Embed Size (px)
Citation preview

Copyright 0 1994 by the Genetics Society of America
Co-Segregation of Intermale Aggression With the Pseudoautosomal Region of the Y Chromosome in Mice
Pierre L. Roubertoux, Michele Carlier, Herve Degrelle, Marie-Claude Haas-Dupertuis, John Phillips and Rene Moutier
U. R.A. 1294 C. N. R.S. Ginitique, Neuroginitique, Comportement, Uniuersiti Paris-V Reni Descartes, 75270 Paras, Cedex 06, France
Manuscript received March 23, 1993 Accepted for publication September 1 1, 1993
ABSTRACT The sexual dimorphism of aggression has led to a search for its Y chromosomal correlates. We have
previously confirmed that initiation of attack behavior against a conspecific male is Y-dependent in two strains of laboratory mice (NZB and CBA/H). We provide evidence that the non-pseudoautosomal region of the Y is not involved and that only the pseudoautosomal region of the Y is correlated with initiation of attack behavior. The autosomal correlates also contribute to this behavior in an additive or interactive manner with the pseudoautosomal correlates.
T HE association of differences in agonistic behav- ior, defined as the initiation of attack behavior
against a conspecific male, with the variants of the Y chromosome in mice is the subject of a long-standing debate (CARLIER et al. 1990; CARLIER, ROUBERTOUX and PASTORET 199 1 ; MAXSON 1992a,b). On the one hand, three independent research teams using inbred strains of laboratory mice have implicated the Y chro- mosome in initiation of attack behavior against con- specific males, either from the comparison of strains congenic for the Y chromosome (MAXSON, GINSBURG and TRATTNER 1979; STEWART, MANNING and BATTY 1980) or from results of mendelian crosses (CARLIER, ROUBERTOUX and PASTORET 199 1). More- over the Y chromosome has been implicated in differ- ences between lines selected for short vs. long attack latency from a population of feral mice (Mus musculus domesticus; VAN OORTMERSSEN 1984; VAN OORT- MERSSEN, BENUS and SLUYTER 1992). On the other hand, a correlated response for female aggression in lines differentially selected only for male aggression has been reported (HOOD and CAIRNS 1988) suggest- ing that this behavior may not be transmitted by a locus linked to the Y chromosome. It is worth noting that in the studies using inbred strains, the Y effect always acts concomitantly with the autosomal back- ground (MAXSON, GINSBURG and TRATTNER 1979; CARLIER et al. 1990).
The opposing conclusions drawn between the re- sults of HOOD and CAIRNS and the other teams could be due to differences in behavioral testing, rearing conditions, or in differences in the genetic informa- tion borne either on the autosomes or on the Y chro- mosome. Obligatory crossing over between the X and the Y chromosomes (SIMLER et al. 1985; ELLIS and GOODFELLOW 1989) at the male meiosis leads to a
Genetics 135 225-230 (January, 1994)
parsimonious hypothesis to reconcile both arguments: a non-pseudoautosomal region of the Y chromosome (Y”’””) is transmitted from father to son, whereas all or part of the other region of the Y chromosome recombines with the X chromosome and is thus termed “pseudoautosomal region” (Y‘””). This shar- ing of the YPAR by both males and females is compatible with a common Y correlate for attack behavior in both sexes, and the so-called “autosomal” co-acting or in- teracting genes would, in fact, be “pseudoautosomal,” i.e., located on the YPAR as previously suggested (CAR- LIER et al. 1990). Male mice from strain NZB/BI- NJGnc (abbreviated N hereafter) display more intense and frequent spontaneous offensive behavior against conspecific males than CBA/HGnc mice (abbreviated H) (CARLIER and ROUBERTOUX 1986). The frequency of males initiating attack behavior is higher in N than in H for dyadic encounters with a standard opponent. Moreover, when the maternal environment is kept constant, this difference is not modified, showing that the observed strain difference has genetic and/or cy- toplasmic correlates. We have previously demon- strated that at least one locus on the Y chromosome is associated with attack behavior in these populations (CARLIER, ROUBERTOUX and PASTORET 1991), but it was not known whether this locus is located on the YNPAR or on the YPAR. In the present study we describe a co-segregation of the YPAR with the initiation of attack behavior.
MATERIALS AND METHODS
Mice: Identified breeders from two highly inbred strains of mice were used: H and N. They were supplied by the CSEAL (CNRS, Orlians la Source, France) at 127 and 1 14 brother X sister generations of mating, respectively, and had been maintained using the same mating system, in our

226 P. L. Roubertoux et al.
laboratory, for a further 8 generations at the beginning of the experiment.
