Overproduction of TorD in the strainexpressing the TorA::HybO fusion protein led to the returnof cellular hydrogenase activity because the HybOC dimerwas now correctly assembled before export [80].
The ability of the TorD/TorA signal peptide couple todirect assembly of the hydrogenase provided a facile in vivoassay for TorD activity. Moreover, using this assay, it waspossible to concentrate solely on any activity associatedspecifically with signal peptide binding. TorD family proteinscontain a highly conserved EPXDH amino acid motif wherethe aspartate residue is invariable and the histidine residueis only very occasionally replaced by tyrosine [92,101,102].Variant TorD proteins in which alanine had been substitutedfor either the histidine or the aspartate residue were nolonger able to rescue hydrogenase activity in the strainproducing a TorA::HybO fusion [80]. However, the in vitro
C©The Authors Journal compilation C©2007 Biochemical Society
The twin-arginine transport system: moving folded proteins across membranes 841
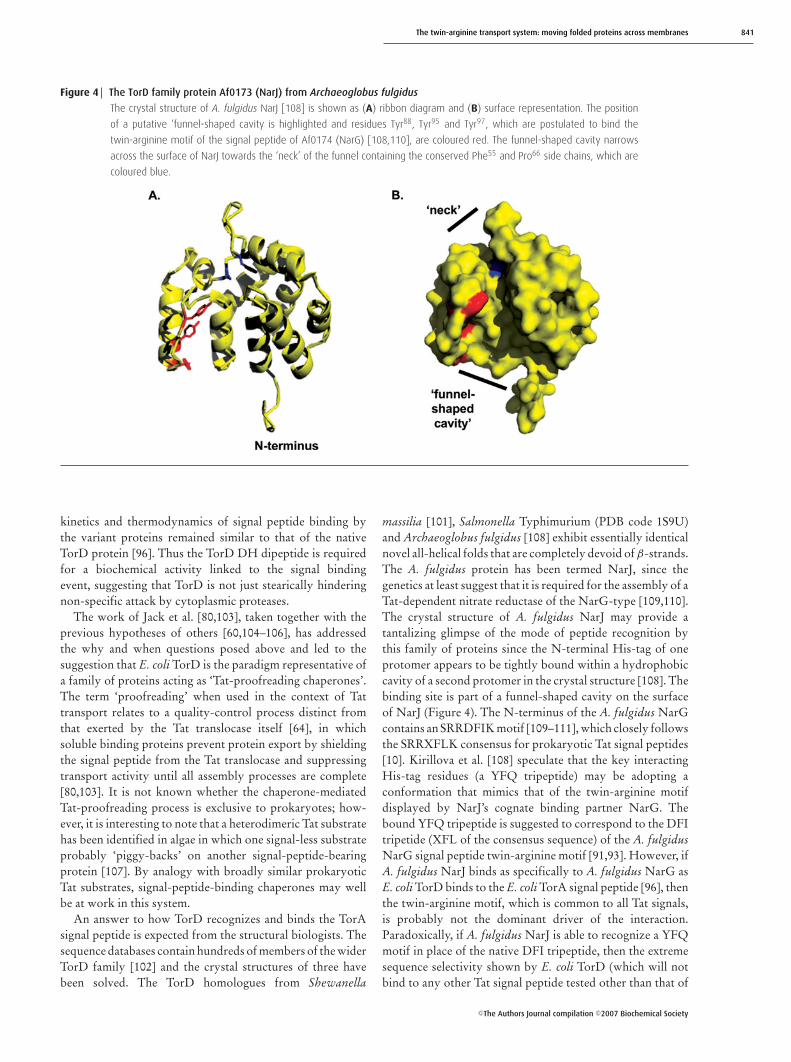
Figure 4 The TorD family protein Af0173 (NarJ) from Archaeoglobus fulgidus
The crystal structure of A. fulgidus NarJ [108] is shown as (A) ribbon diagram and (B) surface representation. The position
of a putative ‘funnel-shaped cavity is highlighted and residues Tyr88, Tyr95 and Tyr97, which are postulated to bind the
twin-arginine motif of the signal peptide of Af0174 (NarG) [108,110], are coloured red. The funnel-shaped cavity narrows
across the surface of NarJ towards the ‘neck’ of the funnel containing the conserved Phe55 and Pro66 side chains, which are
coloured blue.
kinetics and thermodynamics of signal peptide binding bythe variant proteins remained similar to that of the nativeTorD protein [96]. Thus the TorD DH dipeptide is requiredfor a biochemical activity linked to the signal bindingevent, suggesting that TorD is not just stearically hinderingnon-specific attack by cytoplasmic proteases.
The work of Jack et al. [80,103], taken together with theprevious hypotheses of others [60,104–106], has addressedthe why and when questions posed above and led to thesuggestion that E. coli TorD is the paradigm representative ofa family of proteins acting as ‘Tat-proofreading chaperones’.The term ‘proofreading’ when used in the context of Tattransport relates to a quality-control process distinct fromthat exerted by the Tat translocase itself [64], in whichsoluble binding proteins prevent protein export by shieldingthe signal peptide from the Tat translocase and suppressingtransport activity until all assembly processes are complete[80,103]. It is not known whether the chaperone-mediatedTat-proofreading process is exclusive to prokaryotes; how-ever, it is interesting to note that a heterodimeric Tat substratehas been identified in algae in which one signal-less substrateprobably ‘piggy-backs’ on another signal-peptide-bearingprotein [107]. By analogy with broadly similar prokaryoticTat substrates, signal-peptide-binding chaperones may wellbe at work in this system.
