Embed Size (px)
Citation preview

Aquaculture 242 (2004) 137–155
www.elsevier.com/locate/aqua-online
Classification of jinjiang oysters
Crassostrea rivularis (Gould, 1861) from China,
based on morphology and phylogenetic analysis
Haiyan Wanga,b, Ximing Guoa,c,*, Guofan Zhanga, Fusui Zhanga
aInstitute of Oceanology, Chinese Academy of Sciences, Shandong 266071, PR ChinabThe Graduate School of Chinese Academy of Sciences, Beijing, PR China
cHaskin Shellfish Research Laboratory, Institute of Marine and Coastal Sciences,
Rutgers University, NJ 08349, USA
Received 25 May 2004; received in revised form 10 September 2004; accepted 21 September 2004
Abstract
The jinjiang oyster Crassostrea rivularis [Gould, 1861. Descriptions of shells collected in the
North Pacific Exploring Expedition under Captains Ringgold and Rodgers. Proc. Boston Soc. Nat.
Hist. 8 (April), 33-40] is one of the most important and best-known oysters in China. Based on the
color of its flesh, two forms of C. rivularis are recognized and referred to as the bwhite meatQ andbred meatQ oysters. The classification of white and red forms of this species has been a subject of
confusion and debate in China. To clarify the taxonomic status of the two forms of C. rivularis, we
collected and analyzed oysters from five locations along China’s coast using both morphological
characters and DNA sequences from mitochondrial 16S rRNA and cytochrome oxidase I, and the
nuclear 28S rRNA genes. Oysters were classified as white or red forms according to their
morphological characteristics and then subjected to DNA sequencing. Both morphological and DNA
sequence data suggest that the red and white oysters are two separate species. Phylogenetic analysis
of DNA sequences obtained in this study and existing sequences of reference species show that the
red oyster is the same species as C. ariakensis Wakiya [1929. Japanese food oysters. Jpn. J. Zool. 2,
0044-8486/$ -
doi:10.1016/j.
* Corresp
Rutgers Unive
fax: +1 856 7
E-mail add
see front matter D 2004 Elsevier B.V. All rights reserved.
aquaculture.2004.09.014
onding author. Haskin Shellfish Research Laboratory, Institute of Marine and Coastal Sciences,
rsity, 6959 Miller Avenue, Port Norris, NJ 08349, USA. Tel.: +1 856 785 0074;
85 1544.
ress: [email protected] (X. Guo).

H. Wang et al. / Aquaculture 242 (2004) 137–155138
359–367.], albeit the red oysters from north and south China are genetically distinctive. The white
oyster is the same species as a newly described species from Hong Kong, C. hongkongensis Lam and
Morton [2003. Mitochondrial DNA and identification of a new species of Crassostrea (Bivalvia:
Ostreidae) cultured for centuries in the Pearl River Delta, Hong Kong, China. Aqua. 228, 1–13].
Although the name C. rivularis has seniority over C. ariakensis and C. hongkongensis, the original
description of Ostrea rivularis by Gould [1861] does not fit shell characteristics of either the red or
the white oysters. We propose that the name of C. rivularis Gould [1861] should be suspended, the
red oyster should take the name C. ariakensis, and the white oyster should take the name C.
hongkongensis.
D 2004 Elsevier B.V. All rights reserved.
Keywords: Cytochrome oxidase I; 16S rRNA; 28S rRNA; Crassostrea rivularis; C. ariakensis; C.
hongkongensis; Taxonomy
1. Introduction
Oyster classification remains problematic and taxonomic status for many species has not
been satisfactorily resolved. The problem is most pronounced in China, where diverse
groups of oysters exist. According to various records, there are 15–24 species of Ostreidae
naturally distributed along the coast of China. While most of the species are well described,
there are several major confusions and disagreements. One of the problems is the
taxonomic status of jinjiang oyster Crassostrea (Ostrea) rivularisGould (1861) (Li et al.,
1988; Li, 1989; Li and Qi, 1994; Zhou and Allen, 2003).
Jinjiang oyster is probably the most common and best-known oyster in China (Wang
et al., 1993). It has been cultured in China for centuries. It is widely distributed, ranging
from the Lizijiang (oyster river) on the border with Korea in the north (Zhang and Lou,
1956) to Beihai near the border with Vietnam (Guo et al., 1999; Zhou and Allen, 2003). It
is mostly found in rivers and estuaries with salinities of 10–25 ppt. It is one of most
important oyster species cultured in China (Guo et al., 1999). C. rivularis was first
described by Gould (1861) based on a specimen collected from southern China (Johnson,
1964) and subsequently used to describe oysters in Japan and China (Hirase, 1930; Zhang
and Lou, 1956). In China, the name jinjiang (close to river) oyster or C. rivularis has been
consistently used in all taxonomic literature (Li and Qi, 1994). However, two forms of C.
rivularis are recognized in China, initially based on meat color, one is referred to as bredmeatQ and the other as bwhite meatQ. The two oysters are similar in shell morphology and
sympatric in southern China. Oyster farmers prefer the white oyster because of its flavor,
fast growth and high market price. Anatomic and allozyme analyses have demonstrated that
they are two different species, but there are disagreements about which one is C. rivularis
(Li et al., 1988; Li and Qi, 1994). In Japan, the name C. rivularis was rejected and C.
ariakensis (Wakiya, 1929) was used to describe an oyster from the Ariake Bay (Torigoe,
1981). At the present time, the relationship among the red and white oysters from China and
C. ariakensis from Japan is not clear. C. ariakensis is being considered for possible
introduction to Chesapeake Bay in the United States, where the native oyster (C. virginica
Gmelin) has been devastated by two diseases: MSX and Dermo (Mackenzie et al., 1997). C.