Rearing conditions: The following conditions have been kept constant over the years: temperature 23.5" & 0.5"; photoperiod, 12:12 with lights on at 8:30 AM; food (IM UAR) and tap water ad libitum; dust-free sawdust bedding; weaning 30 & 1 days after birth. Each pregnant female was isolated from the mating cages. The litters having fewer than four pups were discarded and the others culled to six pups. At weaning, each male was housed with one female (generally a littermate) in an opaque cage (42 X 27 X 17 cm) until testing (66 If: 4 days after birth).
Behavioral testing: The conditions of observation and measurement (described by CARLIER and ROUBERTOUX 1986) were maintained unchanged for all groups over the years. Briefly, the test was performed in a transparent cage (42 X 26 X 18 cm) with a transparent lid. The floor of the cage was covered with sawdust from several cages housing males of the tested groups. This procedure accelerates the appearance of the first attack (CARLIER and ROUBERTOUX 1986). Several observations were carried out on a given day and the groups were matched for order of observation. Each test was a dyadic encounter with an A/JOrl (A) male as the opponent. This strain had been chosen for its low scores of aggression and, in fact, the A males rarely initiated attack. The males tested in such a situation were discarded. The A opponent came from a group male cage. The behavioral records started when the tested male sniffed the A opponent for the first time, and the test lasted 6 min in cases where no attack occurred. Several variables were measured but only the frequency of attacking males will be considered here.
Crosses: Only crosses informative for the demonstration of a co-segregation between the YPAR and initiation of attack behavior are reported here.
First, congenic strains for YNPAR were developed in 1983 by CARLIER. The H-YNPAR was substituted by the N-YNPAR in the H strain to obtain its congenic H.N-YNPAR for this region of the Y chromosome and secondly by substituting the H- YNPAR in the place of the N-YNPAR in the N strain to obtain its congenic N.H-YNPAR. The congenic N.H-YNpAR was devel- oped with N as recipient strain and H as donor, the N females being sired by NHF, males (in each cross the moth- er's genotype will be given first, separated by x from the father's genotype; within each genotype the first letter+) indicate the strain of the mother) and the backcross males subsequently sired again with N. The symmetrical design was used to obtain the second congenic H.N-YNPAR. The N.H-YNpAR and the H.N-YNPAR reached 29 and 30 backcross generations, respectively, and were observed between the 2nd and 21st repeated backcross. At every generation it is assumed that the congenic progeny lose 50% of the alleles from the congenic donor. Thus more than 99.99% of the allelic forms of the parental recipient strain, located throughout the genotype, including the YPAR, are expected in the congenic, at the 21st backcross. The isogenicity of the genetic background was directly confirmed by skin graft acceptance and by the similarity of mandible shape. First, tail skin reciprocal graftings were performed in females from H and H.N-YNPAR on one hand, and from N and N.H-YNPAR on the other, at the 9th backcross generation. One rejection out of 11 was observed in the first group and none in the second, 347 days after the graft. Second, 11 mandible measurements were performed on the same four groups. There is a high similarity for pairwise squared distances for a pair of congenic strains under the hypothesis of isogenicity, since BAILEY (1985) estimated the number of genes respon- sible for morphological changes on murine mandible to be over 100. No significant difference was found within the
same sex between H vs. H.N-YNPAR and between N vs. N.H- YNpAR (MOUTIER and CARLIER 1991).
Second, the parental H and N females were sired with the parental and congenic males, thus providing two sets of reci rocal Fls: HNF,, NHF, on one hand and HN.H- YNp RF1, NH.N-YNPARFI on the other. These last two groups were constituted after the 8th generation of backcross. Results on HNF, and NHF, have already been published (CARLIER and ROUBERTOUX 1986; CARLIER, ROUBERTOUX and PASTORET 199 1). New groups have since been studied and because these gave very similar results, the data of all the males from the same F, were grouped.
Third, two sets of backcrosses were developed with H and N parental males on reciprocal HNF, and NHF, moth- ers: HN X H, HN X N, NH X H, and NH X N.
