Embed Size (px)
Citation preview

Chromatin Dynamics Epigenetic
parameters amp cellular fate
cell circuits with rewiring
Geneviegraveve Almouzni
Chromatin dynamics
Mixte Unit (3664)
CNRS - Institut Curie
Paris - France
IAS HIV Cure amp Cancer Forum Broad
22 amp 23 July 2017 I Paris France
nucleosome formation
(epigenetic marks)
Histone Variants H33
Histone Modification Ac Me P Ub
DNA methylation
Binding partners HP1
Heterochromatin
Nucleosome
formation
variation
eg H312 H33
CenH3hellip
DNA methylation
Histonevariants
Histonemodifications
Nucleosome
Probst Dunleavy amp Almouzni 2009
Building chromatin domains a matter of dynamics an intimate link with DNA-based processes
Defining Chromatin landscapes
Epigenetic Landscapesrsquo after CH Waddington by Dr Paul L Harrison
High-order chromatin organization
Choice of a cell fate during development
More than
packaging
RNA
Binding Factors
HP1 Suv39
Histone variants
Histone
modifications
Nucleosome
Structural RNA
Higher-orderchromatin
Nuclearposition
Nucleus
P
DNA methylation
DNA
Histone modi cations
Histone variants
Me
Ac
Ac
Chromatin-bindingprotein
DNA
methylation
Histone
variants
Histone
modifications
Nucleosome
High-order
chromatin
Nuclear
position
Chromatin-
binding proteinRNA
Probst et al 2009
Different scales
in space and time
during cell cycle development
and in different environmentshellip
ldquoFrom signaling to epigeneticsrdquo
How to assemble maintain or change chromatin in 4D
Nucleosome
Structural RNA
Higher-orderchromatin
Nuclearposition
Nucleus
P
DNA methylation
DNA
Histone modi cations
Histone variants
Me
Ac
Ac
Chromatin-bindingprotein
DNA
methylation
Histone
variants
Histone
modifications
Nucleosome
High-order
chromatin
Nuclear
position
Chromatin-
binding proteinRNA
IV Histone modifying
enzymes
I DNA modifying enzymes
Different scales in space and time
during cell cycle development and
in different environmentshellip
ldquoFrom signaling to epigeneticsrdquo
Parameters working WITH DNA
V Chromatin remodelers
III Histone chaperones
II Histone variants
VI Chromatin-interacting
complexes (higher order)
Probst et al 2009
How to assemble maintain or change chromatin in 4DA circuit linked to genome function cell fate and its environment
Transcription factors
HP1
KMTSuv39
Targeting chromatin defects in selected solid tumors based on
oncogene addiction synthetic lethality and epigenetic antagonism
Morel D1 Almouzni G23456 Soria JC17 Postel-Vinay S17
Ann Oncol 2017
bull Epigenetic targeting is now one of the most active areas in drug
development and could represent novel therapeutic opportunity for
up to 25 of all solid tumors
bull Review of preclinical and clinical studies
In June 2016 the Financial Times titled
ldquoOncology takes the lead in exploring potential of epigeneticsrdquo
epigenetics now have a new chapter in solid tumors and clinical trials
will hopefully soon reveal the extent of its therapeutic potential
CIRCUITS ADAPTATIONS IN CANCER CELLS
Using the expression of histone chaperones(or chromatin regulators)
to better diagnose and guide clinical decisions
R Montes de Oca ZA Gurard-Levin F Berger H Rehman E Martel A Corpet L de Koning I Vassias L Wilson D Meseure F Reyal A Savignoni
B Asselain X Sastre-Garau G Almouzni
(2015) Molecular Oncology 9 657-674
The histone chaperone HJURP is a new independent
prognostic marker for luminal A breast carcinoma
Non nucleosomal protein involved in
nuclear architecture
Nucleosome
High-order
chromatin
Nuclear
position
Chromatin-
binding protein
RNA
HP1 heterochromatin protein 1
In mammals
one of the best illustration
of constitutive heterochromatin
chromocenters
DAPI
mouse ES cells
A model
Domain
the centromere
CENTROMERE
CENP-AHP1
DNA FISHmajor satellitesminor satellites
Pericentric domains enriched for
H3K9me23 H4K20me3 H2AZ DNA methylation
HP1 SENP7 structural RNA component
Guenatri JCB 2004 Maison NRMCB 2004 Probst Differentiation 2007Probst NRMCB 2009
DAPI dense heterochromatin in interphase mouse nuclei = Blocks of constitutive heterochromatin
Clusters of pericentric domains chromocenters
Remain condensed in interphasic cells
2 to 5 centric domains associated to 1 chromocenter
3T3 10 μm
(HETERO) CHROMATIN a model region next to centromere
CENTROMERE
CENP-AHP1
Guenatri JCB 2004 Maison NRMCB 2004 Probst Differentiation 2007Probst NRMCB 2009
HP1 critical for chromosome segregationEkwall et all Science 1995 Allshire et al GampD 1995 Peters et al Cell 2001 Taddei et al 2001 Obuse et al NCB2004 De Koning et al EMBO Mol Med 2010
IFHP1
DAPI dense = pericentric heterochromatin
3T310 μm
(HETERO) CHROMATIN a model region next to centromere
bull A role for the Suv39h12 HTMs since their loss is accompanied by B lymphomas(Peters AH et al Cell 2001)
bull In B lymphocytes H3K9me3 and HP-1β accumulate at pericentric heterochromatin onlyfollowing stimulation but no enrichment of H3K9me3 was observed in Suv39h dn B cells(Baxter J et al EMBO J 2004 )
Interest in the lab for the constitutive heterochromatin the Suv39h1-HP1-H3K9me3 loop
Maison et al 2010
Involvement in T cells collaboration with S Amigorena
(HETERO) CHROMATIN BASED EPIGENETICS
amp
MODULATION OF IMMUNE SYSTEM RESPONSE
A question of neighbourhood
(HETERO) CHROMATIN BASED EPIGENETICS
amp
MODULATION OF IMMUNE SYSTEM RESPONSE
Meeting with Seb hellip at Curie
Zueva Quivy Almouzni amp Amigorena 2013
A schematic view of lymphocyte differentiation
CD4+ T helperIL producing clearance ofpathogens
CD8+ cytotoxic
Zueva Quivy Almouzni and Amigorena 2013
CD4+ T helper Th1 and Th2 plasticity
Normal conditions
Th1 Interferon gamma producing (Ifng)Th2 Interleukin 4 interleukin-4 (IL-4)
H3K9me3 and H3K9ac mark the promoters of silenced and active TH lineage genes
H3K9me3 and H3K9ac ChIP and q-PCR
Allan et al 2012 Nature
loss of SUV39H1 does not affect differentiation into TH1 or TH2 cells
Naiumlve CD4+ T from wt or Suv39h1 KO cells cultured under Th1 or Th2 conditions on day 7 intracellular staining for IL4 and Ifn gamma Th1 no IL4 but Ifng
Th2 IL4 but no IFng
Allan et al 2012 Nature
SUV39H1 deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Primary culture Naive CD41 T cells from wild- type or Suv39h1-knockout (Suv39h1 KO) mice differentiated for 7 days under TH2-promoting conditions Secondary culture recultured for 2 days in TH1-promoting conditions
TH2 to TH1 plasticity is shown by the increase in the percentage of cells that wereIFN-g positive in Suv39h1 KO when compared to wild-type
Allan et al 2012 Nature
Flow cytometry
HP1a deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Similar results but with HP1a KO
Allan et al 2012 Nature
SUV39H1 deficiency results in impaired ratio of H3K9 modifications in the IFNg promoter in TH2 cells
Primary culture TH2 cells analysed by ChIP using antibodies specific for histone modifications Quantitative PCR with primers specific for the promoters of the indicated genes(in TH2 Ifn is repressed in TH1 Ifn is produced)
plasticity plasticity
Model for a heterochromatin axis (SUV39H1 dependent) to control TH2 gt TH1 plasticity
Disruption of the heterochromatin axis increases plasticity
Zueva Quivy Almouzni and Amigorena 2013
Suv39h1-sustained heterochromatin organization plays a critical role in the expansion and programming of short-lived effector CD8+ T cells (ongoing)
NEXT
Stay tuned The next episodeWith Seb
RNA
Domains
Interacting
partnersMethylated K9 of H3
H3 histone fold
RNADNAChromatin
HP1a HP1 b
p150 CAF-1 KAP1
Suv39
Dnmt1 Dnmt3a Orc1amp 3Lachner et al Nature 2001
Bannister et al Nature 2001
Nielsen et al Mol Cell 2001
Maison et al Nat Genet 2002
Muchardt et al EMBO Rep 2002 Brasher et al EMBO J 2000
Cowieson et al Curr Biol 2000
Murzina et al Mol Cell 1999
Aagaard et al EMBO J 1999
Prasanth et al PNAS 2010
Nosawa et al NCB 2010
Ronoblet et al BBRC 2011
Mouse HP1a and partners
HP1a stabilityaccumulation
at pericentric
heterochromatin
go and run
MORE TOLEARN
TAKE HOME MESSAGE letrsquos consider
the heterochromatin axis as an interesting target in
tumorigenesis - immune response and
possible role in mediated HIV latency
bull The examples of Suv39h1 and HP1a today gt more
bull An illustration with the TH2- TH1 reprogrammingndash Notion of stemness
bull Further characterization needed both for in vivo models and target
--------------------------------------------------------
bull gt how is Suv39h1 contributing ndash Novel functions for Suv39 ndash connection with Sumoylation Maison et al 2016
bull gt Potential importance of CAF-1
More to learn with this modelhellip
GENERAL CONCLUSIONS
bull The parallel development of novel epigenetic drugs and the discovery of multiple oncogenic dependencies epigenetic antagonisms and synthetic lethal relationships has open a new era in epigenetic targeting of solid tumors (CONTEXT)
bull Therapeutic targeting of epigenetic dysfunction may not be limited to the use of epigenetic drugs the role of epigenetic regulation in orchestrating other essential cellular processes such as DNA repair or immunity and connection with metabolism opens additional therapeutic opportunities (BROADER IMPACT)
bull Connections with suppression of tumor-induced inflammation suggests that approaches targeting the anti-tumor immunity or tumor micro-environment might deserve further exploration
FUTURE CHALLENGES
bull tissue- andor subunit-dependent and epigenetic antagonism or synthetic lethal relationships observed in one system may not be translatable to another gt evaluation in the tumor type of interest
bull epigenetic drugs as monotherapy for oligoclonal diseases where the epigenetic alteration serves as oncogenic driver (mostly pediatric hematologic or selected solid malignancies) in polyclonal tumors where the epigenetic alteration is a favoring permissive or secondary event combinations with conventional chemotherapies targeted therapies or immune therapies may be required
bull inhibiting the catalytic activity of some subunitsmdashsuch as EZH2mdashmight not be sufficient to reach a full anti-tumor activity important to discriminate the contribution of the non-catalytic activity of chromatin remodelers to oncogenic events
bull primary and acquired resistance will need to be studied (notably by re-biopsing and molecularly characterizing patients tumors at the occurrence of resistance) and combinatorial approaches should be considered to address this issue
bull It should be remembered that epigenetic modifications are usually reversible
The Chromatin Dynamics Group Institut Curie Paris
La vie de Chacircteau Retreat Nouan le Fuselier 2016
Institut Curie Hospital
Translational ResearchPaul Cottu
Roman Rouzier
Anne-Vincent Salomon
Sergio Roman-Roman
Elisabetta Marangoni
David Gentien
Cecile Reyes
Collaborators
Vera Pancaldi
Alfonso Valencia
Genevieve Almouzni
Team
Jean-Pierre Quivy
Dominique Ray-Gallet
Christele Maison
Guillaume Orsi
Ekaterina Boyartchuk
Delphine Bailly
Daniel Jeffrey
Yann Louault
Audrey Forest
Camille Clement
David Sitbon
Julia Torne
Katrina Podsypanina
NCI NIH
USA
Yves Pommier
Fabricio G Sousa
EP CHPEECABOO
ACKNOWLEDGMENTS
Sebastian Amigorena
Lucia Pace Rhys Allan
Elina Zueva Heidi Schreiber
Vanessa Masson
Florence Cammas team
Peter Adams
Stanford USALaura Attardi
Univ Munich
GermanyAxel Imhof
CNIO Spain
Fundacion la
scienca di vida
Chile
Alejandra
Loyola
Beatson Inst
Scotland
Sophie Postel-
Vinay amp JC Soria
Gustave Roussy

nucleosome formation
(epigenetic marks)
Histone Variants H33
Histone Modification Ac Me P Ub
DNA methylation
Binding partners HP1
Heterochromatin
Nucleosome
formation
variation
eg H312 H33
CenH3hellip
DNA methylation
Histonevariants
Histonemodifications
Nucleosome
Probst Dunleavy amp Almouzni 2009
Building chromatin domains a matter of dynamics an intimate link with DNA-based processes
Defining Chromatin landscapes
Epigenetic Landscapesrsquo after CH Waddington by Dr Paul L Harrison
High-order chromatin organization
Choice of a cell fate during development
More than
packaging
RNA
Binding Factors
HP1 Suv39
Histone variants
Histone
modifications
Nucleosome
Structural RNA
Higher-orderchromatin
Nuclearposition
Nucleus
P
DNA methylation
DNA
Histone modi cations
Histone variants
Me
Ac
Ac
Chromatin-bindingprotein
DNA
methylation
Histone
variants
Histone
modifications
Nucleosome
High-order
chromatin
Nuclear
position
Chromatin-
binding proteinRNA
Probst et al 2009
Different scales
in space and time
during cell cycle development
and in different environmentshellip
ldquoFrom signaling to epigeneticsrdquo
How to assemble maintain or change chromatin in 4D
Nucleosome
Structural RNA
Higher-orderchromatin
Nuclearposition
Nucleus
P
DNA methylation
DNA
Histone modi cations
Histone variants
Me
Ac
Ac
Chromatin-bindingprotein
DNA
methylation
Histone
variants
Histone
modifications
Nucleosome
High-order
chromatin
Nuclear
position
Chromatin-
binding proteinRNA
IV Histone modifying
enzymes
I DNA modifying enzymes
Different scales in space and time
during cell cycle development and
in different environmentshellip
ldquoFrom signaling to epigeneticsrdquo
Parameters working WITH DNA
V Chromatin remodelers
III Histone chaperones
II Histone variants
VI Chromatin-interacting
complexes (higher order)
Probst et al 2009
How to assemble maintain or change chromatin in 4DA circuit linked to genome function cell fate and its environment
Transcription factors
HP1
KMTSuv39
Targeting chromatin defects in selected solid tumors based on
oncogene addiction synthetic lethality and epigenetic antagonism
Morel D1 Almouzni G23456 Soria JC17 Postel-Vinay S17
Ann Oncol 2017
bull Epigenetic targeting is now one of the most active areas in drug
development and could represent novel therapeutic opportunity for
up to 25 of all solid tumors
bull Review of preclinical and clinical studies
In June 2016 the Financial Times titled
ldquoOncology takes the lead in exploring potential of epigeneticsrdquo
epigenetics now have a new chapter in solid tumors and clinical trials
will hopefully soon reveal the extent of its therapeutic potential
CIRCUITS ADAPTATIONS IN CANCER CELLS
Using the expression of histone chaperones(or chromatin regulators)
to better diagnose and guide clinical decisions
R Montes de Oca ZA Gurard-Levin F Berger H Rehman E Martel A Corpet L de Koning I Vassias L Wilson D Meseure F Reyal A Savignoni
B Asselain X Sastre-Garau G Almouzni
(2015) Molecular Oncology 9 657-674
The histone chaperone HJURP is a new independent
prognostic marker for luminal A breast carcinoma
Non nucleosomal protein involved in
nuclear architecture
Nucleosome
High-order
chromatin
Nuclear
position
Chromatin-
binding protein
RNA
HP1 heterochromatin protein 1
In mammals
one of the best illustration
of constitutive heterochromatin
chromocenters
DAPI
mouse ES cells
A model
Domain
the centromere
CENTROMERE
CENP-AHP1
DNA FISHmajor satellitesminor satellites
Pericentric domains enriched for
H3K9me23 H4K20me3 H2AZ DNA methylation
HP1 SENP7 structural RNA component
Guenatri JCB 2004 Maison NRMCB 2004 Probst Differentiation 2007Probst NRMCB 2009
DAPI dense heterochromatin in interphase mouse nuclei = Blocks of constitutive heterochromatin
Clusters of pericentric domains chromocenters
Remain condensed in interphasic cells
2 to 5 centric domains associated to 1 chromocenter
3T3 10 μm
(HETERO) CHROMATIN a model region next to centromere
CENTROMERE
CENP-AHP1
Guenatri JCB 2004 Maison NRMCB 2004 Probst Differentiation 2007Probst NRMCB 2009
HP1 critical for chromosome segregationEkwall et all Science 1995 Allshire et al GampD 1995 Peters et al Cell 2001 Taddei et al 2001 Obuse et al NCB2004 De Koning et al EMBO Mol Med 2010
IFHP1
DAPI dense = pericentric heterochromatin
3T310 μm
(HETERO) CHROMATIN a model region next to centromere
bull A role for the Suv39h12 HTMs since their loss is accompanied by B lymphomas(Peters AH et al Cell 2001)
bull In B lymphocytes H3K9me3 and HP-1β accumulate at pericentric heterochromatin onlyfollowing stimulation but no enrichment of H3K9me3 was observed in Suv39h dn B cells(Baxter J et al EMBO J 2004 )
Interest in the lab for the constitutive heterochromatin the Suv39h1-HP1-H3K9me3 loop
Maison et al 2010
Involvement in T cells collaboration with S Amigorena
(HETERO) CHROMATIN BASED EPIGENETICS
amp
MODULATION OF IMMUNE SYSTEM RESPONSE
A question of neighbourhood
(HETERO) CHROMATIN BASED EPIGENETICS
amp
MODULATION OF IMMUNE SYSTEM RESPONSE
Meeting with Seb hellip at Curie
Zueva Quivy Almouzni amp Amigorena 2013
A schematic view of lymphocyte differentiation
CD4+ T helperIL producing clearance ofpathogens
CD8+ cytotoxic
Zueva Quivy Almouzni and Amigorena 2013
CD4+ T helper Th1 and Th2 plasticity
Normal conditions
Th1 Interferon gamma producing (Ifng)Th2 Interleukin 4 interleukin-4 (IL-4)
H3K9me3 and H3K9ac mark the promoters of silenced and active TH lineage genes
H3K9me3 and H3K9ac ChIP and q-PCR
Allan et al 2012 Nature
loss of SUV39H1 does not affect differentiation into TH1 or TH2 cells
Naiumlve CD4+ T from wt or Suv39h1 KO cells cultured under Th1 or Th2 conditions on day 7 intracellular staining for IL4 and Ifn gamma Th1 no IL4 but Ifng
Th2 IL4 but no IFng
Allan et al 2012 Nature
SUV39H1 deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Primary culture Naive CD41 T cells from wild- type or Suv39h1-knockout (Suv39h1 KO) mice differentiated for 7 days under TH2-promoting conditions Secondary culture recultured for 2 days in TH1-promoting conditions
TH2 to TH1 plasticity is shown by the increase in the percentage of cells that wereIFN-g positive in Suv39h1 KO when compared to wild-type
Allan et al 2012 Nature
Flow cytometry
HP1a deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Similar results but with HP1a KO
Allan et al 2012 Nature
SUV39H1 deficiency results in impaired ratio of H3K9 modifications in the IFNg promoter in TH2 cells
Primary culture TH2 cells analysed by ChIP using antibodies specific for histone modifications Quantitative PCR with primers specific for the promoters of the indicated genes(in TH2 Ifn is repressed in TH1 Ifn is produced)
plasticity plasticity
Model for a heterochromatin axis (SUV39H1 dependent) to control TH2 gt TH1 plasticity
Disruption of the heterochromatin axis increases plasticity
Zueva Quivy Almouzni and Amigorena 2013
Suv39h1-sustained heterochromatin organization plays a critical role in the expansion and programming of short-lived effector CD8+ T cells (ongoing)
NEXT
Stay tuned The next episodeWith Seb
RNA
Domains
Interacting
partnersMethylated K9 of H3
H3 histone fold
RNADNAChromatin
HP1a HP1 b
p150 CAF-1 KAP1
Suv39
Dnmt1 Dnmt3a Orc1amp 3Lachner et al Nature 2001
Bannister et al Nature 2001
Nielsen et al Mol Cell 2001
Maison et al Nat Genet 2002
Muchardt et al EMBO Rep 2002 Brasher et al EMBO J 2000
Cowieson et al Curr Biol 2000
Murzina et al Mol Cell 1999
Aagaard et al EMBO J 1999
Prasanth et al PNAS 2010
Nosawa et al NCB 2010
Ronoblet et al BBRC 2011
Mouse HP1a and partners
HP1a stabilityaccumulation
at pericentric
heterochromatin
go and run
MORE TOLEARN
TAKE HOME MESSAGE letrsquos consider
the heterochromatin axis as an interesting target in
tumorigenesis - immune response and
possible role in mediated HIV latency
bull The examples of Suv39h1 and HP1a today gt more
bull An illustration with the TH2- TH1 reprogrammingndash Notion of stemness
bull Further characterization needed both for in vivo models and target
--------------------------------------------------------
bull gt how is Suv39h1 contributing ndash Novel functions for Suv39 ndash connection with Sumoylation Maison et al 2016
bull gt Potential importance of CAF-1
More to learn with this modelhellip
GENERAL CONCLUSIONS
bull The parallel development of novel epigenetic drugs and the discovery of multiple oncogenic dependencies epigenetic antagonisms and synthetic lethal relationships has open a new era in epigenetic targeting of solid tumors (CONTEXT)
bull Therapeutic targeting of epigenetic dysfunction may not be limited to the use of epigenetic drugs the role of epigenetic regulation in orchestrating other essential cellular processes such as DNA repair or immunity and connection with metabolism opens additional therapeutic opportunities (BROADER IMPACT)
bull Connections with suppression of tumor-induced inflammation suggests that approaches targeting the anti-tumor immunity or tumor micro-environment might deserve further exploration
FUTURE CHALLENGES
bull tissue- andor subunit-dependent and epigenetic antagonism or synthetic lethal relationships observed in one system may not be translatable to another gt evaluation in the tumor type of interest
bull epigenetic drugs as monotherapy for oligoclonal diseases where the epigenetic alteration serves as oncogenic driver (mostly pediatric hematologic or selected solid malignancies) in polyclonal tumors where the epigenetic alteration is a favoring permissive or secondary event combinations with conventional chemotherapies targeted therapies or immune therapies may be required
bull inhibiting the catalytic activity of some subunitsmdashsuch as EZH2mdashmight not be sufficient to reach a full anti-tumor activity important to discriminate the contribution of the non-catalytic activity of chromatin remodelers to oncogenic events
bull primary and acquired resistance will need to be studied (notably by re-biopsing and molecularly characterizing patients tumors at the occurrence of resistance) and combinatorial approaches should be considered to address this issue
bull It should be remembered that epigenetic modifications are usually reversible
The Chromatin Dynamics Group Institut Curie Paris
La vie de Chacircteau Retreat Nouan le Fuselier 2016
Institut Curie Hospital
Translational ResearchPaul Cottu
Roman Rouzier
Anne-Vincent Salomon
Sergio Roman-Roman
Elisabetta Marangoni
David Gentien
Cecile Reyes
Collaborators
Vera Pancaldi
Alfonso Valencia
Genevieve Almouzni
Team
Jean-Pierre Quivy
Dominique Ray-Gallet
Christele Maison
Guillaume Orsi
Ekaterina Boyartchuk
Delphine Bailly
Daniel Jeffrey
Yann Louault
Audrey Forest
Camille Clement
David Sitbon
Julia Torne
Katrina Podsypanina
NCI NIH
USA
Yves Pommier
Fabricio G Sousa
EP CHPEECABOO
ACKNOWLEDGMENTS
Sebastian Amigorena
Lucia Pace Rhys Allan
Elina Zueva Heidi Schreiber
Vanessa Masson
Florence Cammas team
Peter Adams
Stanford USALaura Attardi
Univ Munich
GermanyAxel Imhof
CNIO Spain
Fundacion la
scienca di vida
Chile
Alejandra
Loyola
Beatson Inst
Scotland
Sophie Postel-
Vinay amp JC Soria
Gustave Roussy