H. Wang et al. / Aquaculture 242 (2004) 137–155 139
ariakensis apparently resists both diseases of the native oyster and grows well in
Chesapeake Bay (Calvo et al., 2001). The introduction of a non-native species is highly
controversial and environmental impact studies are being conducted. The clarification on
taxonomic status of C. rivularis in relation to C. ariakensis is important to our
understanding of the distribution and ecology of C. ariakensis, and relevant to the
proposed introduction.
Difficulties in oyster classification have partly rooted in the lack of well-defined
morphological characters. Shell morphology, the main character used in oyster
classification, is known to be plastic and subject to environmental variation. Anatomic
studies are technically challenging and not always possible. Phylogenetic analysis of
molecular data has shown great promise in oyster classification (Banks et al., 1993;
Littlewood, 1994; O Foighil et al., 1995, 1998; O Foighil and Taylor, 2000; Lapegue et
al., 2002). However, phylogenetic analysis alone cannot resolve disputes that originated
from morphological characterization. The combination of the two approaches is needed
to resolve some of the confusions in oyster classification. In this study, we focused on
the taxonomic status of two forms of C. rivularis from China, using both morphological
data and phylogenetic analysis of DNA sequences from mitochondrial 16S rRNA gene
and cytochrome oxidase I (COI), and the nuclear 28S rRNA genes. Here, we provide
evidence that the bred meatQ of C. rivularis is the same species as C. ariakensis from
Japan, and the bwhite meatQ is the same as a newly described species from Hong Kong
C. hongkongensis (Lam and Morton, 2003).
2. Materials and methods
2.1. Sample collection and morphological analyses
Oysters were collected from five typical habitats of Crassostrea rivularis (Gould, 1861)
in China: Weifang in Bohai Sea in the north, and Shenzhen, Yangjiang, Zhanjiang and
Beihai in the south (Fig. 1). At Weifang, oysters were collected by divers at a depth of about
eight meters. Oysters from Shenzhen were obtained from a local market and they were
cultured oysters using natural seed. Oysters from Zhanjiang and Yangjiang were collected
from intertidal zones. Oysters from Beihai were collected from an oyster farm that uses
natural seed. Crassostrea sikamea (Amemiya, 1928) was also used for comparisons.
Specimens for C. sikamea were from a stock maintained at Rutgers University, which
originated from Japan, via introduction to Washington State.
Collected oysters were first separated into bred meatQ or bwhite meatQ according to
morphological characters (Li, 1989; Li and Qi, 1994). The following shell characteristics
were recorded and measured: shell height, shell length, shell width, ligament channel (LC)
length and width, shell dry weight, shell volume and shell cavity volume. Shell parameters
were defined following definitions of Moore et al. (1971). Size measurements were made
to the nearest 0.1 mm using a vernier caliper. Shell volume was measured by the amount of
water that the shells displaced. Shell cavity volume was determined by measuring the
volume of the sand needed to fill the cavity. Shell and LC shape were characterized with a
height to length ratio. Shell density (dry shell weight/shell volume) was also calculated for

Fig. 1. A map of China’s coast showing sites of oyster collection and sites referenced.
H. Wang et al. / Aquaculture 242 (2004) 137–155140
each oyster. Differences between red and white meats in shell measurements were tested
by two-sample t-test.
2.2. DNA extraction and PCR amplification and sequencing
DNAwas extracted from fresh adductor muscle tissue by a phenol/chloroform method as
described by Moore (1993). Primers (Invitrogen, USA) of 16sar and 16sbr (Palumbi et al.,
1991) were used to amplify a segment of the mitochondrial 16S rRNA gene. A segment of
cytochrome oxidase subunit I (COI) was amplified using LCO1490 and HCO2198 primers

H. Wang et al. / Aquaculture 242 (2004) 137–155 141
(Folmer et al., 1994). A 28S ribosomal fragment was amplified using primers (D1F and
D6R) as described by Park and O Foighil (2000). PCR amplification was performed using
a Biometra T1 thermal cycler. Reactions were performed in 50 Al with final
concentrations of: 2.0 mM MgCl2, 150 AM of each dNTP, 0.2 AM each primer, 20 ng
of template DNA, 2 units of Taq polymerase (Promega, USA) in 5 Al of 10� buffer. COI
and 16S were amplified using the following protocol: initial denature at 95 8C for 2 min,
30 cycles of 95 8C for 1 min, 51 8C (COI) or 57 8C (16S) for 1 min and 72 8C for 1 min,
with a final extension at 72 8C for 5 min. The 28S fragment was amplified by an initial
denaturing at 94 8C for 4 min, addition of Taq polymerase, 30 cycles of denaturing at 94
8C for 40 s, annealing at 60 8C for 40 s and extension at 72 8C for 1.25 min, plus a final
extension at 72 8C for 10 min. A negative control (no template) was included during each
PCR run.
PCR products were purified using EZ Spin Column PCR Product Purification Kit (BBI,
Canada). Sequencing was performed in both directions on an ABI PRISM 377XL DNA
Sequencer using the ABI PRISM BigDyek Terminator Cycle Sequencing Ready
Reaction Kit (Perkin-Elmer, USA).
2.3. Phylogenetic analysis
The COI, 16S and 28S sequences obtained in this study and those of other Crassostrea,
Ostrea and Saccostrea species available from GenBank were subjected to phylogenetic
analysis. Initial alignments were performed using CLUSTALW (Tompson et al., 1994). The
sequences were trimmed to the same length as other published sequences after alignment.
Maximum parsimony (MP) and neighbor-joining (NJ) analyses was made using PHYLIP
(Ver. 3.56C, Felsenstein, 1989). Bootstrap analysis with 1000 replications was performed
using the SEQBOOT and CONSENSE programs. Consensus phylogenetic trees were
drawn with DRAWGRAM program in the PHYLIP package. Pair-wise sequence
divergences among haplotypes and reference species were calculated using the DNADIST
program of PHYLIP according to Kimura’s two-parameter model (Kimura, 1980).
3. Results
3.1. Distribution and morphology of red and white C. rivularis
All oysters collected in this study were unambiguously classified as red or white meat
according to morphological characters defined by Li (1989). Among the 16 oysters
collected from Beihai, 7 were red oysters and nine were white (Table 1). Sixteen oysters
were collected from Zhanjiang, and three of them were red and the other 13 were white.
Among 52 oysters from Shenzhen, 3 were red and the other 49 were white. Six oysters,
three red and three white, were obtained from Yangjiang for sequencing, but they were not
from a random sampling of oysters from the field. For the northern China site, however, all
19 oysters collected from Weifang were red meats.
The structural relationship among the gill, soft-tissue, mantle and the promyal chamber
provided the most useful character in separating red and white oysters. In the bwhite

Table 1
Number of C. rivularis collected, number of red and white types, number of oysters measured and sequenced at
five locations
Locations Total Red White Measured Sequenced
BH: Beihai, Guangxi province 16 9 7 16 3 red, 3 white
ZJ: Zhanjiang, Guangdong province 16 3 13 14 3 red, 3 white
SZ: Shenzhen, Guangdong province 52 3 49 52 3 red, 3 white
YJ: Yangjiang, Guangdong province 6 3 3 0 3 red, 3 white
WF: Weifang, Shandong province 19 19 0 19 6 red
H. Wang et al. / Aquaculture 242 (2004) 137–155142
meatsQ, the central part of the gill was detached from the soft tissue and both epibranchial
chambers are open to the promyal chamber (Fig. 2A). After the removal of the mantle, four
rows of gill water tubes were visible. In the bred meatsQ, the gill is attached to the soft
tissue and only the right epibranchial chamber is open to the promyal chamber. Thus,
when the right mantle is lifted, only two rows of gill water tubes were exposed (Fig. 2D).
Fig. 2. Morphological characteristics of red and white forms of C. rivularis. (A) Separation of both gills from
viscera in the white oyster; (B) external shell morphology of the white oyster; (C) internal shell morphology of the
white oyster; (D) opening of the right epibranchial chamber to the promyal chamber in the red oyster; (E) external
shell morphology of the red oyster from southern China; (F) internal shell morphology of the red oyster from
southern China; (G) external shell view of the red oyster from northern China (Weifang); (H) external shell view
of C. ariakensis from Japan (Okutani, 2000); (I) internal shell view of the red oyster from northern China
(Weifang). PC, promyal chamber; GWT, gill water tubes.

