Embed Size (px)
Citation preview

Cloning, Structural Analysis and Expression of Fast Twitch Skeletal Muscle TroponinC2 (TNNC2) Gene in Goat
Haolin Chen, Daihua Wang, *Gangyi Xu, Tintin Wu, Chengli Zheng College of Animal Science and Technology, Sichuan Agricultural University, Ya’an, Sichuan, 625014 China
Haolin Chen E-mail: [email protected] * Corresponding author E-mail: [email protected]
Abstract—TroponinC2 (TNNC2), a constituent of the troponin complex located on the thin filament, plays a critical role in skeletal muscle contraction and improving meat quality traits of livestock and poultry. The TNNC2 gene has been identified in many mammals, but very little known about goat. In the study, we cloned TNNC2 gene in goat for the first time and analyzed its tissue expression. Results indicated that TNNC2 contains 483bp CDS and encodes 160 amino acids, and it was selectively expressed in the muscular tissues of goat. Its nucleotide sequence was high similar with sheep (93.38%) and alpaca (90.48%), and the identity of encoded amino acids which was varied from 100% (sheep) to 70.81% (Atlantic salmon) in animal. We found that the Ala113 and leu132 was distributed very law in mammals. The amino acid residue of Ca2+ binding sitesⅡwas identical in mammals, Aves and amphibian. This study may provide more insight into the molecular structure and expression patterns of the vertebrate TNNC2 gene.
Keywords-TroponinC2 gene; Molecular cloning; Structural analysis; Expression; Goat
I. INTRODUCTION The TNNC proteins belong to a member of multigene
family of Ca2+ binding proteins[1].There are two isoforms of TNNC proteins found in vertebrate striated muscle, troponinC2 (TNNC2) expressed in fast skeletal muscle and troponinC1 (TNNC1) expressed in slow skeletal muscle and cardiac muscle. The difference of the two isoforms is that TNNC2 is activated by Ca2+ binding siteⅠandⅡon the N-terminal, while TNNC1 is activated by Ca2+ binding siteⅡ.
TroponinC2, which has four Ca2+ binding sites, is a prominent member of the troponin complex. Each of these Ca2+ binding sites is characterized by a 12-residue Ca2+ binding loop that is interposed between a pair of a-helices. Each of the Ca2+ binding loops is rich in acidic amino acids (Asp and Glu) that are responsible for the coordination of a single Ca2+ [2]. The integral structure of TNNC2 consists of a N-terminal and a C-terminal globular domains connected by a flexible central helix, giving it a dumbbell-like shape, and each globular domain occupies a pair of EF-hand motifs which can bind to metal ions[3].
The diameter and type of muscle fiber will directly influence on meat tenderness, in which the types of myofiber can be classified according to their contractile nature[4]. As
a subunit of Troponin C (TNNC), fast skeletal troponinC2 (TNNC2) plays a critical role in skeletal muscle contraction[5]. TNNC2 gene has expressed during the myoblast differentiation and skeletal muscle development[6]. The allele T have positive effects on tenderness and marbling score, and suggest that the allele could be used as a molecular marker to study the meat quality in sheep[7]. Thus, the gene involved in skeletal muscle growth, myoblast differentiation and skeletal muscle contraction and was considered usually as potential candidate gene for meat quality[8], and identified to be an excellent model system for studies of developmentally regulated gene expression in skeletal muscle[9].
Tianfu goat is an emerging breed in China, which formed by multiple crossbreeding with the local breeding Chengdu Ma goat for maternal and imported England Saanen, Nubian, Toggenburg and Boer for paternal. This goat breed is noted for good quality meat, rapid growth, roughage resistance, strong adaptability [10]. For this reason, we cloned the TNNC2 gene in goat and analyzed its structural and the tissue expression.
II. MATERRIALS AND METHODS
A. Experimental animals and tissues preparation Tianfu goats were slaughtered at six month, one year,
two years and adult. The tissue samples of the carcasses (heart, liver, lung, kidney, abdominal muscle, longissimus muscle, gluteus maximus and biceps brachii) were collected immediately after slaughter and frozen in liquid nitrogen jar for extracting total RNA.