An answer to how TorD recognizes and binds the TorAsignal peptide is expected from the structural biologists. Thesequence databases contain hundreds of members of the widerTorD family [102] and the crystal structures of three havebeen solved. The TorD homologues from Shewanella
massilia [101], Salmonella Typhimurium (PDB code 1S9U)and Archaeoglobus fulgidus [108] exhibit essentially identicalnovel all-helical folds that are completely devoid of β-strands.The A. fulgidus protein has been termed NarJ, since thegenetics at least suggest that it is required for the assembly of aTat-dependent nitrate reductase of the NarG-type [109,110].The crystal structure of A. fulgidus NarJ may provide atantalizing glimpse of the mode of peptide recognition bythis family of proteins since the N-terminal His-tag of oneprotomer appears to be tightly bound within a hydrophobiccavity of a second protomer in the crystal structure [108]. Thebinding site is part of a funnel-shaped cavity on the surfaceof NarJ (Figure 4). The N-terminus of the A. fulgidus NarGcontains an SRRDFIK motif [109–111], which closely followsthe SRRXFLK consensus for prokaryotic Tat signal peptides[10]. Kirillova et al. [108] speculate that the key interactingHis-tag residues (a YFQ tripeptide) may be adopting aconformation that mimics that of the twin-arginine motifdisplayed by NarJ’s cognate binding partner NarG. Thebound YFQ tripeptide is suggested to correspond to the DFItripetide (XFL of the consensus sequence) of the A. fulgidusNarG signal peptide twin-arginine motif [91,93]. However, ifA. fulgidus NarJ binds as specifically to A. fulgidus NarG asE. coli TorD binds to the E. coli TorA signal peptide [96], thenthe twin-arginine motif, which is common to all Tat signals,is probably not the dominant driver of the interaction.Paradoxically, if A. fulgidus NarJ is able to recognize a YFQmotif in place of the native DFI tripeptide, then the extremesequence selectivity shown by E. coli TorD (which will notbind to any other Tat signal peptide tested other than that of
C©The Authors Journal compilation C©2007 Biochemical Society
842 Biochemical Society Transactions (2007) Volume 35, part 5
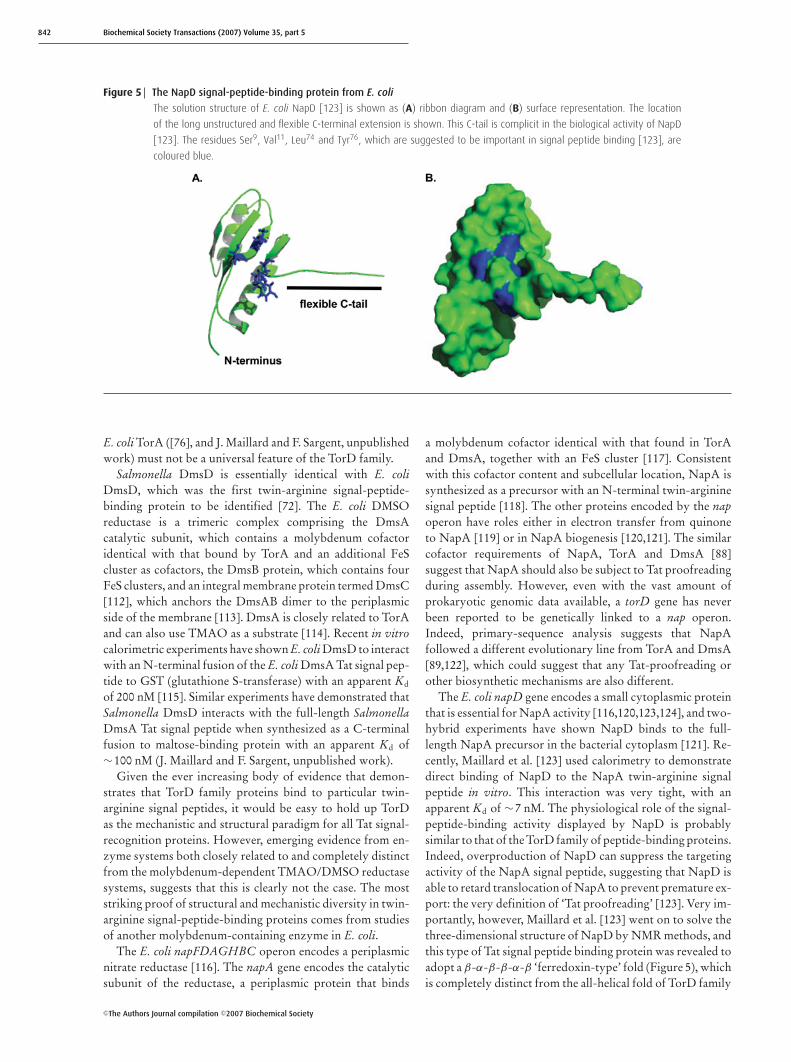
Figure 5 The NapD signal-peptide-binding protein from E. coli
The solution structure of E. coli NapD [123] is shown as (A) ribbon diagram and (B) surface representation. The location
of the long unstructured and flexible C-terminal extension is shown. This C-tail is complicit in the biological activity of NapD
[123]. The residues Ser9, Val11, Leu74 and Tyr76, which are suggested to be important in signal peptide binding [123], are
coloured blue.
E. coli TorA ([76], and J. Maillard and F. Sargent, unpublishedwork) must not be a universal feature of the TorD family.
Salmonella DmsD is essentially identical with E. coliDmsD, which was the first twin-arginine signal-peptide-binding protein to be identified [72]. The E. coli DMSOreductase is a trimeric complex comprising the DmsAcatalytic subunit, which contains a molybdenum cofactoridentical with that bound by TorA and an additional FeScluster as cofactors, the DmsB protein, which contains fourFeS clusters, and an integral membrane protein termed DmsC[112], which anchors the DmsAB dimer to the periplasmicside of the membrane [113]. DmsA is closely related to TorAand can also use TMAO as a substrate [114]. Recent in vitrocalorimetric experiments have shown E. coli DmsD to interactwith an N-terminal fusion of the E. coli DmsA Tat signal pep-tide to GST (glutathione S-transferase) with an apparent Kd
of 200 nM [115]. Similar experiments have demonstrated thatSalmonella DmsD interacts with the full-length SalmonellaDmsA Tat signal peptide when synthesized as a C-terminalfusion to maltose-binding protein with an apparent Kd of∼100 nM (J. Maillard and F. Sargent, unpublished work).
Given the ever increasing body of evidence that demon-strates that TorD family proteins bind to particular twin-arginine signal peptides, it would be easy to hold up TorDas the mechanistic and structural paradigm for all Tat signal-recognition proteins. However, emerging evidence from en-zyme systems both closely related to and completely distinctfrom the molybdenum-dependent TMAO/DMSO reductasesystems, suggests that this is clearly not the case. The moststriking proof of structural and mechanistic diversity in twin-arginine signal-peptide-binding proteins comes from studiesof another molybdenum-containing enzyme in E. coli.
The E. coli napFDAGHBC operon encodes a periplasmicnitrate reductase [116]. The napA gene encodes the catalyticsubunit of the reductase, a periplasmic protein that binds
a molybdenum cofactor identical with that found in TorAand DmsA, together with an FeS cluster [117]. Consistentwith this cofactor content and subcellular location, NapA issynthesized as a precursor with an N-terminal twin-argininesignal peptide [118]. The other proteins encoded by the napoperon have roles either in electron transfer from quinoneto NapA [119] or in NapA biogenesis [120,121]. The similarcofactor requirements of NapA, TorA and DmsA [88]suggest that NapA should also be subject to Tat proofreadingduring assembly. However, even with the vast amount ofprokaryotic genomic data available, a torD gene has neverbeen reported to be genetically linked to a nap operon.Indeed, primary-sequence analysis suggests that NapAfollowed a different evolutionary line from TorA and DmsA[89,122], which could suggest that any Tat-proofreading orother biosynthetic mechanisms are also different.
