Embed Size (px)
Citation preview

International Journal of Food Microbiology 133 (2009) 148–153
Contents lists available at ScienceDirect
International Journal of Food Microbiology
j ourna l homepage: www.e lsev ie r.com/ locate / i j foodmicro
Coculture of Bifidobacterium longum and Bifidobacterium breve alters their proteinexpression profiles and enzymatic activities
Lorena Ruiz a, Borja Sánchez b,⁎, Clara G. de los Reyes-Gavilán a, Miguel Gueimonde a, Abelardo Margolles a
a Instituto de Productos Lácteos de Asturias, Consejo Superior de Investigaciones Científicas (IPLA-CSIC). Ctra. Infiesto s/n, 33300, Villaviciosa, Asturias, Spainb UMR 5248 Chimie et Biologie des membranes et des nanoobjets, Laboratoire de Microbiologie et Biochimie Appliquée, ENITA-Université de Bordeaux 2, 1 cours du Général de Gaulle,33175 Gradignan, France
⁎ Corresponding author. Laboratoire de MicrobioloENITAB. 1, cours du Général de Gaulle, 33175 Gradigna92; fax: +33 557 35 07 39.
E-mail address: [email protected] (B. Sánchez).
0168-1605/$ – see front matter © 2009 Elsevier B.V. Adoi:10.1016/j.ijfoodmicro.2009.05.014
a b s t r a c t
a r t i c l e i n f oArticle history:Received 30 November 2008Received in revised form 8 May 2009Accepted 11 May 2009
Keywords:CocultureBifidobacteriaProteomics
Some strains of the genus Bifidobacterium are probiotic bacteria commonly added to functional dairyproducts. The influence of coculturing Bifidobacterium longum NCIMB8809 and Bifidobacterium breveNCIMB8807 on their physiology was studied. 2DE separation of protein extracts, coupled to MS proteinanalysis allowed the identification of 16 proteins whose expression drastically changed when cells weregrown in compartmentalized coculture, compared to monoculture. These included ribosomal proteins andproteins involved in carbohydrate metabolism, gene regulation, cell envelope biogenesis and transportprocesses. Significant changes in some glycoside-hydrolysing activities (β-D-xylopyranosidase, α-L-arabinofuranosidase and β-D-glucopyranosidase) were also detected. Furthermore, qRT-PCR experimentsusing as targets the B. breve genes clgR (transcriptional regulator) clpP1, clpP2 and clpC (chaperone- andprotease-encoding genes positively regulated by clgR) supported the proteomic results, the four genesdisplaying a higher expression level in coculture. This study provides new insights to understand thecommunication among Bifidobacterium species.
© 2009 Elsevier B.V. All rights reserved.
1. Introduction
Bacteria release a wide range of compounds in the externalenvironment in order to communicate and coordinate their activities.One of the first signalling mechanisms was described as “autoinduc-tion”, in which bacterial communication involves producing, releas-ing, detecting and responding to small signalling hormone-likemolecules, called autoinducers (Visick and Fuqua, 2005; Waters andBassler, 2005). In Gram positive bacteria this communication isusually mediated by three-component systems (Hellingwerf et al.,1998; Nakayama et al., 2001; Gobbetti et al., 2007), which control alarge variety of phenotypes, such as bacteriocin synthesis in lactic acidbacteria (Brurberg et al., 1997), expression of virulence factors instaphylococci (Novick et al., 1995), biofilm formation, and stressresponses (Hoch, 2000), among others. However, some exceptions tothis general system have also been reported, in which γ-butyrolac-tones, similar to Gram negative acyl-homoserine lactones, appear tobe involved in antibiotic production within the genus Streptomyces(Takano et al., 2001).
Bifidobacteria are among the first species to colonise the humangastrointestinal tract, where they are thought to exert health-
gie et Biochimie Appliquée,n, France. Tel.: +33 557 35 59
ll rights reserved.
promoting activities, the most prevalent of which are the modulationof the immune response and the maintenance of the intestinal barrierintegrity (Leahy et al., 2005). Some strains of Bifidobacterium, mainlyfrom the species Bifidobacterium longum, Bifidobacterium breve, Bifi-dobacterium animalis, and Bifidobacterium bifidum, are attractiveprobiotics, and they are normally consumed as food supplements oradjunct cultures in fermented dairy products, in which several Bifi-dobacterium species can be found (Brigidi et al., 2003; Gueimondeet al., 2004a; Masco et al., 2005). However, the factors involved in thebifidobacterial communication networks are completely unknown.Only a recent report has suggested that the phosphorylatedautoinducer-2 production protein LuxS of B. longum NCC2705 mayplay a key role in the regulation of quorum sensing, but there is nophysiological data supporting this hypothesis (Yuan et al., 2008).However, some methods have recently been developed in order tostudy the influence of coculturing different bacteria in theirphysiological state. Cocultivation of different sourdough lactobacillispecies (Lactobacillus sanfranciscensis with Lactobacillus plantarum orLactobacillus brevis) into double culture vessels separated by amembrane filter allowed the identification of new signalling mole-cules using a proteomic approach, including proteins involved in theLuxS autoinduction system, with furanones being considered aspossible signalling molecules (Di Cagno et al., 2007). Also, cocultureof Escherichia coli and Vibrio harveyi showed that specific autoinducerproteins can mediate two-way communication between differentbacterial species (Xavier and Bassler, 2005). Our previous studies have
149L. Ruiz et al. / International Journal of Food Microbiology 133 (2009) 148–153
shown that B. longum NCIMB8809 is able to secrete proteins into thesupernatant with potential intercellular signalling activity (Sánchezet al., 2008). In this respect, we have described in this article for thefirst time a method for the cocultivation of two different Bifidobac-terium species, B. longum and B. breve, which allows diffusion ofmetabolic products, but not cells, to occur, and have examined itsusefulness for the study of the changes in their proteomic profiles andmetabolic activities.