B. Cloning of TNNC2 gene and construction of recombinant plasmid Total RNA was extracted from goat by using Trizol
(TaKaRa, Dalian, Chian). According to a procedure of the synthesis kit, the first strand cDNA was synthesised from total RNA. According to Bos taurus TNNC2 gene sequence (GenBank Accession No. NM001076373), a pair of primer was designed with software primer 5.0 in the conserved region. The forward primer was 5′-TGATGACGGACCAGCAGG-3′, and the reverse primer was 5′-CTTCTTACTGCACGCCCTC-3′. Then using the primers, a part of cDNA fragment was amplified by RT-PCR with first strand cDNA as templates. The PCR started with pre-denature at 95 °C for 5 min, followed by 35 cycles
283
2011 International Conference on Bioscience, Biochemistry and Bioinformatics IPCBEE vol.5 (2011) © (2011) IACSIT Press, Singapore

(94 °C, 40 s; 53 °C, 40 s; 72 °C, 50 s),and ended with final extension at 72 °C for 10 min. Then the PCR products were separated by electrophoresis on a 1.5% agarose gel, and purified by using the PCR Rapid Purification Kit (BioDev-Tech). Purified PCR products were cloned by pMD18-T vector (TaKaRa, Dalian) and were sent to TaKaRa Biotechnology (Dalian) Co. Ltd. for sequencing.
C. Sequence analysis Using BLAST software, we performed a sequence
similarity analysis for the TNNC2 nucleotide and amino acid sequences (http://www.ncbi.nlm.nih.gov/BLAST). Also, we determined a theoretical molecular weight and isoelectric point by using Peptide Mass (http://us. expasy.org/tools/peptide-mass.html).The phosphorylation site was estimated by the neural network (http://www.cbs.dtu.dk/services/NetPhos). Multiple alignment analysis was conducted by the program CLUSTAL X and Bio-Edit software, and the phylogenetic analysis was performed by MEGA version 4.0 with Neighbor-Joining method. The SWISS-MODEL(http://swissmodel.expasy.org) was used to model the protein conformation, which was viewed in the Swiss Pdb Viewer[11]
D. RT-PCR expression analysis Tissue-specific expression pattern of TNNC2 was
performed by semi-quantitative (RT)-PCR using goat β-actin gene as an internal expression control. The primers of β-actin, which were used to adjust the concentration of cDNA, was designed based on the nucleotide sequence deposited in the Gene Bank (Accession No. U39357). TNNC2 primer pairs: 5′-GCCAGACACCCACCAAAG-3′ and 5′-CGAAGATGCGGAAACACT-3′. β-actin primer
pairs: 5′-GTCACCAACTGGGACGACA-3′ and 5′-AGGCGTACAGGGACAGCA-3′. Total RNA was extracted from the heart, liver, lung, kidney, abdominal muscle, longissimus muscle, gluteus maximus and biceps brachii respectively. The amplifications were performed in a 20μL reaction volume containing 12.5μL of 2× SYBR Premix ExTaq (TaKaRa, Dalian), 5μL of each primer, 2μL of diluted cDNA, 4.5μL ddH2O. The PCR amplification was carried out as follows: 95℃ for 10 s followed by 40 cycles of 95℃ for 5 s and 60℃ for 34 s. The 2 – Ct △△ method was used to analyze the expression level of the gene in goat[12]
III. RESULT
A. cDNA cloning and sequence analysis of TNNC2 A 530bp sequence of TNNC2 gene was obtained by
homologous cloning using the cDNA from the longissimus of Tianfu goat (GenBank accession No. HQ640745). The sequence analysis showed that CDS of TNNC2 was 483bp with 160 encode amino acids residues (Fig. 1) and 18.1kDa predicted molecular weight and 4.06 isoelectric points. The total number of negatively charged residues (Asp + Glu) was 45 and positively charged residues (Arg+ Lys) was 16, and it indicated the protein took negative electricity. There were seven phosphoric sites successfully predicted by the neural network (Fig. 1). There was no signal peptide in the protein. Hydrophobic correlation analysis showed that the minimum was -3.12 and the maximum was 1.18 (Fig. 2). The result indicated that the protein hydrophobicity ability was strong. The secondary structure of the protein was mainly with α-helix, random coil and β-sheet, and the β-sheet only exists in the area of EF-Hand SiteⅠ(Fig. 3)
Figure 1. The Nucleotide and deduced amino acid sequences of TNNC2 from goat. The emerald green indicated initiation codon; *indicated terminal
codon; the red indicated phosphoric sites (Ser, Thr, Tyr)
284

Figure 2. The hydrophobicity profile of TNNC2. The horizontal scale indicated the number of amino acid residues and the vertical one was the relative hydrophobic scale. Points below the zero horizontal line corresponded to hydrophobic region, and points above the line were hydrophilic.