Fourth, reciprocal backcrosses on FI mothers whose ova- ries had been removed and replaced in situ by ovaries from parental strains were obtained. As NHF, females are histo- compatible with H and N tissues, their ovaries were removed and replaced by ovaries from N or H females according to a technique described previously (ROUBERTOUX and CAR- LIER 1988; ROUBERTOUX, NOSTEN-BERTRAND and CARLIER 1990; CARLIER, NOSTEN-BERTRAND and MICHARD 1992). The females were labeled NoFl (for those providing N ova) or HoFl (for those providing H ova). Two controls were performed: first, to test for a transplantation effect per se, intrastrain ovary transplantations were used: males born from NoN females (N females whose ovaries had been removed and replaced in situ by ovaries from the N strain) were compared to those with identical genotype born from ungrafted N females, a symmetrical procedure being em- ployed for HoH and H females. No effect due to grafting has been shown for initiation of attack behavior (ROUBER- TOUX and CARLIER 1988). Second, controls for the genotype of the pups were required because part of the ovary of the host female can remain after ovarectomy. Four independent genetic markers (coat color and three electrophoretic mark- ers) were used to detect discrepancies between the expected and observed genotypes. Individuals were first observed in the behavioral test, they were then typed employing the four genetic polymorphic markers. Only those for which the observed genotype fitted the expected were considered for the subsequent statistical analysis. One subject was dis- carded for this reason in the NoFl X HN backcross (leading to an observed error rate of 1/65).
Genetic polymorphism for the YPAR; measurement of liver steroid sulfatase (STS) activity: The hypothesis of co- segregation of the YpAR and initiation of offense behavior, can be tested using known differences between N-YPAR and H-YPAR. The liver STS (EC 3.1.6.2) activity is controlled by the Sts locus (mapped on the YPAR; KEITCES et al. 1985) and was therefore used as a marker. The technique of measure- ment of the initial production rate of ['Hlestrone from ['HI estrone sulfate (expressed as picomoles of ['Hlestrone pro- duced per min and per mg protein; PROST and ADFSSI 1983) was modified from BURSTEIN and DORFMAN (1 963). Briefly, (1) the substrate concentration must not be rate-limiting since the catalysis is inhibited by an excess of substrate. Thus, to obtain the V,,, and K , from the Lineweaver-Burk reciprocal plot, we performed three substrate concentra- tions and a zero time to detect possible free steroid (each in duplicate). (2) Chromatographic purification of the radio- active substrate was needed since it can be partially hydro- lyzed and/or radiolyzed. (3) The reaction was stopped with NanCOs (and not NaOH) to prevent ionization of free estrone, which was extracted using petroleum benzine.
The liver STS activity was measured in males from the parental and their congenic strains, and also in NH.N-
R
yNPAR FI and HN.H-YNpARFI males.

Intermale Aggression 227
Statistics and experimental design: The differences be- tween proportions of attacking males were tested here with a logit model analysis using the SAS CATMOD procedure. All the main effects and interactions are included in this model and tested according to an analysis of variance design as indicated in the SAS/STAT Guide for Personal Computers (1 987).
The contribution of the YNPAR to initiation of attack behavior was tested by comparing males from each parental and its congenic strain. The behavioral observations were not performed at each generation of the congenics after backcross 4. The statistical analysis was performed by pool- ing the generations 2-6, 7-1 3 and 14-21 within each con- genic. A group of each corresponding parental strain was observed each year allowing a comparison between the congenic and their parental strains. The proportions of attacking males were thus compared with a 3 X 2 X 2 desi n; three groups of generations and parental strains, two YN 9 A R
(H and N) and two backgrounds (H and N). The two sets of reciprocal Fls from parental females sired
with parental and congenic males constitute a segregating population for the separation of the possible contribution of YN AR and YPAR. The HNFl and HN.H-YNPARFI share the same YpAR from N and have different non-pseudoautosomal re ions. Similarly, NHFl and NH.N-YNPARF1 share the same Yp B from H but have different non-pseudoautosomal re- gions. A logit model analysis of the attack proportions following a 2 x 2 design was thus performed to test for the effect of each part of the Y (pseudoautosomal and non- pseudoautosomal). However, other candidate factors (the maternal environment, differential genomic imprinting, mi- tochondrial DNA and the X chromosome) also covary with
and YPAR, in these crosses, and thus might be respon- sible for an apparent co-segregation of one part of the Y with initiation of attack behavior in these reciprocal Fls. It was thus necessary to eliminate the possible effects of these factors.