2. Materials and methods
2.1. Microorganims and growth conditions
B. longum NCIMB8809 and B. breve NCIMB8807 (National Collec-tions of Industrial and Marine Bacteria, Aberdeen, Scotland, UK) wereroutinely grown in deMan–Rogosa–Sharpe broth (deMan et al., 1960)
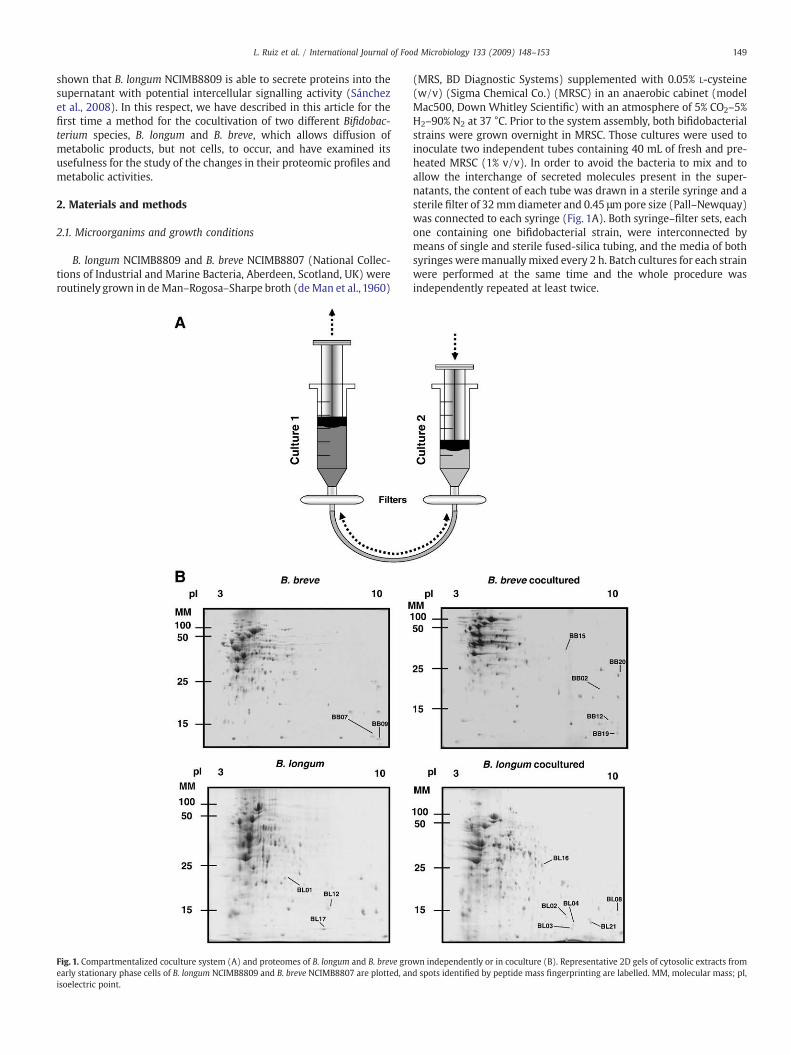
Fig. 1. Compartmentalized coculture system (A) and proteomes of B. longum and B. breve groearly stationary phase cells of B. longum NCIMB8809 and B. breve NCIMB8807 are plotted, anisoelectric point.
(MRS, BD Diagnostic Systems) supplemented with 0.05% L-cysteine(w/v) (Sigma Chemical Co.) (MRSC) in an anaerobic cabinet (modelMac500, Down Whitley Scientific) with an atmosphere of 5% CO2–5%H2–90% N2 at 37 °C. Prior to the system assembly, both bifidobacterialstrains were grown overnight in MRSC. Those cultures were used toinoculate two independent tubes containing 40 mL of fresh and pre-heated MRSC (1% v/v). In order to avoid the bacteria to mix and toallow the interchange of secreted molecules present in the super-natants, the content of each tube was drawn in a sterile syringe and asterile filter of 32mmdiameter and 0.45 μmpore size (Pall–Newquay)was connected to each syringe (Fig. 1A). Both syringe–filter sets, eachone containing one bifidobacterial strain, were interconnected bymeans of single and sterile fused-silica tubing, and the media of bothsyringes weremanually mixed every 2 h. Batch cultures for each strainwere performed at the same time and the whole procedure wasindependently repeated at least twice.
wn independently or in coculture (B). Representative 2D gels of cytosolic extracts fromd spots identified by peptide mass fingerprinting are labelled. MM, molecular mass; pI,

150 L. Ruiz et al. / International Journal of Food Microbiology 133 (2009) 148–153
The maximum specific growth rates were estimated from thegrowth curve according to the method of Baranyi and Roberts (1994)using the software MicroFit 1.0 (http://www.ifr.ac.uk/microfit/). Thegrowth rate was determined for the interconnected cultures as well asfor the individual, unconnected batch cultures (mean±standarddeviations).