The E. coli napD gene encodes a small cytoplasmic proteinthat is essential for NapA activity [116,120,123,124], and two-hybrid experiments have shown NapD binds to the full-length NapA precursor in the bacterial cytoplasm [121]. Re-cently, Maillard et al. [123] used calorimetry to demonstratedirect binding of NapD to the NapA twin-arginine signalpeptide in vitro. This interaction was very tight, with anapparent Kd of ∼7 nM. The physiological role of the signal-peptide-binding activity displayed by NapD is probablysimilar to that of the TorD family of peptide-binding proteins.Indeed, overproduction of NapD can suppress the targetingactivity of the NapA signal peptide, suggesting that NapD isable to retard translocation of NapA to prevent premature ex-port: the very definition of ‘Tat proofreading’ [123]. Very im-portantly, however, Maillard et al. [123] went on to solve thethree-dimensional structure of NapD by NMR methods, andthis type of Tat signal peptide binding protein was revealed toadopt a β-α-β-β-α-β ‘ferredoxin-type’ fold (Figure 5), whichis completely distinct from the all-helical fold of TorD family
C©The Authors Journal compilation C©2007 Biochemical Society

The twin-arginine transport system: moving folded proteins across membranes 843
proteins [110,123]. The four β-strands assemble together togenerate a single antiparallel β-sheet that forms one face of theprotein, while the two α-helices together form the oppositeface (Figure 5). Clearly, the huge structural differencesbetween E. coli NapD and A. fulgidus NarJ means that thereis no analogous signal-peptide-binding funnel identifiable onNapD. Further NMR experiments instead identified a patchof residues within the β-sheet face of NapD where the localenvironment was disrupted upon peptide binding, suggestingstrongly that the peptide-binding site lies within the β-sheet(Figure 5) [123]. This is consistent with what is knownabout the functions of other members of the ferredoxin-likefamily; indeed, so often is the β-sheet region of such proteinsimplicated in mediating protein–protein or protein–ligand in-teractions that it has been described as a ‘supersite’ for binding[125].
E. coli TorD and E. coli NapD are therefore theparadigm members of two large and different families oftwin-arginine signal-peptide-binding proteins. The utterlydissimilar structures of these two families probably points tounrelated molecular mechanisms of signal peptide recogni-tion and demonstrates convergent evolution at work in a keybiological process in the most dramatic fashion. Indeed, it isvery likely that further families of specific Tat signal-bindingproteins exist in the prokaryotic world. Genomic analysishas predicted possible Tat-proofreading chaperones for anumber of complex bacterial Tat substrates [6,18]. Althoughhard biochemical data on signal peptide binding is not yetavailable, structural genomics consortia are beginning torelease structural information on some of these proteins. Forexample, the NMR solution structure of E. coli HyaE reveals athioredoxin-like fold (PDB code 2HFD), unlike that of TorDor NapD. There is some genetic, but as yet no biochemical,evidence that this protein interacts with the precursor form ofa Tat-dependent hydrogenase [106]. In addition, the crystalstructure of the FdhE protein from Pseudomonas aeruginosahas been solved (PDB code 2FIY). FdhE is a rubredoxinhomodimer and has a fold unrelated to that of NapD, TorD orHyaE. In E. coli, the fdhE gene is essential for the biosynthesisof Tat-dependent formate dehydrogenases (multi-subunitmolybdenum-containing enzymes), but is not required forthe assembly of non-exported formate dehydrogenases [126].There is currently no evidence that this protein binds directlyto a twin-arginine signal peptide or even to a Tat-dependentprecursor protein.
Taken together, these studies highlight the amazingstructural diversity of chaperone-mediated Tat-proofreadingsystems either known or predicted to be operating on thebacterial Tat pathway. It is worth reiterating that all twin-arginine signal peptides must interact efficiently with the Tatmachinery to facilitate export, and the twin-arginine motif isone of the keys to the transport activity. Where, then, doesthe very strict chaperone specificity come from? How are theessential structural features for transport balanced withthe equally essential structural features for chaperone bind-ing? One future challenge will be to isolate each activityfor a selection of signal peptides and attempt to unravel
the molecular basis of this dual functionality. Indeed, thesequestions also overlap with understanding how a Tat signalpeptide is handed over from a cytoplasmic chaperone to theTat translocase.
Post-transport events: peptide processingThe vast majority of known and predicted Tat substratesare water-soluble globular proteins. Although most, if notall, will have adopted their native folds before the exportevent, there are still biosynthetic hurdles to overcome aftertranslocation. Some Tat-dependent bacterial proteins, inparticular those that contain certain types of copper cofactor[127,128], do not bind their prosthetic groups until aftertranslocation. In addition, some Tat-dependent prokarytoticproteins may undergo a significant covalent modification bythe addition of a lipid group following transport [17,129].The Shewanella DMSO reductases, for example, are Tat-dependent molybdenum-containing enzymes that arepredicted to be lipidated following export across the cytoplas-mic membrane and then must be secreted across the outermembrane (probably via the Type II apparatus [130]) tothe cell surface [131]. However, far and away the mostcommon, and the most significant, post-translational eventin the biosynthesis of the vast majority of Tat substrates isproteolytic cleavage of the twin-arginine signal peptide.
Signal peptide cleavage probably occurs at a late stage in thetransport process, probably well after complete transport ofthe globular domain. What is the evidence for this? First, notall Tat substrates have cleaved signal peptides. The Rieske FeSproteins, which form part of the cytochrome b6f complex inplant thylakoids, and the cytochrome bc1 and some nitratereductase complexes in prokaryotes, are synthesized withfunctional N-terminal Tat signals that then act as transmem-brane signal anchors [9,132–135]. Clearly, if signal removalwas a critical early step in the export process, biosynthesisof this type of protein would be impossible. Secondly, bothprokaryotic and thylakoidal Tat substrates that are known topossess cleavable signal peptides contain obvious protease-recognition sequences within their c-regions. In the case ofprokaryotes, this is either an AxA ‘type-I’ signal peptidasemotif [10] or an [L/I/G/A][A/G/S]C ‘lipo box’ motif for‘type-II’ signal peptidases (reviewed in [136]), and in the caseof thylakoid Tat substrates there is usually an AXA TPP(thylakoidal-processing protease) recognition motif [137].The active site of each of these proteases is located on thetrans side of the membrane [137,138], which means cleavablesignal peptides must be fully extended, either through the Tatchannel or across the bilayer, to be accessible. Again, giventhat the signal peptide first interacts with the Tat apparatus ina loop conformation [43], full extension of the signal peptidewould only be expected near the end of the transport process.Indeed, recent evidence suggests that signal processing occursat such a late stage that the signal has probably beenreleased from the Tat apparatus following translocation of thepassenger domain. In the study of Gerard and Cline [34],the N-terminus of a twin-arginine signal peptide was firstcovalently cross-linked to thylakoidal TatC before transport
C©The Authors Journal compilation C©2007 Biochemical Society

844 Biochemical Society Transactions (2007) Volume 35, part 5
was initiated. Despite tethering the N-terminus signal peptideat the cis side of the membrane, the passenger domain couldstill be translocated [34]. Significantly, however, the signalpeptide remained unprocessed, suggesting strongly that TPPhad no access to this very-late-stage complex between the Tatmachinery and a Tat-dependent precursor [34].