Figure 3. The secondary structure of TNNC2 of goat .The blue line represents alpha helix, the red line represents extended strand, and the purple line represents random coil.
B. Characteristics of deduced protein and phylogenetic analysis of TNNC2
Comparison of TNNC2 protein with fifteen mammals and three other vertebrates (aves, amphibian, fish) revealed that caprine TNNC2 had a high degree of similarity with these reported animals (Fig. 4). The nucleotide similarities of caprine TNNC2 with these animals was varied from 93.38%
(sheep) to 90.48% (alpaca), 85.37%(aves), 79.42%(fish) to 78.66%(amphibian,).The amino acid sequence of goat TNNC2, had an 100% identity with sheep, giant panda and mustang, and 87.73%, 88.34%, 70.81% to aves, amphibian, fish respectively (Table. I).
There were eight missense mutation in mammals, Residue 113 is a Pro in primates (human gorilla and monkey)
285
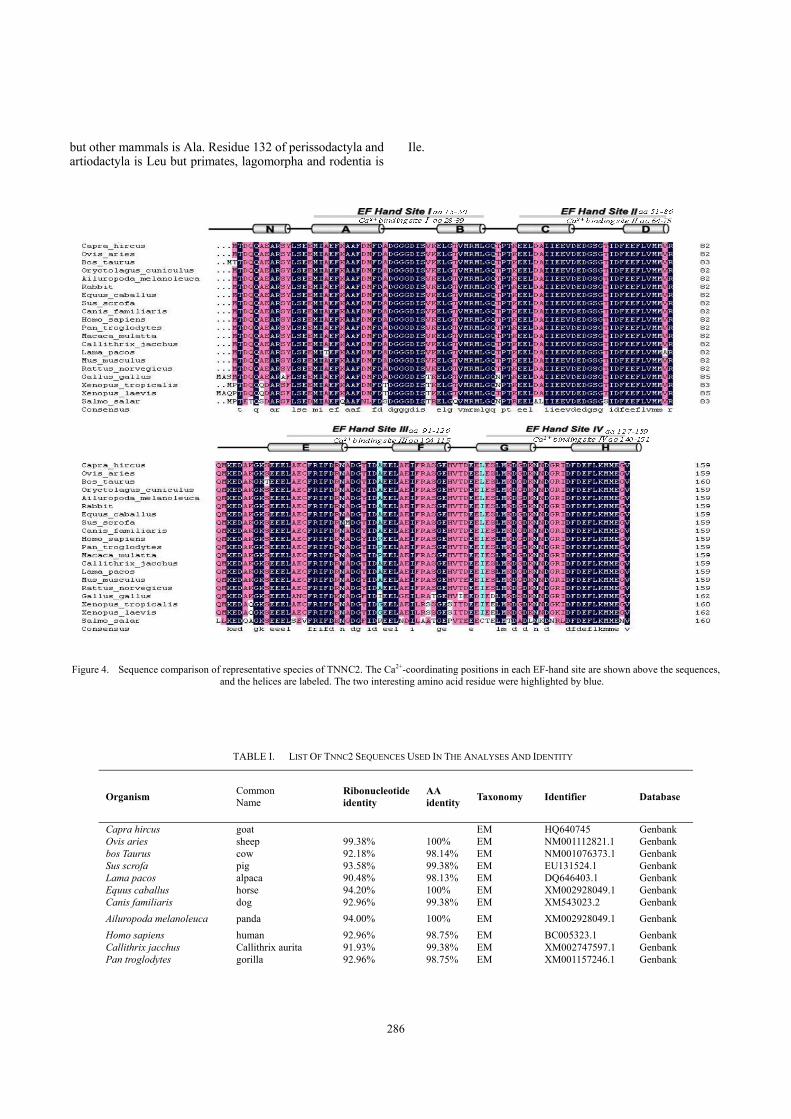
but other mammals is Ala. Residue 132 of perissodactyla and artiodactyla is Leu but primates, lagomorpha and rodentia is
Ile.
Figure 4. Sequence comparison of representative species of TNNC2. The Ca2+-coordinating positions in each EF-hand site are shown above the sequences, and the helices are labeled. The two interesting amino acid residue were highlighted by blue.
