Embed Size (px)
Citation preview

1
Circulation and Water properties and Their Relationship to the Oyster Disease, 1 MSX, in Delaware Bay 2
3
Zhiren Wang1 ([email protected]), 4
Dale Haidvogel1, David Bushek2, Susan Ford2, Eileen Hofmann3, Eric Powell2, and John Wilkin1 5
1. Institute of Marine and Coastal Sciences, Rutgers University, New Brunswick, NJ, 6 2. Haskin Shellfish Research Laboratory, Rutgers University, Port Norris, NJ 7
3. Department of Ocean, Earth and Atmospheric Sciences, Old Dominion University, Norfolk,VA 8
9
Abstract 10
We apply a high-resolution hydro-dynamical model to investigate the role of physical factors 11
influencing infection prevalence of Haplosporidium nelsoni, causative agent of MSX disease in the 12
eastern oyster (Crassostrea virginica), in Delaware Bay, USA. Validation studies are first conducted in 13
the target year 2000, for which good observational data are available. The Regional Ocean Modeling 14
System (ROMS) shows quantitative agreement between the model simulations and concurrent 15
observations of water level, temperature and salinity, matching well for both high-frequency (including 16
tidal) and low-frequency (e.g., seasonal) signals in their phase and amplitude. Multi-year simulations 17
of the river flow, salinity, water temperature and particle transport for periods representing temporal and 18
spatial variations in H. nelsoni infection prevalence (1974-76, 1979-81, 1984-86, 1990-92, and 2006-09) 19
are then conducted to explore the relationships between the variations of prevalence and physical 20
conditions in the Bay. We assess the degree to which the variations in water properties and particle 21
transport are temporally and spatially correlated with infection prevalence variations. Results show 22
statistically significant correlations between the observed prevalence of MSX and multiple physical 23
factors including river flow and salinity (themselves highly correlated), temperature, as well as the 24
co-occurrence of elevated temperature and salinity values. Observed occurrences of extreme values of 25

2
MSX at up-Bay locations do appear to correspond to periods of enhanced up-Bay transport together with 1
hospitable temperature and salinity conditions. 2
3
4
Key words: Delaware Bay, MSX disease, Haplosporidium nelsoni, Crassostrea virginica, Regional 5
Ocean Modeling System 6
7

3
1. Introduction 1
Oysters (Crassostrea virginica) in the mid-Atlantic and northeastern United States are afflicted with a 2
lethal disease caused by a water-borne protozoan parasite, Haplosporidium nelsoni. The disease is 3
called MSX disease after the acronym first given to the parasite (Multinucleated Sphere of unknown 4
affinity). The first recorded outbreak of MSX disease was in 1957 in Delaware Bay, USA, where 5
prevalence has continued to display significant spatial and temporal variability (Ford and Bushek, this 6
volume). Early surveys showed that infection prevalence decreased along a decreasing salinity 7
gradient (Andrews, 1966; Haskin and Ford, 1982). Subsequent studies demonstrated that the 8
plasmodial stage of H. nelsoni, which is most common in oysters, was very sensitive to low salinity and 9
could be eliminated from oysters at salinities <10 (Ford, 1985). On the other hand, the spore stage of 10
what is presumed to be H. nelsoni is widely distributed throughout Delaware Bay (Barber et al., 1991) 11
and recent molecular evidence shows that the parasite can be present in very up-Bay and up-river sites 12
(Ford et al., 2009; Ford et al., this volume). 13
Previous analyses of the relationship between salinity and H. nelsoni infection prevalence have been 14
based on periodic salinity measurements or river flow records (Andrews, 1964 and 1983; Haskin and 15
Ford, 1982; Ford and Bushek, this volume). The availability of circulation models based upon the 16
Regional Ocean Modeling System (ROMS) now makes it possible to investigate the relationship of 17
water properties to MSX disease variability in Delaware Bay more precisely than before. Salinity and 18
temperature are the two physical factors most likely to influence infection patterns in the Bay. In the 19
ROMS implementation described below, salinity in the oyster-growing area is simulated from input of 20
six rivers (not just the Delaware), as well as precipitation, evaporation, mixing, and advection. 21
Temperature, which is estimated from heating (including solar radiation), mixing, and advection, is the 22
predominant correlate with seasonal infection patterns in oysters. 23

4
With ROMS, inter-annual variations in temperature and salinity can be investigated to explore the 1
separate and interactive effects of these two variables. In addition, ROMS permits us to investigate the 2
role of circulation patterns in potentially moving infective stages from down-Bay (where prevalence is 3
higher) into up-Bay regions. The method of transmission of H. nelsoni is unknown. To date, 4
attempts to experimentally transmit the parasite from infected to uninfected oysters have failed and an 5
alternate host or a vector may be involved in the life cycle (Haskin and Andrews, 1988; Ford and Tripp, 6
1996). The circulation model provides a means to test the hypothesis that H. nelsoni is distributed as a 7
passive particle as opposed to a mobile host or vector with specific, non-passive behavior. 8
The outline of this paper is as follows. In Section 2, the implementation of the Delaware Bay model 9
is described. Validation of the model for datasets collected in the year 2000 is discussed in Section 3. 10
Section 4 considers the inter-annual variability of, and correlations among, H. nelsoni infection 11
prevalence in oysters, river flow, temperature, and salinity. Section 5 examines variability in particle 12
transport from the lower to the upper Bay. Conclusions and discussion are offered in Section 6. 13
14
2. The Delaware Bay model 15
The circulation model used in this study is the Regional Ocean Modeling System (ROMS, 16
www.myroms.org). ROMS solves the three-dimensional hydrostatic primitive equations, and has been 17
developed for, and applied to, a wide variety of coastal marine applications, including many studies of 18
coastal and estuarine circulation. Recent examples include the idealized studies of the Hudson River 19
by Zhang et al. (2009) and the realistic simulations of the Columbia River plume by Liu et al. (2009). 20
21
ROMS utilizes consistent temporal averaging of the barotropic mode to guarantee both exact 22
conservation and constancy preservation properties for tracers and therefore more accurately resolves 23