Taken together, it is conceivable that all signal peptidesbehave, albeit fleetingly, as N-terminal signal anchors at a latestage of translocation. In this model, once fully extended, thesignal peptide would escape laterally from the Tat complexthrough a designated ‘side-gate’ in either the Tat channel orthe signal recognition complex, or perhaps be released tolipid if dissipation of the Tat subunits occurs at the end oftranslocation. The hydrophobicity of the lipid would inducespontaneous α-helix formation in the Tat signal peptide,which would then be subjected to proteolysis if appropriate.The escape of the signal peptide must follow on very quicklyafter full extension of the signal peptide, since overloadingof the Tat pathway with substrate can lead to processing ofthe signal before complete translocation of the passengerdomain, resulting in back-sliding of processed protein [139].This model also suggests that any mutagenesis of the signalpeptidase-recognition motifs within the signal peptides, orinactivation of the signal peptidases themselves, could resultin signal-anchored membrane-bound Tat substrates. In stud-ies of the thylakoid Tat system using isolated chloroplasts,removal of the TPP-recognition sequence from the signalpeptide of a heterologous Tat substrate (GFP) preventedproteolytic processing but did not impair Tat transport of thereporter protein [139]. In the E. coli system, mutagenesis ofthe AXA motif of the DmsA Tat signal peptide had no effecton the assembly or physiological activity of the DmsABCDMSO reductase complex [79]. In this case, however, itappeared that the signal peptide was either being opportunist-ically cleaved at a different location or the NXN substitutionused was not sufficient to block processing by LepB [79].
Why is it so important to know what happens to thesignal peptide at the end of translocation? As well as possiblemedical and biotechnological applications (discussed below),the route of escape taken by Tat signals from the Tatapparatus may be related to the molecular mechanism ofmembrane protein integration adopted by the Tat system.In both bacteria and chloroplasts, a subset of Tat-targetedproteins are genuine integral membrane proteins synthesizedwith single internal or C-terminal transmembrane helices[140,141]. In all cases, precursors are synthesized with N-terminal twin-arginine signal peptides that are immediatelylinked to globular water-soluble domains, which are followedby hydrophobic transmembrane helices. Integration of thetransmembrane helices into the lipid bilayer is strictlyTat-dependent [140,141]. However, the very existence of Tat-targeted integral membrane proteins poses a number oftricky questions. For instance, early on in the biosyntheticlife of these types of Tat substrates, there are clear issues ofsolubility: how, for example, is aggregation of such exposedhydrophobic helices prevented? Indeed, in the case of thechloroplast proteins, how is such a protein successfully
negotiated through the envelope membranes before beingintegrated into the thylakoid membrane? There appears to benothing unusual in terms of overall hydrophobicity or aminoacid content about Tat-targeted transmebrane helices whencompared with those of other membrane proteins [140]. It ispossible that cytoplasmic chaperonins or dedicated accessoryproteins could mask such helices before transport, in muchthe same way that a Tat-proofreading chaperone wouldmask an exposed signal peptide. However, such bindingproteins are yet to be found. The successful Tat-dependenttranslocation of an otherwise folded protein bearing a longexposed hydrophobic helix raises questions, again, about theability of the Tat translocase to recognize and actively rejectunfolded polypeptides: the ‘quality-control’ mechanism [64].Clearly, Tat-dependent membrane proteins are not rejected.Unfortunately, very little is known about the molecularmechanisms of either process to enable further comment.
Medical and biotechnological applicationsInvading bacterial plant and animal pathogens often evadehost cell responses by the expression of numerous and diverse‘virulence factors’ (reviewed by Finlay and Falkow [142]).The bacterial Tat system itself has been found to be an import-ant virulence factor in plant [143–146] and animal [147–152]pathogens. Since the Tat translocase is not a feature of humanor other higher animal physiologies, the role of the Tat path-way in the virulence of animal pathogens has attracted someinterest from the biomedical sector as a possible target fornovel drug development. Why does the loss of Tat transportcompromise bacterial virulence? There may well turn out tobe a different explanation for each bacterium under investiga-tion. For example, disruption of Tat transport in the E. coli canhave gross pleiotropic effects on outer membrane integrityand cell division [153,154], which would undoubtedly lead toincreased susceptibility to host defences. Alternatively, thenon-export of a single important Tat-dependent virulencefactor, such as the Tat-dependent hydrogenase of Helico-bacter pylori [155], would also be sufficient to compromisevirulence. Furthermore, in Ps. aeruginosa, the Tat translocasehas been shown to operate as the initial export machineryin the two-step secretion of phospholipase virulence factors[130].
Because most Tat mutant strains that have been engineeredin bacteria are generally still viable, a drug that couldspecifically inhibit the Tat translocase would probably notkill a pathogenic bacterium outright, but may serve to preventinfection and/or proliferation. In addition, commensalbacteria that can survive under fermentative conditions, orthat do not utilize Tat-dependent transport (e.g. lactobacilli),would, in principle, not be adversely affected by such ananti-infective agent. A drug that would specifically inhibitTat transport could act in a number of ways, for exampleto affect the mechanism of energy transduction (which iscurrently unknown, but may be related to that used by themitochondrial F1Fo ATPase [136,156]), the mechanism oftwin-arginine signal peptide recognition, or perhaps themechanism of disengaging signal peptides or transmembrane
C©The Authors Journal compilation C©2007 Biochemical Society

The twin-arginine transport system: moving folded proteins across membranes 845
helices from the TatA channel. Since these latter two keyactivities involve peptide recognition, a peptide antimicrobialcould be the answer to providing specific inhibition of thebacterial Tat pathway. Antimicrobial peptides are usually lessthan 40 amino acids long and are produced naturally by allanimals and plants [157]. It is conceivable that, by combiningresearch into the physiological role of the Tat pathway inmodel laboratory organisms such as E. coli, Salmonella orPseudomonas with the latest peptide synthesis and rapidscreening technologies, a powerful selection protocol couldbe developed for identifying anti-Tat transport peptides thatwould mimic a Tat-dependent signal or C-tail.
In addition to attracting interest as an antibacterial target,the Tat system is also gaining increasing attention from the bi-otechnology sector, principally because of its remarkable abil-ity to move folded proteins across biological membranes. Theapplication of this system for biotechnological purposes hasrecently been extensively reviewed by Bruser [158]; however,there are a few interesting advances worth reiterating here.Harnessing the ability of the Tat system to actively select forfolded substrates would undoubtedly increase the efficiencyof large-scale protein production protocols. Fisher et al. [159]addressed this very issue and, using an ingenious tripartitefusion with a Tat signal at the N-terminus, a β-lactamaseat the C-terminus and the protein of interest sandwiched inthe middle, were able to actively select for variant fusionproteins with increased water solubility. The Tat systemhas also been exploited in several other protein-engineeringprojects, including a novel two-hybrid system for screeningfor protein–protein interactions [160], production andengineering of single-chain antibodies [64,161] and for phagedisplay [162,163]. Some bacteria, in particular Gram-positiveStreptomyces spp., export large numbers of diverse substratesby the Tat system and might be ideal hosts for the heterolog-ous production of pharmaceutically important proteins thatare incompatible with secretion by the Sec pathway [17].