2.2. Two-dimensional gel electrophoresis, statistical analysis, and proteinidentification
Samples for 2DE were taken at 11–13 h after inoculation, whichcorresponded in all cases with the early stationary phase of growth(data not shown). Cell-free extracts and 2DE were obtained andperformed, respectively, as previously described (Sánchez et al., 2005)with minor modifications. Briefly, 500 μg of protein from bifidobacter-ial extracts were loaded onto a strip with a non-linear immobilized pHrange of 3 to 10 (GEHealthcare) and focused at 70,000 V/h. Separationin the second dimensionwas carried out in 12.5% polyacrylamide-SDSgels and protein spots were visualized with Bio-Safe Coomassiestaining (Bio-Rad). Proteins were identified by peptide mass finger-printing after trypsinolysis and matrix-assisted laser desorptionionization/time of flight/mass spectrometry (MALDI-TOF-MS) analy-sis at the Proteomics Unit of the Centro de Investigaciones Cardio-vasculares (Madrid, Spain). Protein identification was achieved bycombining MS data through BioTools software (Bruker-Daltonics) tosearch a non-redundant protein database (NR; 4.106 entries; NationalCenter for Biotechnology Information) using the Mascot software(Matrix Science). Spot detection and volume quantification wereperformed with ImageMaster Platinum software (version 5.00, GEHealthcare). All the growth experiments were carried out in duplicate(biological replicates) and at least three independent gels (technicalreplicates) were performed for each extract.
2.3. Real-Time PCR analysis
Lysis, total RNA extraction, and quantitative PCR analysis were carriedout as previously described (Gueimonde et al., 2007), with an ABI Prism7500 Fast Real-Time PCR System (Applied Biosystems). Primers targetingthe B. breve genes clgR, clpC, clpP1 and clpP2 were designed and theirspecificity was tested before the quantitative analysis. The specificoligonucleotides ClgRR (GGCGACACCCGGCTGTA) and ClgRF (CGAGT-GAACGAACAAATGGAAGT) (amplicon of 63 bp in clgR; accession numberAY837843), ClpCR (CGCATGAAGGATCAGGTGAGTT) and ClpCF(TCGTCCAAACGGTTGAGGAA) (amplicon of 71 bp in clpC, accessionnumber AY722390), ClpP1R (GGCTTCCTGAGCGGTGAAC) and ClpP1F(GCCATACCGTCGAGGAAATCT) (amplicon of 65 bp in clpP1; accessionnumber AY955251) and ClpP2R (GGTTTCCTCCAGCCATTCG) and ClpP2F(CAAGGCCACGGAGATTGAAAT) (amplicon of 67 bp in clpP2; accessionnumber AY955251) were used. Expression levels were determined byrelative quantification using the ΔΔCt method in which the expressionlevel in the control culture (independent culture) is arbitrarily set to 1 andthe expression levels in the samples from the coculture are calculatedrelative to that control. The 16S rRNA was used as endogenous controlusing previously described primers (Gueimonde et al., 2004b). Threeindependent cultures (and cocultures) were analysed, and for eachculture two separate PCR reactions were performed.
2.4. Determination of enzymatic activities
Glycosidase activities were measured spectrophotometrically oncell free extracts from the release of p-Nitrophenol (pNP) produced bythe enzymatic hydrolysis of the corresponding pNP-glycoside sub-strates (Sigma), as previously described (Noriega et al., 2004). Also,fructose-6-phosphate phosphoketolase (F6PPK) activity was mea-sured according to Sánchez et al. (2004) as the ferric acetylhydroxamate produced from the enzymatically generated acetyl
phosphate. One unit of enzymatic activity was defined as the amountof protein that releases 1 nmol of pNP or acetyl phosphate per minute.Specific activities were measured in duplicate for each culture, andexpressed as units per mg protein.
3. Results and discussion
Only a few previous reports have dealt with the communicationmechanisms among probiotic bacteria, but the input and advantagesof high throughput genomic and proteomic techniques currentlymake possible to undertake this kind of studies. Thus the influence ofcompartmentalized cocultivation on the physiology of different Bifi-dobacterium species was investigated in this work.
B. longum NCIMB8809 and B. breve NCIMB8807, when grownindependently, consistently reached the maximum OD600 afterapproximately 12 h of incubation. For all the experiments performedin this paper cells were collected in early stationary phase afterapproximately 11–13 h of incubation (OD600 3.63±0.25). Under theseconditions, the final pH of the cultures was between 4.7 and 4.8. MICdeterminations of both strains showed that this pH was slightlyinhibitory for the growth of B. longum and B. breve, but both strainswere able to grow in MRSC at pH 4.8 (data not shown). Thus, all theexperiments were carried out in unbuffered medium in order tomimic the in vivo situation that these bacteria can find, for example, infermented dairy products or in the gastrointestinal ecosystem.