Nucleosome
Structural RNA
Higher-orderchromatin
Nuclearposition
Nucleus
P
DNA methylation
DNA
Histone modi cations
Histone variants
Me
Ac
Ac
Chromatin-bindingprotein
DNA
methylation
Histone
variants
Histone
modifications
Nucleosome
High-order
chromatin
Nuclear
position
Chromatin-
binding proteinRNA
Probst et al 2009
Different scales
in space and time
during cell cycle development
and in different environmentshellip
ldquoFrom signaling to epigeneticsrdquo
How to assemble maintain or change chromatin in 4D
Nucleosome
Structural RNA
Higher-orderchromatin
Nuclearposition
Nucleus
P
DNA methylation
DNA
Histone modi cations
Histone variants
Me
Ac
Ac
Chromatin-bindingprotein
DNA
methylation
Histone
variants
Histone
modifications
Nucleosome
High-order
chromatin
Nuclear
position
Chromatin-
binding proteinRNA
IV Histone modifying
enzymes
I DNA modifying enzymes
Different scales in space and time
during cell cycle development and
in different environmentshellip
ldquoFrom signaling to epigeneticsrdquo
Parameters working WITH DNA
V Chromatin remodelers
III Histone chaperones
II Histone variants
VI Chromatin-interacting
complexes (higher order)
Probst et al 2009
How to assemble maintain or change chromatin in 4DA circuit linked to genome function cell fate and its environment
Transcription factors
HP1
KMTSuv39
Targeting chromatin defects in selected solid tumors based on
oncogene addiction synthetic lethality and epigenetic antagonism
Morel D1 Almouzni G23456 Soria JC17 Postel-Vinay S17
Ann Oncol 2017
bull Epigenetic targeting is now one of the most active areas in drug
development and could represent novel therapeutic opportunity for
up to 25 of all solid tumors
bull Review of preclinical and clinical studies
In June 2016 the Financial Times titled
ldquoOncology takes the lead in exploring potential of epigeneticsrdquo
epigenetics now have a new chapter in solid tumors and clinical trials
will hopefully soon reveal the extent of its therapeutic potential
CIRCUITS ADAPTATIONS IN CANCER CELLS
Using the expression of histone chaperones(or chromatin regulators)
to better diagnose and guide clinical decisions
R Montes de Oca ZA Gurard-Levin F Berger H Rehman E Martel A Corpet L de Koning I Vassias L Wilson D Meseure F Reyal A Savignoni
B Asselain X Sastre-Garau G Almouzni
(2015) Molecular Oncology 9 657-674
The histone chaperone HJURP is a new independent
prognostic marker for luminal A breast carcinoma
Non nucleosomal protein involved in
nuclear architecture
Nucleosome
High-order
chromatin
Nuclear
position
Chromatin-
binding protein
RNA
HP1 heterochromatin protein 1
In mammals
one of the best illustration
of constitutive heterochromatin
chromocenters
DAPI
mouse ES cells
A model
Domain
the centromere
CENTROMERE
CENP-AHP1
DNA FISHmajor satellitesminor satellites
Pericentric domains enriched for
H3K9me23 H4K20me3 H2AZ DNA methylation
HP1 SENP7 structural RNA component
Guenatri JCB 2004 Maison NRMCB 2004 Probst Differentiation 2007Probst NRMCB 2009
DAPI dense heterochromatin in interphase mouse nuclei = Blocks of constitutive heterochromatin
Clusters of pericentric domains chromocenters
Remain condensed in interphasic cells
2 to 5 centric domains associated to 1 chromocenter
3T3 10 μm
(HETERO) CHROMATIN a model region next to centromere
CENTROMERE
CENP-AHP1
Guenatri JCB 2004 Maison NRMCB 2004 Probst Differentiation 2007Probst NRMCB 2009
HP1 critical for chromosome segregationEkwall et all Science 1995 Allshire et al GampD 1995 Peters et al Cell 2001 Taddei et al 2001 Obuse et al NCB2004 De Koning et al EMBO Mol Med 2010
IFHP1
DAPI dense = pericentric heterochromatin
3T310 μm
(HETERO) CHROMATIN a model region next to centromere
bull A role for the Suv39h12 HTMs since their loss is accompanied by B lymphomas(Peters AH et al Cell 2001)
bull In B lymphocytes H3K9me3 and HP-1β accumulate at pericentric heterochromatin onlyfollowing stimulation but no enrichment of H3K9me3 was observed in Suv39h dn B cells(Baxter J et al EMBO J 2004 )
Interest in the lab for the constitutive heterochromatin the Suv39h1-HP1-H3K9me3 loop
Maison et al 2010
Involvement in T cells collaboration with S Amigorena
(HETERO) CHROMATIN BASED EPIGENETICS
amp
MODULATION OF IMMUNE SYSTEM RESPONSE
A question of neighbourhood
(HETERO) CHROMATIN BASED EPIGENETICS
amp
MODULATION OF IMMUNE SYSTEM RESPONSE
Meeting with Seb hellip at Curie
Zueva Quivy Almouzni amp Amigorena 2013
A schematic view of lymphocyte differentiation
CD4+ T helperIL producing clearance ofpathogens
CD8+ cytotoxic
Zueva Quivy Almouzni and Amigorena 2013
CD4+ T helper Th1 and Th2 plasticity
Normal conditions
Th1 Interferon gamma producing (Ifng)Th2 Interleukin 4 interleukin-4 (IL-4)
H3K9me3 and H3K9ac mark the promoters of silenced and active TH lineage genes
H3K9me3 and H3K9ac ChIP and q-PCR
Allan et al 2012 Nature
loss of SUV39H1 does not affect differentiation into TH1 or TH2 cells
Naiumlve CD4+ T from wt or Suv39h1 KO cells cultured under Th1 or Th2 conditions on day 7 intracellular staining for IL4 and Ifn gamma Th1 no IL4 but Ifng
Th2 IL4 but no IFng
Allan et al 2012 Nature
SUV39H1 deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Primary culture Naive CD41 T cells from wild- type or Suv39h1-knockout (Suv39h1 KO) mice differentiated for 7 days under TH2-promoting conditions Secondary culture recultured for 2 days in TH1-promoting conditions
TH2 to TH1 plasticity is shown by the increase in the percentage of cells that wereIFN-g positive in Suv39h1 KO when compared to wild-type
Allan et al 2012 Nature
Flow cytometry
HP1a deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Similar results but with HP1a KO
Allan et al 2012 Nature
SUV39H1 deficiency results in impaired ratio of H3K9 modifications in the IFNg promoter in TH2 cells
Primary culture TH2 cells analysed by ChIP using antibodies specific for histone modifications Quantitative PCR with primers specific for the promoters of the indicated genes(in TH2 Ifn is repressed in TH1 Ifn is produced)
plasticity plasticity
Model for a heterochromatin axis (SUV39H1 dependent) to control TH2 gt TH1 plasticity
Disruption of the heterochromatin axis increases plasticity
Zueva Quivy Almouzni and Amigorena 2013
Suv39h1-sustained heterochromatin organization plays a critical role in the expansion and programming of short-lived effector CD8+ T cells (ongoing)
NEXT
Stay tuned The next episodeWith Seb
RNA
Domains
Interacting
partnersMethylated K9 of H3
H3 histone fold
RNADNAChromatin
HP1a HP1 b
p150 CAF-1 KAP1
Suv39
Dnmt1 Dnmt3a Orc1amp 3Lachner et al Nature 2001
Bannister et al Nature 2001
Nielsen et al Mol Cell 2001
Maison et al Nat Genet 2002
Muchardt et al EMBO Rep 2002 Brasher et al EMBO J 2000
Cowieson et al Curr Biol 2000
Murzina et al Mol Cell 1999
Aagaard et al EMBO J 1999
Prasanth et al PNAS 2010
Nosawa et al NCB 2010
Ronoblet et al BBRC 2011
Mouse HP1a and partners
HP1a stabilityaccumulation
at pericentric
heterochromatin
go and run
MORE TOLEARN
TAKE HOME MESSAGE letrsquos consider
the heterochromatin axis as an interesting target in
tumorigenesis - immune response and
possible role in mediated HIV latency
bull The examples of Suv39h1 and HP1a today gt more
bull An illustration with the TH2- TH1 reprogrammingndash Notion of stemness
bull Further characterization needed both for in vivo models and target
--------------------------------------------------------
bull gt how is Suv39h1 contributing ndash Novel functions for Suv39 ndash connection with Sumoylation Maison et al 2016
bull gt Potential importance of CAF-1
More to learn with this modelhellip
GENERAL CONCLUSIONS
bull The parallel development of novel epigenetic drugs and the discovery of multiple oncogenic dependencies epigenetic antagonisms and synthetic lethal relationships has open a new era in epigenetic targeting of solid tumors (CONTEXT)
bull Therapeutic targeting of epigenetic dysfunction may not be limited to the use of epigenetic drugs the role of epigenetic regulation in orchestrating other essential cellular processes such as DNA repair or immunity and connection with metabolism opens additional therapeutic opportunities (BROADER IMPACT)
bull Connections with suppression of tumor-induced inflammation suggests that approaches targeting the anti-tumor immunity or tumor micro-environment might deserve further exploration
FUTURE CHALLENGES
bull tissue- andor subunit-dependent and epigenetic antagonism or synthetic lethal relationships observed in one system may not be translatable to another gt evaluation in the tumor type of interest
bull epigenetic drugs as monotherapy for oligoclonal diseases where the epigenetic alteration serves as oncogenic driver (mostly pediatric hematologic or selected solid malignancies) in polyclonal tumors where the epigenetic alteration is a favoring permissive or secondary event combinations with conventional chemotherapies targeted therapies or immune therapies may be required
bull inhibiting the catalytic activity of some subunitsmdashsuch as EZH2mdashmight not be sufficient to reach a full anti-tumor activity important to discriminate the contribution of the non-catalytic activity of chromatin remodelers to oncogenic events
bull primary and acquired resistance will need to be studied (notably by re-biopsing and molecularly characterizing patients tumors at the occurrence of resistance) and combinatorial approaches should be considered to address this issue
bull It should be remembered that epigenetic modifications are usually reversible
The Chromatin Dynamics Group Institut Curie Paris
La vie de Chacircteau Retreat Nouan le Fuselier 2016
Institut Curie Hospital
Translational ResearchPaul Cottu
Roman Rouzier
Anne-Vincent Salomon
Sergio Roman-Roman
Elisabetta Marangoni
David Gentien
Cecile Reyes
Collaborators
Vera Pancaldi
Alfonso Valencia
Genevieve Almouzni
Team
Jean-Pierre Quivy
Dominique Ray-Gallet
Christele Maison
Guillaume Orsi
Ekaterina Boyartchuk
Delphine Bailly
Daniel Jeffrey
Yann Louault
Audrey Forest
Camille Clement
David Sitbon
Julia Torne
Katrina Podsypanina
NCI NIH
USA
Yves Pommier
Fabricio G Sousa
EP CHPEECABOO
ACKNOWLEDGMENTS
Sebastian Amigorena
Lucia Pace Rhys Allan
Elina Zueva Heidi Schreiber
Vanessa Masson
Florence Cammas team
Peter Adams
Stanford USALaura Attardi
Univ Munich
GermanyAxel Imhof
CNIO Spain
Fundacion la
scienca di vida
Chile
Alejandra
Loyola
Beatson Inst
Scotland
Sophie Postel-
Vinay amp JC Soria
Gustave Roussy