Concluding remarksThe Tat protein transport system, which was given its namejust over 9 years ago [4], has flourished into a large anddynamic research field, as is evident from the number ofpapers cited in this review, and the equal number of papersregrettably left out owing to space constraints. There areemerging signs that scientists may be beginning to agree onthe basic principles by which the Tat system operates. Thefuture acquisition of high-resolution structural informationwill pave the way for molecular understanding of Tattransport and for further exploitation of this remarkableprotein transport pathway.
I dedicate the 2007 Colworth Medal Lecture to my wife Tracy
Palmer and our boys James and Jack. I thank David J. Richardson
(University of East Anglia) and Ben C. Berks (University of Oxford)
for guidance, support and many useful discussions. I acknowledge
the hard work and dedication of the numerous postdocs, students
and technical staff who have worked with me in Norwich. You have
all contributed something important to this prize award. Thanks also
to members of Tracy Palmer’s and Ben Berks’s research groups,
past and present, for their valuable input over the years. In addition
to Tracy, Ben and David, I also thank Stuart Ferguson (Oxford), Tony
Pugsley (Paris), Ray Dixon (Norwich), George Georgiou (Austin) and
Dave Kelly (Sheffield) for supporting me in my early research career.
I also acknowledge The Royal Society, the BBSRC (Biotechnology
and Biological Sciences Research Council), and the John and Pamela
Salter Charitable Trust for financial support.
References1 Wang, X. and Lavrov, D.V. (2007) Mol. Biol. Evol. 24, 363–3732 Settles, A.M., Yonetani, A., Baron, A., Bush, D.R., Cline, K. and
Martienssen, R. (1997) Science 278, 1467–14703 Weiner, J.H., Bilous, P.T., Shaw, G.M., Lubitz, S.P., Frost, L., Thomas,
G.H., Cole, J.A. and Turner, R.J. (1998) Cell 93, 93–1014 Sargent, F., Bogsch, E.G., Stanley, N.R., Wexler, M., Robinson, C.,
Berks, B.C. and Palmer, T. (1998) EMBO J. 17, 3640–36505 Bogsch, E.G., Sargent, F., Stanley, N.R., Berks, B.C., Robinson, C. and
Palmer, T. (1998) 273, 18003–180066 Berks, B.C., Palmer, T. and Sargent, F. (2005) Curr. Opin. Microbiol. 8,
174–1817 Stevenson, L.G., Strisovsky, K., Clemmer, K.M., Bhatt, S., Freeman, M.
and Rather, P.N. (2007) Proc. Natl. Acad. Sci. U.S.A. 104, 1003–10088 Mould, R.M. and Robinson, C. (1991) J. Biol. Chem. 266,
12189–121939 Mølik, S., Karnauchov, I., Weidlich, C., Herrmann, R.G. and Klosgen,
R.B. (2001) J. Biol. Chem. 276, 42761–4276610 Berks, B.C. (1996) Mol. Microbiol. 22, 393–40411 Cristobal, S., de Gier, J.-W., Nielsen, H. and von Heijne, G. (1999)
EMBO J. 18, 2982–299012 Hinsley, A.P., Stanley, N.R., Palmer, T. and Berks, B.C. (2001) FEBS
Lett. 497, 45–4913 Ignatova, Z., Hornle, C., Nurk, A. and Kasche, V. (2002)
Biochem. Biophys. Res. Commun. 291, 146–14914 Rose, R.W., Bruser, T., Kissinger, J.C. and Pohlschroder, M. (2002)
Mol. Microbiol. 45, 943–95015 Bendtsen, J.D., Nielsen, H., Widdick, D., Palmer, T. and Brunak, S.
(2005) BMC Bioinform. 6, 167–17516 Tullman-Ercek, D., DeLisa, M., Kawarasaki, Y., Iranpour, P., Ribnicky, B.,
Palmer, T. and Georgiou, G. (2007) J. Biol. Chem. 282, 8309–831617 Widdick, D.A., Dilks, K., Chandra, G., Bottrill, A., Naldrett, M.,
Pohlschroder, M. and Palmer, T. (2006) Proc. Natl. Acad. Sci. U.S.A.103, 17927–17932
18 Berks, B.C., Palmer, T. and Sargent, F. (2003) Adv. Microb. Physiol. 47,187–254
19 Lee, P.A., Tullman-Ercek, D. and Georgiou, G. (2006) Annu. Rev.Microbiol. 60, 373–395
20 Cline, K. and Theg, S.M. (2007) in The Enzymes 25: MolecularMachines Involved in Protein Transport across Cellular Membranes(Tamanoi, F., Dalbey, R.E. and Kohler, C., eds.), Academic Press,San Diego
21 Chanal, A., Santini, C.-L. and Wu, L.-F. (1998) Mol. Microbiol. 30,674–676
22 Yen, M.-R., Tseng, Y.H., Nguyen, E.H., Wu, L.-F. and Saier, Jr, M.H.(2002) Arch. Microbiol. 177, 441–450
23 Behrendt, J., Standar, K., Lindenstrauss, U. and Bruser, T. (2004)FEMS Microbiol. Lett. 234, 303–308
24 Ki, J.J., Kawarasaki, Y., Gam, J., Harvey, B.R., Iverson, B.L. andGeorgiou, G. (2004) J. Mol. Biol. 341, 901–909
25 Buchanan, G., de Leeuw, E., Stanley, N.R., Wexler, M., Berks, B.C.,Sargent, F. and Palmer, T. (2002) Mol. Microbiol. 43, 1457–1470
26 Punginelli, C., Maldonado, B., Grahl, S., Jack, R., Alami, M., Schroder, J.,Berks, B.C. and Palmer, T. (2007) J. Bacteriol. 189, 5482–5494
27 Pop, O., Martin, U., Abel, C. and Muller, J.P. (2002) J. Biol. Chem. 277,3268–3273
28 Jongbloed, J.D.H., Grieger, U., Antelmann, H., Hecker, M., Nijland, R.,Bron, S. and van Dijl, J.M. (2004) Mol. Microbiol. 54, 1319–1325
C©The Authors Journal compilation C©2007 Biochemical Society

846 Biochemical Society Transactions (2007) Volume 35, part 5
29 Gohlke, U., Pullan, L., McDevitt, C.A., Porcelli, I., de Leeuw, E., Palmer,T., Saibil, H.R. and Berks, B.C. (2005) Proc. Natl. Acad. Sci. U.S.A. 102,10482–10486
30 Dabney-Smith, C., Mori, H. and Cline, K. (2006) J. Biol. Chem. 281,5476–5483
31 Cline, K. and Mori, H. (2001) J. Cell Biol. 154, 719–72932 Alami, M., Luke, I., Deitermann, S., Eisner, G., Koch, H.G., Brunner, J.
and Muller, M. (2003) Mol. Cell 12, 937–94633 Bolhuis, A., Mathers, J.E., Thomas, J.D., Barrett, C.M. and Robinson, C.