5
barotropic processes, while preventing aliasing of unresolved barotropic signals into the slower 1
baroclinic motions. Accuracy of the mode-splitting is further enhanced using redefined barotropic 2
pressure-gradient terms to account for the local variations in the density field (i.e., the pressure-gradient 3
truncation error is greatly reduced). Vertical interpolation is based on conservative parabolic splines. 4
Finally, ROMS uses orthogonal curvilinear coordinates in the horizontal, an essential feature here, as 5
described below. Shchepetkin and McWilliams (2005) describe in detail the algorithms that comprise 6
the ROMS computational kernel. ROMS has been shown to produce quantitatively accurate 7
simulations of circulation and water properties in a variety of large-scale and regional settings including 8
the Hudson River (Warner et al., 2005), The Barents Sea (Budgell, 2005), the California Current System 9
(Powell e al., 2006), and the North Pacific Ocean (Curchitser et al., 2005). 10
11
2-1. Model configuration 12
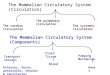
The Delaware Bay model has the following attributes. A horizontal grid of 98386 cells uses 13
orthogonal curvilinear stretching to enhance horizontal resolution in the Bay and tidal river (~230 m) 14
compared to the adjacent coastal ocean (~1500 m) where the domain extends to approximately the 50-m 15
isobath (Figure 1a,b)1. The modeled Delaware River extends as far as Trenton, NJ. There are 20 vertical 16
levels in a generalized topography-following coordinate weighted to give highest resolution near the sea 17
surface. The minimum water depth is 2 m (Figure 1c). The time step is 60 seconds and the barotropic 18
mode is temporally integrated 25 times within each baroclinic step using the split-explicit method 19
(Shchepetkin and McWilliams, 2005). Other settings include: advection via the recursive MPDATA 20
algorithm (Smolarkiewicz and Grabowski, 1990), turbulent mixing using the Generic Length Scale 21
(GLS) scheme (Umlauf and Burchard, 2003) with k-kl closure parameters (Mellor and Yamada, 1982), 22
1 To minimize the number of “land” cells within the ROMS grid, the river cells above 39.8o N have been folded back onto the grid to the south. The results below will be shown in the correct, unfolded geometry.

6
and radiation conditions (Marchesiello et al., 2001) at the open boundaries where there is no 1
independent salinity or temperature information being provided. 2
3
2-2. Forcing data 4
ROMS is forced by the following inputs: 6-hourly atmospheric fluxes (for the 1974-76 simulation) 5
from the ERA-40 data of European Centre for Medium-Range Weather Forecasting (ECMWF, 6
http://data-portal.ecmwf.int) and 3-hourly atmospheric fluxes (for all the other simulations) from the 7
North American Regional Reanalysis (NARR) (Mesinger 2006) input at six locations (Figure 1c); daily 8
river transport from US Geological Survey (USGS, http://waterdata.usgs.gov) input at six locations 9
(Figure 1b); and tidal elevation and currents at the domain perimeter with seven tidal constituents (M2, 10
N2, S2, K1, O1, M4, and M6) from North Atlantic regional harmonic tidal simulation (Mukai et al. 11
2002). 12
The temperature of the shallow and turbid water column is sensitive to the net heat flux, but also to 13
the depth scale assumed for solar radiation absorption. In ROMS, this absorption profile is represented 14
as two exponential functions following Paulson and Simpson (1977), with parameters determined by an 15
assumed “water type”. Our early simulations showed the Delaware Bay model to be too warm, 16
especially in the summer months. To explore this issue, three water types [Cahill et al., 2008] that 17
define different penetration depths (Table 1) and two specifications for the imposed incoming solar 18
shortwave radiation (SSWR) (Figure 2) were compared to assess model sensitivity. 19
The summer-time discrepancy in heating rate was traced to the incoming solar short-wave radiation. 20
The SSWR from the North American Regional Reanalysis (NARR) was discovered to be systematically 21
higher than regional values given by the Delaware Environmental Observation System (DEOS, 22

7
http://www.deos.udel.edu). Comparing 5-min interval DEOS SSWR and 3-hr interval NARR SSWR 1
within Delaware Bay for the period 2005-2009 when both DEOS and NARR SSWR are available 2
(Figure 2) we find ratios of DEOS to NARR SSWR of 77.4%, 77.1%, 76.7%, 76.2%, and 76.8% 3
respectively for winter, spring, summer, fall, and the whole year. The systematic difference is nearly 4
independent of season, so we make a simple correction in the simulations reported below by reducing 5
NARR SSWR by 20%. 6
7
3. Model validation 8
We choose observations of water level, temperature and salinity in the year 2000 as the datasets for 9
model validation. This time period was chosen because relatively good observations of water properties 10
are available for that year. 11
12
3-1. Data for model validation 13
The water level data are collected from the National Water Level Program (NWLP) and the National 14
Water Level Observation Network (NWLON) at the website: http://tidesandcurrents.noaa.gov. There 15
are twelve stations (Figure 1a) in Delaware Bay where the observed and predicted water levels are 16
available. The recording interval is one hour. The data records occupy 31-100% (most over 89%) of the 17
2-year period (2000-01), with record lengths of 76.4-731 days (1835-17544 samples). 18
The observed water temperature (oC) and salinity are provided by the Pre-Construction Oyster and 19
Water Quality Monitoring Study for the Main Channel Deepening Project [prepared by Versar, Inc., 20
http://www.versar.com]. All these data were collected by deploying in-situ YSI series 6600 data loggers 21
moored one-meter from the bottom at the nine oyster beds (Figure 1b) monitored within Delaware Bay. 22

8
Their recording interval is 15 minutes for the period 13 May-12 November 2000 and from 8 March-12 1
April 2001. The temperature signals are dominated by the seasonal cycle; tidal-frequency signals are 2
small (Figure 3). 3
Salinity is directly affected by many environmental influences, e.g., via precipitation/evaporation, 4
river inputs, and mixing processes. Salinity can therefore be a good indicator of model fidelity in 5
response to both high- and low-frequency environmental changes. The year-2000 data records show 6
the salinity field to be dominated by variations at tidal frequencies, and secondarily by longer-period 7
signals related to episodic changes in fresh water inputs (Figure 3). Salinity variations at a given 8
horizontal location may be large, up to 10. 9
10
3-2. Simulated versus observed water level 11
ROMS does a highly accurate job in simulating water levels. For a period of 204 days (11 June to 31 12
December, 2000) with 4896 hourly samples, all the correlation coefficients between simulated and 13
observed water levels are higher than 0.9, and even higher than 0.95 for 7 out of the 12 stations (Figure 14
4). The variances (evaluated by standard deviation) of the simulated water levels are between 0.8 and 15
1.3 times those observed, even closer for the water levels at stations 1, 3, 4, 5, 6, 7, and 8 located in the 16
Bay proper. The systematic bias (observation minus simulation) in M2 amplitude is small (-0.26 to 0.06 17
meters). Using different water types (5, 6 or 7) and either the original NARR SSWR or the 20%-reduced 18
NARR SSWR makes little difference in water level response. 19
20