The contribution of differences in maternal environment can be eliminated if the co-segregation of one part of the Y and initiation of attack behavior still persists when this environment is kept constant. Moreover, possible differen- tial genomic imprinting (for example, the X from H inhib- iting the expression of N-YNPAR in the subjects having re- ceived H-YPAR) is not compatible with a co-segregation which is always found in the backcrosses, regardless of whether the genotypic contribution of the mother is H or N. The effect of these potential factors were tested employing reciprocal backcrosses on F1 mothers bearing in situ grafted ovaries from N and H.
The mitochondrial DNA (mtDNA) is almost entirely ma- ternally transmitted (GYLLENSTEIN et ul. 1991) and H and N mothers have mtDNA from different origins, domesticus and brwirostris, respective1 (YONEKAWA et al. 1982). Con- sequently, the HN.H-YNPA$, males that bear the N-YPAR have also inherited the mtDNA from H, whereas the NH.N- YNPARFl, bearing the H-yPAR, have received the mtDNA from N. Backcrosses from the reciprocal F1 females provide the opportunity to test for an implication of mtDNA on initia- tion of attack behavior. Offspring of the first set of back- crosses with reciprocal F1 females HN X H and HN X N compared to those of the second NH X H and NH X N, have, in probability, the same number of autosomal or x- linked allelic forms from N and H, an identical Y chromo- some and are exposed to identical maternal effects. They only differ by the origin, H us. N, of mtDNA and were used, here, to test for its implication in initiation ofattack behav- ior.
The HNFI and HN.H-YNPARFI us. the NHFl and NH.N- 1 also differ by the X chromosome from H us. N. The
yNPAR
YNPARF
sources of variation between the backcrosses from F1 females with parental males and the backcrosses from FI females bearing N or H ovaries with HNF, and NHFl males are presented in Table 1.
In each pair, 1 us. 1’ or 2 us. 2’, the two backcrosses share identical autosomal information and maternal environment (uterine and postnatal). They differ by the origin of the
some. A possible involvement of this chromosome in the initiation of attack behavior could be deduced for the within- pair com arisons pending the demonstration of an absence of the YZAR effect, deduced from the congenic strains.
The liver STS activity was compared by analysis of vari- ance.
yNPAR and a parental N or H or a recombinant X chromo-
RESULTS
The parental and their congenic strains: The per- centages of attacking males are presented in Figure 1 for the three groups of congenic and parental strains. The three groups of pooled backcrosses and their corresponding parental groups differ (x& = 8.85, P = 0.01). Inspection of Figure 1 shows that males tend to attack less in the groups of the more recent gener- ations. However, the interactions between this de- crease and the background on one hand and the YNPAR on the other are not significant (x& = 0.36, P = 0.83, and x& = 0.98, P = 0.61). The H and N background effect is significant (x(?) = 52,35, P < 0.0001): the percentage of attacking males is higher in N than in H, as we have previously shown (CARLIER and ROUB- ERTOUX 1986). The YNPAR effect is not significant (x(?, = 0.14, P = 0.70) nor is the interaction between the H and the N background and the YNPAR (x(:, = 0.30, P = 0.58).
The reciprocal Fls: The percentage of attacking males in each group of reciprocal Fls are presented in Figure 2. The effect of the non-pseudoautosomal part of the Y and the interaction between the two parts of the Yare not significiant (x$, = 0.000). However, the effect of the pseudoautosomal region plus maternal effects, including the mtDNA and the X contribution is significant (x(?) = 10.43, P = 0.001): the two groups having the pseudoautosomal part of the Y from the N strain (HNF1 and HN.H-YNPARFl) attack more fre- quently, suggesting that if the difference between the reciprocal Fls is borne on the Y chromosome it is independant of the genetic information borne on the
The backcrosses with reciprocal F1 females: The sample sizes and the percentages of males that initiated attack behavior in the offspring of the backcrosses were: H N X H ( n = 21; 38.09%), HN X N (n = 20; SO%), NH X H ( n = 24; 29.17%) and NH X N (n = 26; 69.23%). Only the origin of the father reached the level of significance (x(?) = 14.80; P < 0.0002). T h e males born from HNFI or NHFl mothers did not differ (x(?) = 1.06; P = 0.30). Consequently, as no effect of the mother was detected, the present data suggest that attack behavior differences are indepen- dant of mtDNA origin in these strains.