The values of μmax were 1.26±0.03 for B. longum and 1.98±0.49for B. breve. However, a decrease in the μmax was observed for B. brevewhen it grew in compartmentalized coculture with B. longum (1.14±0.30). This means that specific growth rate was 42.43% lower for B.brevewhen cocultured with B. longum. Remarkably, such an inhibitionwas not observed for the B. longum strain. It is possible that someinhibitory substances produced by B. longum caused the observedeffect. In fact, a previous report (O'Riordan and Fitzgerald, 1998)showed that B. longum NCIMB8809 was able to produce antimicrobialcompounds that can inhibit the growth of several Gram positivebacteria. However, this inhibitionmay be just a limitation derived fromthe experimental design, and caused by the fact that B. longum simplyoutcompetes B. breve by consuming C- and N-sources. In this way, careshould be taken in the interpretation of these results, as the simplicityof the experimental devicemaycombine the effects caused by nutrientstarvation, production of antimicrobials, etc, on specific growth rates.
In order to analyse possible variations in the level of proteinexpression of Bifidobacterium cells grown in coculture, with respect tomonoculture, a two-dimensional electrophoresis (2DE) analysis wascarried out. To determine the effect of coculturing in a reliable way wefocussed our analysis only in those spots showing a very drastic andreproducible change in their intensity (appearance/disappearance ofnew or existing spots in at least three independent gels). Thus, 16spots (Fig. 1B) were selected and the corresponding proteins wereidentified (Table 1). Among the identified proteins, 10 were ribosomalproteins, 5 in B. longum (spots BL08, BL12, BL16, BL17, and BL21) and 5in B. breve (BB07, BB09, BB12, BB19, and BB20). The general tendencyof ribosomal proteins was a marked up-regulation, although 50Sribosomal subunit protein L17 (RplQ) in B. longum and 50S ribosomalsubunit protein L11 (RplK) and 30S ribosomal subunit protein S8(RpsH) in B. breve showed a marked reduction in concentration in theextracts of the cells grown in coculture. Ribosomal proteins arenecessary for ribosome assembly and stability and in certain bacteriathey have been implicated in sensing environmental changes (Chenet al., 2003; Van Bogelen and Neidhardt, 1990). Some of them, such asRplK, are able to detect the presence of nutrient starvation conditions,modulating the synthesis of metabolic enzymes and the cell fitness(Wilson and Nierhaus, 2005).
ClgR, a transcriptional regulator involved in regulation of the clpCgene and the clpP operon of B. breve (De Dea Lindner et al., 2007), wasstrongly up-regulated. ClpC belongs to the stress response-related Clp

Table 1Proteins whose production was affected by the compartmentalized cocultivation in B. breve and B. longum.
COGa Spot no.b Putative functionc Gene Accesion no. Massd pIe Matched peptidesf Coverageg Prod.h
Translation BL12 50S Rpi L17 rplQ BL1607 19.109 6.88 8 33 ↓BL08 30S Rp S9 rpsl BL1572 17.596 9.86 4 20 ↑BL21 Ribosome-associated protein Y 19.712 6.86 4 21 ↑BL16 hypothetical protein in sigma 54 modulation protein/S30ea Rp family BL1418 24.668 5.85 10 35 ↑BL17 30S Rp S16 rpsP BL0305 16.497 6.43 16 79 ↑BB07 50S Rp L11 rplK BL1290 15.291 9.21 10 62 ↓BB09 30S Rp S8 rpsH BL1594 14.260 9.55 8 45 ↓BB12 30S Rp S13 rpsM BL1604 15.549 10.37 9 47 ↑BB19 50S Rp L13 rplM BL1571 16.613 9.99 7 40 ↑BB20 50S Rp L3 rplC BL1578 22.644 9.79 9 32 ↑
Transport BL01 Probable ATP binding protein of ABC transporter BL0870 28.189 5.54 9 37 ↓Carbohydratemetabolism
BL02 Phosphoketolase xfp BL0959 92.683 5.06 9 8 ↑BL03 Phosphoketolase xfp BL0959 92.683 5.06 10 10 ↑BL04 Phosphoketolase xfp BL0959 92.683 5.06 7 6 ↑
Transcription BB02 Transcriptional regulator clgR AAX52924 18.804 9.34 7 34 ↑Cell envelopebiogenesis
BB15 UDP-N-acetylglucosamine: LPS N-acetylglucosamine transferase murG BL1323 41.600 6.47 4 7 ↑
a COG, cluster of orthologous genes.b Spot numbers refer to the proteins labeled in Fig. 1.c Putative functions were established according to the cluster of orthologous genes for B. longum NCC2705 (Schell et al., 2002).d Theoretical molecular mass expressed in kDa.e Theoretical isoelectric point expressed in pH.f Number of tryptic peptides observed contributing to the percentage of amino acid coverage.g Percentage of sequence coverage.h Prod., production. Up-regulation (↑) or down-regulation (↓) of matching proteins for B. longum (BL) and B. breve (BB) when grown in coculture.i Rp, ribosomal protein.