9
3-3. Simulated versus observed water temperature 1
Figure 5a shows the Taylor diagram (Taylor, 2001) comparing the observed and simulated 2
temperature time series at the nine Versar stations. To distinguish between the tidal-frequency signals 3
and those with longer periods, the Taylor diagram shows the results for both low-passed (periods greater 4
than a day) and high-passed (periods less than a day) signals. 5
Temperature time series at the nine Versar sampling sites are dominated by the seasonal cycle. There 6
are high correlations between the simulated and observed low-passed temperatures (Figure 5a) at all the 7
nine stations, with all the correlation coefficients being over 0.99 and the standard deviations of 8
simulated temperature being about 1.15 times those observed. The systematic bias, which has here 9
been introduced to the standard Taylor diagram by adding color to the symbols, is less than a half oC 10
(-0.25 to 0.4), based on hourly data from 13 May to 12 November 2000 (4416 samples). 11
Though making a much lower contribution to the temperature time series, high-passed signals are 12
nonetheless recovered with measureable skill, with the correlation being between 0.4 and 0.75. For the 13
shallow coastal water column, different incoming solar radiation levels make a significant difference. 14
The original NARR SSWR was initially observed to make the Bay too warm by about 0.6 to 1.4oC (not 15
shown). 16
17
3-4. Simulated versus observed salinity 18
Salinity time series at the nine Versar stations show strong contributions at both tidal frequencies and 19
at longer periods associated with variations in fresh water inputs (e.g., Figure 3). On average, salinity 20
values decrease as the stations move towards the upper bay and are closely tied to river inputs, as shown 21
more quantitatively below. 22

10
The correlation coefficients between simulated and observed low-passed salinities (Figure 5b) are 1
higher than 0.8 at all but one station. The standard deviations of simulated salinity are about 0.85 to 1.2 2
times those observed. The systematic bias is from about 0.41 to -1.75. Bias is greatest at the two most 3
down-Bay locations (Fig. 1b, stations 1 and 2), where the simulated salinities exceed observations by 4
greater than 1.0, suggesting either contamination of the salinity signals from the open boundaries (at 5
which no independent salinity information is being supplied) or a local deficit of fresh water input. 6
Correlations between observed and simulated high-passed salinity time series (therefore including the 7
semi-diurnal tidal signals) vary considerably in space, being between 0.45 and 0.85, with accompanying 8
normalized standard deviations of between 0.5 and 1.5. Simulations using the original and the 20% 9
reduced NARR SSWRs show little difference in seasonal mean salinity. Water type 7 increases the 10
seasonal mean salinity by about 0.1 to 0.3 in summer and fall because of the stronger surface 11
evaporation caused by stronger surface heating (water type 7 concentrates SSWR more strongly in the 12
surface layer). 13
14
4. Inter-annual changes in Haplosporidium nelsoni infection prevalence (MSXP), river flow, 15
temperature, and salinity 16
Haplosporidium nelsoni infection prevalence (hereafter, MSXP) has been assayed one to two times 17
per year on 5 sites in Delaware Bay (Arnolds, Cohansey, New Beds-Bennies, Egg Island and Leased 18
Grounds) since 1958, shortly after the initial outbreak. Between 1958 and 2010, all beds were assayed 19
in fall (October/November). From 1958 through 1991 the beds were also assayed in spring (May). 20
These months represent annual infection peaks (Ford and Haskin, 1982). Figure 6 shows histogram 21
time series of MSXP at five locations within the Bay, accompanied by a similar time series for Delaware 22
River discharge at Trenton. General patterns of MSXP include a systematic down-Bay increase, where 23

11
higher salinity favors the parasite, with lower values at the up-Bay locations. Fresh water inflow at 1
Trenton is larger in winter/spring by a factor of about two over summer/fall. Lastly, note the reduced 2
down-Bay prevalence of MSXP associated with the development of increased resistance after 1987 3
(Ford and Bushek, this volume). 4
Prior studies suggest (Andrews, 1964; Haskin and Ford, 1982) that the up-Bay prevalence of H. 5
nelsoni is highly correlated with fluctuations in salinity, and therefore with fresh water inflow. Visual 6
examination of the time series in Figure 6 suggests an imperfect (anti-)correlation between MSXP and 7
river input. We quantify this correlation in the sub-sections below. 8
Rather than a single determining factor, however, we hypothesize that MSXP at up-Bay locations is 9
related to the co-occurrence of three factors. These are 1) the availability of high infection prevalence 10
at down-Bay locations (i.e., a source region), 2) a transport mechanism from lower- to upper-Bay 11
locations (here envisioned simply as an efficient means of passive transport), and 3) environmental 12
conditions (i.e., temperature and salinity) conducive to H. nelsoni survival and ability to proliferate in 13
oysters.2 14
To investigate these hypothesized relationships, we have selected five multi-year periods for study. 15
The periods selected for simulation represent variations in the temporal and spatial variations of H. 16
nelsoni infections in oysters at these locations in relation to Delaware River flow measured at Trenton, 17
New Jersey (Table 2). The five periods, and their respective overall properties, are: 1974-76 18
(average/above average river flow, low up-Bay MSXP), 1979-81 (average river flow, low up-Bay 19
MSXP), 1984-86 (average river flow, high up-Bay MSXP [especially 1985-86]), 1990-92 (average river 20
2 To begin, we will assume that the first of these conditions is always satisfied, namely that MSX levels in the lower Bay are sufficient to act as a source.

12
flow, high up-Bay MSXP [especially 1992]), and 2006-09 (average/above average river flow, elevated 1
up-Bay MSXP in 2008). Short-term deviations from averages within each period are noted in Table 2. 2
3
4-1. Inter-annual variations in physical properties 4
Following successful model validation, we carried out simulations of the water properties in 5
Delaware Bay for the five multi-year periods (1974-76, 1979-81, 1984-86, 1990-92, and 2006-09). In 6
addition to quantification of salinity and temperature, we developed a new index, the “salty-warm water 7
area” (SWA), which is the surface area of the Bay whose bottom salinity is higher than 17.5 and whose 8
bottom temperature is higher than 12.5oC. These values represent conservative estimates of the lower 9
thresholds for H. nelsoni proliferation in oysters (Ford and Haskin, 1982; Haskin and Ford, 1982). 10
Subsequent statistical analysis was used to explore the relationships among the variations of prevalence 11
and physical conditions in the Bay. 12
Figure 7 displays time series for our five multi-year periods (16 years in total) of the following 13
physical properties: bottom-average salinity at several locations (see also Fig. 8) within the Bay, 14
bottom-average temperature3, salty-warm water area, and the total fresh water inflow of the six river 15
inputs. Also shown are anomalies for each of these quantities. Anomalies for temperature and SWA 16
are obtained by removal of a 16-year mean seasonal cycle; anomalies for salinity and river flow are 17
formed by removal of a 16-year mean value (i.e., by removal of a constant from the time series at a 18
given location). 19
Considerable inter-annual variation in the physical environment is evident. A brief summary of 20
these variations is as follows: 21
3 At any given time, temperature varies little among the various locations within the Bay; hence, we show only a single Bay-wide average for bottom temperature.