Nucleosome
Structural RNA
Higher-orderchromatin
Nuclearposition
Nucleus
P
DNA methylation
DNA
Histone modi cations
Histone variants
Me
Ac
Ac
Chromatin-bindingprotein
DNA
methylation
Histone
variants
Histone
modifications
Nucleosome
High-order
chromatin
Nuclear
position
Chromatin-
binding proteinRNA
IV Histone modifying
enzymes
I DNA modifying enzymes
Different scales in space and time
during cell cycle development and
in different environmentshellip
ldquoFrom signaling to epigeneticsrdquo
Parameters working WITH DNA
V Chromatin remodelers
III Histone chaperones
II Histone variants
VI Chromatin-interacting
complexes (higher order)
Probst et al 2009
How to assemble maintain or change chromatin in 4DA circuit linked to genome function cell fate and its environment
Transcription factors
HP1
KMTSuv39
Targeting chromatin defects in selected solid tumors based on
oncogene addiction synthetic lethality and epigenetic antagonism
Morel D1 Almouzni G23456 Soria JC17 Postel-Vinay S17
Ann Oncol 2017
bull Epigenetic targeting is now one of the most active areas in drug
development and could represent novel therapeutic opportunity for
up to 25 of all solid tumors
bull Review of preclinical and clinical studies
In June 2016 the Financial Times titled
ldquoOncology takes the lead in exploring potential of epigeneticsrdquo
epigenetics now have a new chapter in solid tumors and clinical trials
will hopefully soon reveal the extent of its therapeutic potential
CIRCUITS ADAPTATIONS IN CANCER CELLS
Using the expression of histone chaperones(or chromatin regulators)
to better diagnose and guide clinical decisions
R Montes de Oca ZA Gurard-Levin F Berger H Rehman E Martel A Corpet L de Koning I Vassias L Wilson D Meseure F Reyal A Savignoni
B Asselain X Sastre-Garau G Almouzni
(2015) Molecular Oncology 9 657-674
The histone chaperone HJURP is a new independent
prognostic marker for luminal A breast carcinoma
Non nucleosomal protein involved in
nuclear architecture
Nucleosome
High-order
chromatin
Nuclear
position
Chromatin-
binding protein
RNA
HP1 heterochromatin protein 1
In mammals
one of the best illustration
of constitutive heterochromatin
chromocenters
DAPI
mouse ES cells
A model
Domain
the centromere
CENTROMERE
CENP-AHP1
DNA FISHmajor satellitesminor satellites
Pericentric domains enriched for
H3K9me23 H4K20me3 H2AZ DNA methylation
HP1 SENP7 structural RNA component
Guenatri JCB 2004 Maison NRMCB 2004 Probst Differentiation 2007Probst NRMCB 2009
DAPI dense heterochromatin in interphase mouse nuclei = Blocks of constitutive heterochromatin
Clusters of pericentric domains chromocenters
Remain condensed in interphasic cells
2 to 5 centric domains associated to 1 chromocenter
3T3 10 μm
(HETERO) CHROMATIN a model region next to centromere
CENTROMERE
CENP-AHP1
Guenatri JCB 2004 Maison NRMCB 2004 Probst Differentiation 2007Probst NRMCB 2009
HP1 critical for chromosome segregationEkwall et all Science 1995 Allshire et al GampD 1995 Peters et al Cell 2001 Taddei et al 2001 Obuse et al NCB2004 De Koning et al EMBO Mol Med 2010
IFHP1
DAPI dense = pericentric heterochromatin
3T310 μm
(HETERO) CHROMATIN a model region next to centromere
bull A role for the Suv39h12 HTMs since their loss is accompanied by B lymphomas(Peters AH et al Cell 2001)
bull In B lymphocytes H3K9me3 and HP-1β accumulate at pericentric heterochromatin onlyfollowing stimulation but no enrichment of H3K9me3 was observed in Suv39h dn B cells(Baxter J et al EMBO J 2004 )
Interest in the lab for the constitutive heterochromatin the Suv39h1-HP1-H3K9me3 loop
Maison et al 2010
Involvement in T cells collaboration with S Amigorena
(HETERO) CHROMATIN BASED EPIGENETICS
amp
MODULATION OF IMMUNE SYSTEM RESPONSE
A question of neighbourhood
(HETERO) CHROMATIN BASED EPIGENETICS
amp
MODULATION OF IMMUNE SYSTEM RESPONSE
Meeting with Seb hellip at Curie
Zueva Quivy Almouzni amp Amigorena 2013
A schematic view of lymphocyte differentiation
CD4+ T helperIL producing clearance ofpathogens
CD8+ cytotoxic
Zueva Quivy Almouzni and Amigorena 2013
CD4+ T helper Th1 and Th2 plasticity
Normal conditions
Th1 Interferon gamma producing (Ifng)Th2 Interleukin 4 interleukin-4 (IL-4)
H3K9me3 and H3K9ac mark the promoters of silenced and active TH lineage genes
H3K9me3 and H3K9ac ChIP and q-PCR
Allan et al 2012 Nature
loss of SUV39H1 does not affect differentiation into TH1 or TH2 cells
Naiumlve CD4+ T from wt or Suv39h1 KO cells cultured under Th1 or Th2 conditions on day 7 intracellular staining for IL4 and Ifn gamma Th1 no IL4 but Ifng
Th2 IL4 but no IFng
Allan et al 2012 Nature
SUV39H1 deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Primary culture Naive CD41 T cells from wild- type or Suv39h1-knockout (Suv39h1 KO) mice differentiated for 7 days under TH2-promoting conditions Secondary culture recultured for 2 days in TH1-promoting conditions
TH2 to TH1 plasticity is shown by the increase in the percentage of cells that wereIFN-g positive in Suv39h1 KO when compared to wild-type
Allan et al 2012 Nature
Flow cytometry
HP1a deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Similar results but with HP1a KO
Allan et al 2012 Nature
SUV39H1 deficiency results in impaired ratio of H3K9 modifications in the IFNg promoter in TH2 cells
Primary culture TH2 cells analysed by ChIP using antibodies specific for histone modifications Quantitative PCR with primers specific for the promoters of the indicated genes(in TH2 Ifn is repressed in TH1 Ifn is produced)
plasticity plasticity
Model for a heterochromatin axis (SUV39H1 dependent) to control TH2 gt TH1 plasticity
Disruption of the heterochromatin axis increases plasticity
Zueva Quivy Almouzni and Amigorena 2013
Suv39h1-sustained heterochromatin organization plays a critical role in the expansion and programming of short-lived effector CD8+ T cells (ongoing)
NEXT
Stay tuned The next episodeWith Seb
RNA
Domains
Interacting
partnersMethylated K9 of H3
H3 histone fold
RNADNAChromatin
HP1a HP1 b
p150 CAF-1 KAP1
Suv39
Dnmt1 Dnmt3a Orc1amp 3Lachner et al Nature 2001
Bannister et al Nature 2001
Nielsen et al Mol Cell 2001
Maison et al Nat Genet 2002
Muchardt et al EMBO Rep 2002 Brasher et al EMBO J 2000
Cowieson et al Curr Biol 2000
Murzina et al Mol Cell 1999
Aagaard et al EMBO J 1999
Prasanth et al PNAS 2010
Nosawa et al NCB 2010
Ronoblet et al BBRC 2011
Mouse HP1a and partners
HP1a stabilityaccumulation
at pericentric
heterochromatin
go and run
MORE TOLEARN
TAKE HOME MESSAGE letrsquos consider
the heterochromatin axis as an interesting target in
tumorigenesis - immune response and
possible role in mediated HIV latency
bull The examples of Suv39h1 and HP1a today gt more
bull An illustration with the TH2- TH1 reprogrammingndash Notion of stemness
bull Further characterization needed both for in vivo models and target
--------------------------------------------------------
bull gt how is Suv39h1 contributing ndash Novel functions for Suv39 ndash connection with Sumoylation Maison et al 2016
bull gt Potential importance of CAF-1
More to learn with this modelhellip
GENERAL CONCLUSIONS
bull The parallel development of novel epigenetic drugs and the discovery of multiple oncogenic dependencies epigenetic antagonisms and synthetic lethal relationships has open a new era in epigenetic targeting of solid tumors (CONTEXT)
bull Therapeutic targeting of epigenetic dysfunction may not be limited to the use of epigenetic drugs the role of epigenetic regulation in orchestrating other essential cellular processes such as DNA repair or immunity and connection with metabolism opens additional therapeutic opportunities (BROADER IMPACT)
bull Connections with suppression of tumor-induced inflammation suggests that approaches targeting the anti-tumor immunity or tumor micro-environment might deserve further exploration
FUTURE CHALLENGES
bull tissue- andor subunit-dependent and epigenetic antagonism or synthetic lethal relationships observed in one system may not be translatable to another gt evaluation in the tumor type of interest
bull epigenetic drugs as monotherapy for oligoclonal diseases where the epigenetic alteration serves as oncogenic driver (mostly pediatric hematologic or selected solid malignancies) in polyclonal tumors where the epigenetic alteration is a favoring permissive or secondary event combinations with conventional chemotherapies targeted therapies or immune therapies may be required
bull inhibiting the catalytic activity of some subunitsmdashsuch as EZH2mdashmight not be sufficient to reach a full anti-tumor activity important to discriminate the contribution of the non-catalytic activity of chromatin remodelers to oncogenic events
bull primary and acquired resistance will need to be studied (notably by re-biopsing and molecularly characterizing patients tumors at the occurrence of resistance) and combinatorial approaches should be considered to address this issue
bull It should be remembered that epigenetic modifications are usually reversible
The Chromatin Dynamics Group Institut Curie Paris
La vie de Chacircteau Retreat Nouan le Fuselier 2016
Institut Curie Hospital
Translational ResearchPaul Cottu
Roman Rouzier
Anne-Vincent Salomon
Sergio Roman-Roman
Elisabetta Marangoni
David Gentien
Cecile Reyes
Collaborators
Vera Pancaldi
Alfonso Valencia
Genevieve Almouzni
Team
Jean-Pierre Quivy
Dominique Ray-Gallet
Christele Maison
Guillaume Orsi
Ekaterina Boyartchuk
Delphine Bailly
Daniel Jeffrey
Yann Louault
Audrey Forest
Camille Clement
David Sitbon
Julia Torne
Katrina Podsypanina
NCI NIH
USA
Yves Pommier
Fabricio G Sousa
EP CHPEECABOO
ACKNOWLEDGMENTS
Sebastian Amigorena
Lucia Pace Rhys Allan
Elina Zueva Heidi Schreiber
Vanessa Masson
Florence Cammas team
Peter Adams
Stanford USALaura Attardi
Univ Munich
GermanyAxel Imhof
CNIO Spain
Fundacion la
scienca di vida
Chile
Alejandra
Loyola
Beatson Inst
Scotland
Sophie Postel-
Vinay amp JC Soria
Gustave Roussy

Targeting chromatin defects in selected solid tumors based on
oncogene addiction synthetic lethality and epigenetic antagonism
Morel D1 Almouzni G23456 Soria JC17 Postel-Vinay S17
Ann Oncol 2017
bull Epigenetic targeting is now one of the most active areas in drug
development and could represent novel therapeutic opportunity for
up to 25 of all solid tumors
bull Review of preclinical and clinical studies
In June 2016 the Financial Times titled
ldquoOncology takes the lead in exploring potential of epigeneticsrdquo
epigenetics now have a new chapter in solid tumors and clinical trials
will hopefully soon reveal the extent of its therapeutic potential
CIRCUITS ADAPTATIONS IN CANCER CELLS
Using the expression of histone chaperones(or chromatin regulators)
to better diagnose and guide clinical decisions
R Montes de Oca ZA Gurard-Levin F Berger H Rehman E Martel A Corpet L de Koning I Vassias L Wilson D Meseure F Reyal A Savignoni
B Asselain X Sastre-Garau G Almouzni
(2015) Molecular Oncology 9 657-674
The histone chaperone HJURP is a new independent
prognostic marker for luminal A breast carcinoma
Non nucleosomal protein involved in
nuclear architecture
Nucleosome
High-order
chromatin
Nuclear
position
Chromatin-
binding protein
RNA
HP1 heterochromatin protein 1
In mammals
one of the best illustration
of constitutive heterochromatin
chromocenters
DAPI
mouse ES cells
A model
Domain
the centromere
CENTROMERE
CENP-AHP1
DNA FISHmajor satellitesminor satellites
Pericentric domains enriched for
H3K9me23 H4K20me3 H2AZ DNA methylation
HP1 SENP7 structural RNA component
Guenatri JCB 2004 Maison NRMCB 2004 Probst Differentiation 2007Probst NRMCB 2009
DAPI dense heterochromatin in interphase mouse nuclei = Blocks of constitutive heterochromatin
Clusters of pericentric domains chromocenters
Remain condensed in interphasic cells
2 to 5 centric domains associated to 1 chromocenter
3T3 10 μm
(HETERO) CHROMATIN a model region next to centromere
CENTROMERE
CENP-AHP1
Guenatri JCB 2004 Maison NRMCB 2004 Probst Differentiation 2007Probst NRMCB 2009
HP1 critical for chromosome segregationEkwall et all Science 1995 Allshire et al GampD 1995 Peters et al Cell 2001 Taddei et al 2001 Obuse et al NCB2004 De Koning et al EMBO Mol Med 2010
IFHP1
DAPI dense = pericentric heterochromatin
3T310 μm
(HETERO) CHROMATIN a model region next to centromere
bull A role for the Suv39h12 HTMs since their loss is accompanied by B lymphomas(Peters AH et al Cell 2001)
bull In B lymphocytes H3K9me3 and HP-1β accumulate at pericentric heterochromatin onlyfollowing stimulation but no enrichment of H3K9me3 was observed in Suv39h dn B cells(Baxter J et al EMBO J 2004 )
Interest in the lab for the constitutive heterochromatin the Suv39h1-HP1-H3K9me3 loop
Maison et al 2010
Involvement in T cells collaboration with S Amigorena
(HETERO) CHROMATIN BASED EPIGENETICS
amp
MODULATION OF IMMUNE SYSTEM RESPONSE
A question of neighbourhood
(HETERO) CHROMATIN BASED EPIGENETICS
amp
MODULATION OF IMMUNE SYSTEM RESPONSE
Meeting with Seb hellip at Curie
Zueva Quivy Almouzni amp Amigorena 2013
A schematic view of lymphocyte differentiation
CD4+ T helperIL producing clearance ofpathogens
CD8+ cytotoxic
Zueva Quivy Almouzni and Amigorena 2013
CD4+ T helper Th1 and Th2 plasticity
Normal conditions
Th1 Interferon gamma producing (Ifng)Th2 Interleukin 4 interleukin-4 (IL-4)
H3K9me3 and H3K9ac mark the promoters of silenced and active TH lineage genes
H3K9me3 and H3K9ac ChIP and q-PCR
Allan et al 2012 Nature
loss of SUV39H1 does not affect differentiation into TH1 or TH2 cells
Naiumlve CD4+ T from wt or Suv39h1 KO cells cultured under Th1 or Th2 conditions on day 7 intracellular staining for IL4 and Ifn gamma Th1 no IL4 but Ifng
Th2 IL4 but no IFng
Allan et al 2012 Nature
SUV39H1 deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Primary culture Naive CD41 T cells from wild- type or Suv39h1-knockout (Suv39h1 KO) mice differentiated for 7 days under TH2-promoting conditions Secondary culture recultured for 2 days in TH1-promoting conditions
TH2 to TH1 plasticity is shown by the increase in the percentage of cells that wereIFN-g positive in Suv39h1 KO when compared to wild-type
Allan et al 2012 Nature
Flow cytometry
HP1a deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Similar results but with HP1a KO
Allan et al 2012 Nature
SUV39H1 deficiency results in impaired ratio of H3K9 modifications in the IFNg promoter in TH2 cells
Primary culture TH2 cells analysed by ChIP using antibodies specific for histone modifications Quantitative PCR with primers specific for the promoters of the indicated genes(in TH2 Ifn is repressed in TH1 Ifn is produced)
plasticity plasticity
Model for a heterochromatin axis (SUV39H1 dependent) to control TH2 gt TH1 plasticity
Disruption of the heterochromatin axis increases plasticity
Zueva Quivy Almouzni and Amigorena 2013
Suv39h1-sustained heterochromatin organization plays a critical role in the expansion and programming of short-lived effector CD8+ T cells (ongoing)
NEXT
Stay tuned The next episodeWith Seb
RNA
Domains
Interacting
partnersMethylated K9 of H3
H3 histone fold
RNADNAChromatin
HP1a HP1 b
p150 CAF-1 KAP1
Suv39
Dnmt1 Dnmt3a Orc1amp 3Lachner et al Nature 2001
Bannister et al Nature 2001
Nielsen et al Mol Cell 2001
Maison et al Nat Genet 2002
Muchardt et al EMBO Rep 2002 Brasher et al EMBO J 2000
Cowieson et al Curr Biol 2000
Murzina et al Mol Cell 1999
Aagaard et al EMBO J 1999
Prasanth et al PNAS 2010
Nosawa et al NCB 2010
Ronoblet et al BBRC 2011
Mouse HP1a and partners
HP1a stabilityaccumulation
at pericentric
heterochromatin
go and run
MORE TOLEARN
TAKE HOME MESSAGE letrsquos consider
the heterochromatin axis as an interesting target in
tumorigenesis - immune response and
possible role in mediated HIV latency
bull The examples of Suv39h1 and HP1a today gt more
bull An illustration with the TH2- TH1 reprogrammingndash Notion of stemness
bull Further characterization needed both for in vivo models and target
--------------------------------------------------------
bull gt how is Suv39h1 contributing ndash Novel functions for Suv39 ndash connection with Sumoylation Maison et al 2016
bull gt Potential importance of CAF-1
More to learn with this modelhellip
GENERAL CONCLUSIONS
bull The parallel development of novel epigenetic drugs and the discovery of multiple oncogenic dependencies epigenetic antagonisms and synthetic lethal relationships has open a new era in epigenetic targeting of solid tumors (CONTEXT)
bull Therapeutic targeting of epigenetic dysfunction may not be limited to the use of epigenetic drugs the role of epigenetic regulation in orchestrating other essential cellular processes such as DNA repair or immunity and connection with metabolism opens additional therapeutic opportunities (BROADER IMPACT)
bull Connections with suppression of tumor-induced inflammation suggests that approaches targeting the anti-tumor immunity or tumor micro-environment might deserve further exploration
FUTURE CHALLENGES
bull tissue- andor subunit-dependent and epigenetic antagonism or synthetic lethal relationships observed in one system may not be translatable to another gt evaluation in the tumor type of interest
bull epigenetic drugs as monotherapy for oligoclonal diseases where the epigenetic alteration serves as oncogenic driver (mostly pediatric hematologic or selected solid malignancies) in polyclonal tumors where the epigenetic alteration is a favoring permissive or secondary event combinations with conventional chemotherapies targeted therapies or immune therapies may be required
bull inhibiting the catalytic activity of some subunitsmdashsuch as EZH2mdashmight not be sufficient to reach a full anti-tumor activity important to discriminate the contribution of the non-catalytic activity of chromatin remodelers to oncogenic events
bull primary and acquired resistance will need to be studied (notably by re-biopsing and molecularly characterizing patients tumors at the occurrence of resistance) and combinatorial approaches should be considered to address this issue
bull It should be remembered that epigenetic modifications are usually reversible
The Chromatin Dynamics Group Institut Curie Paris
La vie de Chacircteau Retreat Nouan le Fuselier 2016
Institut Curie Hospital
Translational ResearchPaul Cottu
Roman Rouzier
Anne-Vincent Salomon
Sergio Roman-Roman
Elisabetta Marangoni
David Gentien
Cecile Reyes
Collaborators
Vera Pancaldi
Alfonso Valencia
Genevieve Almouzni
Team
Jean-Pierre Quivy
Dominique Ray-Gallet
Christele Maison
Guillaume Orsi
Ekaterina Boyartchuk
Delphine Bailly
Daniel Jeffrey
Yann Louault
Audrey Forest
Camille Clement
David Sitbon
Julia Torne
Katrina Podsypanina
NCI NIH
USA
Yves Pommier
Fabricio G Sousa
EP CHPEECABOO
ACKNOWLEDGMENTS
Sebastian Amigorena
Lucia Pace Rhys Allan
Elina Zueva Heidi Schreiber
Vanessa Masson
Florence Cammas team
Peter Adams
Stanford USALaura Attardi
Univ Munich
GermanyAxel Imhof
CNIO Spain
Fundacion la
scienca di vida
Chile
Alejandra
Loyola
Beatson Inst
Scotland
Sophie Postel-
Vinay amp JC Soria
Gustave Roussy