H. Wang et al. / Aquaculture 242 (2004) 137–155 143
External characteristics of the shells are somewhat variable, with some common
features and noticeable differences between the two forms. Both red and white oysters
were large and of approximately the same size in the mixed populations. They both had
deeply cupped left valves and slightly convex right valves. The left valves were larger and
thicker than the right valve. Right valves were relatively smooth with multiple layers of
overlapping, concentric and brittle lamellae. No radiant ribs were protruding from the
surface of either valve (in contrast to Gould’s description of Ostrea rivularis). Color of the
right valve was variable in both red and white oysters, ranging from white, pale, gray,
yellow or yellowish brown to purple. Lamellae closer to the edge were usually dark in
color, and those near the umbo were eroded and chalky white. The umbo and ligament
channel were obvious. Internal side of shells was white with D- or kidney-shaped adductor
muscle scar.
There were also some noticeable differences in shell characteristics between the red and
white oysters. One of the most obvious was that the red oysters were generally round or
oval-shaped, while the white oysters were more elongated (Fig. 2B,C,E,F). Another
difference was in the size or shape of the umbo or ligament channel. The white oysters had
noticeably large umbo regions and longer ligament channels than the red oysters (Fig.
2B,C,E,F,G,I). The red oysters had a yellow or brown shell edge, while the white oysters
did not. The red oysters had more lamellae on the right valve than the white oysters. There
were some differences in shell color between northern and southern oysters, which might
be genetic or environmental. Red and white oysters from southern China (BH, ZJ, YJ and
SZ) had dark or purple muscle scars, while red oysters from north China (WF) all had
white muscle scars (Fig. 2C,F,I). The red oysters from northern China had yellowish
brown lamellae on the left valves, while red and white oysters from the south had some
green patches on the right valves.
Statistical analysis of shell measurements provided confirmation of general
observations on shell morphology in the red and white oysters. Oysters sampled from
Beihai, which was the only population that contained about equal numbers of the red
and white oysters, provided a direct comparison of two types from the same
environment and age (because they were from the same spat collection). The red and
white oysters from Beihai had similar shell height and width, but significantly (P=0.001)
different shell length (Table 2). Difference in shell shape was also indicated by shell
height/length (SH/SL) ratio: the red oysters had a significantly (P=0.000) lower SH/SL
ratio than the white oysters (1.4 vs. 2.0), suggesting a rounder shell shape for the red
oyster. Another major difference is the significantly longer ligamental channel (P=0.004)
and larger LCH/LCL ratio (P=0.000) in the white than the red oysters. The white oysters
had deeper umbo cavities than the red oysters (Table 2). Shells of the red oysters were
bigger and heavier than that of the white oysters, but both had exactly the same density.
The combination of all red and white oysters from the four populations provided the
same pattern of differences in shell measurements, providing stronger support of the
differences observed (Table 2).
The largest white oyster observed was a specimen from Shenzhen, which measured
378�299�271 mm (H�L�W). The largest red oyster was from Weifang and measured
250�160�55 mm. The smallest white and red oysters, both from Zhanjiang, measured
75�44�28 and 81�68�28 mm, respectively.

Table 2
Shell measurements (means) and p-values from two-sample test of red and white forms of C. rivularis collected
from Beihai and all four sites (Beihai, Shenzhen, Zhanjiang and Weifang): character names were abbreviated as
SH for shell height, SL for shell length, SW for shell width, SH/SL for shell height/length ratio, LCH for
alimental channel height, LCL for ligamental channel length, LCH/LCL for ligamental channel height/length
ratio, UCD for umbo cavity depth, SCV for shell cavity volume, DSWT for dry shell weight, SV for shell volume
and SD for shell density
Site
form
Sample
size
SH
(mm)
SL
(mm)
SW
(mm)
SH/
SL
LCH
(mm)
LCL
(mm)
LCH/
LCL
UCD
(mm)
SCV
(ml)
DSWT
(g)
SV
(ml)
SD
Beihai
Red 7 99.2 71.1 37.2 1.4 18.0 28.3 0.65 8.8 51.1 123.9 64.4 2.06
White 9 107.1 54.3 33.7 2.0 29.3 24.1 1.22 12.9 37.6 91.0 47.1 2.05
p-value 0.236 0.001 0.289 0.000 0.004 0.081 0.000 0.010 0.019 0.043 0.139 0.956
All
Red 32 157.7 95.4 41.1 1.66 22.2 32.2 0.70 5.8 139.6 376.5 195.7 2.01
White 69 150.8 79.3 49.5 1.98 33.4 27.5 1.23 13.6 97.6 242.2 137.9 2.00
p-value 0.515 0.009 0.072 0.001 0.000 0.001 0.000 0.000 0.007 0.005 0.020 0.933
H. Wang et al. / Aquaculture 242 (2004) 137–155144
3.2. Phylogenetic analysis
3.2.1. Mitochondrial 16S rRNA sequence
A 480-bp fragment of the mitochondrial 16S rRNA gene was sequenced for 30
individuals collected from 5 sites (Table 1). Four haplotypes were identified among the 30
16S sequences. All white oysters from the four southern sites shared a common haplotype
(White A), which is identical in sequence as C. hongkongensis. All 12 red oysters from the
Fig. 3. Phylogenetic trees based on neighbor-joining (A) and parsimony (B) analyses of partial mt16S sequence
using Kimura two-parameter distances with 1000 bootstrap. O. aupouria was used as an outgroup. Numbers at
steps are bootstrap values that are N50.
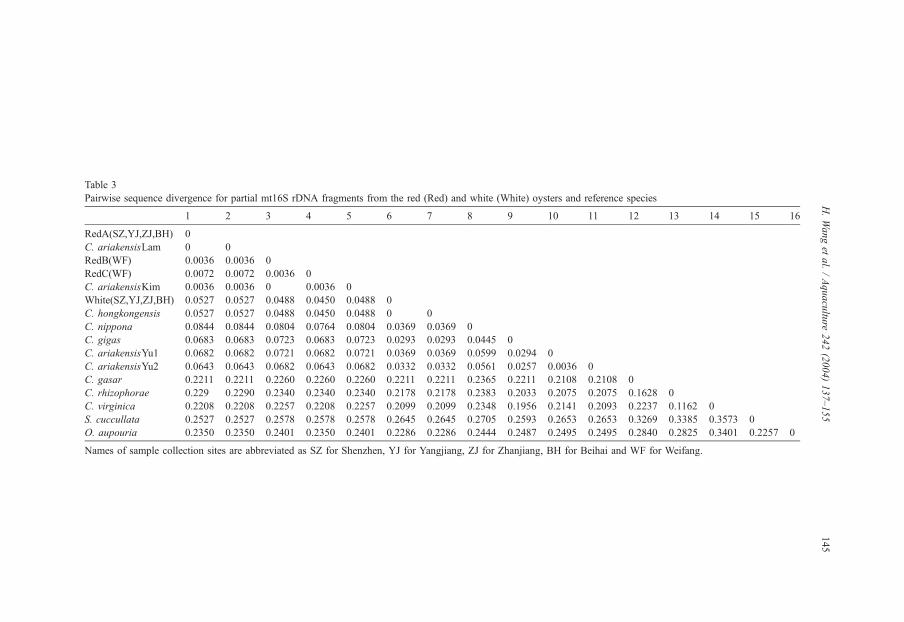
Table 3
Pairwise sequence divergence for partial mt16S rDNA fragments from the red (Red) and white (White) oysters and reference species
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
RedA(SZ,YJ,ZJ,BH) 0
C. ariakensisLam 0 0
RedB(WF) 0.0036 0.0036 0
RedC(WF) 0.0072 0.0072 0.0036 0
C. ariakensisKim 0.0036 0.0036 0 0.0036 0
White(SZ,YJ,ZJ,BH) 0.0527 0.0527 0.0488 0.0450 0.0488 0
C. hongkongensis 0.0527 0.0527 0.0488 0.0450 0.0488 0 0
C. nippona 0.0844 0.0844 0.0804 0.0764 0.0804 0.0369 0.0369 0
C. gigas 0.0683 0.0683 0.0723 0.0683 0.0723 0.0293 0.0293 0.0445 0
C. ariakensisYu1 0.0682 0.0682 0.0721 0.0682 0.0721 0.0369 0.0369 0.0599 0.0294 0
C. ariakensisYu2 0.0643 0.0643 0.0682 0.0643 0.0682 0.0332 0.0332 0.0561 0.0257 0.0036 0
C. gasar 0.2211 0.2211 0.2260 0.2260 0.2260 0.2211 0.2211 0.2365 0.2211 0.2108 0.2108 0
C. rhizophorae 0.229 0.2290 0.2340 0.2340 0.2340 0.2178 0.2178 0.2383 0.2033 0.2075 0.2075 0.1628 0
C. virginica 0.2208 0.2208 0.2257 0.2208 0.2257 0.2099 0.2099 0.2348 0.1956 0.2141 0.2093 0.2237 0.1162 0
S. cuccullata 0.2527 0.2527 0.2578 0.2578 0.2578 0.2645 0.2645 0.2705 0.2593 0.2653 0.2653 0.3269 0.3385 0.3573 0
O. aupouria 0.2350 0.2350 0.2401 0.2350 0.2401 0.2286 0.2286 0.2444 0.2487 0.2495 0.2495 0.2840 0.2825 0.3401 0.2257 0
Names of sample collection sites are abbreviated as SZ for Shenzhen, YJ for Yangjiang, ZJ for Zhanjiang, BH for Beihai and WF for Weifang.
H.Wanget
al./Aquacultu
re242(2004)137–155
145