TABLE I. LIST OF TNNC2 SEQUENCES USED IN THE ANALYSES AND IDENTITY
Organism Common Name
Ribonucleotideidentity
AA identity Taxonomy Identifier Database
Capra hircus goat EM HQ640745 Genbank Ovis aries sheep 99.38% 100% EM NM001112821.1 Genbank bos Taurus cow 92.18% 98.14% EM NM001076373.1 Genbank Sus scrofa pig 93.58% 99.38% EM EU131524.1 Genbank Lama pacos alpaca 90.48% 98.13% EM DQ646403.1 Genbank Equus caballus horse 94.20% 100% EM XM002928049.1 Genbank Canis familiaris dog 92.96% 99.38% EM XM543023.2 Genbank Ailuropoda melanoleuca panda 94.00% 100% EM XM002928049.1 Genbank Homo sapiens human 92.96% 98.75% EM BC005323.1 Genbank Callithrix jacchus Callithrix aurita 91.93% 99.38% EM XM002747597.1 Genbank Pan troglodytes gorilla 92.96% 98.75% EM XM001157246.1 Genbank
286

Macaca mulatta monkey 92.75% 99.38% EM XM001108623.2 Genbank Rabbit hare 95.45% 99.38% EM Y00760.1 Genbank Oryctolagus cuniculus rabbit 95.03% 99.38% EM NM001082645.1 Genbank Rattus norvegicus rat 90.83% 99.38% EM NM001037351.1 Genbank Mus musculus mouse 89.44% 98.75% EM BC024390.1 Genbank Gallus gallus chicken 85.37% 87.73% bird NM205450.1 Genbank Xenopus tropicalis Western clawed frog 79.42% 88.82% AM BC061286.1 Genbank
Xenopus laevis African clawed frog 78.66% 88.34% AM NM001085939.1 Genbank Salmo salar Atlantic salmon 79.42% 70.81% fish BT049981.2 Genbank
Homo-sapiens
Bos-taurus
Pan-troglodytes
Macac-amulatta
Callithrix-jacchus
Equus-caballus
Ailuropoda-melanoleuca
Canis-familiaris
Sus-scrofa
Rabbit
Oryctolagus-cuniculus
Ovis-aries
Capra-hircus
Mus-musculus
Rattus-norvegicus
Lama-pacos
Gallus-gallus
Xenopus-laevis
Salmo-salar
Xenopus-tropicalis100
100
100
99
100
100
74
98
99
89
84
48
8035
30
14
49
Figure 5. Phylogenic tree of TNNC2. The GenBank accession Nos of the sequences were listed in the table I.
C. The structural model of TNNC2 SWISS-MODEL server, a fully automatic procedure,
was used to construct a 3D structural model. It was constructed using bovine TNNC2 as the template (PDB
accession, 1a2x) (Fig. 6A). And the Ca2+ binding sites were marked out by the SPDBV 4.01 molecular-graphics program (Fig. 6B).
287

(A)
(B) Figure 6. The 3D-structural model of TNNC2. A. The structure of alpha-helix, beta-strand and random-coils. B. The structure was shown in backbone, The
Ca2+ binding sitesⅠ, Ⅱ, Ⅲ, Ⅳ were highlighted by white, blue, red and yellow respectively.
D. Relative Expression pattern of TNNC2 in goat We performed semi-quantitative (RT)-PCR to analyze its
mRNA transcription levels. Results indicated there were some differences in TNNC2 transcription levels between the four developmental stages of the goat (Fig. 7A) The
transcription level of TNNC2 was highest in six-month old goats. The expression amount of TNNC2 was highest in longissimus muscle. There were no expression of TNNC2 in the heart and other Non-muscle tissue (Fig. 7B).
288

(A)
(B)
Figure 7. Expression of TNNC2 gene in goat. The vertica1 axis indicated the value of the TNNC2 mRNA. A. The horizontal axis indicated the various growth stages of TNNC2 in longissimus muscle. B. The horizonta1 axis indicated the various tissues of yearling. Line1-8 was heart, liver, lung,
kidney, gluteus maximus, longissimus muscle, abdominal muscle and biceps brachii, respectively.
IV. DISCUSSION In the study, the cloning, structural analysis and
expression of troponinC2 (TNNC2) gene were completed for the first in fast twitch skeletal muscle of goat. In ruminants, meat tenderness was influenced by the pattern of the animal’s growth, Carcass weight of slow-growing ruminants is lower and more tenderness than that of faster-growing [13].The TNNC2 was considered as a candidate gene with meat quality in our study.