Using the expression of histone chaperones(or chromatin regulators)
to better diagnose and guide clinical decisions
R Montes de Oca ZA Gurard-Levin F Berger H Rehman E Martel A Corpet L de Koning I Vassias L Wilson D Meseure F Reyal A Savignoni
B Asselain X Sastre-Garau G Almouzni
(2015) Molecular Oncology 9 657-674
The histone chaperone HJURP is a new independent
prognostic marker for luminal A breast carcinoma
Non nucleosomal protein involved in
nuclear architecture
Nucleosome
High-order
chromatin
Nuclear
position
Chromatin-
binding protein
RNA
HP1 heterochromatin protein 1
In mammals
one of the best illustration
of constitutive heterochromatin
chromocenters
DAPI
mouse ES cells
A model
Domain
the centromere
CENTROMERE
CENP-AHP1
DNA FISHmajor satellitesminor satellites
Pericentric domains enriched for
H3K9me23 H4K20me3 H2AZ DNA methylation
HP1 SENP7 structural RNA component
Guenatri JCB 2004 Maison NRMCB 2004 Probst Differentiation 2007Probst NRMCB 2009
DAPI dense heterochromatin in interphase mouse nuclei = Blocks of constitutive heterochromatin
Clusters of pericentric domains chromocenters
Remain condensed in interphasic cells
2 to 5 centric domains associated to 1 chromocenter
3T3 10 μm
(HETERO) CHROMATIN a model region next to centromere
CENTROMERE
CENP-AHP1
Guenatri JCB 2004 Maison NRMCB 2004 Probst Differentiation 2007Probst NRMCB 2009
HP1 critical for chromosome segregationEkwall et all Science 1995 Allshire et al GampD 1995 Peters et al Cell 2001 Taddei et al 2001 Obuse et al NCB2004 De Koning et al EMBO Mol Med 2010
IFHP1
DAPI dense = pericentric heterochromatin
3T310 μm
(HETERO) CHROMATIN a model region next to centromere
bull A role for the Suv39h12 HTMs since their loss is accompanied by B lymphomas(Peters AH et al Cell 2001)
bull In B lymphocytes H3K9me3 and HP-1β accumulate at pericentric heterochromatin onlyfollowing stimulation but no enrichment of H3K9me3 was observed in Suv39h dn B cells(Baxter J et al EMBO J 2004 )
Interest in the lab for the constitutive heterochromatin the Suv39h1-HP1-H3K9me3 loop
Maison et al 2010
Involvement in T cells collaboration with S Amigorena
(HETERO) CHROMATIN BASED EPIGENETICS
amp
MODULATION OF IMMUNE SYSTEM RESPONSE
A question of neighbourhood
(HETERO) CHROMATIN BASED EPIGENETICS
amp
MODULATION OF IMMUNE SYSTEM RESPONSE
Meeting with Seb hellip at Curie
Zueva Quivy Almouzni amp Amigorena 2013
A schematic view of lymphocyte differentiation
CD4+ T helperIL producing clearance ofpathogens
CD8+ cytotoxic
Zueva Quivy Almouzni and Amigorena 2013
CD4+ T helper Th1 and Th2 plasticity
Normal conditions
Th1 Interferon gamma producing (Ifng)Th2 Interleukin 4 interleukin-4 (IL-4)
H3K9me3 and H3K9ac mark the promoters of silenced and active TH lineage genes
H3K9me3 and H3K9ac ChIP and q-PCR
Allan et al 2012 Nature
loss of SUV39H1 does not affect differentiation into TH1 or TH2 cells
Naiumlve CD4+ T from wt or Suv39h1 KO cells cultured under Th1 or Th2 conditions on day 7 intracellular staining for IL4 and Ifn gamma Th1 no IL4 but Ifng
Th2 IL4 but no IFng
Allan et al 2012 Nature
SUV39H1 deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Primary culture Naive CD41 T cells from wild- type or Suv39h1-knockout (Suv39h1 KO) mice differentiated for 7 days under TH2-promoting conditions Secondary culture recultured for 2 days in TH1-promoting conditions
TH2 to TH1 plasticity is shown by the increase in the percentage of cells that wereIFN-g positive in Suv39h1 KO when compared to wild-type
Allan et al 2012 Nature
Flow cytometry
HP1a deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Similar results but with HP1a KO
Allan et al 2012 Nature
SUV39H1 deficiency results in impaired ratio of H3K9 modifications in the IFNg promoter in TH2 cells
Primary culture TH2 cells analysed by ChIP using antibodies specific for histone modifications Quantitative PCR with primers specific for the promoters of the indicated genes(in TH2 Ifn is repressed in TH1 Ifn is produced)
plasticity plasticity
Model for a heterochromatin axis (SUV39H1 dependent) to control TH2 gt TH1 plasticity
Disruption of the heterochromatin axis increases plasticity
Zueva Quivy Almouzni and Amigorena 2013
Suv39h1-sustained heterochromatin organization plays a critical role in the expansion and programming of short-lived effector CD8+ T cells (ongoing)
NEXT
Stay tuned The next episodeWith Seb
RNA
Domains
Interacting
partnersMethylated K9 of H3
H3 histone fold
RNADNAChromatin
HP1a HP1 b
p150 CAF-1 KAP1
Suv39
Dnmt1 Dnmt3a Orc1amp 3Lachner et al Nature 2001
Bannister et al Nature 2001
Nielsen et al Mol Cell 2001
Maison et al Nat Genet 2002
Muchardt et al EMBO Rep 2002 Brasher et al EMBO J 2000
Cowieson et al Curr Biol 2000
Murzina et al Mol Cell 1999
Aagaard et al EMBO J 1999
Prasanth et al PNAS 2010
Nosawa et al NCB 2010
Ronoblet et al BBRC 2011
Mouse HP1a and partners
HP1a stabilityaccumulation
at pericentric
heterochromatin
go and run
MORE TOLEARN
TAKE HOME MESSAGE letrsquos consider
the heterochromatin axis as an interesting target in
tumorigenesis - immune response and
possible role in mediated HIV latency
bull The examples of Suv39h1 and HP1a today gt more
bull An illustration with the TH2- TH1 reprogrammingndash Notion of stemness
bull Further characterization needed both for in vivo models and target
--------------------------------------------------------
bull gt how is Suv39h1 contributing ndash Novel functions for Suv39 ndash connection with Sumoylation Maison et al 2016
bull gt Potential importance of CAF-1
More to learn with this modelhellip
GENERAL CONCLUSIONS
bull The parallel development of novel epigenetic drugs and the discovery of multiple oncogenic dependencies epigenetic antagonisms and synthetic lethal relationships has open a new era in epigenetic targeting of solid tumors (CONTEXT)
bull Therapeutic targeting of epigenetic dysfunction may not be limited to the use of epigenetic drugs the role of epigenetic regulation in orchestrating other essential cellular processes such as DNA repair or immunity and connection with metabolism opens additional therapeutic opportunities (BROADER IMPACT)
bull Connections with suppression of tumor-induced inflammation suggests that approaches targeting the anti-tumor immunity or tumor micro-environment might deserve further exploration
FUTURE CHALLENGES
bull tissue- andor subunit-dependent and epigenetic antagonism or synthetic lethal relationships observed in one system may not be translatable to another gt evaluation in the tumor type of interest
bull epigenetic drugs as monotherapy for oligoclonal diseases where the epigenetic alteration serves as oncogenic driver (mostly pediatric hematologic or selected solid malignancies) in polyclonal tumors where the epigenetic alteration is a favoring permissive or secondary event combinations with conventional chemotherapies targeted therapies or immune therapies may be required
bull inhibiting the catalytic activity of some subunitsmdashsuch as EZH2mdashmight not be sufficient to reach a full anti-tumor activity important to discriminate the contribution of the non-catalytic activity of chromatin remodelers to oncogenic events
bull primary and acquired resistance will need to be studied (notably by re-biopsing and molecularly characterizing patients tumors at the occurrence of resistance) and combinatorial approaches should be considered to address this issue
bull It should be remembered that epigenetic modifications are usually reversible
The Chromatin Dynamics Group Institut Curie Paris
La vie de Chacircteau Retreat Nouan le Fuselier 2016
Institut Curie Hospital
Translational ResearchPaul Cottu
Roman Rouzier
Anne-Vincent Salomon
Sergio Roman-Roman
Elisabetta Marangoni
David Gentien
Cecile Reyes
Collaborators
Vera Pancaldi
Alfonso Valencia
Genevieve Almouzni
Team
Jean-Pierre Quivy
Dominique Ray-Gallet
Christele Maison
Guillaume Orsi
Ekaterina Boyartchuk
Delphine Bailly
Daniel Jeffrey
Yann Louault
Audrey Forest
Camille Clement
David Sitbon
Julia Torne
Katrina Podsypanina
NCI NIH
USA
Yves Pommier
Fabricio G Sousa
EP CHPEECABOO
ACKNOWLEDGMENTS
Sebastian Amigorena
Lucia Pace Rhys Allan
Elina Zueva Heidi Schreiber
Vanessa Masson
Florence Cammas team
Peter Adams
Stanford USALaura Attardi
Univ Munich
GermanyAxel Imhof
CNIO Spain
Fundacion la
scienca di vida
Chile
Alejandra
Loyola
Beatson Inst
Scotland
Sophie Postel-
Vinay amp JC Soria
Gustave Roussy

Non nucleosomal protein involved in
nuclear architecture
Nucleosome
High-order
chromatin
Nuclear
position
Chromatin-
binding protein
RNA
HP1 heterochromatin protein 1
In mammals
one of the best illustration
of constitutive heterochromatin
chromocenters
DAPI
mouse ES cells
A model
Domain
the centromere
CENTROMERE
CENP-AHP1
DNA FISHmajor satellitesminor satellites
Pericentric domains enriched for
H3K9me23 H4K20me3 H2AZ DNA methylation
HP1 SENP7 structural RNA component
Guenatri JCB 2004 Maison NRMCB 2004 Probst Differentiation 2007Probst NRMCB 2009
DAPI dense heterochromatin in interphase mouse nuclei = Blocks of constitutive heterochromatin
Clusters of pericentric domains chromocenters
Remain condensed in interphasic cells
2 to 5 centric domains associated to 1 chromocenter
3T3 10 μm
(HETERO) CHROMATIN a model region next to centromere
CENTROMERE
CENP-AHP1
Guenatri JCB 2004 Maison NRMCB 2004 Probst Differentiation 2007Probst NRMCB 2009
HP1 critical for chromosome segregationEkwall et all Science 1995 Allshire et al GampD 1995 Peters et al Cell 2001 Taddei et al 2001 Obuse et al NCB2004 De Koning et al EMBO Mol Med 2010
IFHP1
DAPI dense = pericentric heterochromatin
3T310 μm
(HETERO) CHROMATIN a model region next to centromere
bull A role for the Suv39h12 HTMs since their loss is accompanied by B lymphomas(Peters AH et al Cell 2001)
bull In B lymphocytes H3K9me3 and HP-1β accumulate at pericentric heterochromatin onlyfollowing stimulation but no enrichment of H3K9me3 was observed in Suv39h dn B cells(Baxter J et al EMBO J 2004 )
Interest in the lab for the constitutive heterochromatin the Suv39h1-HP1-H3K9me3 loop
Maison et al 2010
Involvement in T cells collaboration with S Amigorena
(HETERO) CHROMATIN BASED EPIGENETICS
amp
MODULATION OF IMMUNE SYSTEM RESPONSE
A question of neighbourhood
(HETERO) CHROMATIN BASED EPIGENETICS
amp
MODULATION OF IMMUNE SYSTEM RESPONSE
Meeting with Seb hellip at Curie
Zueva Quivy Almouzni amp Amigorena 2013
A schematic view of lymphocyte differentiation
CD4+ T helperIL producing clearance ofpathogens
CD8+ cytotoxic
Zueva Quivy Almouzni and Amigorena 2013
CD4+ T helper Th1 and Th2 plasticity
Normal conditions
Th1 Interferon gamma producing (Ifng)Th2 Interleukin 4 interleukin-4 (IL-4)
H3K9me3 and H3K9ac mark the promoters of silenced and active TH lineage genes
H3K9me3 and H3K9ac ChIP and q-PCR
Allan et al 2012 Nature
loss of SUV39H1 does not affect differentiation into TH1 or TH2 cells
Naiumlve CD4+ T from wt or Suv39h1 KO cells cultured under Th1 or Th2 conditions on day 7 intracellular staining for IL4 and Ifn gamma Th1 no IL4 but Ifng
Th2 IL4 but no IFng
Allan et al 2012 Nature
SUV39H1 deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Primary culture Naive CD41 T cells from wild- type or Suv39h1-knockout (Suv39h1 KO) mice differentiated for 7 days under TH2-promoting conditions Secondary culture recultured for 2 days in TH1-promoting conditions
TH2 to TH1 plasticity is shown by the increase in the percentage of cells that wereIFN-g positive in Suv39h1 KO when compared to wild-type
Allan et al 2012 Nature
Flow cytometry
HP1a deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Similar results but with HP1a KO
Allan et al 2012 Nature
SUV39H1 deficiency results in impaired ratio of H3K9 modifications in the IFNg promoter in TH2 cells
Primary culture TH2 cells analysed by ChIP using antibodies specific for histone modifications Quantitative PCR with primers specific for the promoters of the indicated genes(in TH2 Ifn is repressed in TH1 Ifn is produced)
plasticity plasticity
Model for a heterochromatin axis (SUV39H1 dependent) to control TH2 gt TH1 plasticity
Disruption of the heterochromatin axis increases plasticity
Zueva Quivy Almouzni and Amigorena 2013
Suv39h1-sustained heterochromatin organization plays a critical role in the expansion and programming of short-lived effector CD8+ T cells (ongoing)
NEXT
Stay tuned The next episodeWith Seb
RNA
Domains
Interacting
partnersMethylated K9 of H3
H3 histone fold
RNADNAChromatin
HP1a HP1 b
p150 CAF-1 KAP1
Suv39
Dnmt1 Dnmt3a Orc1amp 3Lachner et al Nature 2001
Bannister et al Nature 2001
Nielsen et al Mol Cell 2001
Maison et al Nat Genet 2002
Muchardt et al EMBO Rep 2002 Brasher et al EMBO J 2000
Cowieson et al Curr Biol 2000
Murzina et al Mol Cell 1999
Aagaard et al EMBO J 1999
Prasanth et al PNAS 2010
Nosawa et al NCB 2010
Ronoblet et al BBRC 2011
Mouse HP1a and partners
HP1a stabilityaccumulation
at pericentric
heterochromatin
go and run
MORE TOLEARN
TAKE HOME MESSAGE letrsquos consider
the heterochromatin axis as an interesting target in
tumorigenesis - immune response and
possible role in mediated HIV latency
bull The examples of Suv39h1 and HP1a today gt more
bull An illustration with the TH2- TH1 reprogrammingndash Notion of stemness
bull Further characterization needed both for in vivo models and target
--------------------------------------------------------
bull gt how is Suv39h1 contributing ndash Novel functions for Suv39 ndash connection with Sumoylation Maison et al 2016
bull gt Potential importance of CAF-1
More to learn with this modelhellip
GENERAL CONCLUSIONS
bull The parallel development of novel epigenetic drugs and the discovery of multiple oncogenic dependencies epigenetic antagonisms and synthetic lethal relationships has open a new era in epigenetic targeting of solid tumors (CONTEXT)
bull Therapeutic targeting of epigenetic dysfunction may not be limited to the use of epigenetic drugs the role of epigenetic regulation in orchestrating other essential cellular processes such as DNA repair or immunity and connection with metabolism opens additional therapeutic opportunities (BROADER IMPACT)
bull Connections with suppression of tumor-induced inflammation suggests that approaches targeting the anti-tumor immunity or tumor micro-environment might deserve further exploration
FUTURE CHALLENGES
bull tissue- andor subunit-dependent and epigenetic antagonism or synthetic lethal relationships observed in one system may not be translatable to another gt evaluation in the tumor type of interest
bull epigenetic drugs as monotherapy for oligoclonal diseases where the epigenetic alteration serves as oncogenic driver (mostly pediatric hematologic or selected solid malignancies) in polyclonal tumors where the epigenetic alteration is a favoring permissive or secondary event combinations with conventional chemotherapies targeted therapies or immune therapies may be required
bull inhibiting the catalytic activity of some subunitsmdashsuch as EZH2mdashmight not be sufficient to reach a full anti-tumor activity important to discriminate the contribution of the non-catalytic activity of chromatin remodelers to oncogenic events
bull primary and acquired resistance will need to be studied (notably by re-biopsing and molecularly characterizing patients tumors at the occurrence of resistance) and combinatorial approaches should be considered to address this issue
bull It should be remembered that epigenetic modifications are usually reversible
The Chromatin Dynamics Group Institut Curie Paris
La vie de Chacircteau Retreat Nouan le Fuselier 2016
Institut Curie Hospital
Translational ResearchPaul Cottu
Roman Rouzier
Anne-Vincent Salomon
Sergio Roman-Roman
Elisabetta Marangoni
David Gentien
Cecile Reyes
Collaborators
Vera Pancaldi
Alfonso Valencia
Genevieve Almouzni
Team
Jean-Pierre Quivy
Dominique Ray-Gallet
Christele Maison
Guillaume Orsi
Ekaterina Boyartchuk
Delphine Bailly
Daniel Jeffrey
Yann Louault
Audrey Forest
Camille Clement
David Sitbon
Julia Torne
Katrina Podsypanina
NCI NIH
USA
Yves Pommier
Fabricio G Sousa
EP CHPEECABOO
ACKNOWLEDGMENTS
Sebastian Amigorena
Lucia Pace Rhys Allan
Elina Zueva Heidi Schreiber
Vanessa Masson
Florence Cammas team
Peter Adams
Stanford USALaura Attardi
Univ Munich
GermanyAxel Imhof
CNIO Spain
Fundacion la
scienca di vida
Chile
Alejandra
Loyola
Beatson Inst
Scotland
Sophie Postel-
Vinay amp JC Soria
Gustave Roussy