(2001) J. Biol. Chem. 276, 20213–2021934 Gerard, F. and Cline, K. (2006) J. Biol. Chem. 281, 6130–613535 de Leeuw, E., Granjon, T., Porcelli, I., Alami, M., Carr, S.B., Muller, M.,
Sargent, F., Palmer, T. and Berks, B.C. (2002) J. Mol. Biol. 322,1135–1146
36 McDevitt, C.A., Buchanan, G., Sargent, F., Palmer, T. and Berks, B.C.(2006) FEBS J. 273, 5656–5668
37 Richter, S. and Bruser, T. (2005) J. Biol. Chem. 280, 42723–4273038 Lee, P.A., Orriss, G., Buchanan, G., Greene, N.P., Bond, P.J., Punginelli,
C., Jack, R.L., Sansom, M.S.P., Berks, B.C. and Palmer, T. (2006)J. Biol. Chem. 281, 34072–34085
39 Oates, J., Mathers, J., Mangels, D., Kuhlbrandt, W., Robinson, C. andModel, K. (2003) J. Mol. Biol. 330, 277–286
40 Gerard, F. and Cline, K. (2007) J. Biol .Chem. 282, 5263–752241 Mori, H. and Cline, K. (2002) J. Cell. Biol. 157, 205–21042 Bruser, T. and Sanders, C. (2003) Microbiol. Res. 158, 7–1743 Fincher, V., McCaffery, M. and Cline, K. (1998) FEBS Lett. 423, 66–7044 Alder, N.N. and Theg, S.M. (2003) Cell 112, 231–24245 Braun, N.A., Davis, A.W. and Theg, S. (2007) Biophys. J. 93,
1993–199846 de Leeuw, E., Porcelli, I., Sargent, F., Palmer, T. and Berks, B.C. (2001)
FEBS Lett. 506, 143–14847 Porcelli, I., de Leeuw, E., Wallis, R., van den Brink-van der Laan, E.,
de Kruijff, B., Wallace, B.A., Palmer, T. and Berks, B.C. (2002)Biochemistry 41, 13690–13697
48 Pop, O.I., Westermann, M., Volkmer-Engert, R., Schulz, D., Lemke, C.,Schreiber, S., Gerlach, R., Wetzker, R. and Muller, J.P. (2003) J. Biol.Chem. 278, 38428–38436
49 Westermann, M., Pop, O.I., Gerlach, R., Appel, T.R., Schlormann, W.,Schreiber, S. and Muller, J.P. (2006) Biochim. Biophys. Acta 1758,443–451
50 De Keersmaeker, S., Van Mellaert, L., Schaerlaekens, K., Van Dessel,W., Vrancken, K., Lammertyn, E., Anne, J. and Geukens, N. (2005)FEBS Lett. 579, 797–802
51 De Keersmaeker, S., Van Mellaert, L., Schaerlaekens, K., Van Dessel,W., Vrancken, K., Lammertyn, E., Anne, J. and Geukens, N. (2005)FEBS Lett. 579, 797–802
52 Chan, C.S., Zlomislic, M.R., Tieleman, D.P. and Turner, R.J. (2007)Biochemistry 46, 7396–7404
53 Sipos, L. and von Heijne, G. (1993) Eur. J. Biochem. 213, 1333–134054 Wang, Y., Zhang, Y. and Ha, Y. (2006) Nature 444, 179–18055 Wu, Z., Yan, N., Feng, L., Oberstein, A., Yan, H., Baker, R.P., Gu, L.,
Jeffrey, P.D., Urban, S. and Shi, Y. (2006) Nat. Struct. Mol. Biol. 13,1084–1091
56 Sargent, F., Stanley, N.R., Berks, B.C. and Palmer, T. (1999)J. Biol. Chem. 274, 36073–36082
57 Blaudeck, N., Kreutzenbeck, P., Muller, M., Sprenger, G.A. and Freudl,R. (2005) J. Biol. Chem. 280, 3426–3432
58 Creighton, A.M., Hulford, A., Mant, A., Robinson, D. and Robinson, C.(1995) J. Biol. Chem. 270, 1663–1669
59 Clark, S.A. and Theg, S.M. (1997) Mol. Biol. Cell 8, 923–93460 Santini, C. L., Ize, B., Chanal, A., Muller, M., Giordano, G. and Wu, L.F.
(1998) EMBO J. 17, 101–11261 Musser, S.M. and Theg, S.M. (2000) Eur. J. Biochem. 167, 2588–259862 Thomas, J.D., Daniel, R.A., Errington, J. and Robinson, C. (2001)
Mol. Microbiol. 39, 47–5263 Santini, C.-L., Bernadac, A., Zhang, M., Chanal, A., Ize, B., Blanco, C.
and Wu, L.-F. (2001) J. Biol. Chem. 276, 8159–816464 DeLisa, M.P., Tullman, D. and Georgiou, G. (2003) Proc. Natl.
Acad. Sci. U.S.A. 100, 6115–612065 Hynds, P.J., Robinson, D. and Robinson, C. (1998) J. Biol. Chem. 273,
34868–3487466 Madueno, F., Napier, J.A. and Gray, J.C. (1993) Plant Cell 5, 1865–187667 Rodrigue, A., Batia, N., Muller, M., Fayet, O., Bohm, R.,
Mandrand-Berthelot, M.A. and Wu, L.F. (1996) J. Bacteriol. 178,4453–4460
68 Graubner, W., Schierhorn, A. and Bruser, T. (2007) J. Biol. Chem. 282,7116–7124
69 Perez-Rodriguez, R., Fisher, A.C., Perlmutter, J.D., Hicks, M.G.,Chanal, A., Santini, C.-L., Wu, L.-F., Palmer, T. and DeLisa, M.P. (2007)J. Mol. Biol. 367, 715–730
70 Rial, D.V. and Ceccarelli, E.A. (2002) Protein Expression Purif. 25,503–507
71 Howell, J.M., Winstone, T.L., Coorssen, J.R. and Turner, R.J. (2006)Proteomics 6, 2050–2069
72 Oresnik, I.J., Ladner, C.L. and Turner, R.J. (2001) Mol. Microbiol. 40,323–331
73 Houry, W.A. (2001) Curr. Protein Pept. Sci. 2, 227–24474 Kipping, M., Lilie, H., Lindenstrauss, U., Andreesen, J.R., Griesinger, C.,
Carlomagno, T. and Bruser, T. (2003) FEBS Lett. 550, 18–2275 San Miguel, M., Marrington, R., Rodger, P.M., Rodger, A. and
Robinson, C. (2003) Eur. J. Biochem. 270, 3345–335276 DeLisa, M.P., Samuelson, P., Palmer, T. and Georgiou, G. (2002)
J. Biol. Chem. 277, 29825–2983177 Ize, B., Gerard, F., Zhang, M., Chanal, A., Voulhoux, R., Palmer, T.,
Filloux, A. and Wu, L.-F. (2002) J. Mol. Biol. 317, 327–33578 Blaudeck, N., Kreutzenbeck, P., Freudl, R. and Sprenger, G.A. (2003)
J. Bacteriol. 185, 2811–281979 Sambasivarao, D., Turner, R.J., Simala-Grant, J.L., Shaw, G., Hu, J. and
Weiner, J.H. (2003) J. Biol. Chem. 275, 22526–2253180 Jack, R.L., Buchanan, G., Dubini, A., Hatzixanthis, K., Palmer, T. and
Sargent, F. (2004) EMBO J. 23, 3962–397281 Blaudeck, N., Sprenger, G.A., Freudl, R. and Wiegert, T. (2001)
J. Bacteriol. 183, 604–61082 Buchanan, G., Sargent, F., Berks, B.C. and Palmer, T. (2001)
Arch. Microbiol. 177, 107–11283 Czjzek, M., Dos Santos, J.-P., Pommier, J., Giordano, G., Mejean, V. and
Haser, R. (1998) J. Mol. Biol. 284, 435–44784 Schneider, F., Lowe, J., Huber, R., Schindelin, H., Kisker, C. and
Knablein, J. (1996) J. Mol. Biol. 263, 53–6985 McAlpine, A.S., McEwan, A.G. and Bailey, S. (1998) J. Mol. Biol. 275,
613–62386 Schindelin, H., Kisker, C., Hilton, J., Rajagopalan, K.V. and Rees, D.C.