H. Wang et al. / Aquaculture 242 (2004) 137–155146
4 southern sites had the same haplotype (RedA), and the 6 red oysters from the northern
site (Weifang) had 2 different haplotypes, RedB (5) and RedC (1).
Phylogenetic analysis was conducted using the four haplotypes identified in this study
and the following sequences from GenBank/reference:C. ariakensis (LamAY160757; Kim
AY007427; Yu1 and Yu2, Yu et al., 2003), C. nippona (AY007426), C. hongkongensis
(AY160756), C. gigas (S66183), C. gasar (AJ312937), C. virginica (AF092285), C.
rhizophorae (AJ312938), S. cuccullata (AF458901) and O. aupouria (AF052064) as an
outgroup. Including the outgroup, 34 nucleotide positions were variable in the 16S data
set. Phylogenetic analysis using NJ and MP procedures produced almost identical results
(Fig. 3A,B). As expected, all four haplotypes identified in this study were clustered with
members of Crassostrea, and away from S. cuccullata and O. aupouria. The white oysters
and C. hongkongensis, as they had the same haplotype, formed one clade. The three
haplotypes for red oysters were closely clustered with C. ariakensis of Lam and Kim. The
C. ariakensis haplotypes from Yu et al. (2003) clustered with C. gigas. A closer
examination indicated that the RedA haplotype from southern China was the same as C.
ariakensis of Lam (Hong Kong), and RedB from the north is the same as C. ariakensis of
Kim (Korea).
Sequence divergence between the red and white haplotypes was about 5%, which was
higher than that between the two known species of Crassostrea–C. gigas and C. nippona
(Table 3). There was no divergence between the white and C. hongkongensis, suggesting
that they are the same species. Also, no divergence was observed between RedA and C.
ariakensis Lam, and between RedB and C. ariakensis Kim. Divergence among all red and
C. ariakensis haplotypes was less than 1%, with the exception of C. ariakensis of Yu et al.
(2003).
3.2.2. Mitochondrial COI DNA sequences and AA sequences
A 640-bp fragment of the COI sequences was sequenced for the 30 oysters selected in
this study, generating 11 haplotypes. Three of these haplotypes occurred in white oysters
and were designated with White A, B and C. Of the 12 white oysters sequenced, 10 oysters
had haplotype WhiteA, 1 oyster had WhiteB and 1 had WhiteC. Eight of the 11 haplotypes
were observed in the red oysters: RedA, B, C, D and E were found in southern red oysters
from Shenzhen, Yangjiang, Zhanjiang and Beihai; and RedF, G and H in red oysters from
the north site (Weifang). For red oysters from southern sites, eight had haplotype RedA
and only one oyster each had haplotypes RedB, C, D and E. Haplotype RedF was shared
by four red oysters from the north, and RedG and H were represented by one oyster each.
The partial COI sequences from the 11 haplotypes obtained in this study were subjected
to phylogenetic analysis along with the following sequences from GenBank: C. ariakensis
(O’Foghil AF152569; Kim AF300617; Lam AY160752, AY160753, AY160754, Yu et al.,
2003), C. hongkongensis (AY160746–AY160751), C. gigas (AF152565), C. virginica
(AF152566), C. iredalei (AY038078), C. belcheri (AY038077), C. nippona (AF300616),
C. sikamea (AF152568), C. angulata (AF152567), S. cuccullata (AY038076) and O.
aupouria (AF112288) as an outgroup. Including the outgroup, 188 nucleotide positions
were variable in the COI data set. Although the NJ and MP trees were different in
appearance, they supported the same general patterns: (1) the white haplotypes clustered
together withC. hongkongensis; (2) the red haplotypes clustered together withC. ariakensis