CENTROMERE
CENP-AHP1
DNA FISHmajor satellitesminor satellites
Pericentric domains enriched for
H3K9me23 H4K20me3 H2AZ DNA methylation
HP1 SENP7 structural RNA component
Guenatri JCB 2004 Maison NRMCB 2004 Probst Differentiation 2007Probst NRMCB 2009
DAPI dense heterochromatin in interphase mouse nuclei = Blocks of constitutive heterochromatin
Clusters of pericentric domains chromocenters
Remain condensed in interphasic cells
2 to 5 centric domains associated to 1 chromocenter
3T3 10 μm
(HETERO) CHROMATIN a model region next to centromere
CENTROMERE
CENP-AHP1
Guenatri JCB 2004 Maison NRMCB 2004 Probst Differentiation 2007Probst NRMCB 2009
HP1 critical for chromosome segregationEkwall et all Science 1995 Allshire et al GampD 1995 Peters et al Cell 2001 Taddei et al 2001 Obuse et al NCB2004 De Koning et al EMBO Mol Med 2010
IFHP1
DAPI dense = pericentric heterochromatin
3T310 μm
(HETERO) CHROMATIN a model region next to centromere
bull A role for the Suv39h12 HTMs since their loss is accompanied by B lymphomas(Peters AH et al Cell 2001)
bull In B lymphocytes H3K9me3 and HP-1β accumulate at pericentric heterochromatin onlyfollowing stimulation but no enrichment of H3K9me3 was observed in Suv39h dn B cells(Baxter J et al EMBO J 2004 )
Interest in the lab for the constitutive heterochromatin the Suv39h1-HP1-H3K9me3 loop
Maison et al 2010
Involvement in T cells collaboration with S Amigorena
(HETERO) CHROMATIN BASED EPIGENETICS
amp
MODULATION OF IMMUNE SYSTEM RESPONSE
A question of neighbourhood
(HETERO) CHROMATIN BASED EPIGENETICS
amp
MODULATION OF IMMUNE SYSTEM RESPONSE
Meeting with Seb hellip at Curie
Zueva Quivy Almouzni amp Amigorena 2013
A schematic view of lymphocyte differentiation
CD4+ T helperIL producing clearance ofpathogens
CD8+ cytotoxic
Zueva Quivy Almouzni and Amigorena 2013
CD4+ T helper Th1 and Th2 plasticity
Normal conditions
Th1 Interferon gamma producing (Ifng)Th2 Interleukin 4 interleukin-4 (IL-4)
H3K9me3 and H3K9ac mark the promoters of silenced and active TH lineage genes
H3K9me3 and H3K9ac ChIP and q-PCR
Allan et al 2012 Nature
loss of SUV39H1 does not affect differentiation into TH1 or TH2 cells
Naiumlve CD4+ T from wt or Suv39h1 KO cells cultured under Th1 or Th2 conditions on day 7 intracellular staining for IL4 and Ifn gamma Th1 no IL4 but Ifng
Th2 IL4 but no IFng
Allan et al 2012 Nature
SUV39H1 deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Primary culture Naive CD41 T cells from wild- type or Suv39h1-knockout (Suv39h1 KO) mice differentiated for 7 days under TH2-promoting conditions Secondary culture recultured for 2 days in TH1-promoting conditions
TH2 to TH1 plasticity is shown by the increase in the percentage of cells that wereIFN-g positive in Suv39h1 KO when compared to wild-type
Allan et al 2012 Nature
Flow cytometry
HP1a deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Similar results but with HP1a KO
Allan et al 2012 Nature
SUV39H1 deficiency results in impaired ratio of H3K9 modifications in the IFNg promoter in TH2 cells
Primary culture TH2 cells analysed by ChIP using antibodies specific for histone modifications Quantitative PCR with primers specific for the promoters of the indicated genes(in TH2 Ifn is repressed in TH1 Ifn is produced)
plasticity plasticity
Model for a heterochromatin axis (SUV39H1 dependent) to control TH2 gt TH1 plasticity
Disruption of the heterochromatin axis increases plasticity
Zueva Quivy Almouzni and Amigorena 2013
Suv39h1-sustained heterochromatin organization plays a critical role in the expansion and programming of short-lived effector CD8+ T cells (ongoing)
NEXT
Stay tuned The next episodeWith Seb
RNA
Domains
Interacting
partnersMethylated K9 of H3
H3 histone fold
RNADNAChromatin
HP1a HP1 b
p150 CAF-1 KAP1
Suv39
Dnmt1 Dnmt3a Orc1amp 3Lachner et al Nature 2001
Bannister et al Nature 2001
Nielsen et al Mol Cell 2001
Maison et al Nat Genet 2002
Muchardt et al EMBO Rep 2002 Brasher et al EMBO J 2000
Cowieson et al Curr Biol 2000
Murzina et al Mol Cell 1999
Aagaard et al EMBO J 1999
Prasanth et al PNAS 2010
Nosawa et al NCB 2010
Ronoblet et al BBRC 2011
Mouse HP1a and partners
HP1a stabilityaccumulation
at pericentric
heterochromatin
go and run
MORE TOLEARN
TAKE HOME MESSAGE letrsquos consider
the heterochromatin axis as an interesting target in
tumorigenesis - immune response and
possible role in mediated HIV latency
bull The examples of Suv39h1 and HP1a today gt more
bull An illustration with the TH2- TH1 reprogrammingndash Notion of stemness
bull Further characterization needed both for in vivo models and target
--------------------------------------------------------
bull gt how is Suv39h1 contributing ndash Novel functions for Suv39 ndash connection with Sumoylation Maison et al 2016
bull gt Potential importance of CAF-1
More to learn with this modelhellip
GENERAL CONCLUSIONS
bull The parallel development of novel epigenetic drugs and the discovery of multiple oncogenic dependencies epigenetic antagonisms and synthetic lethal relationships has open a new era in epigenetic targeting of solid tumors (CONTEXT)
bull Therapeutic targeting of epigenetic dysfunction may not be limited to the use of epigenetic drugs the role of epigenetic regulation in orchestrating other essential cellular processes such as DNA repair or immunity and connection with metabolism opens additional therapeutic opportunities (BROADER IMPACT)
bull Connections with suppression of tumor-induced inflammation suggests that approaches targeting the anti-tumor immunity or tumor micro-environment might deserve further exploration
FUTURE CHALLENGES
bull tissue- andor subunit-dependent and epigenetic antagonism or synthetic lethal relationships observed in one system may not be translatable to another gt evaluation in the tumor type of interest
bull epigenetic drugs as monotherapy for oligoclonal diseases where the epigenetic alteration serves as oncogenic driver (mostly pediatric hematologic or selected solid malignancies) in polyclonal tumors where the epigenetic alteration is a favoring permissive or secondary event combinations with conventional chemotherapies targeted therapies or immune therapies may be required
bull inhibiting the catalytic activity of some subunitsmdashsuch as EZH2mdashmight not be sufficient to reach a full anti-tumor activity important to discriminate the contribution of the non-catalytic activity of chromatin remodelers to oncogenic events
bull primary and acquired resistance will need to be studied (notably by re-biopsing and molecularly characterizing patients tumors at the occurrence of resistance) and combinatorial approaches should be considered to address this issue
bull It should be remembered that epigenetic modifications are usually reversible
The Chromatin Dynamics Group Institut Curie Paris
La vie de Chacircteau Retreat Nouan le Fuselier 2016
Institut Curie Hospital
Translational ResearchPaul Cottu
Roman Rouzier
Anne-Vincent Salomon
Sergio Roman-Roman
Elisabetta Marangoni
David Gentien
Cecile Reyes
Collaborators
Vera Pancaldi
Alfonso Valencia
Genevieve Almouzni
Team
Jean-Pierre Quivy
Dominique Ray-Gallet
Christele Maison
Guillaume Orsi
Ekaterina Boyartchuk
Delphine Bailly
Daniel Jeffrey
Yann Louault
Audrey Forest
Camille Clement
David Sitbon
Julia Torne
Katrina Podsypanina
NCI NIH
USA
Yves Pommier
Fabricio G Sousa
EP CHPEECABOO
ACKNOWLEDGMENTS
Sebastian Amigorena
Lucia Pace Rhys Allan
Elina Zueva Heidi Schreiber
Vanessa Masson
Florence Cammas team
Peter Adams
Stanford USALaura Attardi
Univ Munich
GermanyAxel Imhof
CNIO Spain
Fundacion la
scienca di vida
Chile
Alejandra
Loyola
Beatson Inst
Scotland
Sophie Postel-
Vinay amp JC Soria
Gustave Roussy

CENTROMERE
CENP-AHP1
Guenatri JCB 2004 Maison NRMCB 2004 Probst Differentiation 2007Probst NRMCB 2009
HP1 critical for chromosome segregationEkwall et all Science 1995 Allshire et al GampD 1995 Peters et al Cell 2001 Taddei et al 2001 Obuse et al NCB2004 De Koning et al EMBO Mol Med 2010
IFHP1
DAPI dense = pericentric heterochromatin
3T310 μm
(HETERO) CHROMATIN a model region next to centromere
bull A role for the Suv39h12 HTMs since their loss is accompanied by B lymphomas(Peters AH et al Cell 2001)
bull In B lymphocytes H3K9me3 and HP-1β accumulate at pericentric heterochromatin onlyfollowing stimulation but no enrichment of H3K9me3 was observed in Suv39h dn B cells(Baxter J et al EMBO J 2004 )
Interest in the lab for the constitutive heterochromatin the Suv39h1-HP1-H3K9me3 loop
Maison et al 2010
Involvement in T cells collaboration with S Amigorena
(HETERO) CHROMATIN BASED EPIGENETICS
amp
MODULATION OF IMMUNE SYSTEM RESPONSE
A question of neighbourhood
(HETERO) CHROMATIN BASED EPIGENETICS
amp
MODULATION OF IMMUNE SYSTEM RESPONSE
Meeting with Seb hellip at Curie
Zueva Quivy Almouzni amp Amigorena 2013
A schematic view of lymphocyte differentiation
CD4+ T helperIL producing clearance ofpathogens
CD8+ cytotoxic
Zueva Quivy Almouzni and Amigorena 2013
CD4+ T helper Th1 and Th2 plasticity
Normal conditions
Th1 Interferon gamma producing (Ifng)Th2 Interleukin 4 interleukin-4 (IL-4)
H3K9me3 and H3K9ac mark the promoters of silenced and active TH lineage genes
H3K9me3 and H3K9ac ChIP and q-PCR
Allan et al 2012 Nature
loss of SUV39H1 does not affect differentiation into TH1 or TH2 cells
Naiumlve CD4+ T from wt or Suv39h1 KO cells cultured under Th1 or Th2 conditions on day 7 intracellular staining for IL4 and Ifn gamma Th1 no IL4 but Ifng
Th2 IL4 but no IFng
Allan et al 2012 Nature
SUV39H1 deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Primary culture Naive CD41 T cells from wild- type or Suv39h1-knockout (Suv39h1 KO) mice differentiated for 7 days under TH2-promoting conditions Secondary culture recultured for 2 days in TH1-promoting conditions
TH2 to TH1 plasticity is shown by the increase in the percentage of cells that wereIFN-g positive in Suv39h1 KO when compared to wild-type
Allan et al 2012 Nature
Flow cytometry
HP1a deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Similar results but with HP1a KO
Allan et al 2012 Nature
SUV39H1 deficiency results in impaired ratio of H3K9 modifications in the IFNg promoter in TH2 cells
Primary culture TH2 cells analysed by ChIP using antibodies specific for histone modifications Quantitative PCR with primers specific for the promoters of the indicated genes(in TH2 Ifn is repressed in TH1 Ifn is produced)
plasticity plasticity
Model for a heterochromatin axis (SUV39H1 dependent) to control TH2 gt TH1 plasticity
Disruption of the heterochromatin axis increases plasticity
Zueva Quivy Almouzni and Amigorena 2013
Suv39h1-sustained heterochromatin organization plays a critical role in the expansion and programming of short-lived effector CD8+ T cells (ongoing)
NEXT
Stay tuned The next episodeWith Seb
RNA
Domains
Interacting
partnersMethylated K9 of H3
H3 histone fold
RNADNAChromatin
HP1a HP1 b
p150 CAF-1 KAP1
Suv39
Dnmt1 Dnmt3a Orc1amp 3Lachner et al Nature 2001
Bannister et al Nature 2001
Nielsen et al Mol Cell 2001
Maison et al Nat Genet 2002
Muchardt et al EMBO Rep 2002 Brasher et al EMBO J 2000
Cowieson et al Curr Biol 2000
Murzina et al Mol Cell 1999
Aagaard et al EMBO J 1999
Prasanth et al PNAS 2010
Nosawa et al NCB 2010
Ronoblet et al BBRC 2011
Mouse HP1a and partners
HP1a stabilityaccumulation
at pericentric
heterochromatin
go and run
MORE TOLEARN
TAKE HOME MESSAGE letrsquos consider
the heterochromatin axis as an interesting target in
tumorigenesis - immune response and
possible role in mediated HIV latency
bull The examples of Suv39h1 and HP1a today gt more
bull An illustration with the TH2- TH1 reprogrammingndash Notion of stemness
bull Further characterization needed both for in vivo models and target
--------------------------------------------------------
bull gt how is Suv39h1 contributing ndash Novel functions for Suv39 ndash connection with Sumoylation Maison et al 2016
bull gt Potential importance of CAF-1
More to learn with this modelhellip
GENERAL CONCLUSIONS
bull The parallel development of novel epigenetic drugs and the discovery of multiple oncogenic dependencies epigenetic antagonisms and synthetic lethal relationships has open a new era in epigenetic targeting of solid tumors (CONTEXT)
bull Therapeutic targeting of epigenetic dysfunction may not be limited to the use of epigenetic drugs the role of epigenetic regulation in orchestrating other essential cellular processes such as DNA repair or immunity and connection with metabolism opens additional therapeutic opportunities (BROADER IMPACT)
bull Connections with suppression of tumor-induced inflammation suggests that approaches targeting the anti-tumor immunity or tumor micro-environment might deserve further exploration
FUTURE CHALLENGES
bull tissue- andor subunit-dependent and epigenetic antagonism or synthetic lethal relationships observed in one system may not be translatable to another gt evaluation in the tumor type of interest
bull epigenetic drugs as monotherapy for oligoclonal diseases where the epigenetic alteration serves as oncogenic driver (mostly pediatric hematologic or selected solid malignancies) in polyclonal tumors where the epigenetic alteration is a favoring permissive or secondary event combinations with conventional chemotherapies targeted therapies or immune therapies may be required
bull inhibiting the catalytic activity of some subunitsmdashsuch as EZH2mdashmight not be sufficient to reach a full anti-tumor activity important to discriminate the contribution of the non-catalytic activity of chromatin remodelers to oncogenic events
bull primary and acquired resistance will need to be studied (notably by re-biopsing and molecularly characterizing patients tumors at the occurrence of resistance) and combinatorial approaches should be considered to address this issue
bull It should be remembered that epigenetic modifications are usually reversible
The Chromatin Dynamics Group Institut Curie Paris
La vie de Chacircteau Retreat Nouan le Fuselier 2016
Institut Curie Hospital
Translational ResearchPaul Cottu
Roman Rouzier
Anne-Vincent Salomon
Sergio Roman-Roman
Elisabetta Marangoni
David Gentien
Cecile Reyes
Collaborators
Vera Pancaldi
Alfonso Valencia
Genevieve Almouzni
Team
Jean-Pierre Quivy
Dominique Ray-Gallet
Christele Maison
Guillaume Orsi
Ekaterina Boyartchuk
Delphine Bailly
Daniel Jeffrey
Yann Louault
Audrey Forest
Camille Clement
David Sitbon
Julia Torne
Katrina Podsypanina
NCI NIH
USA
Yves Pommier
Fabricio G Sousa
EP CHPEECABOO
ACKNOWLEDGMENTS
Sebastian Amigorena
Lucia Pace Rhys Allan
Elina Zueva Heidi Schreiber
Vanessa Masson
Florence Cammas team
Peter Adams
Stanford USALaura Attardi
Univ Munich
GermanyAxel Imhof
CNIO Spain
Fundacion la
scienca di vida
Chile
Alejandra
Loyola
Beatson Inst
Scotland
Sophie Postel-
Vinay amp JC Soria
Gustave Roussy

bull A role for the Suv39h12 HTMs since their loss is accompanied by B lymphomas(Peters AH et al Cell 2001)
bull In B lymphocytes H3K9me3 and HP-1β accumulate at pericentric heterochromatin onlyfollowing stimulation but no enrichment of H3K9me3 was observed in Suv39h dn B cells(Baxter J et al EMBO J 2004 )
Interest in the lab for the constitutive heterochromatin the Suv39h1-HP1-H3K9me3 loop
Maison et al 2010
Involvement in T cells collaboration with S Amigorena
(HETERO) CHROMATIN BASED EPIGENETICS
amp
MODULATION OF IMMUNE SYSTEM RESPONSE
A question of neighbourhood
(HETERO) CHROMATIN BASED EPIGENETICS
amp
MODULATION OF IMMUNE SYSTEM RESPONSE
Meeting with Seb hellip at Curie
Zueva Quivy Almouzni amp Amigorena 2013
A schematic view of lymphocyte differentiation
CD4+ T helperIL producing clearance ofpathogens
CD8+ cytotoxic
Zueva Quivy Almouzni and Amigorena 2013
CD4+ T helper Th1 and Th2 plasticity
Normal conditions
Th1 Interferon gamma producing (Ifng)Th2 Interleukin 4 interleukin-4 (IL-4)
H3K9me3 and H3K9ac mark the promoters of silenced and active TH lineage genes
H3K9me3 and H3K9ac ChIP and q-PCR
Allan et al 2012 Nature
loss of SUV39H1 does not affect differentiation into TH1 or TH2 cells
Naiumlve CD4+ T from wt or Suv39h1 KO cells cultured under Th1 or Th2 conditions on day 7 intracellular staining for IL4 and Ifn gamma Th1 no IL4 but Ifng
Th2 IL4 but no IFng
Allan et al 2012 Nature
SUV39H1 deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Primary culture Naive CD41 T cells from wild- type or Suv39h1-knockout (Suv39h1 KO) mice differentiated for 7 days under TH2-promoting conditions Secondary culture recultured for 2 days in TH1-promoting conditions
TH2 to TH1 plasticity is shown by the increase in the percentage of cells that wereIFN-g positive in Suv39h1 KO when compared to wild-type
Allan et al 2012 Nature
Flow cytometry
HP1a deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Similar results but with HP1a KO
Allan et al 2012 Nature
SUV39H1 deficiency results in impaired ratio of H3K9 modifications in the IFNg promoter in TH2 cells
Primary culture TH2 cells analysed by ChIP using antibodies specific for histone modifications Quantitative PCR with primers specific for the promoters of the indicated genes(in TH2 Ifn is repressed in TH1 Ifn is produced)
plasticity plasticity
Model for a heterochromatin axis (SUV39H1 dependent) to control TH2 gt TH1 plasticity
Disruption of the heterochromatin axis increases plasticity
Zueva Quivy Almouzni and Amigorena 2013
Suv39h1-sustained heterochromatin organization plays a critical role in the expansion and programming of short-lived effector CD8+ T cells (ongoing)
NEXT
Stay tuned The next episodeWith Seb
RNA
Domains
Interacting
partnersMethylated K9 of H3
H3 histone fold
RNADNAChromatin
HP1a HP1 b
p150 CAF-1 KAP1
Suv39
Dnmt1 Dnmt3a Orc1amp 3Lachner et al Nature 2001
Bannister et al Nature 2001
Nielsen et al Mol Cell 2001
Maison et al Nat Genet 2002
Muchardt et al EMBO Rep 2002 Brasher et al EMBO J 2000
Cowieson et al Curr Biol 2000
Murzina et al Mol Cell 1999
Aagaard et al EMBO J 1999
Prasanth et al PNAS 2010
Nosawa et al NCB 2010
Ronoblet et al BBRC 2011
Mouse HP1a and partners
HP1a stabilityaccumulation
at pericentric
heterochromatin
go and run
MORE TOLEARN
TAKE HOME MESSAGE letrsquos consider
the heterochromatin axis as an interesting target in
tumorigenesis - immune response and
possible role in mediated HIV latency
bull The examples of Suv39h1 and HP1a today gt more
bull An illustration with the TH2- TH1 reprogrammingndash Notion of stemness
bull Further characterization needed both for in vivo models and target
--------------------------------------------------------
bull gt how is Suv39h1 contributing ndash Novel functions for Suv39 ndash connection with Sumoylation Maison et al 2016
bull gt Potential importance of CAF-1
More to learn with this modelhellip
GENERAL CONCLUSIONS
bull The parallel development of novel epigenetic drugs and the discovery of multiple oncogenic dependencies epigenetic antagonisms and synthetic lethal relationships has open a new era in epigenetic targeting of solid tumors (CONTEXT)
bull Therapeutic targeting of epigenetic dysfunction may not be limited to the use of epigenetic drugs the role of epigenetic regulation in orchestrating other essential cellular processes such as DNA repair or immunity and connection with metabolism opens additional therapeutic opportunities (BROADER IMPACT)
bull Connections with suppression of tumor-induced inflammation suggests that approaches targeting the anti-tumor immunity or tumor micro-environment might deserve further exploration
FUTURE CHALLENGES
bull tissue- andor subunit-dependent and epigenetic antagonism or synthetic lethal relationships observed in one system may not be translatable to another gt evaluation in the tumor type of interest
bull epigenetic drugs as monotherapy for oligoclonal diseases where the epigenetic alteration serves as oncogenic driver (mostly pediatric hematologic or selected solid malignancies) in polyclonal tumors where the epigenetic alteration is a favoring permissive or secondary event combinations with conventional chemotherapies targeted therapies or immune therapies may be required
bull inhibiting the catalytic activity of some subunitsmdashsuch as EZH2mdashmight not be sufficient to reach a full anti-tumor activity important to discriminate the contribution of the non-catalytic activity of chromatin remodelers to oncogenic events
bull primary and acquired resistance will need to be studied (notably by re-biopsing and molecularly characterizing patients tumors at the occurrence of resistance) and combinatorial approaches should be considered to address this issue
bull It should be remembered that epigenetic modifications are usually reversible
The Chromatin Dynamics Group Institut Curie Paris
La vie de Chacircteau Retreat Nouan le Fuselier 2016
Institut Curie Hospital
Translational ResearchPaul Cottu
Roman Rouzier
Anne-Vincent Salomon
Sergio Roman-Roman
Elisabetta Marangoni
David Gentien
Cecile Reyes
Collaborators
Vera Pancaldi
Alfonso Valencia
Genevieve Almouzni
Team
Jean-Pierre Quivy
Dominique Ray-Gallet
Christele Maison
Guillaume Orsi
Ekaterina Boyartchuk
Delphine Bailly
Daniel Jeffrey
Yann Louault
Audrey Forest
Camille Clement
David Sitbon
Julia Torne
Katrina Podsypanina
NCI NIH
USA
Yves Pommier
Fabricio G Sousa
EP CHPEECABOO
ACKNOWLEDGMENTS
Sebastian Amigorena
Lucia Pace Rhys Allan
Elina Zueva Heidi Schreiber
Vanessa Masson
Florence Cammas team
Peter Adams
Stanford USALaura Attardi
Univ Munich
GermanyAxel Imhof
CNIO Spain
Fundacion la
scienca di vida
Chile
Alejandra
Loyola
Beatson Inst
Scotland
Sophie Postel-
Vinay amp JC Soria
Gustave Roussy