13
1974-76: River input is the second highest among the five multi-year periods (61 m3/s higher 1
mean than the 16-year average). Salinity is slightly below the 16-year average, with the most 2
negative anomalies at Arnolds (-0.6) and Cohansey (-0.5), the two uppermost sites sampled. The 3
three-year-mean temperature is close to the longer-term mean. SWA is near the long-term 4
average. 5
1979-81: River flow is low (-67 m3/s relative to the three-year average) for an extended period 6
from May 1980 to January 1981 and from August to October of 1981. Mean conditions in 7
1979-81 are both the saltiest (Arnolds, +1.6; Cohansey, +1.3; Egg Island, +1.1) and coolest (-0.8 8
oC) of our five time windows. SWA is the lowest of any other of the multi-year periods. 9
1984-86: River input in this three-year period is near the 16-year mean; however, there is an 10
extended period of below-average input between July 1984 and August 1985. Both salinity and 11
temperature are “spot on” the 16-year mean values. SWA is also near the mean 16-year level. 12
Although the three-year means are near the long-term average, there are identifiable periods of 13
above- and below-average salinities. An extended period of above-average up-Bay salinity from 14
approximately July 1984 to September 1985 corresponds to the period of reduced river inflow. 15
1990-92: On average, this three-year period is a bit warmer (+0.2 oC) and saltier (Arnolds, +0.5; 16
Cohansey, +0.5; Egg Island, +0.3). The latter is associated with drier conditions (3-year mean 17
river flow -72 m3/s relative to the long-term mean). SWA is near normal. 18
2006-09 is the warmest period (+0.5 oC), and is accompanied by the largest river flow (+78 m3/s), 19
suggestive of a climate warming signal. Given the increased river inputs, salinities are lower 20
than in any other period (Arnolds, -1.0; Cohansey, -1.1; Egg Island, -0.8). Despite this 21

14
freshening, the combined inventory of warm/salty water is enhanced above the long-term mean 1
(+38 km2). 2
3
4-2. Correlation analysis 4
We have performed a systematic correlation analysis, seeking significant correlations between 5
various factors including: fall, spring and mean yearly MSXP; monthly, seasonal, and yearly river 6
discharge (from all 6 rivers and, separately, from only the Delaware River); simulated salinity; simulated 7
temperature; and simulated salty-warm area (SWA). We find statistically significant correlations among 8
several simulated physical properties and MSXP. Table 3 shows some of these correlations at Arnolds, 9
Cohansey, New Beds-Bennies, Egg Island, and Leased Grounds (Fig. 8). We define a significant 10
correlation as one whose correlation confidence is 95% and rejecting probability less than 5%. The 11
primary results of this correlation analysis are as follows. 12
River flow, temperature and salinity: As expected, river flow and simulated salinity are highly 13
negatively correlated at all locations (Table 3a, column 1). There is also a statistically 14
significant positive correlation between river flow and temperature in both winter/spring and 15
summer/fall (Table 3a, columns 2 and 3), presumably a reflection of a positive relationship 16
between temperature and winter/spring snow melt and summer/fall precipitation. The negative 17
river flow/salinity and positive river flow/temperature correlations are consistent with a negative 18
temperature/salinity correlation (Table 3a, column 4). 19
Correlations of MSXP with salinity and river flow (Table 3b, columns 1-4): Observed levels of 20
MSXP are statistically significantly correlated with spring river flow everywhere except at 21
Leased Grounds (Table 3b, columns 1 and 2). Correlation coefficients between spring MSXP and 22

15
spring river flow at the four up-Bay sites vary between -31% to -52%. In contrast, fall MSXP 1
observations are essentially uncorrelated with concurrent (i.e., fall) river flow values; however, 2
fall MSXP levels do appear to have a “memory” of river flow in the prior Spring, with negative 3
correlations of -27% to -39% at the four upper-most Bay locations. Correlations between 4
simulated salinity and spring MSXP are also high, especially in the upper Bay (Table 3b, columns 5
3 and 4). However, fall MSXP values do not show a significant correlation with simulated 6
salinity at any time of year (spring or fall). 7
Temperature versus MSXP (Table 3b, columns 5-6): In contrast to the influences of river flow 8
(and therefore salinity), which are highly significant, especially at the up-Bay locations, the 9
correlation analysis suggests that variations in temperature are of lesser importance. There 10
appears to be a weak (anti-)correlation between spring MSXP and concurrent temperature at 11
Arnolds. In addition, fall values of MSXP are weakly correlated (coefficients -0.45 to -0.47) 12
with temperature anomalies in the prior spring at New-Bennies, Egg Island and Leased Grounds. 13
Warm-salty water versus MSXP (Table 3b, column 7): Strong positive correlations are also found 14
between spring MSXP and the co-occurrence of warm and salty conditions at mid-Bay locations 15
(Cohansey and New-Bennies Beds; Table 3b, column 7). 16
17
5. Variability in particle transport between the lower and upper Bay 18
The preceding section establishes a close relationship (i.e., significant correlations) between 19
physical conditions in all Bay regions and the observed MSXP. Notwithstanding their statistical 20
significance, these correlations clearly do not explain completely the observed temporal variations in 21
MSXP. Above, we have hypothesized that additional factors – such as the availability of an effective 22
lower-Bay source region, and an efficient up-Bay transport mechanism – might also be related to the 23

16
presence or absence of H. nelsoni in upper-Bay regions. 1
During a 3-year (2007-2009) study of potential disease refuges in the upper Bay, the presence of H. 2
nelsoni was monitored using both histological and molecular methods (Ford et al., this volume). The 3
very sensitive molecular method (PCR) was able to detect the presence of the parasite even when 4
infections in oysters were not detectable by histology. These methods detected a particularly striking 5
example of up-Bay incursion of H. nelsoni that occurred in summer/fall 2008 (see Figure 6). 6
Although the average up-Bay conditions in the overall period 2006-2009 were somewhat fresher 7
and warmer than their respective longer-term means, the summer/fall months (June through November) 8
of 2008 were relatively salty and cool (Figure 7). These conditions would be hospitable to H. nelsoni if a 9
source to the upper Bay were available. 10
Here, we explore the simplest possibility: that the pulse of H. nelsoni infective elements apparently 11
introduced into the upper Bay in the summer of 2008 is related to enhanced up-Bay transport of passive 12
particles. To do so, we release passive particles4 near and at the sea bed at the horizontal locations 13
shown in Figure 8. The particles are released at five locations within the Bay (labeled 1 through 5 in 14
Figure 8), and during each of three years (2007-2009) on nine different days (1, 15 and 29 May; 12 and 15
26 June; 15 and 29 August; 12 and 26 September; and 10 October). On each of these days the particles 16
are released five times over the course of 24 hours (i.e., at three-hour intervals). The number of particles 17
released in each region 1-5 is proportional to the area shown in Figure 8. 18
The passive particles are tracked until the end of the simulation (unless they leave the Bay or [via 19
numerical round-off error] enter the land mass or sea bed). The result is a set of time series of particle 20
locations for passively redistributed water parcels. These time series may then be subjected to analysis 21
4 Passive particles are both transported by the simulated three-dimensional velocity field, and mixed vertically by a random walk process whose amplitude is obtained from the ROMS turbulent closure sub-model. The passive particles have no active behavior.