Fig. 4. Phylogenetic trees based on neighbor-joining (A) and parsimony (B) analyses of partial mtCOI sequences
using Kimura two-parameter distances with 1000 bootstrap. O. aupouria was used as an outgroup. Numbers at
steps are bootstrap values that are N50.
H. Wang et al. / Aquaculture 242 (2004) 137–155 147
of Lam, Kim and O Foighil; and (3)C. ariakensis of Yu fell into a clade withC. sikamea and
C. gigas (Fig. 4A,B).
Sequence divergence between white and red haplotypes ranged from 13.6% to 14.4%,
which were similar to that between C. ariakensis and C. nippona and considerably higher
than that between C. gigas and C. sikamea (Table 4). Divergences among the white and C.
hongkongensis haplotypes were less than 1%. Similarly, divergences among the red and C.
ariakensis (except that of Yu et al., 2003) were also less than 1%.
Despite the high levels of variation in nucleotide sequence, the amino acid (AA)
sequence of the COI fragment was conserved. WhiteA and WhiteB halotypes had the same
AA sequence (WhiteA), White C had another AA sequence (WhiteC) and all eight
haplotypes of the red oysters shared one AA sequence (RedA). Among the three AA
halotypes (WhiteA, WhiteC and RedA), four amino acids were variable due to
transversions. RedA differed from WhiteA and WhiteC in two and four AA positions,
respectively. WhiteA and WhiteC differed in two AA positions.
3.2.3. Nuclear 28S rRNA sequence
Sequences of an 812-bp fragment of the nuclear 28S rRNA gene from the 30 oysters
sequenced in this study produced only 2 haplotypes: 1 for the 12 white oysters and 1 for
the 18 red oysters (Table 1).

Table 4
Pair-wise sequence divergence for partial mt cytochrome oxidase I fragments from the red (Red) and white (White) oysters and reference species
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
RedA 0
RedB(SZ) 0.0017 0
RedC(ZJ) 0.0017 0.0035 0
RedD(YJ) 0.0017 0.0035 0.0035 0
RedE(BH) 0.0017 0.0035 0.0035 0.0035 0
C. ariaLamA 0 0.0017 0.0017 0.0017 0.0017 0
RedF(WF) 0.0017 0.0035 0.0035 0.0035 0.0035 0.0017 0
RedG(WF) 0.0035 0.0052 0.0017 0.0052 0.0052 0.0035 0.0017 0
RedH(WF) 0.0035 0.0052 0.0052 0.0052 0.0035 0.0035 0.0017 0.0035 0
C. ariaFoighil 0.0017 0.0035 0.0035 0.0035 0.0035 0.0017 0 0.0017 0.0017 0
C. ariaKim 0.0087 0.0104 0.0104 0.0104 0.0104 0.0087 0.0069 0.0087 0.0087 0.0069 0
WhiteA 0.1378 0.1378 0.1358 0.1399 0.1399 0.1378 0.1399 0.1378 0.1421 0.1399 0.1481 0
WhiteB(ZJ) 0.1399 0.1399 0.1378 0.1419 0.1419 0.1399 0.1419 0.1399 0.1441 0.1419 0.1501 0.0017 0
WhiteC(SZ) 0.1401 0.1401 0.1380 0.1421 0.1421 0.1401 0.1421 0.1401 0.1443 0.1421 0.1503 0.0035 0.0052 0
C. honkon.A 0.1378 0.1378 0.1358 0.1399 0.1399 0.1378 0.1399 0.1378 0.1421 0.1399 0.1481 0 0.0017 0.0035 0
C. nippona 0.1475 0.1455 0.1455 0.1496 0.1496 0.1475 0.1496 0.1475 0.1518 0.1496 0.1578 0.1274 0.1254 0.1296 0.1274 0
C. ariaYu1 0.1634 0.1613 0.1655 0.1655 0.1613 0.1634 0.1655 0.1676 0.1657 0.1655 0.1739 0.1458 0.1438 0.1481 0.1458 0.1625 0
C. sikamea 0.1657 0.1636 0.1678 0.1678 0.1636 0.1657 0.1678 0.1699 0.1680 0.1678 0.1762 0.1501 0.1481 0.1524 0.1501 0.1648 0.0192 0
C. gigas 0.1598 0.1577 0.1619 0.1619 0.1619 0.1598 0.1619 0.164 0.1642 0.1619 0.1703 0.1324 0.1304 0.1346 0.1324 0.1615 0.1130 0.0992 0
C. iredalei 0.1800 0.1778 0.1821 0.1821 0.1778 0.1800 0.1778 0.1800 0.178 0.1778 0.1864 0.1750 0.1729 0.1752 0.1750 0.1646 0.1824 0.1913 0.1814 0
Names of sample collection sites are abbreviated as SZ for Shenzhen, YJ for Yangjiang, ZJ for Zhanjiang, BH for Beihai and WF for Weifang.
H.Wanget
al./Aquacultu
re242(2004)137–155
148

Fig. 5. Phylogenetic trees based on neighbor-joining (A) and parsimony (B) analyses of partial 28S rRNA
sequences using Kimura two-parameter distances with 1000 bootstrap. O. aupouria was used as an outgroup.
Numbers at steps are bootstrap values that are N50.
H. Wang et al. / Aquaculture 242 (2004) 137–155 149
For phylogenetic analysis, the two haplotypes identified in this study was compared to
available sequences from GenBank: C. ariakensis (O’Foghil AF137052), C. gigas
(AF137051), C. virginica (AF137050), C. belcheri (Z29545), C. nippona (AB110095), S.
cuccullata (Z29553) C. rhizophorae (AF137049) and O. angasi (AF137046) as an
outgroup. Including the outgroup, 24 nucleotide positions were variable in the 28S data
set. In both NJ and MP trees, the white oyster was in the same clade with C. gigas and the
red oyster was grouped with C. ariakensis (Fig. 5). The sequence divergence between the
Table 5
Pair-wise sequence divergence for partial 28S rRNA fragments from the red (Red) and white (White) oysters and
reference species
1 2 3 4 5 6 7 8 9 10 11
Red
(SZ,YJ,ZJ,BH,WF)
0
C. ariakensis 0 0
White
(SZ,YJ,ZJ,BH)
0.0115 0.0115 0
C. nippona 0.0232 0.0232 0.0166 0
C. gigas 0.0127 0.0127 0.0063 0.0179 0
C. sikamea 0.0115 0.0115 0.0051 0.0166 0.0038 0
C. belcheri 0.0313 0.0313 0.0273 0.0286 0.0286 0.0273 0
C. rhizophorae 0.0842 0.0842 0.0884 0.0901 0.0871 0.0886 0.0921 0
C. virginica 0.0828 0.0828 0.0870 0.0886 0.0857 0.0872 0.0906 0.0038 0
O. angasi 0.0852 0.0852 0.0852 0.0938 0.0837 0.0867 0.0991 0.0917 0.0903 0
S. cuccullata 0.0798 0.0798 0.0769 0.0843 0.0770 0.0799 0.0879 0.0917 0.0932 0.0694 0
Names of sample collection sites are abbreviated as SZ for Shenzhen, YJ for Yangjiang, ZJ for Zhanjiang, BH for
Beihai and WF for Weifang.