TNNC2 consists of two globular domains connected by a α-helixes linker (fig. 6). Both N-terminal and C-terminal domains contain two EF-hand Ca2+ binding sites. It is similar to other Ca2+ binding proteins, EF-hands consist of two α-helixes which connected by a flexible loop, forming what is known as a helix-loop-helix. The helices and loop in the motif each contain 12 amino acid residues[14-15] This motif is also present in the deduced amino acid sequence of the TNNC2 in goat (Fig. 1) Therefore, we consider that the protein of TNNC2 of goat include four Ca2+ binding sites. Comparison of calcium-binding site residues in the amino
acid sequences of various species indicated that TNNC2 of goat is a member of the EF-hand superfamily (Fig. 2)
The analysis of encoded amino acids revealed that the caprine TNNC2 was highly homologous with that of other species, such as sheep (100%), human (92.96%), aves (87.73%) (table.1). It suggests that there are rigid structure-functional requirements for TNNC2 operating within the mammalian striated muscle.
There were eight amino acids missense mutation in 16 species of mammals. All of this missense mutation is conservative substitution except Thr18 in alpaca. Alpaca live on in highlands of South America, with an average altitude above 4, 000 meters[16]. As Thr is a polar, hydrophilic residue and Ala is a nonpolar, hydrophobic residue, this represents a non-conservative properties substitution. The replacement of a hydrophilic residue (Thr) with a hydrophobic residue (Ala) at position 18,and the presence of a hydrophobic residue at position 17 (Ile),represent an adaptation of TNNC2 functioning of alpaca to circumstances.
289

The Pro113 present only to primates (human,gorilla and monkey). The replacement may play a key role in the evolution of primates. The Leu or Ile at 132 locus distribute regularly in the 6 types of mammals. Moreover, Avian and ectothermic species (Xenopus laevis and Xenopus tropicalis) is also Ile at 132 locus. In other words, Leu exist only in perissodactyla and Artiodactyla animals, and these may represent an adaptation to the characteristics of horned animals.
Binding of Ca2+ to the low-affinity sites of TNNC (particularly Ca2+binding site II, which is conserved in TNNC1 and TNNC2) is the trigger event for contraction[17]. This hypothesis is supported by the finding that TNNC1, which contains only a single low-affinity Ca2+ binding site (site II), can regulate fast skeletal muscle contraction in response to Ca2+ after substitution of TNNC1 for TNNC2 in permeabilized fast skeletal muscle fibers[18]. Compared to siteⅠ, Ⅲ and Ⅳ, the sequence of site II of TNNC2 is the most conservative, only a conservative substitution Ser70in fish in the 20 species(fig. 4) as Thr and Ser both belong to polar and hydrophilic amino acid residues. Therefore, it will specify the importance of site II further.
We know that there are three distinct sites of the TNNC2 molecule and make important contacts with TNI [19-26]. The regions of TNNC2 (AA 50-60 and AA 90-100) interacts with TNI (AA 104-115). This binding requires Ca2+ or Mg2+ occupancy of the high-affinity COOH-terminal Ca2+ binding sites. and a peptide (comprising AA 126-136) of TNNC2 interacts with TNI in a Ca2+ independent manner[17]. We found the amino acid is identical of TNNC2 (AA 50-60 and AA 90-100) in mammals, Aves and amphibian. For all mammalian, amphibian and fish, amino acid residue at 128 locus is a Thr, but that is Ile in Aves. As Ile is a nonpolar, hydrophobicity residue and Thr is a polar, hydrophilic residue, this represents a non-conservative properties substitution. This substitution may play a key role in the evolution of birds flying.
The cross-linking studies have also examined the interaction of TNNC2 with TNT in either heterodimeric complexes or trimeric complexes that also contained TNI[27]. These studies demonstrated an interaction between Cys98 of TNNC (located in the COOH-terminal half of the molecule) and residues 175-178 of TNT (Met-Lys-Lys-Lys)[28]. Residue 98 is a Cys in all mammalian, amphibian and avian TNNC2 but is a Val in fish (Fig. 6). This may be the key that the terrestrial animal adapt to the movement on land,what is more, it illustrate the importance of touch the residue 98 with TNT.