151L. Ruiz et al. / International Journal of Food Microbiology 133 (2009) 148–153
ATPase family, whose members act as chaperones and regulators ofproteolysis (Ventura et al., 2005a). On the other hand, the clpP operonis constituted by clpP1 and clpP2, paralogous genes that form abicistronic transcriptional unit, whose deduced protein productsdisplay significant homology to characterised ClpP peptidases (Ven-tura et al., 2005b). The three genes (clpC, clpP1 and clpP2) have beenshown to be involved in the response of B. breve to some environ-mental stimuli, such as moderate heat (De Dea Lindner et al., 2007).We demonstrated, using quantitative Real-Time PCR analysis, that thedifferent concentration of B. breve ClgR was due to an induction of thegene. The clgR transcript was about 15 times more abundant incoculture, compared to monoculture (15.15±5.31). Furthermore, wealso found a clear induction of the genes clpP1 (4.53±0.40), clpP2 andclpC, although for clpP2 and clpC a quantitative determination of theinduction could not be performed since the transcripts of the geneswere only found in coculture, while in monoculture they were alwaysunder the detection limit (data not shown). These results suggest anactivation of a stress-related response in B. brevewhen grown togetherwith B. longum. Interestingly, several ribosomal proteins have beenshown to be substrates of Clp proteases in bacteria (Neher et al., 2006),and this fact could be related to the mismatch of the theoreticalmolecularmass (Table 1) and the experimentalmolecularmass (Fig.1)found for some ribosomal proteins in this study, such as BB19.
Three different proteolytic products of the enzyme Xfp (Fructose6-phosphate phosphoketolase; BL02, BL03, and BL04) showed up in B.longum when grown together with B. breve. Xfp is the key enzyme ofthe glucose catabolism in Bifidobacterium (Meile et al., 2001). A recentreport indicated that 21 different spots of Xfp differing in charge andmass were identified in B. longum NCC2705 (Yuan et al., 2006),prompting the authors to suggest that the different Xfp variants couldbe the consequence of posttranslational modifications which mayinfluence the properties and the function of the enzyme. Furthermore,several spots corresponding to the isoforms of this protein werepreviously detected as a response to bile or low pH in Bifidobacterium(Sánchez et al., 2007a,b), suggesting that environmental conditionscould influence sugar metabolism through Xfp posttranslationalmodifications. These results appear to indicate that the strains senseeach other rearranging their carbohydrate metabolism in a way thatmay increase their ability to compete for sugars.
On the other hand MurG, an essential bacterial glycosyltransferaseinvolved in the biosynthesis of peptidoglycan and cell division(Mohammadi et al., 2007), was drastically up-regulated in B. brevecocultures, which could indicate that B. breve enhances its cell wallbiosynthesis as a response to the inhibition effect exerted by B. longum.In relation to this, the cell wall degrading activity N-acetil-β-D-glucosaminidase was increased in B. longumwhen grown in coculture(Table 2), suggesting an activation of autolysis mechanisms in thepresence of B. breve. The autolytic activity of this kind of enzymes hasalready been reported for other Gram positive bacteria (Buist et al.,1995; Harty et al., 2004). In this respect, the up-regulation of murG inB. breve could be also directed to counteract the harmful effect causedby the increased N-acetil-β-D-glucosaminidase activity of B. longum.
One of the most common results of coculturing bacteria is ashift in the enzymatic activity profiles (Di Cagno et al., 2007). Also,previous studies showed that shifts in the environmental condi-tions significantly influence F6PPK activity and sugar metabolism inB. longum (Sánchez et al., 2005). Therefore, in order to evaluate theinfluence of secreted molecules on cell metabolism, and to supportthe proteomic changes with physiological data, glycosidase activ-ities were measured. No statistically significant changes weredetected for the F6PPK activity, and most of the activities testeddid not significantly change when the strains were grown incompartmentalized coculture (Table 2). However, it is remarkablethat some activities responsible for the degradation of complexcarbohydrates (β-D-xylopyranosidase, α-L-arabinofuranosidase andβ-D-glucopyranosidase) were increased in B. longum, B. breve orboth, which could be the consequence of the different substratespecificities of the 2 species and a direct nutrient competition inthe medium. Interestingly, some of these enzymatic activities, suchas α-L-arabinofuranosidase, have been found to be inducible bysome carbohydrates in Bifidobacterium (Gueimonde et al., 2007). Inthis respect, a whole genome transcriptional profiling of B. longumand Bacteroides thetaiotaomicron in germ-free mice revealed thatthe presence of B. longum elicits an expansion in the diversity ofpolysaccharides targeted for degradation by B. thetaiotaomicron(Sonnenburg et al., 2006).