p P A R
228 P. L. R o u b e r t o u x et al.
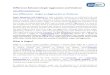
TABLE 1
Sources of variation between reciprocal backcrosses on F1 mothers bearing H and N ovaries. NoFI:FI females bearing N ovaries; HoFI:FI females bearing H ovaries; NPAR, non-pseudoautosomal region; PAR, pseudoautosomal region
PAR
Backcross generations environment mtDNA Autosomes y N P A R On X On Y X N P A R
Maternal
NOFI X N H ( 1 ) FI N 314 N 114 H H N FI N NHFl X N (1 ') FI N 314 N 114 H N FI N FI HoFl X H N (2) FI H 114 N 314 H N H FI H HNF1 X H (2') FI H 114 N 314 H H FI H FI
EN.H-Y NPAR 1z1 H.N-Y NPAR
;I x, 51) 128 -li I O I n 51) oo 1; 1 1 , 02
2 t o 6 7 to 13 1 4 t o 2 1 Generations of backcrosses
FIGURE 1 .-Percentage (+sE) of attacking males for the congenic and parental strains at three stages of backcrosses. N , number of subjects
[ ? z:
c
5:
38
-n. z
151
FIGURE 2.-Percentage (+sE) of attacking males for four groups of reciprocal Fls between parental strains H and N and between parental and congenic strains H.N-ppAR and N.H-ppAR. Number of subjects are indicated under the x axis.
Reciprocal backcrosses from HNFl, NHFI, NoFl and HoFl females: There was no effect of the YNPAR on the phenotype under study and thus the equations 1 , 1 ', 2, 2' could be used to test for the effect of the parental (H or N) us. X chromosome recombining H and N information. The comparisons for proportions of attacking males differ neither between ( 1 ) offspring of NoFl X NH (n = 15; 93.33%) and ( 1 ') offspring of NHFl X N (n = 26; 69.23%) (x:(1,2 = 1.97; P = 0. lo), nor between (2) offspring of HoFl X HN (n = 22;
13 I
FIGURE S."Percentage (+SE) of attacking males in offspring of reciprocal backcrosses using FI mothers bearing H or N ovaries and FI males. Number of subjects are indicated under the x axis (from CARLIER, ROUBERTOUX and PASTORET 199 1).
36.36%) and (2') offspring of HNFl X H (n = 21; 38.10%) (x(?, = 0.14; P = 0.9 1 ) .
Reciprocal backcrosses on F1 mothers bearing H or N ovaries: The four groups were: NoF, X NH (n
and HoFl X HN (n = 22). Here the maternal environ- ment effects are kept constant. Under these conditions (Figure 3), there is an effect of the genotype of the mother H us. N (x(?) = 5.03; P < 0.03): i .e. , the male progeny attack less when born from H rather than from N females. An effect of the genotype of the father is also shown (x:l) = 5.94; P < 0.02). The partial comparisons demonstrate that the male offspring from NHF, fathers that have received the H-YNPAR and the N-YPAR attack more frequently than the males which have received the N-YNPAR and the H-YPAR from HNFl fathers, whatever the genotype or the maternal environment provided by the mother (CARLIER, ROUBERTOUX and PASTORET 199 1).
The liver STS activity : The activity of this enzyme (Table 2) had a significantly lower value in the H and H.N-YNPAR which share the same H-YPAR compared to the N and N.H-YNPAR which share the same N-YPAR, the other sources of variation, including the interac- tion between the YNPAR and the autosomes, were not significant. This result indicates that two different allelic forms of the Sts gene corresponding to different levels of enzymatic activity are borne on the N-YPAR
= 15), HoFl X NH (n = 14), NoFl X HN (n = 13)

Intermale Aggression 229
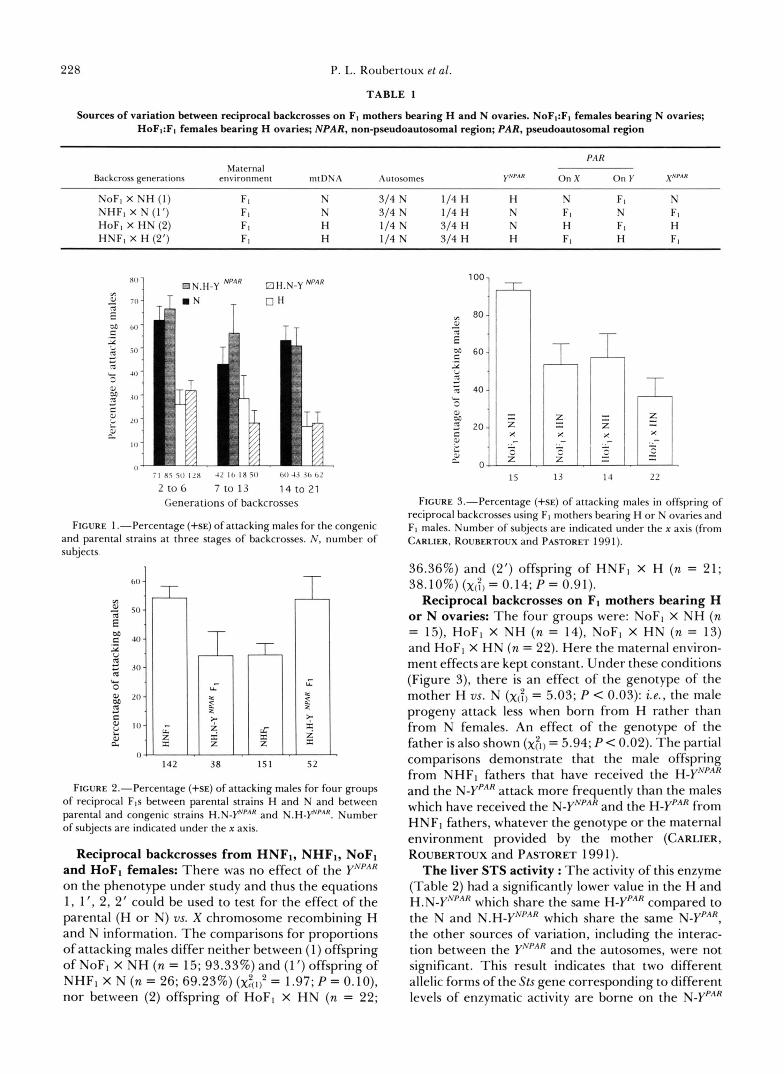
TABLE 2
Steroid sulfatase activity (pmol/min/mg protein) in parental and congenic strains and one of their reciprocal hybrids (n, number of subjects; see text for other abbreviations)
Genotypes N N.H-PpAR H H.N-PpAR N H . N - Y N ~ ~ ~ F , H N . H - Y N ~ ~ ~ F ,