Initial immunohistochemical studies showed that TNNC1 only in cardiac and slow skeletal muscle, whereas, TNNC2 was found exclusively in fast skeletal myocytes[9, 24, 29]. Research suggested that a TNC-like protein was present in chick embryo brain[30]. Normally, the TNNC2 gene is not expressed in ventricular myocytes. But inappropriate expression of TNNC2 gene may be appeared in cardiac muscle in some pathological state, and might be related to the clinical[31]. In invertebrates, TNNC is expressed in a variety of non-muscle tissues[32]. Semi-(RT)-PCR analysis
has indicated that expression of the porcine gene is restricted to skeletal muscle (biceps brachii, quadriceps femoris, longissimus dorsi)[33]. In our study, we have also found that TNNC2 of goat was expressed in gluteus maximus, longissimus muscle, abdominal muscle and biceps brachii. This result implied that the TNC gene may be an excellent model system for elucidating the molecular mechanisms that regulate skeletal versus cardiac muscle-specific gene expression.
V. CONCLUSION In the present study, the full-length cDNA of TNNC2
gene was cloned with analyzed the amino acids sequence and space structure in goats. We found that the TNNC2 gene of goat had high degree of identity with other mammals. By comparing the sequences of TNNC2, we have discovered a number of amino acid residues which may have a critical impact for their functional studies. The TNNC2 showed a high level of expression in skeletal muscle of goat. The results of expression profiles of TNNC2 gene will help to explain molecular mechanisms of regulating skeletal and development.
ACKNOWLEDGMENT This work was financially supported by the National
Modern Industry Technology System on Sheep and Goat for Meat (NYCYTX-39) and the Projects of Selecting New Breed on Livestock in Sichuan Province (01NG002-06). Research Instistute of Animal Genetics&Breeding, College of Animal Science & Technology of Sichuan Agricultural University, 46# Xinkang Road, Yucheng district, Yaan 625014, Sichuan province of China. Tel: +86 (0835) 288-6096; Fax: +86 (0835) 288-6080; E-mail: goatsxgy @ 263.net.
REFERENCE [1] R. Kretsinger, "Structure and evolution of calcium modulated
proteins. CRC Crit," Rev. Biochem, vol. 8, pp. 119-174, 1980. [2] J. Johnson, et al., "A fluorescent probe study of Ca2+ binding to the
Ca2+-specific sites of cardiac troponin and troponin C," Journal of Biological Chemistry, vol. 255, p. 9635, 1980.
[3] A. Babu, et al., "Determination of residue specificity in the EF-hand of troponin C for Ca2+ coordination, by genetic engineering," Journal of Biological Chemistry, vol. 267, p. 15469, 1992.
[4] S. Fonseca, et al., "Slow fiber cluster pattern in pig longissimus thoracis muscle: implications for myogenesis," Journal of animal science, vol. 81, p. 973, 2003.
[5] C. Farah and F. Reinach, "The troponin complex and regulation of muscle contraction," The FASEB Journal, vol. 9, p. 755, 1995.
[6] E. Bucher, et al., "Expression of the troponin complex genes: transcriptional coactivation during myoblast differentiation and independent control in heart and skeletal muscles," Molecular and cellular biology, vol. 8, p. 4134, 1988.
[7] Q. XU, et al., "Characterization of the fast skeletal troponin C (TNNC2) gene in three Chinese native sheep breeds."
[8] M. Te Pas and A. Soumillion, "Improvement of livestock breeding strategies using physiologic and functional genomic information of the muscle regulatory factors gene family for skeletal muscle development," Current Genomics, vol. 2, pp. 285-304, 2001.
290

[9] M. Parmacek, et al., "The structure and regulation of expression of the murine fast skeletal troponin C gene. Identification of a developmentally regulated, muscle-specific transcriptional enhancer," Journal of Biological Chemistry, vol. 265, p. 15970, 1990.
[10] D. Wang, et al., "Characteristics and production performance of Tianfu goat, a new breed population," Small Ruminant Research, 2010.
[11] T. Schwede, et al., "SWISS-MODEL: an automated protein homology-modeling server," Nucleic Acids Research, vol. 31, p. 3381, 2003.
[12] K. Livak and T. Schmittgen, "Analysis of relative gene expression data using real-time quantitative PCR and the 2-[Delta][Delta] CT method," Methods, vol. 25, pp. 402-408, 2001.
[13] G. Harper, "Trends in skeletal muscle biology and the understanding of toughness in beef," Australian Journal of Agricultural Research, vol. 50, pp. 1105-1129.