(1996) Science 272, 1615–162187 Mejean, V., Iobbi-Nivol, C., Lepelletie, M., Giordano, G., Chippaux, M.
and Pascal, M.C. (1994) Mol. Microbiol. 11, 1169–117988 Kisker, C., Schindelin, H. and Rees, D.C. (1997) Annu. Rev. Biochem.
66, 233–26789 McDevitt, C.A., Hugenholtz, P., Hanson, G.R. and McEwan, A.G. (2002)
Mol. Microbiol. 44, 1575–8790 Pommier, J., Mejean, V., Giordano, G. and Iobbi-Nivol, C. (1998)
J. Biol. Chem. 273, 16615–1662091 Ilbert, M., Mejean, V., Giudici-Orticoni, M.T., Samama, J.-P. and
Iobbi-Nivol, C. (2003) J. Biol. Chem. 278, 28787–2879292 Ilbert, M., Mejean, V. and Iobbi-Nivol, C. (2004) Microbiology 150,
935–94393 Genest, O., Ilbert, M., Mejean, V. and Iobbi-Nivol, C. (2005)
J. Biol. Chem. 280, 15644–1564894 Genest, O., Seduk, F., Ilbert, M., Mejean, V. and Iobbi-Nivol, C. (2006)
Biochem. Biophys. Res. Commun. 339, 991–99595 Genest, O., Seduk, F., Theraulaz, L., Mejean, V. and Iobbi-Nivol, C.
(2006) FEMS Microbiol. Lett. 265, 51–5596 Hatzixanthis, K., Clarke, T.A., Oubrie, A., Richardson, D.J., Turner, R.J.
and Sargent, F. (2005) Proc. Natl. Acad. Sci. 102, 8460–846597 Li, S.Y., Chang, B.Y. and Lin, S.C. (2006) J. Biotechnol. 122, 412–42198 Sargent, F., Ballantine, S.P., Rugman, P.A., Palmer, T. and Boxer, D.H.
(1998) Eur. J. Biochem. 255, 746–75499 Rodrigue, A., Chanal, A., Beck, K., Muller, M. and Wu, L.-F. (1999)
J. Biol. Chem. 274, 13223–13228100 Butland, G., Zhang, J.W., Yang, W., Sheung, A., Wong, P., Greenblatt,
J.F., Emili, A. and Zamble, D.B. (2006) FEBS Lett. 580, 677–681101 Tranier, S., Iobbi-Nivol, C., Birck, C., Ilbert, M., Mortier-Barriere, I.,
Mejean, V. and Samama, J.-P. (2003) Structure 11, 165–174102 Turner, R.J., Papish, A.L. and Sargent, F. (2004) Can. J. Microbiol. 50,
225–238103 Jack, R.L., Dubini, A., Palmer, T. and Sargent, F. (2005)
Biochem. Soc. Trans. 33, 105–107104 Wu, L.-F., Chanal, A. and Rodrigue, A. (2000) Arch. Microbiol. 173,
319–324105 Sargent, F., Berks, B.C. and Palmer, T. (2002) Arch. Microbiol. 178,
77–84106 Dubini, A. and Sargent, F. (2003) FEBS Lett. 549, 141–146
C©The Authors Journal compilation C©2007 Biochemical Society

The twin-arginine transport system: moving folded proteins across membranes 847
107 Broughton, M.J., Howe, C.J. and Hiller, R.G. (2006) Gene 369, 72–79108 Kirillova, O., Chruszcz, M., Shumilin, I.A., Skarina, T.,
Gorodichtchenskaia, E., Cymborowski, M., Savchenko, A., Edwards, A.and Minor, W. (2007) Acta Crystallogr. Sect. D Biol. Crystallogr. 63,348–354
109 Dridge, E.J., Richardson, D.J., Lewis, R.J. and Butler, C.S. (2006)Biochem. Soc. Trans. 34, 118–121
110 Sargent, F. (2007) Microbiology 153, 633–651111 Richardson, D.J., Berks, B.C., Russell, D.A., Spiro, S. and Taylor, C.J.
(2001) Cell. Mol. Life Sci. 58, 165–178112 Bilous, P.T., Cole, S.T., Anderson, W.F. and Weiner, J.H. (1988)
Mol. Microbiol. 2, 785–795113 Stanley, N.R., Sargent, F., Buchanan, G., Shi, J., Stewart, V., Palmer, T.
and Berks, B.C. (2002) Mol. Microbiol. 43, 1005–1021114 Simala-Grant, J.L. and Weiner, J.H. (1996) Microbiology 142,
3231–3239115 Winstone, T.L., Workentine, M.L., Sarfo, K.J., Binding, A.J., Haslam, B.D.