A question of neighbourhood
(HETERO) CHROMATIN BASED EPIGENETICS
amp
MODULATION OF IMMUNE SYSTEM RESPONSE
Meeting with Seb hellip at Curie
Zueva Quivy Almouzni amp Amigorena 2013
A schematic view of lymphocyte differentiation
CD4+ T helperIL producing clearance ofpathogens
CD8+ cytotoxic
Zueva Quivy Almouzni and Amigorena 2013
CD4+ T helper Th1 and Th2 plasticity
Normal conditions
Th1 Interferon gamma producing (Ifng)Th2 Interleukin 4 interleukin-4 (IL-4)
H3K9me3 and H3K9ac mark the promoters of silenced and active TH lineage genes
H3K9me3 and H3K9ac ChIP and q-PCR
Allan et al 2012 Nature
loss of SUV39H1 does not affect differentiation into TH1 or TH2 cells
Naiumlve CD4+ T from wt or Suv39h1 KO cells cultured under Th1 or Th2 conditions on day 7 intracellular staining for IL4 and Ifn gamma Th1 no IL4 but Ifng
Th2 IL4 but no IFng
Allan et al 2012 Nature
SUV39H1 deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Primary culture Naive CD41 T cells from wild- type or Suv39h1-knockout (Suv39h1 KO) mice differentiated for 7 days under TH2-promoting conditions Secondary culture recultured for 2 days in TH1-promoting conditions
TH2 to TH1 plasticity is shown by the increase in the percentage of cells that wereIFN-g positive in Suv39h1 KO when compared to wild-type
Allan et al 2012 Nature
Flow cytometry
HP1a deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Similar results but with HP1a KO
Allan et al 2012 Nature
SUV39H1 deficiency results in impaired ratio of H3K9 modifications in the IFNg promoter in TH2 cells
Primary culture TH2 cells analysed by ChIP using antibodies specific for histone modifications Quantitative PCR with primers specific for the promoters of the indicated genes(in TH2 Ifn is repressed in TH1 Ifn is produced)
plasticity plasticity
Model for a heterochromatin axis (SUV39H1 dependent) to control TH2 gt TH1 plasticity
Disruption of the heterochromatin axis increases plasticity
Zueva Quivy Almouzni and Amigorena 2013
Suv39h1-sustained heterochromatin organization plays a critical role in the expansion and programming of short-lived effector CD8+ T cells (ongoing)
NEXT
Stay tuned The next episodeWith Seb
RNA
Domains
Interacting
partnersMethylated K9 of H3
H3 histone fold
RNADNAChromatin
HP1a HP1 b
p150 CAF-1 KAP1
Suv39
Dnmt1 Dnmt3a Orc1amp 3Lachner et al Nature 2001
Bannister et al Nature 2001
Nielsen et al Mol Cell 2001
Maison et al Nat Genet 2002
Muchardt et al EMBO Rep 2002 Brasher et al EMBO J 2000
Cowieson et al Curr Biol 2000
Murzina et al Mol Cell 1999
Aagaard et al EMBO J 1999
Prasanth et al PNAS 2010
Nosawa et al NCB 2010
Ronoblet et al BBRC 2011
Mouse HP1a and partners
HP1a stabilityaccumulation
at pericentric
heterochromatin
go and run
MORE TOLEARN
TAKE HOME MESSAGE letrsquos consider
the heterochromatin axis as an interesting target in
tumorigenesis - immune response and
possible role in mediated HIV latency
bull The examples of Suv39h1 and HP1a today gt more
bull An illustration with the TH2- TH1 reprogrammingndash Notion of stemness
bull Further characterization needed both for in vivo models and target
--------------------------------------------------------
bull gt how is Suv39h1 contributing ndash Novel functions for Suv39 ndash connection with Sumoylation Maison et al 2016
bull gt Potential importance of CAF-1
More to learn with this modelhellip
GENERAL CONCLUSIONS
bull The parallel development of novel epigenetic drugs and the discovery of multiple oncogenic dependencies epigenetic antagonisms and synthetic lethal relationships has open a new era in epigenetic targeting of solid tumors (CONTEXT)
bull Therapeutic targeting of epigenetic dysfunction may not be limited to the use of epigenetic drugs the role of epigenetic regulation in orchestrating other essential cellular processes such as DNA repair or immunity and connection with metabolism opens additional therapeutic opportunities (BROADER IMPACT)
bull Connections with suppression of tumor-induced inflammation suggests that approaches targeting the anti-tumor immunity or tumor micro-environment might deserve further exploration
FUTURE CHALLENGES
bull tissue- andor subunit-dependent and epigenetic antagonism or synthetic lethal relationships observed in one system may not be translatable to another gt evaluation in the tumor type of interest
bull epigenetic drugs as monotherapy for oligoclonal diseases where the epigenetic alteration serves as oncogenic driver (mostly pediatric hematologic or selected solid malignancies) in polyclonal tumors where the epigenetic alteration is a favoring permissive or secondary event combinations with conventional chemotherapies targeted therapies or immune therapies may be required
bull inhibiting the catalytic activity of some subunitsmdashsuch as EZH2mdashmight not be sufficient to reach a full anti-tumor activity important to discriminate the contribution of the non-catalytic activity of chromatin remodelers to oncogenic events
bull primary and acquired resistance will need to be studied (notably by re-biopsing and molecularly characterizing patients tumors at the occurrence of resistance) and combinatorial approaches should be considered to address this issue
bull It should be remembered that epigenetic modifications are usually reversible
The Chromatin Dynamics Group Institut Curie Paris
La vie de Chacircteau Retreat Nouan le Fuselier 2016
Institut Curie Hospital
Translational ResearchPaul Cottu
Roman Rouzier
Anne-Vincent Salomon
Sergio Roman-Roman
Elisabetta Marangoni
David Gentien
Cecile Reyes
Collaborators
Vera Pancaldi
Alfonso Valencia
Genevieve Almouzni
Team
Jean-Pierre Quivy
Dominique Ray-Gallet
Christele Maison
Guillaume Orsi
Ekaterina Boyartchuk
Delphine Bailly
Daniel Jeffrey
Yann Louault
Audrey Forest
Camille Clement
David Sitbon
Julia Torne
Katrina Podsypanina
NCI NIH
USA
Yves Pommier
Fabricio G Sousa
EP CHPEECABOO
ACKNOWLEDGMENTS
Sebastian Amigorena
Lucia Pace Rhys Allan
Elina Zueva Heidi Schreiber
Vanessa Masson
Florence Cammas team
Peter Adams
Stanford USALaura Attardi
Univ Munich
GermanyAxel Imhof
CNIO Spain
Fundacion la
scienca di vida
Chile
Alejandra
Loyola
Beatson Inst
Scotland
Sophie Postel-
Vinay amp JC Soria
Gustave Roussy

Zueva Quivy Almouzni amp Amigorena 2013
A schematic view of lymphocyte differentiation
CD4+ T helperIL producing clearance ofpathogens
CD8+ cytotoxic
Zueva Quivy Almouzni and Amigorena 2013
CD4+ T helper Th1 and Th2 plasticity
Normal conditions
Th1 Interferon gamma producing (Ifng)Th2 Interleukin 4 interleukin-4 (IL-4)
H3K9me3 and H3K9ac mark the promoters of silenced and active TH lineage genes
H3K9me3 and H3K9ac ChIP and q-PCR
Allan et al 2012 Nature
loss of SUV39H1 does not affect differentiation into TH1 or TH2 cells
Naiumlve CD4+ T from wt or Suv39h1 KO cells cultured under Th1 or Th2 conditions on day 7 intracellular staining for IL4 and Ifn gamma Th1 no IL4 but Ifng
Th2 IL4 but no IFng
Allan et al 2012 Nature
SUV39H1 deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Primary culture Naive CD41 T cells from wild- type or Suv39h1-knockout (Suv39h1 KO) mice differentiated for 7 days under TH2-promoting conditions Secondary culture recultured for 2 days in TH1-promoting conditions
TH2 to TH1 plasticity is shown by the increase in the percentage of cells that wereIFN-g positive in Suv39h1 KO when compared to wild-type
Allan et al 2012 Nature
Flow cytometry
HP1a deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Similar results but with HP1a KO
Allan et al 2012 Nature
SUV39H1 deficiency results in impaired ratio of H3K9 modifications in the IFNg promoter in TH2 cells
Primary culture TH2 cells analysed by ChIP using antibodies specific for histone modifications Quantitative PCR with primers specific for the promoters of the indicated genes(in TH2 Ifn is repressed in TH1 Ifn is produced)
plasticity plasticity
Model for a heterochromatin axis (SUV39H1 dependent) to control TH2 gt TH1 plasticity
Disruption of the heterochromatin axis increases plasticity
Zueva Quivy Almouzni and Amigorena 2013
Suv39h1-sustained heterochromatin organization plays a critical role in the expansion and programming of short-lived effector CD8+ T cells (ongoing)
NEXT
Stay tuned The next episodeWith Seb
RNA
Domains
Interacting
partnersMethylated K9 of H3
H3 histone fold
RNADNAChromatin
HP1a HP1 b
p150 CAF-1 KAP1
Suv39
Dnmt1 Dnmt3a Orc1amp 3Lachner et al Nature 2001
Bannister et al Nature 2001
Nielsen et al Mol Cell 2001
Maison et al Nat Genet 2002
Muchardt et al EMBO Rep 2002 Brasher et al EMBO J 2000
Cowieson et al Curr Biol 2000
Murzina et al Mol Cell 1999
Aagaard et al EMBO J 1999
Prasanth et al PNAS 2010
Nosawa et al NCB 2010
Ronoblet et al BBRC 2011
Mouse HP1a and partners
HP1a stabilityaccumulation
at pericentric
heterochromatin
go and run
MORE TOLEARN
TAKE HOME MESSAGE letrsquos consider
the heterochromatin axis as an interesting target in
tumorigenesis - immune response and
possible role in mediated HIV latency
bull The examples of Suv39h1 and HP1a today gt more
bull An illustration with the TH2- TH1 reprogrammingndash Notion of stemness
bull Further characterization needed both for in vivo models and target
--------------------------------------------------------
bull gt how is Suv39h1 contributing ndash Novel functions for Suv39 ndash connection with Sumoylation Maison et al 2016
bull gt Potential importance of CAF-1
More to learn with this modelhellip
GENERAL CONCLUSIONS
bull The parallel development of novel epigenetic drugs and the discovery of multiple oncogenic dependencies epigenetic antagonisms and synthetic lethal relationships has open a new era in epigenetic targeting of solid tumors (CONTEXT)
bull Therapeutic targeting of epigenetic dysfunction may not be limited to the use of epigenetic drugs the role of epigenetic regulation in orchestrating other essential cellular processes such as DNA repair or immunity and connection with metabolism opens additional therapeutic opportunities (BROADER IMPACT)
bull Connections with suppression of tumor-induced inflammation suggests that approaches targeting the anti-tumor immunity or tumor micro-environment might deserve further exploration
FUTURE CHALLENGES
bull tissue- andor subunit-dependent and epigenetic antagonism or synthetic lethal relationships observed in one system may not be translatable to another gt evaluation in the tumor type of interest
bull epigenetic drugs as monotherapy for oligoclonal diseases where the epigenetic alteration serves as oncogenic driver (mostly pediatric hematologic or selected solid malignancies) in polyclonal tumors where the epigenetic alteration is a favoring permissive or secondary event combinations with conventional chemotherapies targeted therapies or immune therapies may be required
bull inhibiting the catalytic activity of some subunitsmdashsuch as EZH2mdashmight not be sufficient to reach a full anti-tumor activity important to discriminate the contribution of the non-catalytic activity of chromatin remodelers to oncogenic events
bull primary and acquired resistance will need to be studied (notably by re-biopsing and molecularly characterizing patients tumors at the occurrence of resistance) and combinatorial approaches should be considered to address this issue
bull It should be remembered that epigenetic modifications are usually reversible
The Chromatin Dynamics Group Institut Curie Paris
La vie de Chacircteau Retreat Nouan le Fuselier 2016
Institut Curie Hospital
Translational ResearchPaul Cottu
Roman Rouzier
Anne-Vincent Salomon
Sergio Roman-Roman
Elisabetta Marangoni
David Gentien
Cecile Reyes
Collaborators
Vera Pancaldi
Alfonso Valencia
Genevieve Almouzni
Team
Jean-Pierre Quivy
Dominique Ray-Gallet
Christele Maison
Guillaume Orsi
Ekaterina Boyartchuk
Delphine Bailly
Daniel Jeffrey
Yann Louault
Audrey Forest
Camille Clement
David Sitbon
Julia Torne
Katrina Podsypanina
NCI NIH
USA
Yves Pommier
Fabricio G Sousa
EP CHPEECABOO
ACKNOWLEDGMENTS
Sebastian Amigorena
Lucia Pace Rhys Allan
Elina Zueva Heidi Schreiber
Vanessa Masson
Florence Cammas team
Peter Adams
Stanford USALaura Attardi
Univ Munich
GermanyAxel Imhof
CNIO Spain
Fundacion la
scienca di vida
Chile
Alejandra
Loyola
Beatson Inst
Scotland
Sophie Postel-
Vinay amp JC Soria
Gustave Roussy

Zueva Quivy Almouzni and Amigorena 2013
CD4+ T helper Th1 and Th2 plasticity
Normal conditions
Th1 Interferon gamma producing (Ifng)Th2 Interleukin 4 interleukin-4 (IL-4)
H3K9me3 and H3K9ac mark the promoters of silenced and active TH lineage genes
H3K9me3 and H3K9ac ChIP and q-PCR
Allan et al 2012 Nature
loss of SUV39H1 does not affect differentiation into TH1 or TH2 cells
Naiumlve CD4+ T from wt or Suv39h1 KO cells cultured under Th1 or Th2 conditions on day 7 intracellular staining for IL4 and Ifn gamma Th1 no IL4 but Ifng
Th2 IL4 but no IFng
Allan et al 2012 Nature
SUV39H1 deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Primary culture Naive CD41 T cells from wild- type or Suv39h1-knockout (Suv39h1 KO) mice differentiated for 7 days under TH2-promoting conditions Secondary culture recultured for 2 days in TH1-promoting conditions
TH2 to TH1 plasticity is shown by the increase in the percentage of cells that wereIFN-g positive in Suv39h1 KO when compared to wild-type
Allan et al 2012 Nature
Flow cytometry
HP1a deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Similar results but with HP1a KO
Allan et al 2012 Nature
SUV39H1 deficiency results in impaired ratio of H3K9 modifications in the IFNg promoter in TH2 cells
Primary culture TH2 cells analysed by ChIP using antibodies specific for histone modifications Quantitative PCR with primers specific for the promoters of the indicated genes(in TH2 Ifn is repressed in TH1 Ifn is produced)
plasticity plasticity
Model for a heterochromatin axis (SUV39H1 dependent) to control TH2 gt TH1 plasticity
Disruption of the heterochromatin axis increases plasticity
Zueva Quivy Almouzni and Amigorena 2013
Suv39h1-sustained heterochromatin organization plays a critical role in the expansion and programming of short-lived effector CD8+ T cells (ongoing)
NEXT
Stay tuned The next episodeWith Seb
RNA
Domains
Interacting
partnersMethylated K9 of H3
H3 histone fold
RNADNAChromatin
HP1a HP1 b
p150 CAF-1 KAP1
Suv39
Dnmt1 Dnmt3a Orc1amp 3Lachner et al Nature 2001
Bannister et al Nature 2001
Nielsen et al Mol Cell 2001
Maison et al Nat Genet 2002
Muchardt et al EMBO Rep 2002 Brasher et al EMBO J 2000
Cowieson et al Curr Biol 2000
Murzina et al Mol Cell 1999
Aagaard et al EMBO J 1999
Prasanth et al PNAS 2010
Nosawa et al NCB 2010
Ronoblet et al BBRC 2011
Mouse HP1a and partners
HP1a stabilityaccumulation
at pericentric
heterochromatin
go and run
MORE TOLEARN
TAKE HOME MESSAGE letrsquos consider
the heterochromatin axis as an interesting target in
tumorigenesis - immune response and
possible role in mediated HIV latency
bull The examples of Suv39h1 and HP1a today gt more
bull An illustration with the TH2- TH1 reprogrammingndash Notion of stemness
bull Further characterization needed both for in vivo models and target
--------------------------------------------------------
bull gt how is Suv39h1 contributing ndash Novel functions for Suv39 ndash connection with Sumoylation Maison et al 2016
bull gt Potential importance of CAF-1
More to learn with this modelhellip
GENERAL CONCLUSIONS
bull The parallel development of novel epigenetic drugs and the discovery of multiple oncogenic dependencies epigenetic antagonisms and synthetic lethal relationships has open a new era in epigenetic targeting of solid tumors (CONTEXT)
bull Therapeutic targeting of epigenetic dysfunction may not be limited to the use of epigenetic drugs the role of epigenetic regulation in orchestrating other essential cellular processes such as DNA repair or immunity and connection with metabolism opens additional therapeutic opportunities (BROADER IMPACT)
bull Connections with suppression of tumor-induced inflammation suggests that approaches targeting the anti-tumor immunity or tumor micro-environment might deserve further exploration
FUTURE CHALLENGES
bull tissue- andor subunit-dependent and epigenetic antagonism or synthetic lethal relationships observed in one system may not be translatable to another gt evaluation in the tumor type of interest
bull epigenetic drugs as monotherapy for oligoclonal diseases where the epigenetic alteration serves as oncogenic driver (mostly pediatric hematologic or selected solid malignancies) in polyclonal tumors where the epigenetic alteration is a favoring permissive or secondary event combinations with conventional chemotherapies targeted therapies or immune therapies may be required
bull inhibiting the catalytic activity of some subunitsmdashsuch as EZH2mdashmight not be sufficient to reach a full anti-tumor activity important to discriminate the contribution of the non-catalytic activity of chromatin remodelers to oncogenic events
bull primary and acquired resistance will need to be studied (notably by re-biopsing and molecularly characterizing patients tumors at the occurrence of resistance) and combinatorial approaches should be considered to address this issue
bull It should be remembered that epigenetic modifications are usually reversible
The Chromatin Dynamics Group Institut Curie Paris
La vie de Chacircteau Retreat Nouan le Fuselier 2016
Institut Curie Hospital
Translational ResearchPaul Cottu
Roman Rouzier
Anne-Vincent Salomon
Sergio Roman-Roman
Elisabetta Marangoni
David Gentien
Cecile Reyes
Collaborators
Vera Pancaldi
Alfonso Valencia
Genevieve Almouzni
Team
Jean-Pierre Quivy
Dominique Ray-Gallet
Christele Maison
Guillaume Orsi
Ekaterina Boyartchuk
Delphine Bailly
Daniel Jeffrey
Yann Louault
Audrey Forest
Camille Clement
David Sitbon
Julia Torne
Katrina Podsypanina
NCI NIH
USA
Yves Pommier
Fabricio G Sousa
EP CHPEECABOO
ACKNOWLEDGMENTS
Sebastian Amigorena
Lucia Pace Rhys Allan
Elina Zueva Heidi Schreiber
Vanessa Masson
Florence Cammas team
Peter Adams
Stanford USALaura Attardi
Univ Munich
GermanyAxel Imhof
CNIO Spain
Fundacion la
scienca di vida
Chile
Alejandra
Loyola
Beatson Inst
Scotland
Sophie Postel-
Vinay amp JC Soria
Gustave Roussy