17
to determine the likelihood that particles released in one location will be transported by either advection 1
or mixing into other regions. The sub-regions for which this analysis is performed are labeled A 2
through F in Figure 8. We are particularly interested in the pathways by which, and effectiveness with 3
which, particles reach the up-Bay regions labeled E and F. 4
5
5-1. General patterns of passive particle movement 6
Figure 9 summarizes the general patterns of particle redistribution in the passive particle release 7
experiments. The two panels display the three-year average for particles released on 1 May (left) and 29 8
August (right) and subsequently tracked for 31 and 30 days, respectively. We choose these release 9
dates to represent early- and late-season dates when infective elements might be released during the 10
infection period (~June through early October) for H. nelsoni in Delaware Bay (Ford and Haskin, 1982). 11
Passive particle trajectories may be summarized as follows. 12
The effectiveness of up-Bay particle transport varies considerably with the location of particle release. 13
Direct communication of particles from the lower Bay to the upper Bay (i.e., from release site 1 to 14
regions C through F) is rare. Rather, delivery of passive particles to the upper-Bay (regions E and F) is 15
mainly from the Delaware side of the Bay at release location 4. 16
Simulations indicate that particles released in the lower Bay are retained there to a far greater extent 17
than particles released from more upper Bay sites. Of the particles released from site 1 in the Leased 18
Grounds, 73~77% remain in the surrounding region B, while 31%-49% of particles released from sites 2 19
and 3 remain in the surrounding region C (Figure 9). This result is consistent with the hypothesis, based 20
on elevated infection prevalence in Leased Ground oysters, that H. nelsoni infective elements are 21
concentrated in the lower Bay and become diluted in an up-Bay direction (Ford and Haskin, 1982). 22

18
Lastly, it may be noted from Figure 9 that retention of passive particles in mid-Bay and transport of 1
particles to upper-Bay regions are both enhanced in Fall (September) relative to Spring (May). This is 2
consistent with the seasonal variations in river inflow noted above. 3
4
5-2. Inter-annual variability in particle redistribution 5
The type of information shown in geographical perspective in Figure 9 can be compactly 6
summarized in the form of a location probability matrix (LPM). In the LPM, the probability that 7
particles released at each of the five sites (1 through 5) will spend time in one of the six Delaware Bay 8
sub-domains (A through F) is tabulated in a 5-by-6 matrix. Figure 10 shows a suite of LPMs for each 9
of the months May through September for each of the years 2007, 2008 and 2009. 10
Perhaps not surprisingly, particle residence patterns undergo large inter-annual changes. From 11
Figure 10, we note in particular the following: 12
Connectivity between release site 4 and regions E and F: As noted above, the primary route for 13
passive particles to reach up-Bay regions is via release site 45. However, the effectiveness of this 14
transfer pathway is highly variable. For the periods studied here, transfers from release site 4 to 15
regions E and F are robust (13%-34%) for all months examined except May 2007 and August 16
through September 2008. Particularly effective periods for transfer of passive particles to 17
up-Bay regions are May through July 2008 and July 2009. 18
Retention rates: Particles released at site 1 in the lower Bay are retained in the lower Bay (region 19
B) to a high degree. However the retention rate varies by month and year between 57 and 94%. 20
Similar retention rate variations are found at the other release sites. 21
5 Note that passive parcels released elsewhere, once arriving at release site 4, will thereafter be further re-distributed as if they had been released at site 4; that is, they have no memory of their prior locations.

19
Figure 11 summarizes the temporal variation in MSX prevalence in the up-Bay regions E and F for 1
the period June 2007 through September 2009. Super-imposed on Figure 11 are curves showing 2
estimates of up-Bay particle transport efficiency (the green curves). Although meaningful correlations 3
cannot be derived from these few monthly estimates, there is at least a visual correspondence between 4
periods of high observed MSXP and periods of enhanced up-Bay transport. In particular, the 5
anomalous period of high up-Bay MSXP (fall 2008) does in fact correspond to a period of enhanced 6
up-Bay receipt of passive particles. 7
8
6. Summary and discussion 9
We have developed a high-resolution hydro-dynamical model for the Delaware Bay forced by 10
hydrological inputs from six rivers and atmospheric inputs of momentum, heat and moisture. Model 11
validation against intensive in situ datasets in the year 2000 shows the model to possess significant skill 12
in predicting variations in water level, temperature and salinity. Both seasonal and tidal-frequency 13
signals are accurately reproduced with little bias. 14
The resulting Delaware Bay model has been applied to quantify the relationship between observed 15
Haplosporidium nelsoni infection prevalence (MSXP) and concurrent physical conditions in the Bay. 16
To do so, simulations of circulation and environmental conditions in Delaware Bay were performed for 17
five multi-year periods covering a total of sixteen years. Subsequent statistical analysis and passive 18
particle releases were used to explore the connections between MSXP and (e.g.) salinity and temperature. 19
The hypothesis under-lying these analyses has been that elevated levels of MSXP at low-salinity, 20
up-Bay locations depend upon a source of infective stages, an effective means of up-Bay transport, and 21
environmental conditions hospitable to MSX survival. We further hypothesized that the source of 22

20
infection comes from lower-Bay regions where infection prevalence is relatively high and that the 1
abundance of infective stages in any given year is proportional to that prevalence. 2
Results from the correlation analysis suggest that MSXP may be related to multiple physical factors, 3
depending on location and time of year. As has been hypothesized (but not quantified) previously, 4
MSXP is significantly correlated with river flow and salinity, especially at up-Bay locations. The 5
correlation of observed MSXP with simulated temperature is less strong, although there is a weak 6
negative correlation between fall MSXP and the previous spring temperature at down-Bay sites. 7
Finally, at mid-Bay locations the combined influence of temperature and salinity – via the salty-warm 8
water area index – is particularly well correlated with observed levels of MSXP. 9
We can examine our three-part hypothesis by inspecting disease and water properties in 2007-2009 10
when we have data on the prevalence of H. nelsoni in the upper and lower Bay regions from both 11
molecular and histological assays as well as particle transport simulations (Fig. 11 and Ford et al., Fig. 4 12
this volume). In 2007, prevalence at two lower-Bay sites where the parasite has always been enzootic 13
(New Beds and Cape Shore, see Ford et al., Figs. 1 and 4 this volume,) was moderately high. 14
Transport potential to Region E was strong and prevalence there was relatively high (Fig. 11). More 15
instructive is a comparison between 2008 and 2009. In 2008, down-bay prevalence was very high, 16
upbay transport potential was strong to upbay Regions E and F, and H. nelsoni prevalence spiked in both. 17
In contrast, H. nelsoni was not detected in either up-bay region in 2009, despite relatively strong, albeit 18
late, up-bay transport potential that year (Fig. 11). As judged by the near absence of infections at the 19
Cape Shore site during the summer and fall of 2009 and in September and November at New Beds (Ford 20
et al., this volume), we conclude that infective stages were scarce in the lower-Bay in 2009, which 21
would have limited their availability for up-bay transport even though the mechanism existed. In 22