n 5 9 8 8 6 5 R f SE 167.2 f 29.1 141.4 * 14.5 77.8 f 16.7 56.8 f 5.6 32.0 f 9.3 65.43 f 5.1
( 4 (a) (b) (b) ( 4 ( 4
The strains with the same letter in the last row did not differ. The difference between (a) and (b) is significant: F(2,26) = 23.48 (P < 0.001), as is the difference between (c) and (d): t(9) = 2.97 (P < 0.05).
and H-YPAR. Moreover, the significantly lower STS activity found in the NH.N-YNPARFI compared to the HN.H-YNPARF1, suggests an under-expression of Sts when carried on the XPAR in these strains. The iden- tical activities for the Y- and X-borne loci reported by JONES et al. (1 989) are most probably limited to the Sts silent allele employed by these authors.
DISCUSSION
The percentage of males initiating attack behavior is higher in strain N than in H for dyadic encounters with an A male as standard opponent as shown in Figure 1. We know that this difference has at least one genetic correlate (CARLIER, ROUBERTOUX and PASTORET 1991). The present results indicate that one locus is located on the YPAR.
Co-segregation of initiation of attack behavior with YNPAR was first examined using the permutation of the
ent segregation between initiation of attack behavior and YNPAR appears as soon as the first backcross and still persists at the 2 1 st. This clearly demonstrates that the locus is not on the YNPAR. On the contrary, the absence of dissociation between the initiation of attack behavior and the YPAR in different segregating gener- ations, strongly suggests that the genetic correlates for attack behavior are located on the YPAR chromo- somal region.
The groups HN.H-YNPARF1, and HNFl (sharing the same YPAR from N and having different Y non-pseu- doautosomal regions) have a high attack behavior, whereas NH.N-YNPARFl and NHFl (sharing the same
regions) have a low attack behavior (Figure 2). Other factors, presented in the experimental design, also covary with YPAR but the results presented here show that they have no significant effect. (1) Initiation of attack behavior is independent of the origin of mtDNA, as shown here by the comparison of back- crosses with reciprocal F1 females. (2) For several reasons, the X chromosome cannot contribute to the difference in frequency of initiation of attack behav- ior. First, there is a poor likelihood of its involvement since the individuals which have received the X from H, the less attacking strain, attack more frequently than those which have received the X from N, the
yNPAR between strains N and H (Figure 1). Independ-
yPAR from H but different Y non-pseudoautosomal
more attacking strain. Second, the association of N- YPAR with a high incidence of attack behavior is always present in the reciprocal backcrosses on F1 mothers whether they bear H or N ovaries (and consequently an H or an N X chromosome). Third, the inheritance of an X chromosome either from H or N compared to an X recombining H and N information, did not produce any difference in the incidence of attack behavior. (3) The covariation of the frequency of attack behavior with YPAR still persists when the ma- ternal environment is kept constant, in the reciprocal backcrosses on F1 mothers bearing H or N ovaries: in the experiment illustrated in Figure 3 only the pseu- doautosomal region was correlated with the attack behavior in the expected direction. (4) Possible differ- ential genomic imprinting (X from H inhibiting the expression of N-YNPAR) in the subjects having received H-YPAR, is not compatible with the frequency of attack behavior differences always found to be in the same direction in the backcrosses, regardless of whether the genotypic contribution of the mother is H or N.