Normal conditions
Th1 Interferon gamma producing (Ifng)Th2 Interleukin 4 interleukin-4 (IL-4)
H3K9me3 and H3K9ac mark the promoters of silenced and active TH lineage genes
H3K9me3 and H3K9ac ChIP and q-PCR
Allan et al 2012 Nature
loss of SUV39H1 does not affect differentiation into TH1 or TH2 cells
Naiumlve CD4+ T from wt or Suv39h1 KO cells cultured under Th1 or Th2 conditions on day 7 intracellular staining for IL4 and Ifn gamma Th1 no IL4 but Ifng
Th2 IL4 but no IFng
Allan et al 2012 Nature
SUV39H1 deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Primary culture Naive CD41 T cells from wild- type or Suv39h1-knockout (Suv39h1 KO) mice differentiated for 7 days under TH2-promoting conditions Secondary culture recultured for 2 days in TH1-promoting conditions
TH2 to TH1 plasticity is shown by the increase in the percentage of cells that wereIFN-g positive in Suv39h1 KO when compared to wild-type
Allan et al 2012 Nature
Flow cytometry
HP1a deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Similar results but with HP1a KO
Allan et al 2012 Nature
SUV39H1 deficiency results in impaired ratio of H3K9 modifications in the IFNg promoter in TH2 cells
Primary culture TH2 cells analysed by ChIP using antibodies specific for histone modifications Quantitative PCR with primers specific for the promoters of the indicated genes(in TH2 Ifn is repressed in TH1 Ifn is produced)
plasticity plasticity
Model for a heterochromatin axis (SUV39H1 dependent) to control TH2 gt TH1 plasticity
Disruption of the heterochromatin axis increases plasticity
Zueva Quivy Almouzni and Amigorena 2013
Suv39h1-sustained heterochromatin organization plays a critical role in the expansion and programming of short-lived effector CD8+ T cells (ongoing)
NEXT
Stay tuned The next episodeWith Seb
RNA
Domains
Interacting
partnersMethylated K9 of H3
H3 histone fold
RNADNAChromatin
HP1a HP1 b
p150 CAF-1 KAP1
Suv39
Dnmt1 Dnmt3a Orc1amp 3Lachner et al Nature 2001
Bannister et al Nature 2001
Nielsen et al Mol Cell 2001
Maison et al Nat Genet 2002
Muchardt et al EMBO Rep 2002 Brasher et al EMBO J 2000
Cowieson et al Curr Biol 2000
Murzina et al Mol Cell 1999
Aagaard et al EMBO J 1999
Prasanth et al PNAS 2010
Nosawa et al NCB 2010
Ronoblet et al BBRC 2011
Mouse HP1a and partners
HP1a stabilityaccumulation
at pericentric
heterochromatin
go and run
MORE TOLEARN
TAKE HOME MESSAGE letrsquos consider
the heterochromatin axis as an interesting target in
tumorigenesis - immune response and
possible role in mediated HIV latency
bull The examples of Suv39h1 and HP1a today gt more
bull An illustration with the TH2- TH1 reprogrammingndash Notion of stemness
bull Further characterization needed both for in vivo models and target
--------------------------------------------------------
bull gt how is Suv39h1 contributing ndash Novel functions for Suv39 ndash connection with Sumoylation Maison et al 2016
bull gt Potential importance of CAF-1
More to learn with this modelhellip
GENERAL CONCLUSIONS
bull The parallel development of novel epigenetic drugs and the discovery of multiple oncogenic dependencies epigenetic antagonisms and synthetic lethal relationships has open a new era in epigenetic targeting of solid tumors (CONTEXT)
bull Therapeutic targeting of epigenetic dysfunction may not be limited to the use of epigenetic drugs the role of epigenetic regulation in orchestrating other essential cellular processes such as DNA repair or immunity and connection with metabolism opens additional therapeutic opportunities (BROADER IMPACT)
bull Connections with suppression of tumor-induced inflammation suggests that approaches targeting the anti-tumor immunity or tumor micro-environment might deserve further exploration
FUTURE CHALLENGES
bull tissue- andor subunit-dependent and epigenetic antagonism or synthetic lethal relationships observed in one system may not be translatable to another gt evaluation in the tumor type of interest
bull epigenetic drugs as monotherapy for oligoclonal diseases where the epigenetic alteration serves as oncogenic driver (mostly pediatric hematologic or selected solid malignancies) in polyclonal tumors where the epigenetic alteration is a favoring permissive or secondary event combinations with conventional chemotherapies targeted therapies or immune therapies may be required
bull inhibiting the catalytic activity of some subunitsmdashsuch as EZH2mdashmight not be sufficient to reach a full anti-tumor activity important to discriminate the contribution of the non-catalytic activity of chromatin remodelers to oncogenic events
bull primary and acquired resistance will need to be studied (notably by re-biopsing and molecularly characterizing patients tumors at the occurrence of resistance) and combinatorial approaches should be considered to address this issue
bull It should be remembered that epigenetic modifications are usually reversible
The Chromatin Dynamics Group Institut Curie Paris
La vie de Chacircteau Retreat Nouan le Fuselier 2016
Institut Curie Hospital
Translational ResearchPaul Cottu
Roman Rouzier
Anne-Vincent Salomon
Sergio Roman-Roman
Elisabetta Marangoni
David Gentien
Cecile Reyes
Collaborators
Vera Pancaldi
Alfonso Valencia
Genevieve Almouzni
Team
Jean-Pierre Quivy
Dominique Ray-Gallet
Christele Maison
Guillaume Orsi
Ekaterina Boyartchuk
Delphine Bailly
Daniel Jeffrey
Yann Louault
Audrey Forest
Camille Clement
David Sitbon
Julia Torne
Katrina Podsypanina
NCI NIH
USA
Yves Pommier
Fabricio G Sousa
EP CHPEECABOO
ACKNOWLEDGMENTS
Sebastian Amigorena
Lucia Pace Rhys Allan
Elina Zueva Heidi Schreiber
Vanessa Masson
Florence Cammas team
Peter Adams
Stanford USALaura Attardi
Univ Munich
GermanyAxel Imhof
CNIO Spain
Fundacion la
scienca di vida
Chile
Alejandra
Loyola
Beatson Inst
Scotland
Sophie Postel-
Vinay amp JC Soria
Gustave Roussy

H3K9me3 and H3K9ac mark the promoters of silenced and active TH lineage genes
H3K9me3 and H3K9ac ChIP and q-PCR
Allan et al 2012 Nature
loss of SUV39H1 does not affect differentiation into TH1 or TH2 cells
Naiumlve CD4+ T from wt or Suv39h1 KO cells cultured under Th1 or Th2 conditions on day 7 intracellular staining for IL4 and Ifn gamma Th1 no IL4 but Ifng
Th2 IL4 but no IFng
Allan et al 2012 Nature
SUV39H1 deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Primary culture Naive CD41 T cells from wild- type or Suv39h1-knockout (Suv39h1 KO) mice differentiated for 7 days under TH2-promoting conditions Secondary culture recultured for 2 days in TH1-promoting conditions
TH2 to TH1 plasticity is shown by the increase in the percentage of cells that wereIFN-g positive in Suv39h1 KO when compared to wild-type
Allan et al 2012 Nature
Flow cytometry
HP1a deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Similar results but with HP1a KO
Allan et al 2012 Nature
SUV39H1 deficiency results in impaired ratio of H3K9 modifications in the IFNg promoter in TH2 cells
Primary culture TH2 cells analysed by ChIP using antibodies specific for histone modifications Quantitative PCR with primers specific for the promoters of the indicated genes(in TH2 Ifn is repressed in TH1 Ifn is produced)
plasticity plasticity
Model for a heterochromatin axis (SUV39H1 dependent) to control TH2 gt TH1 plasticity
Disruption of the heterochromatin axis increases plasticity
Zueva Quivy Almouzni and Amigorena 2013
Suv39h1-sustained heterochromatin organization plays a critical role in the expansion and programming of short-lived effector CD8+ T cells (ongoing)
NEXT
Stay tuned The next episodeWith Seb
RNA
Domains
Interacting
partnersMethylated K9 of H3
H3 histone fold
RNADNAChromatin
HP1a HP1 b
p150 CAF-1 KAP1
Suv39
Dnmt1 Dnmt3a Orc1amp 3Lachner et al Nature 2001
Bannister et al Nature 2001
Nielsen et al Mol Cell 2001
Maison et al Nat Genet 2002
Muchardt et al EMBO Rep 2002 Brasher et al EMBO J 2000
Cowieson et al Curr Biol 2000
Murzina et al Mol Cell 1999
Aagaard et al EMBO J 1999
Prasanth et al PNAS 2010
Nosawa et al NCB 2010
Ronoblet et al BBRC 2011
Mouse HP1a and partners
HP1a stabilityaccumulation
at pericentric
heterochromatin
go and run
MORE TOLEARN
TAKE HOME MESSAGE letrsquos consider
the heterochromatin axis as an interesting target in
tumorigenesis - immune response and
possible role in mediated HIV latency
bull The examples of Suv39h1 and HP1a today gt more
bull An illustration with the TH2- TH1 reprogrammingndash Notion of stemness
bull Further characterization needed both for in vivo models and target
--------------------------------------------------------
bull gt how is Suv39h1 contributing ndash Novel functions for Suv39 ndash connection with Sumoylation Maison et al 2016
bull gt Potential importance of CAF-1
More to learn with this modelhellip
GENERAL CONCLUSIONS
bull The parallel development of novel epigenetic drugs and the discovery of multiple oncogenic dependencies epigenetic antagonisms and synthetic lethal relationships has open a new era in epigenetic targeting of solid tumors (CONTEXT)
bull Therapeutic targeting of epigenetic dysfunction may not be limited to the use of epigenetic drugs the role of epigenetic regulation in orchestrating other essential cellular processes such as DNA repair or immunity and connection with metabolism opens additional therapeutic opportunities (BROADER IMPACT)
bull Connections with suppression of tumor-induced inflammation suggests that approaches targeting the anti-tumor immunity or tumor micro-environment might deserve further exploration
FUTURE CHALLENGES
bull tissue- andor subunit-dependent and epigenetic antagonism or synthetic lethal relationships observed in one system may not be translatable to another gt evaluation in the tumor type of interest
bull epigenetic drugs as monotherapy for oligoclonal diseases where the epigenetic alteration serves as oncogenic driver (mostly pediatric hematologic or selected solid malignancies) in polyclonal tumors where the epigenetic alteration is a favoring permissive or secondary event combinations with conventional chemotherapies targeted therapies or immune therapies may be required
bull inhibiting the catalytic activity of some subunitsmdashsuch as EZH2mdashmight not be sufficient to reach a full anti-tumor activity important to discriminate the contribution of the non-catalytic activity of chromatin remodelers to oncogenic events
bull primary and acquired resistance will need to be studied (notably by re-biopsing and molecularly characterizing patients tumors at the occurrence of resistance) and combinatorial approaches should be considered to address this issue
bull It should be remembered that epigenetic modifications are usually reversible
The Chromatin Dynamics Group Institut Curie Paris
La vie de Chacircteau Retreat Nouan le Fuselier 2016
Institut Curie Hospital
Translational ResearchPaul Cottu
Roman Rouzier
Anne-Vincent Salomon
Sergio Roman-Roman
Elisabetta Marangoni
David Gentien
Cecile Reyes
Collaborators
Vera Pancaldi
Alfonso Valencia
Genevieve Almouzni
Team
Jean-Pierre Quivy
Dominique Ray-Gallet
Christele Maison
Guillaume Orsi
Ekaterina Boyartchuk
Delphine Bailly
Daniel Jeffrey
Yann Louault
Audrey Forest
Camille Clement
David Sitbon
Julia Torne
Katrina Podsypanina
NCI NIH
USA
Yves Pommier
Fabricio G Sousa
EP CHPEECABOO
ACKNOWLEDGMENTS
Sebastian Amigorena
Lucia Pace Rhys Allan
Elina Zueva Heidi Schreiber
Vanessa Masson
Florence Cammas team
Peter Adams
Stanford USALaura Attardi
Univ Munich
GermanyAxel Imhof
CNIO Spain
Fundacion la
scienca di vida
Chile
Alejandra
Loyola
Beatson Inst
Scotland
Sophie Postel-
Vinay amp JC Soria
Gustave Roussy
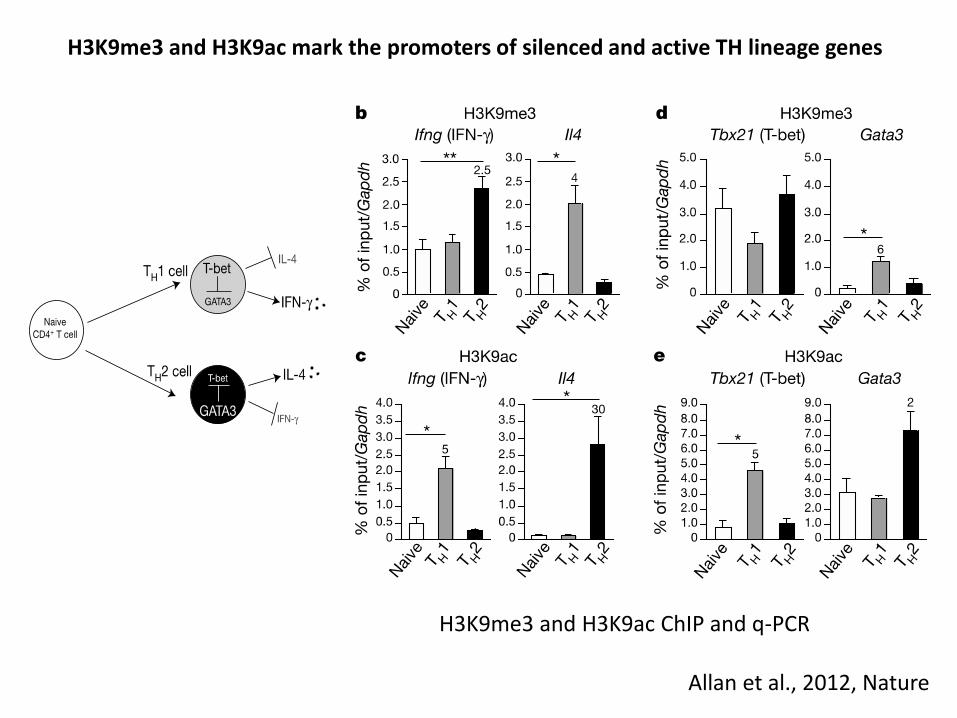
loss of SUV39H1 does not affect differentiation into TH1 or TH2 cells
Naiumlve CD4+ T from wt or Suv39h1 KO cells cultured under Th1 or Th2 conditions on day 7 intracellular staining for IL4 and Ifn gamma Th1 no IL4 but Ifng
Th2 IL4 but no IFng
Allan et al 2012 Nature
SUV39H1 deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Primary culture Naive CD41 T cells from wild- type or Suv39h1-knockout (Suv39h1 KO) mice differentiated for 7 days under TH2-promoting conditions Secondary culture recultured for 2 days in TH1-promoting conditions
TH2 to TH1 plasticity is shown by the increase in the percentage of cells that wereIFN-g positive in Suv39h1 KO when compared to wild-type
Allan et al 2012 Nature
Flow cytometry
HP1a deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Similar results but with HP1a KO
Allan et al 2012 Nature
SUV39H1 deficiency results in impaired ratio of H3K9 modifications in the IFNg promoter in TH2 cells
Primary culture TH2 cells analysed by ChIP using antibodies specific for histone modifications Quantitative PCR with primers specific for the promoters of the indicated genes(in TH2 Ifn is repressed in TH1 Ifn is produced)
plasticity plasticity
Model for a heterochromatin axis (SUV39H1 dependent) to control TH2 gt TH1 plasticity
Disruption of the heterochromatin axis increases plasticity
Zueva Quivy Almouzni and Amigorena 2013
Suv39h1-sustained heterochromatin organization plays a critical role in the expansion and programming of short-lived effector CD8+ T cells (ongoing)
NEXT
Stay tuned The next episodeWith Seb
RNA
Domains
Interacting
partnersMethylated K9 of H3
H3 histone fold
RNADNAChromatin
HP1a HP1 b
p150 CAF-1 KAP1
Suv39
Dnmt1 Dnmt3a Orc1amp 3Lachner et al Nature 2001
Bannister et al Nature 2001
Nielsen et al Mol Cell 2001
Maison et al Nat Genet 2002
Muchardt et al EMBO Rep 2002 Brasher et al EMBO J 2000
Cowieson et al Curr Biol 2000
Murzina et al Mol Cell 1999
Aagaard et al EMBO J 1999
Prasanth et al PNAS 2010
Nosawa et al NCB 2010
Ronoblet et al BBRC 2011
Mouse HP1a and partners
HP1a stabilityaccumulation
at pericentric
heterochromatin
go and run
MORE TOLEARN
TAKE HOME MESSAGE letrsquos consider
the heterochromatin axis as an interesting target in
tumorigenesis - immune response and
possible role in mediated HIV latency
bull The examples of Suv39h1 and HP1a today gt more
bull An illustration with the TH2- TH1 reprogrammingndash Notion of stemness
bull Further characterization needed both for in vivo models and target
--------------------------------------------------------
bull gt how is Suv39h1 contributing ndash Novel functions for Suv39 ndash connection with Sumoylation Maison et al 2016
bull gt Potential importance of CAF-1
More to learn with this modelhellip
GENERAL CONCLUSIONS
bull The parallel development of novel epigenetic drugs and the discovery of multiple oncogenic dependencies epigenetic antagonisms and synthetic lethal relationships has open a new era in epigenetic targeting of solid tumors (CONTEXT)
bull Therapeutic targeting of epigenetic dysfunction may not be limited to the use of epigenetic drugs the role of epigenetic regulation in orchestrating other essential cellular processes such as DNA repair or immunity and connection with metabolism opens additional therapeutic opportunities (BROADER IMPACT)
bull Connections with suppression of tumor-induced inflammation suggests that approaches targeting the anti-tumor immunity or tumor micro-environment might deserve further exploration
FUTURE CHALLENGES
bull tissue- andor subunit-dependent and epigenetic antagonism or synthetic lethal relationships observed in one system may not be translatable to another gt evaluation in the tumor type of interest
bull epigenetic drugs as monotherapy for oligoclonal diseases where the epigenetic alteration serves as oncogenic driver (mostly pediatric hematologic or selected solid malignancies) in polyclonal tumors where the epigenetic alteration is a favoring permissive or secondary event combinations with conventional chemotherapies targeted therapies or immune therapies may be required
bull inhibiting the catalytic activity of some subunitsmdashsuch as EZH2mdashmight not be sufficient to reach a full anti-tumor activity important to discriminate the contribution of the non-catalytic activity of chromatin remodelers to oncogenic events
bull primary and acquired resistance will need to be studied (notably by re-biopsing and molecularly characterizing patients tumors at the occurrence of resistance) and combinatorial approaches should be considered to address this issue
bull It should be remembered that epigenetic modifications are usually reversible
The Chromatin Dynamics Group Institut Curie Paris
La vie de Chacircteau Retreat Nouan le Fuselier 2016
Institut Curie Hospital
Translational ResearchPaul Cottu
Roman Rouzier
Anne-Vincent Salomon
Sergio Roman-Roman
Elisabetta Marangoni
David Gentien
Cecile Reyes
Collaborators
Vera Pancaldi
Alfonso Valencia
Genevieve Almouzni
Team
Jean-Pierre Quivy
Dominique Ray-Gallet
Christele Maison
Guillaume Orsi
Ekaterina Boyartchuk
Delphine Bailly
Daniel Jeffrey
Yann Louault
Audrey Forest
Camille Clement
David Sitbon
Julia Torne
Katrina Podsypanina
NCI NIH
USA
Yves Pommier
Fabricio G Sousa
EP CHPEECABOO
ACKNOWLEDGMENTS
Sebastian Amigorena
Lucia Pace Rhys Allan
Elina Zueva Heidi Schreiber
Vanessa Masson
Florence Cammas team
Peter Adams
Stanford USALaura Attardi
Univ Munich
GermanyAxel Imhof
CNIO Spain
Fundacion la
scienca di vida
Chile
Alejandra
Loyola
Beatson Inst
Scotland
Sophie Postel-
Vinay amp JC Soria
Gustave Roussy