21
addition, salinity on the upper-Bay sites was considerably lower in 2009 than in 2008 so that the 1
environment was less conducive to H. nelsoni survival and its ability to infect and proliferate in oysters. 2
Although the above comparison appears to be consistent with a scenario in which “free” infective 3
stages are moved up-bay from a lower Bay source, we urge caution in coming to such a conclusion. 4
For instance, another host, such as a weakly swimming zooplankter, could equally well have been 5
transported up-bay in the same manner and where infective stages could be released “on site”. 6
Alternatively, there may not be a single source region, but sources throughout the Bay that respond 7
similarly to factors that influence the abundance of infective particles in all areas at the same time. In 8
this regard, it is important to note that Figures 9 and 10 show relative potential transport from release 9
locations rather than absolute values. The absolute transport will depend upon the abundance of 10
particles released at a particular location. 11
Finally, the Delaware Bay model has proven to be an effective platform with which to examine past 12
environmental influences on H. nelsoni prevalence. A natural next step is to inquire into the potential 13
impacts of future climate variability on circulation, tracer fields, and MSXP in Delaware Bay. A 14
sequence of climate sensitivity studies addressing these issues is underway. 15
16

22
Acknowledgement: This study is supported by the National Science Foundation Ecology of Infectious 1
Diseases (OCE06-22672) program. The first author is also supported by a Fellowship from the DuPont 2
Corporation. Dr. David Robinson (Rutgers University), Dr. Kevin Brinson (University of Delaware), 3
and Dr. Daniel Leathers (University of Delaware) provided the DEOS solar radiation data. 4
5

23
References 1
Andrews, J. D. 1964. Oyster mortality studies in Virginia. IV. MSX in James River public seed beds. 2
Proc. Nat. Shellfish. Assoc., 53, 65-84. 3
Andrews, J. D. 1966. Oyster mortality studies in Virginia V. Epizootiology of MSX, a protistan 4
pathogen of oysters. Ecology 47:19-31. 5
Andrews, J. D. 1983. Minchinia nelsoni (MSX) infections in the James River seed-oyster area and their 6
expulsion in spring. Estuar. Coast. Shelf Sci., 16, 255-269. 7
Barber, R.D., S.A. Kanaley, and S.E. Ford. 1991. Evidence for regular sporulation by Haplosporidium 8
nelsoni (=MSX) (Ascetospora: Haplosporidiidae) in spat of the American oyster, Crassostrea virginica . 9
J. Protozool. 38(4):305-306. 10
W. P. Budgell, 2005. Numerical simulation of ice-ocean variability in the Barents Sea region: Towards 11
dynamical downscaling, Ocean Dynamics, DOI 10.1007/s10236-005-0008-3. 12
Cahill, B., O. Schofield, R. Chant, J. Wilkin, E. Hunter, S. Glenn, and P. Bissett, 2008. Dynamics of 13
turbid buoyant plumes and the feedbacks on near-shore biogeochemistry and physics, Geophys. Res. 14
Lett., . 35, L10605, doi:10.1029/2008GL033595. 15
Curchitser, E. N., D. B. Haidvogel, A. J. Hermann, E. L. Dobbins, T. M. Powell, and A. Kaplan, 2005. 16
Multi-scale modeling of the North Pacific Ocean: Assessment and analysis of simulated basin-scale 17
variability (1996–2003). J. Geophys. Res., 110, 11021, doi:101029/2005JC002902. 18
Ford, S. E. 1985. Effects of salinity on survival of the MSX parasite Haplosporidium nelsoni (Haskin, 19
Stauber, and Mackin) in oysters. J. Shellfish Res. 5:85-90 20
Ford, S. E. and H. H. Haskin. 1982. History and epizootiology of Haplosporidium nelsoni (MSX), an 21
oyster pathogen, in Delaware Bay, 1957-1980. J. Invertebr. Pathol., 40, 118-141. 22

24
Ford, S.E. and M.R. Tripp, 1996. Disease and Defense Mechanisms. In: Kennedy, V.S., R.I.E Newell, 1
and A.F. Eble, Eds. The Eastern Oyster: Crassostrea virginica . Maryland Sea 2
Ford, S.E and D. Bushek. (this volume) Development of resistance to an introduced marine pathogen by 3
a native host 4
Ford, S.E., E. Scarpa and D. Bushek, (this volume). Spatial and temporal variability in disease refuges in 5
an estuary: implications for the development of resistance. 6
Ford, S. E., B. Allam and Z. Xu (2009) Using bivalves as particle collectors and PCR detection to 7
investigate the environmental distribution of Haplosporidium nelsoni., Dis. Aquatic Org. 83:159-168. 8
Haskin, H. H. and J. D. Andrews. 1988. Uncertainties and speculations about the life cycle of the eastern 9
oyster pathogen Haplosporidium nelsoni (MSX), in Disease Processes in Marine Bivalve Molluscs, 10
Fisher, W. S., ed., American Fisheries Society, 5-22. 11
Haskin, H. H., and S. E. Ford. 1982. Haplosporidium nelsoni (MSX) on Delaware Bay seed oyster beds: 12
a host-parasite relationship along a salinity gradient. J. Invertebr. Pathol. 40:388-405. 13
Liu, Y., MacCready, P., Hickey, B. M., Dever, E. P., Kosro, P. M., Banas, N. S., 2009. Evaluation of a 14
coastal ocean circulation model for the columbia river plume in summer 2004. J. Geophys. Res. 114. 15
Marchesiello, P., J.C. McWilliams, and A. Shehepetkin (2001), Open boundary conditions for long-term 16
integration of regional ocean models, Ocean Modell., 3, 1-20. 17
Mellor,G.L. and T. Yamada (1982), Development of a turbulent closure model for geophysical fluid 18
problems. Reviews of Geophysics and Space Physics, 20, 851-875. 19
Mesinger, F., G. DiMego, E. Kalnay, K. Mitchell, P. C. Shafran, W. Ebisuzaki, D. Jovi, an, J. 20
Woollen, E. Rogers, E. H. Berbery, M. B. Ek, Y. Fan, R. Grumbine, W. Higgins, H. Li, Y. Lin, G. 21