H. Wang et al. / Aquaculture 242 (2004) 137–155150
white and red is about 1.2%, which was higher than the divergence between C. virginica
and C. rhizophorae (Table 5).
Sequences obtained in this study have been deposited in GenBank (accession number
AY632546–AY632568).
4. Discussion
4.1. Classification of the white and red forms of C. rivularis
The taxonomic status of the two forms of C. rivularis (Gould, 1861) has been the
subject of much confusion and debate in China. Although differences between the two
forms of C. rivularis have long been recognized (Zhang and Qi, 1960; Huang, 1962), there
has been no agreement on the taxonomic status of either form. Currently, the two forms are
being referred to as the bred meatQ and bwhite meatQ of C. rivularis. Using allozyme
markers, Li et al. (1988) was the first to show that the two forms are genetically distinct
and should be considered as two separate species. In a careful analysis of anatomical
characters, Li (1989) demonstrated that these oysters are two different species. This study
provides DNA sequence data for the white and red oysters, which support the conclusion
that the red and white oysters are separate species. In all three genes analyzed, sequence
divergence between the white and red oysters is higher than that between some of well-
established species. Further, sequence analysis confirms that both species are members of
Crassostrea, as they clustered with known members of the genus and away from members
of Ostrea and Saccostrea.
Although there was general agreement that the two oysters were separate species, there
was no consensus on what species they were. Li et al. (1988) argued that the white meat was
C. rivularis and the red meat was probably C. iredalei. On the other hand, Li and Qi (1994)
concluded that the redmeat wasC. rivularis and they named the white meat as bjinjiang-likeQoyster (Crassostrea sp.), giving a full and specific description. Apparently unaware of the
literature on the red and white ofC. rivularis in China, Lam andMorton (2003) described an
oyster from Hong Kong and named it C. hongkongensis. Boudry et al. (2003) also
recognized an oyster from Hong Kong as different from other oysters, based on the existing
molecular database, but refrained from naming it as a new species until further studies.
Our sequence data clearly demonstrate that C. hongkongensis is the same oyster as the
bwhite meatQ of C. rivularis that have been recognized in China for many decades and
recently named bjinjiang-likeQ oyster by Li and Qi (1994). The 16S sequence of the white
oyster is identical to that of C. hongkongensis. The COI sequence had more haplotypes,
but sequence divergence between haplotypes of the white and C. hongkongensis ranged
between 0% and 0.5%, significantly lower than that between any pair of known species. C.
gigas and C. sikamea, two of the most closely related species included in our analysis, had
a divergence of 1.9% in the COI fragment.
Our sequence data also show that the red oyster is the same species as C. ariakensis.
Divergences among all red and C. ariakensis haplotypes in 16S and COI fragment were
lower than that between any known species. All red oysters from this study had one
haplotype for the 28S fragment, which is identical to that of C. ariakensis.

H. Wang et al. / Aquaculture 242 (2004) 137–155 151
4.2. Taxonomic history
Since the white oyster had been described as C. rivularis Gould (1861) (Zhang and Qi,
1960; Li et al., 1988) and Crassostrea sp. (Li and Qi, 1994) before being named as C.
hongkongensis, the question becomes what is the proper name for this species. The name
C. rivularis (Gould, 1861) clearly has seniority; however, the white oyster does not fit the
original description of C. rivularis by Gould (1861). Gould’s original description of O.
rivularis is in Latin, which can be translated as bdisk-like, round, flat; lower valve thick
and purple with distant radiant ribs and sub-tubular rays; upper valve simple with branchy
and brook-like purple veins; cavity minimally deep, oval, with ash-white margin; hinge
weakQ. The type specimen, which was measured as 60 mm in diameter and 10 mm thick,
was collected from southern China as part of the north Pacific Exploring Expedition
(Johnson, 1964). Gould stressed that bthe rays of little tubes below, and the veins above,
are usually clear and distinctive charactersQ, which is absent from the white oysters
described here. Also, the shape of the white oyster is elongated, rather than round. The
height/width ratio of the 69 white oysters sampled in this study averaged 3.4 (ranging from
1.7 to 5.4), compared to 6.0 of Gould’s type specimen. The conclusion by Li et al. (1988)
that the white oyster was C. rivularis was without reference to Gould’s description and
based on the finding that the genetic distance between white and C. gigas is similar to that
between C. gigas and C. rivularis by Buroker et al. (1979), which is probably incidental.
Li and Qi (1994) recognized that the white oyster should not be called C. rivularis. They
provided a full description of the white oyster as an independent species, jinjiang-like
oyster (Crassostrea sp.), but did not give a full scientific name. Consequently, in our view,
C. hongkongensis (Lam and Morton, 2003) becomes the only valid name for the white
oyster from south China.
The round and disk-like shell of Gould’s specimen is somewhat similar to that of the red
oyster described in our study. Li and Qi (1994) concluded that the red meat is C. rivularis,
probably based on the shell shape and the fact that red oyster was widely distributed in
China and the subject of most traditional literature on C. rivularis in China. However, our
observation of the red oysters found no brays of little tubes below and the veins aboveQ thatwas stressed by Gould (1861) as characteristic of O. rivularis. Japanese researchers
initially used the name O. rivularis for an oyster from Ariake Bay and later replaced it with
C. ariakensis (Wakiya, 1929), because the shell characters of the Ariake oyster did not fit
Gould’s description (Habe and Kosuge, 1967; Torigoe, 1981). Our sequence data show
that the red oyster, particularly that from north China, is unquestionably the same species
as C. ariakensis. Also, the shell morphology of the red oysters in our study closely
resembles that of C. ariakensis from Japan (Fig. 2G,H). Therefore, we propose that the red
oyster from China should take the name C. ariakensis and the name C. rivularis should
not be used for the red oysters from China in the future. The taxonomic history of the
white and red oysters China is summarized in Table 6. Our study suggests that neither the
red nor white oysters from China can be classified as C. rivularis based on the
morphological descriptions given by Gould (1861). It is most likely that O. rivularis of
Gould (1861) referred to a species that is different from C. ariakensis (Habe and Kosuge,
1967; Torigoe, 1981) as well as C. hongkongensis. The name C. rivularis was adopted in
China probably based on early Japanese literature, which was incorrect.