and Turner, R.J. (2006) Arch. Biochem. Biophys. 455, 89–97116 Grove, J., Tanapongpipat, S., Thomas, G., Griffiths, L., Crooke, H. and
Cole, J. (1996) Mol. Microbiol. 19, 467–481117 Jepson, B.J., Mohan, S., Clarke, T.A., Gates, A.J., Cole, J.A., Butler, C.S.,
Butt, J.N., Hemmings, A.M. and Richardson, D.J. (2007) J. Biol. Chem.282, 425–437
118 Thomas, G., Potter, L. and Cole, J.A. (1999) FEMS Microbiol. Lett. 174,167–171
119 Brondijk, T.H., Fiegen, D., Richardson, D.J. and Cole, J.A. (2002)Mol. Microbiol. 44, 245–255
120 Potter, L.C. and Cole, J.A. (1999) Biochem. J. 344, 69–76121 Nilavongse, A., Brondijk, T.H., Overton, T.W., Richardson, D.J., Leach,
E.R. and Cole, J.A. (2006) Microbiology 152, 3227–3237122 Jormakka, M., Richardson, D.J., Byrne, B. and Iwata, S. (2004)
Structure 12, 95–104123 Maillard, J., Spronk, C.A.E.M., Buchanan, G., Lyall, V., Richardson, D.J.,
Palmer, T., Vuister, G.W. and Sargent, F. (2007) Proc. Natl. Acad.Sci. U.S.A. 104, 15641–15646
124 Reyes, F., Gavira, M., Castillo, F. and Moreno-Vivian, C. (1998)Biochem. J. 331, 897–904
125 Russell, R.B., Sasieni, P.D. and Sternberg, M.J. (1998) J. Mol. Biol. 282,903–918
126 Mandrand-Berthelot, M.-A., Couchoux-Luthaud, G., Santini, C.-L. andGiordano, G. (1988) J. Gen. Microbiol. 134, 3129–3139
127 Chen, L.Y., Leu, W.M., Wang, K.T. and Lee, Y.H. (1992) J. Biol. Chem.267, 20100–20107
128 Dreusch, A., Burgisser, D.M., Heizmann, C.W. and Zumft, W.G. (1997)Biochim. Biophys. Acta 1319, 311–318
129 McDonough, J.A., Hacker, K.E., Flores, A.R., Pavelka, Jr, M.S. andBraunstein, M. (2005) J. Bacteriol. 187, 7667–7679
130 Voulhoux, R., Ball, G., Ize, B., Vasil, M.L., Lazdunski, A., Wu, L.-F. andFilloux, A. (2001) EMBO J. 20, 6735–6741
131 Gralnick, J.A., Vali, H., Lies, D.P. and Newman, D.K. (2006) Proc. Natl.Acad. Sci. U.S.A. 103, 4669–4674
132 Kurisu, G., Zhang, H., Smith, J.L. and Cramer, W.A. (2003) Science 302,1009–1014
133 Bachmann, J., Bauer, B., Zwicker, K., Ludwig, B. and Anderka, O.(2006) FEBS J. 273, 4817–4830
134 De Buck, E., Vranckx, L., Meyen, E., Maes, L., Vandersmissen, L.,Anne, J. and Lammertyn, E. (2007) FEBS Lett. 581, 259–264
135 Martinez-Espinosa, R.M., Bonete, M.J., Dridge, E.J., Butler, C.S.,Sargent, F. and Richardson, D.J. (2007) FEMS Microbiol. Lett.,doi:10.1111/j.1574-6968.2007.00887.x
136 Sargent, F., Berks, B.C. and Palmer, T. (2006) FEMS Microbiol. Lett.254, 198–207
137 Kirwin, P.M., Elderfield, P.D., Williams, R.S. and Robinson, C. (1988)J. Biol. Chem. 263, 18128–18132
138 Paetzel, M., Dalbey, R.E. and Strynadka, N.C. (1998) Nature 396,186–190
139 Di Cola, A. and Robinson, C. (2005) J. Cell. Biol. 171, 281–289140 Hatzixanthis, K., Palmer, T. and Sargent, F. (2003) Mol. Microbiol. 49,
1377–1390141 Summer, E.J., Hori, H., Settles, M.A. and Cline, K. (2000) J. Biol. Chem.
275, 23483–23490142 Finlay, B.B. and Falkow, S. (1997) Microbiol. Mol. Biol. Rev. 61,
136–169143 Ding, Z. and Christie, P.J. (2003) J. Bacteriol. 185, 760–771144 Caldelari, I., Mann, S., Crooks, C. and Palmer, T. (2006) Mol. Plant
Microbe Interact. 19, 200–212145 Bronstein, P.A., Marrichi, M., Cartinhour, S., Schneider, D.J. and DeLisa,
M.P. (2005) J. Bacteriol. 187, 8450–8461146 Gonzalez, E.T., Brown, D.G., Swanson, J.K. and Allen, C. (2007)
Appl. Environ. Microbiol. 73, 3779–3786147 Pradel, N., Ye, C., Livrelli, V., Xu, J., Joly, B. and Wu, L.F. (2003)
Infect. Immun. 71, 4908–4916148 Ochsner, U.A., Snyder, A., Vasil, A.I. and Vasil, M.L. (2002) Proc. Natl.
Acad. Sci. U.S.A. 99, 8312–8317149 Lavander, M., Ericsson, S.K., Broms, J.E. and Forsberg, A. (2006)
Infect. Immun. 74, 1768–1776150 Rossier, O. and Cianciotto, N.P. (2005) Infect. Immun. 73,
2020–2032151 De Buck, E., Maes, L., Meyen, E., Van Mellaert, L., Geukens, N., Anne,
J. and Lammertyn, E. (2005) Biochem. Biophys. Res. Commun. 331,1413–1420
152 Saint-Joanis, B., Demangel, C., Jackson, M., Brodin, P., Marsollier, L.,Boshoff, H. and Cole, S.T. (2006) J. Bacteriol. 188, 6669–6679
153 Stanley, N.R., Findlay, K., Berks, B.C. and Palmer, T. (2001) J. Bacteriol.183, 139–144
154 Ize, B., Stanley, N.R., Buchanan, G. and Palmer, T. (2003)Mol. Microbiol. 48, 1183–1193
155 Olson, J.W. and Maier, R.J. (2002) Science 298, 1788–1790156 Theg, S.M., Cline, K., Finazzi, G. and Wollman, F.A. (2005)
Trends Plant. Sci. 10, 153–154157 Zasloff, M. (2002) Nature 415, 389–395158 Bruser, T. (2007) Appl. Microbiol. Biotechnol. 76, 35–45159 Fisher, A.C., Kim, W. and DeLisa, M.P. (2006) Protein Sci. 15,
449–458160 Strauch, E.M. and Georgiou, G. (2007) Protein Sci. 16, 1001–1008161 Ribnicky, B., Van Blarcom, T. and Georgiou, G. (2007) J. Mol. Biol. 369,
631–639162 Paschke, M. and Hohne, W. (2005) Gene 350, 79–88163 Droge, M.J., Boersma, Y.L., Braun, P.G., Buining, R.J., Julsing, M.K.,
Selles, K.G., van Dijl, J.M. and Quax, W.J. (2006) Appl. Environ.Microbiol. 72, 4589–4595
164 Bogsch, E., Brink, S. and Robinson, C. (1997) EMBO J. 16, 3851–3859165 Dilks, K., Gimenez, M.I. and Pohlschroder, M. (2005) J. Bacteriol. 187,
8104–8113166 Gouffi, K., Gerard, F., Santini, C.L. and Wu, L.F. (2004) J. Biol. Chem.
279, 11608–11615
Received 14 June 2007doi:10.1042/BST0350835
C©The Authors Journal compilation C©2007 Biochemical Society