25
Manikin, D. Parrish, and W. Shi, 2006. North American Regional Reanalysis, Bulletin of the 1
American Meteorological Society, 87(3), 343-360. 2
Mukai, A. Y., J. J. Westerink, R. A. Luettich, and D. Mark, 2002. Eastcoast 2001, A tidal constituent 3
database for the western North Atlantic, Gulf of Mexico and Caribbean Sea, Tech. Rep. ERDC/CHL 4
TR-02-24, 196 pp. 5
Paulson, C. A. and J. J. Simpson, 1977. Irradiance measuremens in the upper ocean. J. Phys. 6
Oceanogr., 7, 952-956. 7
T. M. Powell, C. V. Lewis, E. N. Curchitser, D. B. Haidvogel, A. J. Herman, E. L. Dobbins, 2006. 8
Results from a three-dimensional, nested biological-physical model of the California Current System 9
and comparisons with statistics from satellite imagery, Journal of Geophysical Research, 111, 10
doi:10.1029/2004JC002506. 11
Shchepetkin, A.F., McWilliams, J.C. (2005), The Regional Ocean Modeling System: A split-explicit, 12
free-surface, topography-following coordinate oceanic model. Ocean Modelling, 9, 347-404. 13
Smolarkiewicz, P.K. and Grabowski, W.W., 1990. The multidimensional positive definite advection 14
transport algorithm: nonoscillatory option. J. Comput. Phys. v86. 355-375. 15
Taylor, Karl E. (2001), Summarizing multiple aspects of model performance in a single diagram. J. 16
Geophys. Res., Vol.110, C11021, doi:10.1029/2005JC002902. 17
Umlauf, L., and H. Burchard (2003), A generic length-scale equation for geophysical turbulence models, 18
J. Mar. Res., 61,235-265. 19
J. C. Warner, W. R. Geyer, J. A. Lerczak, 2005. Numerical modeling of an estuary: A comprehensive 20
skill assessment, Journal of Geophysical Research, 110. C05001. doi:10.1029/2004JC002691. 21

26
Zhang, W., J. Wilkin and R. Chant, 2009. Modeling the pathways and mean dynamics of river plume 1
dispersal in New York Bight. J. Phys. Oceanogr., doi:10.1175/2008JPO4082.1, 39, 1167-1183. 2
3

27
Table 1: Parameter values that define the vertical penetrating structure for SSWR for water types 5, 6, 1 and 7. Penetration percentage as a function of depth (z) is given by the formula: 2
100*[a*exp(z/b) + (1-a)*exp(z/c)] 3 where the parameters b and c set the vertical e-folding scales (in meters) and a fixes the relative 4
amplitudes of the two exponential functions. 5 6
Penetration percentage (%) against depth (z, m) Water type 5 a = 0.78 b = 1.4 m c = 7.9 m Water type 6 a = 0.5 b = 0.01 m c = 4.9 m Water type 7 a = 0.5 b = 0.2 m c = 0.2 m
7
8
9
10 Table 2: Rationale for selection of model simulation periods based on Haplosporidium nelsoni 11 prevalence in oysters in Delaware Bay and Delaware River flow measured at Trenton, New Jersey. 12 Prevalence was assessed using tissue-section histology for the first four periods and using both histology 13 and polymerase chain reaction (PCR) for the last. 14
15
Period 1974-76 1979-81 1984-86 1990-92 2006-09
Prevalence elevated in lower
Bay, reduced in upper Bay
(Same as in 1974-76)
very high with extreme
mortality
very high on uppermost beds
in 91-92,low elsewhere
elevated on uppermost bedsin fall08-spring09
River flow average or above average
below average 80-81
drought Sep84-Aug85
below average fall91-spring92
average or above average
16
17 18

28
Table 3: Significant correlations between Haplosporidium nelsoni infection prevalence (MSXP) and 1 total river flow input from the six rivers shown in Figure 1b , salinity, and temperature at Arnolds, 2 Cohansey, New-Bennies, Egg Island, and Leased Ground locations (from upper to lower Bay, as shown 3 in Figure 8). Values for bottom salinity and temperature are averaged within a two-by-two kilometer 4 square around each of the beds. The red/blue numbers indicate the correlations with rejecting probability 5 5% or less/5~10%. The shaded numbers are the correlation coefficients between the MSXP and 6 physical conditions in the prior Spring or fall. Correlations of river flow with Fall and Spring MSXP are 7 based upon 52 and 32 yearly estimates, respectively. Correlations between salinity/temperature/SWA 8 and Fall and Spring MSXP are based upon 16 and 12 years of simulation, respectively. (Spring 9 samples of MSXP are not available for the 2006-2009 period.) 10
11 Table 3a: Correlation among 16-year river flow, simulated salinity, and temperature 12
Factor Bed
Yearly river flow and salinity
Feb river flow and temperature
Nov river flow and temperature
Dec-May salinityand temperature
Arnolds -0.85 0.57 0.43 -0.63 Cohansey -0.86 0.64 0.51 -0.63
New-Bennies -0.81 0.59 0.48 -0.68 Egg Island -0.78 0.64 0.52 -0.67
Leased Grounds -0.69 0.68 0.53 -0.58 13
Table 3b: Correlation between MSXP and water properties 14
MSX
P Factor
Bed
River flow Salinity Temperature SWA
Dec-May Jun-Nov Dec-May Jun-Nov Dec-May Jun-Nov Jun-Nov
Dec
-May
Arnolds -0.31 -0.46 0.71 0.68 -0.48
Cohansey -0.46 0.48 0.91 New-Bennies -0.52 0.59 0.92
Egg Island -0.48 0.24 0.46 0.55 Leased Grounds -0.27 0.36 0.52
Jun-
Nov
Arnolds -0.29 0.43 Cohansey -0.39 -0.25 0.42 0.44
New-Bennies -0.29 -0.24 0.43 -0.47
Egg Island -0.27 -0.45
Leased Grounds -0.46
15 16

1
2
3 4 5 6 7 8 9
Figure 1. Tlevel statiosalinity-temstations (bl
The Delawarons (numbemperature stlack dots). T
re Bay modeers); (middletations (num
The pink bou
el implemene panel) alo
mbers); and (undary lines i
29
ntation: (left ong-bay spa(right panel)indicate the
panel) crossacing (m), ) bathymetryopen bound
s-bay spacin6 river inpy (m) and a
daries.
ng (m) and 1put stations,atmospheric
2 water and 9 forcing

1
2
3
4
5
F6 a7 S8 S9 10
11
Figure 2: Nand NARR oSSWR compSystem, NAR
NARR and observing stputed as an aRR: North A
DEOS solartations; (uppaverage over
American Re
r short waveper panel) mr years 2005
egional Rean
30
e radiation (Wmean annual 5 to 2009. Dnalysis.
Wm-2): (lowvariation in
DEOS: Delaw
wer panel) lon NARR (reware Enviro
cations of thed) and DEOnmental Ob
he DEOS OS (blue) servation