Table 6
Taxonomic history of the white and red forms of C. rivularis from China
Date and events Reference
1861, Ostrea rivularis, specimen from south China Gould, 1861
1869, Ostrea rivularis, specimen from Japan Lischke, 1869
1913, Ostrea ariakensis Wakiya MS, specimen from Japan Wakiya, 1929
1929, Ostrea rivularis, specimen from China Lamy, 1929
1954, Ostrea rivularis, specimen from south China Ye, 1954
1956, Ostrea (Crassostrea) rivularis, specimen from north China Zhang and Lou, 1956
1960, Ostrea rivularis, white and red meat forms recognized in China Zhang and Qi, 1960
1967, Ostrea rivularis, invalidated and replaced by C. ariakensis in Japan Habe and Kosuge, 1967
1988, Crassostrea rivularis=white meat; C. iredalei=red meat Li et al., 1988
1994, Crassostrea rivularis=red meat; C. sp.=white meat Li and Qi, 1994
2003, Crassostrea hongkongensis, specimen from Hong Kong Lam and Morton, 2003
2004, C. rivularis replaced; C. hongkongensis=white; C. ariakensis=red This study
H. Wang et al. / Aquaculture 242 (2004) 137–155152
4.3. Distribution of C. hongkongensis and C. ariakensis in China
The white (C. hongkongensis) and red (C. ariakensis) oysters can be clearly separated
by morphological differences. For 30 oysters separated by morphology and subjected to
sequencing, morphological classification was supported by sequence analysis in all cases.
Structural relationships among gill, soft-tissue and mantle provides unambiguous
separation of the two species (Li, 1989). Despite considerable variation and plasticity,
shell characters are also somewhat informative. The red oysters are generally oval shaped,
while the white oysters are elongated. The white oysters have a longer umbo or ligament
channel than the red oyster. Shell morphology alone cannot separate the two species with
certainty.
Our limited sampling of five sites suggests that C. hongkongensis is not limited to
Hong Kong. It dominated samples from Zhanjiang and Shenzhen in Guangdong province
and accounted for about 44% of samples from Beihai, Guangxi province. The white oyster
has been reported from as far north as Xiamen, Fujian province (Li et al., 1988). No white
oyster was found in our samples from Weifang, Shandong province in the north. From
another study, samples from Zhoushan, Zhejiang province did not contain white oysters
(unpublished data). It seems that the white oyster is distributed from Fujian to Guangxi
provinces in south China, with populations centering in Guangdong province. Further
survey is needed to detail its wider geographic range.
C. ariakensis is found at all five sampling sites in this study, ranging from Bohai Sea in
the north to Beihai near the Vietnam border. This distribution probably represents the
natural distribution of this species in China. Aquaculture of this species is limited to
southern China, and there is no known history of introductions between north and south
China. C. ariakensis is the dominant species in the north and probably responsible for the
wide distribution of C. rivularis that is previously reported in China (see review by Zhou
and Allen, 2003), but most previous studies did not separate the red from the white oysters.
Our data suggest that existing literature on C. rivularis from Zhejiang and north should be
considered as that for C. ariakensis. Literature from Guangdong was likely based on C.

H. Wang et al. / Aquaculture 242 (2004) 137–155 153
hongkongensis or a mixture of both. Findings of this study call for a complete
reclassification of major oyster estuaries in China.
Our data also show that C. ariakensis from the north are genetically distinctive from
C. ariakensis from the south. In both 16S and COI sequences, northern C. ariakensis
from Weifang did not share a single common haplotype with the southern C. ariakensis
from the four southern sites. The northern C. ariakensis were more closely related to C.
ariakensis from Korea (Kim et al., 2000) and Japan (O Foighil et al., 1995). The southern
C. ariakensis is more closely related to C. ariakensis from Hong Kong (AY160752–
AY160754). Morphologically, the southern C. ariakensis had purple adductor muscle
scars and deep umbo cavities, while the northern C. ariakensis had white adductor muscle
scars and shallow umbo cavities. The molecular data suggest that biogeographical barriers
exist, and the northern and southern C. ariakensis have been in complete isolation for
sometime. The Yangtze river estuary is known to be a barrier for the distribution of many
marine invertebrates (Xu, 1997).
Whether the northern and southern C. ariakensis should be considered as two
populations or subspecies, has to be determined by future studies with a focus on their
biological differences. In DNA sequences, the level of divergence between the two
geographic populations is less than 1% for both 16S and COI. For AA sequence of COI
and DNA sequence of 28S, the northern and southern C. ariakensis did not show any
divergence. However, it is possible that reproductive isolation has allowed sufficient
biological differences to accumulate that they should be considered two subspecies or
species. Hybridization experiments between the two populations would be most
informative.
Yu et al. (2003) reported 16S and COI sequences for an oyster collected from Yantai
(near our Weifang site) that they referred to as C. ariakensis (without morphological
description). Our analyses show that their specimens are probably not C. ariakensis, but
likely C. gigas (Fig. 5, 7). C. gigas does occur in Bohai Sea. Li and Qi (1994) noted that
C. gigas had be mistakenly classified as C. rivularis. This study demonstrates the
importance of using both morphological characteristics and phylogenetic analysis in oyster
classification. Molecular data alone may not solve confusions originated from
morphological descriptions.
Acknowledgments
We thank Fengshan Xu, Xiaoxu Li and Suping Zhang for advice on oyster
taxonomy. Zhihua Lin, Zhongjie You, Huayong Que, Tao Zhang, Ganggui Zhao and
Shuang Zeng and Xiao Liu provided oyster samples to this study. We thank Yongping
Wang for DNA samples of some species. We are also grateful to Baozhong Liu, Bo
Dong and Xiaodong Zheng for helpful comments on the data analysis. This work is
supported by grants from National Science Foundation of China (no. 39825121 to Guo
and no. 40406032 to Wang) and Chinese Academy of Sciences (ZKCX2-211 to G.
Zhang). Guo’s participation is supported by Rutgers University and a grant from the
NOAA CBO Non-native Oyster Research Program (NA04NMF4570424). This is
Rutgers contribution no. IMCS-2004-15.