The agreement of the data presented here with the hypothesis of a genetic correlate of attack behavior located on the pseudoautosomal part of the Y chro- mosome is reinforced by the demonstration of an enzymatic polymorphism for H-YPAR and N-YPAR. However, high attack behavior is associated with the N genetic information of the pseudoautosomal region only when it is borne on the Y chromosome and not on the X. On the other hand, the low frequency of attack behavior is associated with the genetic infor- mation from H, only when it is borne on the YPAR and not on the XPAR. The description of a polymorphism for the Sts gene, previously mapped on this region, strengthens this hypothesis. We have observed (Table 2) that the STS enzymatic activity is higher when measured in individuals bearing the N-YNPAR and the H-XNPAR compared to those bearing the H-YNPAR and the N-XNPAR, suggesting that the contribution to the phenotype could vary, according to the region (on the X or on the Y) on which it is borne. However, the differences in attack behavior do not seem to be the consequence of a pleiotropic effect of allelic substitu- tions at the Sts locus, as suggested by the positive correlation between attack behavior and STS enzy- matic activity, in N and H strains. This correlation

230 P. L. Roubertoux et al.
becomes negative with other strains: the C57BL/10 males that attack less than DBA/1 (SELMANOFF, MAX- SON and GINSBURG 1976) have a significantly higher STS enzymatic activity (67.9 & 15.2 vs. 41.9 & 6.7; - unpublished).
The YPAR alone cannot be considered responsible for the difference in this behavior since the offspring of the backcrosses from a mother belonging to the more attacking strain N attack more frequently than those of the backcrosses from an H mother (less at- tacking strain), when the strain of the father and the maternal environment are held constant (NoFI X HNFl > HoFl X HNFl and NoFl X NHFl > HoFl X NHF]); see Figure 3. Thus autosomal homozygotic alleles clearly also contribute to the difference either in an additive or interactive manner with the YPAR.
We thank W. E. CRUSIO, B. DRESP, P. MANDEL and B. MARTIN for useful advice and J. HIRSCH and S. C. MAXSON for discussion and suggestions during the preparation of the manuscript. We also thank our anonymous referees for their helpful comments. This work was supported by CNRS (URA 1294), DRED (Universitk Paris V Renk Descartes), Fondation pour la Recherche Mkdicale, Naturalia et Biologia, NATO Grant 0235/89, and Universiti de Reims Champagne Ardenne.
LITERATURE CITED
BAILEY, D. W., 1985 Genes that effect the shape of the murine mandible. Congenic strain analysis. J. Hered. 7 6 107-1 14.
BURSTEIN S., and R. I . DORFMAN, 1963 Determination of mam- malian steroid sulfatase with 7-aSH 36-hydroxyandrost-5-en- 17-one sulfate. J. Biol. Chem. 238 1656-1660.
CARLIER, M., M. NOSTEN-BERTRAND and C. MICHARD, 1992 Separating genetic effects from maternal environmental effects, pp. 1 11-1 26 in Techniques for the Genetic Analysis of the Brain and Behavior: Focus on the Mouse, edited by D. GOLDOW- ITZ, D. WAHLSTEN and R. WIMER. Elsevier, Amsterdam.
CARLIER, M., AND P. L. ROUBERTOUX, 1986 Differences between CBA/H and NZB mice on intermale aggression. I. Comparison between parental strains and reciprocal Fls, pp. 47-57 in Genetic Approaches to Behavioral Phenotypes, edited by J. ME- DIONI and G. VAYSSE. Privat, Toulouse.
CARLIER, M., P. L. ROUBERTOUX and C. PASTORET, 1991 The Y chromosome effect on intermale aggression in mice depends on the maternal environment. Genetics 129 231-236.