In summary, thiswork shows a rapid, suitable, and easy-to-performmethod for any coculture sharing similar growth parameters, and this

Table 2Enzymatic activities of cell-free extracts of B. breve and B. longum grown independentlyor in compartmentalized coculture.
pNP substrates Condition Specific activity (mean±SD)
B. longum B. breve
α-D-glucopyranoside Independent 31.30±1.21 29.46±8.73Coculture 36.68±3.40⁎ 40.90±14.58
β-D-galactopyranoside Independent 150.79±6.77 125.96±47.12Coculture 171.22±2.34⁎⁎ 88.18±25.58
β-D-glucopyranoside Independent 0.12±0.02 3.06±1.40Coculture 0.13±0.15 7.61±2.15⁎⁎
α-L-arabinofuranoside Independent 6.07±1.55 1.61±1.09Coculture 6.61±1.26 4.52±3.25⁎⁎
α-L-rhamnopyranoside Independent 0.21±0.17 0.77±0.46Coculture 0.11±0.03 1.63±1.23
N-acetil-β-D-glucosamine Independent 2.72±0.28 19.71±12.74Coculture 3.64±0.31⁎⁎ 20.05±11.19
β-D-fucopyranoside Independent 5.76±1.50 23.67±11.02Coculture 6.09±0.81 24.83±6.72
β-D-glucuronide Independent 0.29±0.23 0.21±0.07Coculture 0.67±0.72 0.35±0.15
α-D-galactopyranoside Independent 50.65±13.72 65.99±42.82Coculture 45.01±3.74 74.30±30.77
β-D-xylopyranoside Independent 0.51±0.16 0.34±0.14Coculture 0.97±0.20⁎ 0.78±0.32⁎
F6PPK activity Independent 1,955.57±161.17 2,117.66±187.45Coculture 2,202.26±248.48 2,275.26±308.68
A one-way ANOVA analysis was conducted to compare the independent and cocultureconditions. Results are the mean of two independent experiments, and activitymeasurements were made in duplicate.⁎ pb0.05.⁎⁎ pb0.01.
152 L. Ruiz et al. / International Journal of Food Microbiology 133 (2009) 148–153
model could easily be adapted for other bacterial species or generaProteomic changes, enzymatic activities and the induction of fourgenes were determined in this report but other studies, such as high-throughput transcriptomics, metabolomics, and more specific mole-cular biology techniques could also be performed without the need ofexpensive and time-consuming cell separation techniques. In fact, wewere able to detect a remarkable effect of coculturing in the productionof ribosomal proteins, which could reflect a deep impact in theregulatory pathways controlling translation in response to externalfactors. Proteins involved in carbohydrate catabolism, transport,transcription and cell envelope biosynthesis were also affected. Thisin vitro study could also be a complement for more complex coculturestudies in vivo.
Acknowledgements
This work was financed by the Spanish Plan Nacional de I+D(AGL2007-61805). Lorena Ruiz was supported by an I3P predoctoralfellowship granted by CSIC and FEDER funds, Borja Sánchez was therecipient of a Clarín postdoctoral contract from the Principado deAsturias Government, andMiguel Gueimondewas funded by a Ramóny Cajal postdoctoral contract from the Spanish Ministry of Educationand Science.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.ijfoodmicro.2009.05.014.
References
Baranyi, J., Roberts, T.A., 1994. A dynamic approach to predicting bacterial growth infood. International Journal of Food Microbiology 23, 277–294.
Brigidi, P., Swennen, E., Vitali, B., Rossi, M., Matteuzzi, D., 2003. PCR detection of Bifi-dobacterium strains and Streptococcus thermophilus in feces of human subjects afteroral bacteriotherapy and yogurt consumption. International Journal of FoodMicrobiology 81, 203–209.
Brurberg, M.B., Nes, I.F., Eijsink, V.G., 1997. Pheromone-induced production ofantimicrobial peptides in Lactobacillus. Molecular Microbiology 26, 347–360.
Buist, G., Kok, J., Leenhouts, K.J., Dabrowska, M., Venema, G., Haandrikman, A.J., 1995.Molecular cloning and nucleotide sequence of the gene encoding the majorpeptidoglycan hydrolase of Lactococcus lactis, a muramidase needed for cellseparation. Journal of Bacteriology 177, 1554–1563.
Chen, H., Teplitski, M., Robinson, J.B., Rolfe, B.G., Bauer, W.D., 2003. Proteomic analysis ofwild-type Sinorhizobium meliloti responses to N-acyl homoserine lactone quorum-sensing signals and the transition to stationary phase. Journal of Bacteriology 185,5029–5036.
De Dea Lindner, J., Canchaya, C., Zhang, Z., Neviani, E., Fitzgerald, G.F., van Sinderen, D.,Ventura, M., 2007. Exploiting Bifidobacterium genomes: the molecular basis ofstress response. International Journal of Food Microbiology 120, 13–24.
de Man, J., Rogosa, M., Sharpe, M.E., 1960. A medium for the culture of lactobacilli.Journal of Applied Bacteriology 23, 130–135.
Di Cagno, R., De Angelis, M., Limitone, A., Minervini, F., Simonetti, M.C., Buchin, S.,Gobbetti, M., 2007. Cell–cell communication in sourdough lactic acid bacteria: aproteomic study in Lactobacillus sanfranciscensis CB1. Proteomics 7, 2430–2446.
Gobbetti, M., De Angelis, M., Di Cagno, R., Minervini, F., Limitone, A., 2007. Cell–cellcommunication in food related bacteria. International Journal of FoodMicrobiology120, 34–45.