H. Wang et al. / Aquaculture 242 (2004) 137–155154
References
Amemiya, I., 1928. Ecological studies of Japanese oysters with special reference to the salinity of their habitats.
J. Coll. Agri. Univ. Tokyo 9 (5), 332–382.
Banks, M.A., Hedgecock, D., Walters, C., 1993. Discrimination between closely related Pacific oyster species
(Crassostrea) via mitochondrial DNA sequences coding for large subunit rRNA. Mol. Mar. Biol. Biotechnol.
2, 129–136.
Boudry, P., Heurtebise, S., Lapegue, S., 2003. Mitochondrial and nuclear DNA sequence variation of presumed
Crassostrea gigas and Crassostrea angulata specimens: a new oyster species in Hong Kong. Aquaculture
228, 15–25.
Buroker, N.E., Hersbberger, W.K., Chew, K.K., 1979. Population genetics of the family Ostreidae: II.
Interspecific studies of the genera Crassostrea and Saccostrea. Mar. Biol. 54, 171–184.
Calvo, G.W., Luckenbach, M.W., Allen Jr., S.K., Burreson, E.M., 2001. A comparative field study of C.
ariakensis (Fujita 1913) and Crassostrea virginica (Gmelin 1791) in relation to salinity in Virginia.
J. Shellfish Res. 20, 221–229.
Felsenstein, J., 1989. PHYLIP—Phylogeny Inference Package (Version 3.2). Cladistics 5, 164–166.
Folmer, O., Black, M., Hoeh, W., Lutz, R., Vrijenhoek, R., 1994. DNA primers for amplification of mitochondrial
cytochrome C oxidase subunit I from diverse metazoan invertebrate. Mol. Mar. Biol. Biotechnol. 3, 294–299.
Guo, X., Ford, S., Zhang, F., 1999. Molluscan aquaculture in China. J. Shellfish Res. 18, 19–31.
Gould, A.A., 1861. Descriptions of shells collected in the North Pacific Exploring Expedition under Captains
Ringgold and Rodgers. Proc. Boston Soc. Nat. Hist. 8, 33–40 (April).
Habe, T., Kosuge, S., 1967. Common shells of Japan in colour. Hoikusha (Osaka) 18, 233.
Hirase, S., 1930. On the classification of Japanese oysters. Jpn. J. Zool. 3, 1–65.
Huang, J., 1962. Study on the major cultivated species of oyster in Guangdong province. Transactions of Annual
Meeting of Oceanology and Limnology Society of Guangdong, pp. 41–52. (in Chinese).
Johnson, R.I., 1964. The recent Mollusca of Augustus Addison Gould. Bull. - U. S. Natl. Mus. 239, 182.
Kim, S.-H., Lee, S.-Y., Park, D.-W., An, H.-S., 2000. Phylogenetic relationship among four species of Korean
oysters based on mitochondrial 16S rDNA and COI gene. Korean J. Biol. Sci. 16 (2), 203–211.
Kimura, M., 1980. A simple method for estimating evolutionary rates of base substitutions through comparative
studies of nucleotide sequences. J. Mol. Evol. 16, 111–120.
Lam, K., Morton, B., 2003. Mitochondrial DNA and morphological identification of a new species of
Crassostrea (Bivalvia: Ostreidae) cultured for centuries in the Pearl River Delta, Hong Kong, China.
Aquaculture 228, 1–13.
Lamy, E., 1929. Revision des Ostrea vivants du Museum National d’Histoire Naturelle de Paris. J. Conchliol. 73,
1–168.
Lapegue, S., Boutet, I., Leitao, A., 2002. TransAtlantic distribution of a mangrove oyster species revealed by 16S
mtDNA and karyological analyses. Biol. Bull. 202, 232–242.
Li, X., 1989. A comparative morphology of mantle cavity in some Chinese oysters. Oceanol. Limnol. Sin. 20 (6),
502–507 (in Chinese).
Li, X., Qi, Z., 1994. Studies on the comparative anatomy, systematic classification and evolution of Chinese
oysters. Stud. Mar. Sin. 35, 143–173 (in Chinese).
Li, G., Hu, Y., Qing, N., 1988. Population gene pools of big size cultivated oysters (Crassostrea) along the
Guangdong and Fujian coast of China. Proc. Mar. Biol. of South China Sea. China Ocean Press, pp. 51–70.
Lischke, C.E., 1869. Japanische Meeres-Conchlien, vol. 1. Theodor Fischer, Cassel, pp. 192.
Littlewood, D.T., 1994. Molecular phylogenetics of cupped oysters based on partial 28S rRNA gene sequences.
Mol. Phylogenet. Evol. 3 (3), 221–229.
Mackenzie, Jr., C.L., Burrell, Jr., V.G., Rosenfield, A., Hobart, W.L., 1997. The History, Present Condition, and
Future of the Molluscan Fisheries of North and Central American and Europe, Volume 1, Atlantic and Gulf
Coasts. U.S. Department of Commerce, NOAA Technical Report NMFS 127, 234 pp.
Moore, D., 1993. Preparation of genomic DNA from mammalian tissue. In: Ausubel, F.M. (Eds.), Current
Protocols in Molecular Biology, vol. 1. John Wiley, New York, pp. 1–2.
Moore, R.C., Teichert, C., McCormick, L., Williams, R.B., 1971. Treatise on Invertebrate Paleontology, Part N,
3(3). The Geological Society of America, Inc and the University of Kansas.

H. Wang et al. / Aquaculture 242 (2004) 137–155 155
O Foighil, D., Taylor, D.J., 2000. Evolution of parental care and ovulation behavior in oysters. Mol. Phylogenet.
Evol. 15 (2), 301–313.
O Foighil, D., Gaffney, P.M., Hilbish, T.J., 1995. Differences in mitochondrial 16S ribosomal gene sequences
allow discrimination among American (Crassostrea virginica) and Asian (C. gigas, C. ariakensis) oyster
species. J. Exp. Mar. Biol. Ecol. 192, 211–220.
O Foighil, D., Gaffney, P.M., Wilbur, W.A., 1998. Mitochondrial cytochrome oxidase I gene sequence support an
Asian origin for the Portuguese oyster Crassostrea angulata. Mar. Biol. 131, 497–503.
Okutani, T., 2000. Marine Mollusks in Japan. Tokai University, pp. 923–927.
Palumbi, S.R., Martin, A., Romano, S., 1991. The simple fool’s guide to PCR. University of Hawaii Press,
Honolulu, Hawaii.
Park, J.K., O Foighil, D., 2000. Sphaeriid and Corbiculid clam represent separate Heterodont bivalve radiations
into freshwater environments. Mol. Phylogenet. Evol. 14 (1), 75–88.
Tompson, J.D., Higgins, D.G., Gibson, T.J., 1994. CLUSTAL W: improving the sensitivity of progressive
multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix
choice. Nucleic Acids Res. 22, 4672–4680.
Torigoe, K., 1981. Oysters in Japan. J. Sci. Hiroshima Univ., Ser. B, Div. 129, 291–481.
Wakiya, Y., 1929. Japanese food oysters. Jpn. J. Zool. 2, 359–367.
Wang, R., Wang, Z., Zhang, J., 1993. Marine Molluscan Culture. Qingdao Ocean University Press, Qingdao,
China. (in Chinese).
Xu, F., 1997. Bivalves from China Seas. Science Press, Beijing. In Chinese.
Ye, X., 1954. Oysters from Xiamen area. J. Xiamen Univ. 3, 56–80. (in Chinese).
Yu, Z., Kong, X., Zhang, L., Guo, X., Xiang, J., 2003. Taxonomic status of four Crassostrea oysters from China
as inferred from mitochondrial DNA sequences. J. Shellfish Res. 22 (1), 31–38.
Zhang, X., Lou, Z.K., 1956. Studies on oysters in China. Acta Zool. Sin. 8, 65–94. (in Chinese).
Zhang, X., Qi, Z., 1960. Bivalves of Nanhai. Science Press, Beijing, China. (in Chinese).
Zhou, M., Allen Jr., S.K., 2003. A review of published work on Crassostrea ariakensis. J. Shellfish Res. 22 (1),
1–20.





















![Evaluating the impact of oyster (Crassostrea virginica ......sediment N cycling, oyster-free cages and cages with adult or juvenile oysters [Crassostrea virginica (Gmelin, 1791)] were](https://img.dokumen.tips/doc/110x75/60c4a8704d94be71f945d9dd/evaluating-the-impact-of-oyster-crassostrea-virginica-sediment-n-cycling.jpg)