1
2
3
4 5 6
Figure 3: T(below) fo
Time series oor the Arnold
of observed (ds station sho
radi
(red line) anown in Figuriation were u
31
d simulated re 1b. Waterused for this
(blue line) sr type 7 and simulation.
salinity (abov20%-reduce
ve) and temped solar shor
perature rt wave

1
2
3 4 5 6 7 8 9
10 11 12 13 14 15
FigwatJunNAdiagthe the statfrommeabarPRM
gure 4: Tayloter level at t
ne to 31 DecARR SSWR.
gram: 1) theazimuthal aradial dista
tions); 3) tm the pink pan simulatio
rs are those MS=0, and S
or diagram (the 12 statioember 2000 There are
e correlation axis; 2) the sance from ththe pattern rpoint to a “teon minus mefor stations/SB=0.
(Taylor, 200ns shown in. This simulfour basic scoefficient
standard devhe origin toroot mean sest” point; anean data sho/system bias
32
01) summarizn Figure 1, cation uses w
statistical qu(CC) betweeiation norma
o the “test” quare (PRMnd 4) the syown in the cs). For a pe
zing the mocomputed frowater type 7 uantities sumen data and alized to datpoints (colo
MS) error meystem bias (Scolorbar (nuerfect simul
del-data comom hourly dplus a 20%
mmarized in simulation i
ta (NSTD) eor number feasured by tSB, in meterumbers insidlation, CC=1
mparison fordata from 11reduction inthis Taylor
indicated onexpressed byfor differentthe distance
rs here) withde/below the1, NSTD=1,
r
n r n y t e h e ,

1
2
3
4 5 6
p7 8 9
Figure 5: Tand salinityLow-pass (
periods shor
Taylor diagra (b) at the 9 indicated byrter than/no s
co
am (explainestations sho
y squares) anshorter than olors bars ind
ed in Figure wn in Figure
nd high-pass 1 day were
dicate the sy
33
4) showing me 1, based on(circles) temfiltered in lo
ystematic bia
model-data n hourly datamperature anow/high passas (for (a), un
comparisonsa from 13 M
nd salinity arsed temperatnits of oC).
s for temperMay to 12 No
re shown. Siture and salin
ature (a) ov. 2000. ignals of nity. The

1
2 3
F4 D5 A6 17 M8
9
Figure 6: TDelaware RiArnolds, Co1974-76, 19MSXP is def
ime series oiver flow (mohansey, New79-81, 1984fined as 100
of winter/spm3/s, row 1)
w Beds-Ben4-86, 1990-9*number of
ring (Dec-Mand (rows 2
nnies, Egg I92, and 200infected oys
34
May, column2-6) HaplospIsland, and
06-09 are thsters/number
n 1) and sumporidium neLeased Gro
he five scenr of oysters s
mmer/fall (Jelsoni prevalounds. The
narios used fsampled.
Jun-Nov, colence (MSXhighlighted
for our com
olumn 2) XP, %) at d periods mparison.

1 2 3 4 5 6 7 8 9
10 11 12 13 14 15
Figure 7: Tbottom
salty-warm wscales,
temperatur0.981 for tsalinity/te
sixteen yeatemperaturdischarge.continuou
en-day-averm salinity (a,b
water area (S, black lines)re is highly cthe whole peemperature iars (1974-76re and SWA.. Within eachsly for over
raged time seb,c) averagedSWA, km2, ) at 2m abovcorrelated weriod. In (e), is higher than6, 1979-81, 1. The curvesh frame, durtwenty days
eries (left scad within bede), and the to
ve sea surfacwith the botto
the SWA (bn 17.5/12.5o
984-86, 199s of means anring yellow/gs. The blue
eac
35
ales) and the
ds, temperatuotal river dise averaged w
om temperatublue lines) reC. The anom
90-92, and 20nd anomaliegreen-shadede number in ech scenario.
eir anomalieure (oC, d) avscharge (m3/within the Bure (blue linefers to the amalies are re006-09) with
es overlap bud periods theeach frame i
s (right scaleveraged with/s, f). In (d), ay are also p
nes) with corrarea of the welative to theh seasonalityut with differe anomaly isindicates the
es) for the sihin the Bay, air temperat
plotted. The relation coef
water whose b means withy deducted orent scales fos positive/nee total mean
imulated the ture (left air fficient bottom
hin the only for for river egative within

1 2 3 4
s5 6 7 8
i9 10 11
Figure 8: shapes), and
releasing grreleased (fiv
29 May, indicates the
Locations otransport/re
rid points wive at bottom,12and 26 Ju
e releasing dbay
of major oysetention-regioithin the five, five near bone, 15 and 2epth (m) ave, B, C, D, E,
ster collectioons (separate circular areottom, at thr
29 August, 1eraged betwe, and F plus
36
on sites (blaced by straigheas (1-5, respree-hour inte2 and 26 Sepeen the two rupper bay) a
ck squares), rht lines). Thpectively). Oervals) on eaptember, andreleasing layare used to lo
released floahere are 141, On each gridach of the reld 10 Octobeyers. Six reocate floats.
ats (colored 194, 106, 59
d point ten fllease days: 1er. The coloegions (A pl
circular 9, and 50 oats are
1, 15 and or bar lus lower

1
2 3 4 5 6 7 8 9
10
11 12 13
Figure 9:near-bottofollowed in the squsix sub-re
: Particle trom release for 31 days;
uares show thegions (A-F)
ransport andsites (numb; Right: parthe percent o.
d retention pbers in browticles releaseof time spent
37
percent (valuwn circles). ed on 29 Aut by each gro
ues in color Left: part
ugust and foloup of releas
squares) froticles releasllowed for 3sed particles
om five bottosed on 1 M30 days. Ns within each
om and May and Numbers
h of the

1
2 3 4 5 6 7 8
Figure 100: Fraction (pre
percent) of teleased at eac
time spent inch of the site
38
n each of thees 1-5 (x-axi
e six sub-regis) by month
ions A-F( y-h and year.
-axis) for par
rticles

1
2 3 4 5 6 7 8 9
10 11
Figure 11histologicE and F (first 10 daRound Iscalculatedmaximum
1: Monthly Hcal data versu(green lines;ays of each land and H
d from the dm ranges of n
Haplosporidus net up-ba; see Figure month. E in
Hope Creek. different relenet up-bay pa
dium nelsoni ay particle re
8). The parncludes Arno
The error eases that aarticle reside
39
prevalence
esidence timrticle residenolds Bed andbar for a s
are done befence time pe
(%, bars) esme percent fro
nce time perd Smyrna Rspecific monfore or by thercent in that
stimated fromom lower rercent is aver
River, and Renth (June tohat month, t month.
m molecularegions to regraged withinegion F incluo Septemberrepresenting
r and gions n the udes r) is
g the