Gueimonde, M., Delgado, S., Mayo, B., Ruas-Madiedo, P., Margolles, A., de los Reyes-Gavilán, C.G., 2004a. Viability and diversity of probiotic Lactobacillus and Bifido-bacterium included in commercial fermentedmilks. Food Research International 37,839–850.
Gueimonde, M., Tölkkö, S., Korpimäki, T., Salminen, S., 2004b. New real-timequantitative PCR procedure for quantification of bifidobacteria in human fecalsamples. Applied and Environmental Microbiology 70, 4165–4169.
Gueimonde, M., Noriega, L., Margolles, A., de los Reyes-Gavilán, C.G., 2007. Induction ofalpha-L-arabinofuranosidase activity by monomeric carbohydrates in Bifidobacter-ium longum and ubiquity of encoding genes. Archives of Microbiology 187,145–153.
Harty, D.W., Chen, Y., Simpson, C.L., Berg, T., Cook, S.L.,Mayo, J.A., Hunter, N., Jacques, N.A.,2004. Characterisation of a novel homodimeric N-acetyl-beta-D-glucosaminidasefrom Streptococcus gordonii. Biochemical and Biophysical Research Communications319, 439–447.
Hellingwerf, K.J., Crielaard, W.C., Joost Teixeira de Mattos, M., Hoff, W.D., Kort, R.,Verhamme, D.T., Avignone-Rossa, C., 1998. Current topics in signal transduction inbacteria. Antonie van Leeuwenhoek 74, 211–227.
Hoch, J.A., 2000. Two-component and phosphorelay signal transduction. CurrentOpinions in Microbiology 3, 165–170.
Leahy, S.C., Higgins, D.G., Fitzgerald, G.F., van Sinderen, D., 2005. Getting better withbifidobacteria. Journal of Applied Microbiology 98, 1303–1315.
Masco, L., Huys, G., De Brandt, E., Temmerman, R., Swings, J., 2005. Culture-dependentand culture-independent qualitative analysis of probiotic products claimed tocontain bifidobacteria. International Journal of Food Microbiology 102, 221–230.
Meile, L., Rohr, L.M., Geissmann, T.A., Herensperger, M., Teuber, M., 2001. Characteriza-tion of the D-xylulose 5-phosphate/D-fructose 6-phosphate phosphoketolase gene(xfp) from Bifidobacterium lactis. Journal of Bacteriology 183, 2929–2936.
Mohammadi, T., Karczmarek, A., Crouvoisier, M., Bouhss, A., Mengin-Lecreulx, D., denBlaauwen, T., 2007. The essential peptidoglycan glycosyltransferase MurG forms acomplex with proteins involved in lateral envelope growth as well as with proteinsinvolved in cell division in Escherichia coli. Molecular Microbiology 65, 1106–1121.
Nakayama, J., Cao, Y., Horii, T., Sakuda, S., Akkermans, A.D.L., De Vos, W.M., 2001.Gelatinase biosynthesis-activating pheromone: a peptide lactone that mediates aquorum sensing in Enterococcus faecalis. Molecular Microbiology 41, 145–154.
Neher, S.B., Villén, J., Oakes, E.C., Bakalarski, C.E., Sauer, R.T., Gygi, S.P., Baker, T.A., 2006.Proteomic profiling of ClpXP substrates after DNA damage reveals extensiveinstability within SOS regulon. Molecular Cell 22, 193–204.
Noriega, L., Gueimonde, M., Sánchez, B., Margolles, A., de los Reyes-Gavilán, C.G., 2004.Effect of the adaptation to high bile salts concentrations on glycosidic activity,survival at low pH and cross-resistance to bile salts in Bifidobacterium. InternationalJournal of Food Microbiology 94, 79–86.
Novick, R.P., Projan, S.J., Kornblum, J., Ross, H.F., Ji, G., Kreiswirth, B., Vandenesch, F.,Moghazeh, S., 1995. The agr P2 operon: an autocatalytic sensory transductionsystem in Staphylococcus aureus. Molecular Genetics and Genomics 248, 446–458.
O'Riordan, K., Fitzgerald, G.F., 1998. Evaluation of bifidobacteria for the production ofantimicrobial compounds and assessment of performance in Cottage cheese atrefrigeration temperature. Journal of Applied Microbiology 85, 103–114.
Sánchez, B., Noriega, L., Ruas-Madiedo, P., de los Reyes-Gavilán, C.G., Margolles, A., 2004.Acquired resistance to bile increases fructose-6-phosphate phosphoketolaseactivity in Bifidobacterium. FEMS Microbiology Letters 235, 35–41.
Sánchez, B., Champomier-Vergès, M.C., Anglade, P., Baraige, F., de los Reyes-Gavilán, C.G.,Margolles, A., Zagorec, M., 2005. Proteomic analysis of global changes in proteinexpression during bile salt exposure of Bifidobacterium longumNCIMB 8809. Journalof Bacteriology 187, 5799–5808.
Sánchez, B., Champomier-Vergès, M.C., Collado, M.C., Anglade, P., Baraige, F., Sanz, Y., delos Reyes-Gavilán, C.G., Margolles, A., Zagorec, M., 2007a. Low-pH adaptation andthe acid tolerance response of Bifidobacterium longum biotype longum. Applied andEnvironmental Microbiology 73, 6450–6459.