[14] H. Kawasaki and R. Kretsinger, "Ca 2+-binding proteins 1: EF-hands," Protein Profile, vol. 1, pp. 343-517, 1995.
[15] S. Nakayama, et al., "Evolution of EF-hand calcium-modulated proteins. II. Domains of several subfamilies have diverse evolutionary histories," Journal of Molecular Evolution, vol. 34, pp. 416-448, 1992.
[16] C. Novoa and J. Wheeler, "Llamas and Alpacas In: Evolution of domesticated Animals (IL Mason ed.) pp. 116¨C128," ed: Longman London, 1984.
[17] T. Gillis, et al., "Functional and evolutionary relationships of troponin C," Physiological genomics, vol. 32, p. 16, 2007.
[18] R. Moss, et al., "Altered Ca2+ dependence of tension development in skinned skeletal muscle fibers following modification of troponin by partial substitution with cardiac troponin C," Journal of Biological Chemistry, vol. 261, p. 6096, 1986.
[19] P. Cachia, et al., "Calmodulin and troponin C: a comparative study of the interaction of mastoparan and troponin I inhibitory peptide [104-115]," Biochemistry, vol. 25, pp. 3553-3562, 1986.
[20] Z. Grabarek, et al., "Proteolytic fragments of troponin C. Interactions with the other troponin subunits and biological activity," Journal of Biological Chemistry, vol. 256, p. 13121, 1981.
[21] R. Grand, et al., "Proton-magnetic-resonance studies on the interaction of rabbit skeletal-muscle troponin I with troponin C and actin," Biochemical Journal, vol. 203, p. 61, 1982.
[22] J. Lan, et al., "Interactions of troponin I and its inhibitory fragment (residues 104-115) with troponin C and calmodulin," Biochemistry, vol. 28, pp. 7380-7385, 1989.
[23] J. Leszyk, et al., "Crosslinking of rabbit skeletal muscle troponin with the photoactive reagent 4-maleimidobenzophenone: identification of residues in troponin I that are close to cysteine-98 of troponin C," Biochemistry, vol. 26, pp. 7042-7047, 1987.
[24] H. Syska, et al., "The relationship between biological activity and primary structure of troponin I from white skeletal muscle of the rabbit," Biochemical Journal, vol. 153, p. 375, 1976.
[25] T. Tao, et al., "Calcium dependence of the distance between Cys-98 of troponin C and Cys-133 of troponin I in the ternary troponin complex. Resonance energy transfer measurements," Biochemistry, vol. 28, pp. 5902-5908, 1989.
[26] J. Van Eyk and R. Hodges, "The biological importance of each amino acid residue of the troponin I inhibitory sequence 104-115 in the interaction with troponin C and tropomyosin-actin," Journal of Biological Chemistry, vol. 263, p. 1726, 1988.
[27] J. Leszyk, et al., "Characterization of zero-length cross-links between rabbit skeletal muscle troponin C and troponin I: evidence for direct interaction between the inhibitory region of troponin I and the amino-terminal, regulatory domain of troponin C," Biochemistry, vol. 29, pp. 299-304, 1990.
[28] J. Leszyk, et al., "Cross-linking of rabbit skeletal muscle troponin subunits: labeling of cysteine-98 of troponin C with 4-maleimidobenzophenone and analysis of products formed in the
binary complex with troponin T and the ternary complex with troponins I and T," Biochemistry, vol. 27, pp. 6983-6987, 1988.
[29] G. Dhoot, et al., "Polymorphic forms of troponin T and troponin C and their localization in striated muscle cell types* 1," Experimental Cell Research, vol. 122, pp. 339-350, 1979.
[30] R. Fine, et al., "Troponin C in brain," 1975. [31] A. Babu, et al., "Molecular basis for the influence of muscle length on
myocardial performance," Science, vol. 240, p. 74, 1988. [32] J. Chen, et al., "Molecular characterization and immunohistochemical
localization of a novel troponin C during silkworm development," Cell and Tissue Research, vol. 331, pp. 725-738, 2008.
[33] X. Tian, et al., "Cloning and expression of the pig fast skeletal muscle troponin C gene," Yi chuan= Hereditas/Zhongguo yi chuan xue hui bian ji, vol. 28, p. 949, 2006.
291





