CARLIER, M., P. L. ROUBERTOUX, M. L. KOTTLER and H. DE- GRELLE, 1990 Y chromosome and aggression in strains of laboratory mice. Behav. Genet. 20: 137-156.
ELLIS, E., and P. N. GOODFELLOW, 1989 The mammalian pseu- doautosomal region. Trends Genet. 5: 406-4 10.
GYLLENSTEN, U., D. WHARTON, A. JOSEFSSON and A. C. WILSON, 1991 Paternal inheritance of mitochondrial DNA in mice. Nature 352: 255-257.
HOOD, K. E., and R. B. CAIRNS, 1988 A developmental-genetic
analysis of aggressive behavior in mice. 11. cross-sex inheritance. Behav. Genet. 18 605-619
JONES, J., J. P. PETERS, C. RASBERRY and B. M. CATTANACH, 1989 X-inactivation of the Sts locus in mouse: anomaly of the dosage compensation mechanism. Genet. Res. 53: 193-199.
KEITGES, M. RIVEST, M. SINISCALCoandS. M. GARTLER, 1985 X- linkage of steroid sulphatase in the mouse is evidence for a functional Y-linked allele. Nature 315: 226.
MAXSON, S. C., 1992a Methodological issues in genetic analyses of an agonistic behavior (offense) in male mice, pp. 349-374 in Techniques for the Genetic Analysis of the Brain and BehavMr: Focus on the Mouse, edited by D. GQLDOWITZ, D. WAHLSTEN and R. WIMER. Elsevier, Amsterdam.
MAXSON, S. C., 1992b Potential genetic models of aggression and violences in males, pp. 174-188 in Genetically Dejned Animals Models of Neurobehavioral Dysfunctions, edited by P. DRISCOLL. Birkhauser, Boston.
MAXSON, S. C., B. GINSBURG and A. TRATTNER, 1979 Interaction of Y-chromosomal gene@) in the development of intermale aggression in mice. Behav. Genet. 9 219-226.
MOUTIER, R., and M. CARLIER, 1991 Mandible shape analysis in Y-congenic strains of mice. Lab. Anim. 25 303-307.
PROST, O., and G. L. ADESSI, 1983 Estrone and dehydroepian- drosterone sulfatase activities in normal and pathological hu- man endometrium biopsies. J. Clin. Endocrinol. Metab. 56
ROUBERTOUX, P. L., and M. CARLIER, 1988 Differences between CBA/H and NZB mice on intermale aggression. I1 Maternal effects. Behav. Genet. IS: 175-184.
ROUBERTOUX, P. L., M. NOSTEN-BERTRAND and M. CARLIER, 1990 Additive and interactive effects between genotype and maternal environments, concepts and facts. Adv. Study Behav.
SAS/STAT, 1987 Guide for Personal Computers, Version 6 Edition, SAS Institute Inc., Cary, N.C.
SELMANOFF, M. K., S. C. MAXSON and B. GINSBURG, 1976 Chromosomal determinants of intermale aggressive be- havior in inbred mice. Behav. Genet. 6: 53-69.
SIMMLER M. C., F. ROUYER, G. VERGNAUD, M., NYSTROM-LAHTI, K. Y. NGO, A. DE LA CHAPELLE and J. WEISSENBACH, 1985 Pseudoautosomal DNA sequences in the pairing region ofthe human sex chromosomes. Nature 317: 692-697.
STEWART, A. D., A. MANNING and J. BATTY, 1980 Effects on y chromosome variants on the male behaviour of the mouse MUS musculus. Genet. Res. 35: 261-268.
VAN OORTMERSSEN, G. A., 1984 A behavioral-genetic approach to variation in attack latency, serum testosterone level, and other agonistic parameters. Aggressive Behav. 10 177-1 78.
VAN OORTMERSSEN, G. A,, R. F. BENUS and F. SLUYTER, 1992 Studies on wild house mice. IV. On the heredity of testosterone and readiness to attack. Aggressivee Behav. 1s:
YONEKAWA, H., K. MORIWAKI, 0. COTOH, N. MIYASHITA, s. MIG- ITA, F. BONHOMME, J. P. HJORTH, M. L. PETRAS and y. TA- GASHIRA, 1982 Origins of laboratory mice deduced from restriction patterns of mitochondrial DNA. Differentiation 22:
Communicating editor: R. E. GANSCHOW
653-661.
19 205-247.
143-148.
222-226.