Sánchez, B., Champomier-Vergès, M.C., Stuer-Lauridsen, B., Ruas-Madiedo, P., Anglade,P., Baraige, F., de los Reyes-Gavilán, C.G., Johansen, E., Zagorec, M., Margolles, A.,2007b. Adaptation and response of Bifidobacterium animalis subsp. lactis to bile: aproteomic and physiological approach. Applied and Environmental Microbiology73, 6757–6767.
Sánchez, B., Champomier-Vergès, M.C., Anglade, P., Baraige, F., de los Reyes-Gavilán, C.G.,Margolles, A., Zagorec, M., 2008. A preliminary analysis of Bifidobacterium longumexported proteins by two-dimensional electrophoresis. Journal of MolecularMicrobiology and Biotechnology 14, 74–79.

153L. Ruiz et al. / International Journal of Food Microbiology 133 (2009) 148–153
Schell, M.A., Karmirantzou, M., Snel, B., Vilanova, D., Berger, B., Pessi, G., Zwahlen, M.C.,Desiere, F., Bork, P., Delley, M., Pridmore, R.D., Arigoni, F., 2002. The genomesequence of Bifidobacterium longum reflects its adaptation to the humangastrointestinal tract. Proceedings of the National Academy of Sciences USA 99,14422–14427.
Sonnenburg, J.L., Chen, C.T., Gordon, J.I., 2006. Genomic and metabolic studies of theimpact of probiotics on a model gut symbiont and host. Public Library of ScienceBiology 4, e413.
Takano, E., Chakraburtty, R., Nihira, T., Yamada, Y., Bibb, M.J., 2001. A complex role for thegamma-butyrolactone SCB1 in regulating antibiotic production in Streptomycescoelicolor A3(2). Molecular Microbiology 41, 1015–1028.
Van Bogelen, R.A., Neidhardt, F.C., 1990. Ribosomes as sensors of heat and cold shock inEscherichia coli. Proceedings of the National Academy of Sciences USA 87,5589–5593.
Ventura, M., Fitzgerald, G.F., van Sinderen, D., 2005a. Genetic and transcriptionalorganization of the clpC locus in Bifidobacterium breve UCC 2003. Applied andEnvironmental Microbiology 71, 6282–6291.
Ventura, M., Zhang, Z., Cronin, M., Canchaya, C., Kenny, J.G., Fitzgerald, G.F., vanSinderen, D., 2005b. The ClgR protein regulates transcription of the clpP operon inBifidobacterium breve UCC 2003. Journal of Bacteriology 187, 8411–8426.
Visick, K.L., Fuqua, C., 2005. Decoding microbial chatter: cell–cell communication inbacteria. Journal of Bacteriology 187, 5507–5519.
Waters, C.M., Bassler, B.L., 2005. Quorum sensing: cell-to-cell communication inbacteria. Annual Review of Cell and Developmental Biology 21, 319–346.
Wilson, D.N., Nierhaus, K.H., 2005. Ribosomal proteins in the spotlight. Critical Reviewsin Biochemistry & Molecular Biology 40, 243–267.
Xavier, K.B., Bassler, B.L., 2005. Interference with AI-2-mediated bacterial cell–cellcommunication. Nature 437, 750–753.
Yuan, J., Zhu, L., Liu, X., Li, T., Zhang, Y., Ying, T., Wang, B., Wang, J., Dong, H., Feng, E., Li,Q., Wang, J., Wang, H., Wei, K., Zhang, X., Huang, C., Huang, P., Huang, L., Zeng, M.,Wang, H., 2006. A proteome reference map and proteomic analysis of Bifidobac-terium longum NCC2705. Molecular and Cellular Proteomics 5, 1105-1101.
Yuan, J., Wang, B., Sun, Z., Bo, X., Yuan, X., He, X., Zhao, H., Du, X., Wang, F., Jiang, Z.,Zhang, L., Jia, L., Wang, Y., Wei, K., Wang, J., Zhang, X., Sun, Y., Huang, L., Zeng, M.,2008. Analysis of host-inducing proteome changes in Bifidobacterium longumNCC2705 grown in vivo. Journal of Proteome Research 7, 375–385.






![Safety Evaluations of Bifidobacterium bifidum BGN4 and ... · sequence was reported to GenBank [31]. B. longum BORI, also isolated from a healthy breast-fed infant and deposited in](https://img.dokumen.tips/doc/110x75/5e1631ad061df060a06066fb/safety-evaluations-of-bifidobacterium-bifidum-bgn4-and-sequence-was-reported.jpg)




![Effect of Probiotics Lactobacillus and Bifidobacterium on ... · Bifidobacterium animalis NCIMB 702242 [27, 39, 47] Lactobacillus plantarum NCIMB 11974 [32, 41, 48, 49] Bifidobacterium](https://img.dokumen.tips/doc/110x75/5f0da2017e708231d43b51a3/effect-of-probiotics-lactobacillus-and-bifidobacterium-on-bifidobacterium-animalis.jpg)

