SUV39H1 deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Primary culture Naive CD41 T cells from wild- type or Suv39h1-knockout (Suv39h1 KO) mice differentiated for 7 days under TH2-promoting conditions Secondary culture recultured for 2 days in TH1-promoting conditions
TH2 to TH1 plasticity is shown by the increase in the percentage of cells that wereIFN-g positive in Suv39h1 KO when compared to wild-type
Allan et al 2012 Nature
Flow cytometry
HP1a deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Similar results but with HP1a KO
Allan et al 2012 Nature
SUV39H1 deficiency results in impaired ratio of H3K9 modifications in the IFNg promoter in TH2 cells
Primary culture TH2 cells analysed by ChIP using antibodies specific for histone modifications Quantitative PCR with primers specific for the promoters of the indicated genes(in TH2 Ifn is repressed in TH1 Ifn is produced)
plasticity plasticity
Model for a heterochromatin axis (SUV39H1 dependent) to control TH2 gt TH1 plasticity
Disruption of the heterochromatin axis increases plasticity
Zueva Quivy Almouzni and Amigorena 2013
Suv39h1-sustained heterochromatin organization plays a critical role in the expansion and programming of short-lived effector CD8+ T cells (ongoing)
NEXT
Stay tuned The next episodeWith Seb
RNA
Domains
Interacting
partnersMethylated K9 of H3
H3 histone fold
RNADNAChromatin
HP1a HP1 b
p150 CAF-1 KAP1
Suv39
Dnmt1 Dnmt3a Orc1amp 3Lachner et al Nature 2001
Bannister et al Nature 2001
Nielsen et al Mol Cell 2001
Maison et al Nat Genet 2002
Muchardt et al EMBO Rep 2002 Brasher et al EMBO J 2000
Cowieson et al Curr Biol 2000
Murzina et al Mol Cell 1999
Aagaard et al EMBO J 1999
Prasanth et al PNAS 2010
Nosawa et al NCB 2010
Ronoblet et al BBRC 2011
Mouse HP1a and partners
HP1a stabilityaccumulation
at pericentric
heterochromatin
go and run
MORE TOLEARN
TAKE HOME MESSAGE letrsquos consider
the heterochromatin axis as an interesting target in
tumorigenesis - immune response and
possible role in mediated HIV latency
bull The examples of Suv39h1 and HP1a today gt more
bull An illustration with the TH2- TH1 reprogrammingndash Notion of stemness
bull Further characterization needed both for in vivo models and target
--------------------------------------------------------
bull gt how is Suv39h1 contributing ndash Novel functions for Suv39 ndash connection with Sumoylation Maison et al 2016
bull gt Potential importance of CAF-1
More to learn with this modelhellip
GENERAL CONCLUSIONS
bull The parallel development of novel epigenetic drugs and the discovery of multiple oncogenic dependencies epigenetic antagonisms and synthetic lethal relationships has open a new era in epigenetic targeting of solid tumors (CONTEXT)
bull Therapeutic targeting of epigenetic dysfunction may not be limited to the use of epigenetic drugs the role of epigenetic regulation in orchestrating other essential cellular processes such as DNA repair or immunity and connection with metabolism opens additional therapeutic opportunities (BROADER IMPACT)
bull Connections with suppression of tumor-induced inflammation suggests that approaches targeting the anti-tumor immunity or tumor micro-environment might deserve further exploration
FUTURE CHALLENGES
bull tissue- andor subunit-dependent and epigenetic antagonism or synthetic lethal relationships observed in one system may not be translatable to another gt evaluation in the tumor type of interest
bull epigenetic drugs as monotherapy for oligoclonal diseases where the epigenetic alteration serves as oncogenic driver (mostly pediatric hematologic or selected solid malignancies) in polyclonal tumors where the epigenetic alteration is a favoring permissive or secondary event combinations with conventional chemotherapies targeted therapies or immune therapies may be required
bull inhibiting the catalytic activity of some subunitsmdashsuch as EZH2mdashmight not be sufficient to reach a full anti-tumor activity important to discriminate the contribution of the non-catalytic activity of chromatin remodelers to oncogenic events
bull primary and acquired resistance will need to be studied (notably by re-biopsing and molecularly characterizing patients tumors at the occurrence of resistance) and combinatorial approaches should be considered to address this issue
bull It should be remembered that epigenetic modifications are usually reversible
The Chromatin Dynamics Group Institut Curie Paris
La vie de Chacircteau Retreat Nouan le Fuselier 2016
Institut Curie Hospital
Translational ResearchPaul Cottu
Roman Rouzier
Anne-Vincent Salomon
Sergio Roman-Roman
Elisabetta Marangoni
David Gentien
Cecile Reyes
Collaborators
Vera Pancaldi
Alfonso Valencia
Genevieve Almouzni
Team
Jean-Pierre Quivy
Dominique Ray-Gallet
Christele Maison
Guillaume Orsi
Ekaterina Boyartchuk
Delphine Bailly
Daniel Jeffrey
Yann Louault
Audrey Forest
Camille Clement
David Sitbon
Julia Torne
Katrina Podsypanina
NCI NIH
USA
Yves Pommier
Fabricio G Sousa
EP CHPEECABOO
ACKNOWLEDGMENTS
Sebastian Amigorena
Lucia Pace Rhys Allan
Elina Zueva Heidi Schreiber
Vanessa Masson
Florence Cammas team
Peter Adams
Stanford USALaura Attardi
Univ Munich
GermanyAxel Imhof
CNIO Spain
Fundacion la
scienca di vida
Chile
Alejandra
Loyola
Beatson Inst
Scotland
Sophie Postel-
Vinay amp JC Soria
Gustave Roussy

HP1a deficiency results in increased plasticity of TH2 cellsunder TH1-promoting conditions
Similar results but with HP1a KO
Allan et al 2012 Nature
SUV39H1 deficiency results in impaired ratio of H3K9 modifications in the IFNg promoter in TH2 cells
Primary culture TH2 cells analysed by ChIP using antibodies specific for histone modifications Quantitative PCR with primers specific for the promoters of the indicated genes(in TH2 Ifn is repressed in TH1 Ifn is produced)
plasticity plasticity
Model for a heterochromatin axis (SUV39H1 dependent) to control TH2 gt TH1 plasticity
Disruption of the heterochromatin axis increases plasticity
Zueva Quivy Almouzni and Amigorena 2013
Suv39h1-sustained heterochromatin organization plays a critical role in the expansion and programming of short-lived effector CD8+ T cells (ongoing)
NEXT
Stay tuned The next episodeWith Seb
RNA
Domains
Interacting
partnersMethylated K9 of H3
H3 histone fold
RNADNAChromatin
HP1a HP1 b
p150 CAF-1 KAP1
Suv39
Dnmt1 Dnmt3a Orc1amp 3Lachner et al Nature 2001
Bannister et al Nature 2001
Nielsen et al Mol Cell 2001
Maison et al Nat Genet 2002
Muchardt et al EMBO Rep 2002 Brasher et al EMBO J 2000
Cowieson et al Curr Biol 2000
Murzina et al Mol Cell 1999
Aagaard et al EMBO J 1999
Prasanth et al PNAS 2010
Nosawa et al NCB 2010
Ronoblet et al BBRC 2011
Mouse HP1a and partners
HP1a stabilityaccumulation
at pericentric
heterochromatin
go and run
MORE TOLEARN
TAKE HOME MESSAGE letrsquos consider
the heterochromatin axis as an interesting target in
tumorigenesis - immune response and
possible role in mediated HIV latency
bull The examples of Suv39h1 and HP1a today gt more
bull An illustration with the TH2- TH1 reprogrammingndash Notion of stemness
bull Further characterization needed both for in vivo models and target
--------------------------------------------------------
bull gt how is Suv39h1 contributing ndash Novel functions for Suv39 ndash connection with Sumoylation Maison et al 2016
bull gt Potential importance of CAF-1
More to learn with this modelhellip
GENERAL CONCLUSIONS
bull The parallel development of novel epigenetic drugs and the discovery of multiple oncogenic dependencies epigenetic antagonisms and synthetic lethal relationships has open a new era in epigenetic targeting of solid tumors (CONTEXT)
bull Therapeutic targeting of epigenetic dysfunction may not be limited to the use of epigenetic drugs the role of epigenetic regulation in orchestrating other essential cellular processes such as DNA repair or immunity and connection with metabolism opens additional therapeutic opportunities (BROADER IMPACT)
bull Connections with suppression of tumor-induced inflammation suggests that approaches targeting the anti-tumor immunity or tumor micro-environment might deserve further exploration
FUTURE CHALLENGES
bull tissue- andor subunit-dependent and epigenetic antagonism or synthetic lethal relationships observed in one system may not be translatable to another gt evaluation in the tumor type of interest
bull epigenetic drugs as monotherapy for oligoclonal diseases where the epigenetic alteration serves as oncogenic driver (mostly pediatric hematologic or selected solid malignancies) in polyclonal tumors where the epigenetic alteration is a favoring permissive or secondary event combinations with conventional chemotherapies targeted therapies or immune therapies may be required
bull inhibiting the catalytic activity of some subunitsmdashsuch as EZH2mdashmight not be sufficient to reach a full anti-tumor activity important to discriminate the contribution of the non-catalytic activity of chromatin remodelers to oncogenic events
bull primary and acquired resistance will need to be studied (notably by re-biopsing and molecularly characterizing patients tumors at the occurrence of resistance) and combinatorial approaches should be considered to address this issue
bull It should be remembered that epigenetic modifications are usually reversible
The Chromatin Dynamics Group Institut Curie Paris
La vie de Chacircteau Retreat Nouan le Fuselier 2016
Institut Curie Hospital
Translational ResearchPaul Cottu
Roman Rouzier
Anne-Vincent Salomon
Sergio Roman-Roman
Elisabetta Marangoni
David Gentien
Cecile Reyes
Collaborators
Vera Pancaldi
Alfonso Valencia
Genevieve Almouzni
Team
Jean-Pierre Quivy
Dominique Ray-Gallet
Christele Maison
Guillaume Orsi
Ekaterina Boyartchuk
Delphine Bailly
Daniel Jeffrey
Yann Louault
Audrey Forest
Camille Clement
David Sitbon
Julia Torne
Katrina Podsypanina
NCI NIH
USA
Yves Pommier
Fabricio G Sousa
EP CHPEECABOO
ACKNOWLEDGMENTS
Sebastian Amigorena
Lucia Pace Rhys Allan
Elina Zueva Heidi Schreiber
Vanessa Masson
Florence Cammas team
Peter Adams
Stanford USALaura Attardi
Univ Munich
GermanyAxel Imhof
CNIO Spain
Fundacion la
scienca di vida
Chile
Alejandra
Loyola
Beatson Inst
Scotland
Sophie Postel-
Vinay amp JC Soria
Gustave Roussy

SUV39H1 deficiency results in impaired ratio of H3K9 modifications in the IFNg promoter in TH2 cells
Primary culture TH2 cells analysed by ChIP using antibodies specific for histone modifications Quantitative PCR with primers specific for the promoters of the indicated genes(in TH2 Ifn is repressed in TH1 Ifn is produced)
plasticity plasticity
Model for a heterochromatin axis (SUV39H1 dependent) to control TH2 gt TH1 plasticity
Disruption of the heterochromatin axis increases plasticity
Zueva Quivy Almouzni and Amigorena 2013
Suv39h1-sustained heterochromatin organization plays a critical role in the expansion and programming of short-lived effector CD8+ T cells (ongoing)
NEXT
Stay tuned The next episodeWith Seb
RNA
Domains
Interacting
partnersMethylated K9 of H3
H3 histone fold
RNADNAChromatin
HP1a HP1 b
p150 CAF-1 KAP1
Suv39
Dnmt1 Dnmt3a Orc1amp 3Lachner et al Nature 2001
Bannister et al Nature 2001
Nielsen et al Mol Cell 2001
Maison et al Nat Genet 2002
Muchardt et al EMBO Rep 2002 Brasher et al EMBO J 2000
Cowieson et al Curr Biol 2000
Murzina et al Mol Cell 1999
Aagaard et al EMBO J 1999
Prasanth et al PNAS 2010
Nosawa et al NCB 2010
Ronoblet et al BBRC 2011
Mouse HP1a and partners
HP1a stabilityaccumulation
at pericentric
heterochromatin
go and run
MORE TOLEARN
TAKE HOME MESSAGE letrsquos consider
the heterochromatin axis as an interesting target in
tumorigenesis - immune response and
possible role in mediated HIV latency
bull The examples of Suv39h1 and HP1a today gt more
bull An illustration with the TH2- TH1 reprogrammingndash Notion of stemness
bull Further characterization needed both for in vivo models and target
--------------------------------------------------------
bull gt how is Suv39h1 contributing ndash Novel functions for Suv39 ndash connection with Sumoylation Maison et al 2016
bull gt Potential importance of CAF-1
More to learn with this modelhellip
GENERAL CONCLUSIONS
bull The parallel development of novel epigenetic drugs and the discovery of multiple oncogenic dependencies epigenetic antagonisms and synthetic lethal relationships has open a new era in epigenetic targeting of solid tumors (CONTEXT)
bull Therapeutic targeting of epigenetic dysfunction may not be limited to the use of epigenetic drugs the role of epigenetic regulation in orchestrating other essential cellular processes such as DNA repair or immunity and connection with metabolism opens additional therapeutic opportunities (BROADER IMPACT)
bull Connections with suppression of tumor-induced inflammation suggests that approaches targeting the anti-tumor immunity or tumor micro-environment might deserve further exploration
FUTURE CHALLENGES
bull tissue- andor subunit-dependent and epigenetic antagonism or synthetic lethal relationships observed in one system may not be translatable to another gt evaluation in the tumor type of interest
bull epigenetic drugs as monotherapy for oligoclonal diseases where the epigenetic alteration serves as oncogenic driver (mostly pediatric hematologic or selected solid malignancies) in polyclonal tumors where the epigenetic alteration is a favoring permissive or secondary event combinations with conventional chemotherapies targeted therapies or immune therapies may be required
bull inhibiting the catalytic activity of some subunitsmdashsuch as EZH2mdashmight not be sufficient to reach a full anti-tumor activity important to discriminate the contribution of the non-catalytic activity of chromatin remodelers to oncogenic events
bull primary and acquired resistance will need to be studied (notably by re-biopsing and molecularly characterizing patients tumors at the occurrence of resistance) and combinatorial approaches should be considered to address this issue
bull It should be remembered that epigenetic modifications are usually reversible
The Chromatin Dynamics Group Institut Curie Paris
La vie de Chacircteau Retreat Nouan le Fuselier 2016
Institut Curie Hospital
Translational ResearchPaul Cottu
Roman Rouzier
Anne-Vincent Salomon
Sergio Roman-Roman
Elisabetta Marangoni
David Gentien
Cecile Reyes
Collaborators
Vera Pancaldi
Alfonso Valencia
Genevieve Almouzni
Team
Jean-Pierre Quivy
Dominique Ray-Gallet
Christele Maison
Guillaume Orsi
Ekaterina Boyartchuk
Delphine Bailly
Daniel Jeffrey
Yann Louault
Audrey Forest
Camille Clement
David Sitbon
Julia Torne
Katrina Podsypanina
NCI NIH
USA
Yves Pommier
Fabricio G Sousa
EP CHPEECABOO
ACKNOWLEDGMENTS
Sebastian Amigorena
Lucia Pace Rhys Allan
Elina Zueva Heidi Schreiber
Vanessa Masson
Florence Cammas team
Peter Adams
Stanford USALaura Attardi
Univ Munich
GermanyAxel Imhof
CNIO Spain
Fundacion la
scienca di vida
Chile
Alejandra
Loyola
Beatson Inst
Scotland
Sophie Postel-
Vinay amp JC Soria
Gustave Roussy

plasticity plasticity
Model for a heterochromatin axis (SUV39H1 dependent) to control TH2 gt TH1 plasticity
Disruption of the heterochromatin axis increases plasticity
Zueva Quivy Almouzni and Amigorena 2013
Suv39h1-sustained heterochromatin organization plays a critical role in the expansion and programming of short-lived effector CD8+ T cells (ongoing)
NEXT
Stay tuned The next episodeWith Seb
RNA
Domains
Interacting
partnersMethylated K9 of H3
H3 histone fold
RNADNAChromatin
HP1a HP1 b
p150 CAF-1 KAP1
Suv39
Dnmt1 Dnmt3a Orc1amp 3Lachner et al Nature 2001
Bannister et al Nature 2001
Nielsen et al Mol Cell 2001
Maison et al Nat Genet 2002
Muchardt et al EMBO Rep 2002 Brasher et al EMBO J 2000
Cowieson et al Curr Biol 2000
Murzina et al Mol Cell 1999
Aagaard et al EMBO J 1999
Prasanth et al PNAS 2010
Nosawa et al NCB 2010
Ronoblet et al BBRC 2011
Mouse HP1a and partners
HP1a stabilityaccumulation
at pericentric
heterochromatin
go and run
MORE TOLEARN
TAKE HOME MESSAGE letrsquos consider
the heterochromatin axis as an interesting target in
tumorigenesis - immune response and
possible role in mediated HIV latency
bull The examples of Suv39h1 and HP1a today gt more
bull An illustration with the TH2- TH1 reprogrammingndash Notion of stemness
bull Further characterization needed both for in vivo models and target
--------------------------------------------------------
bull gt how is Suv39h1 contributing ndash Novel functions for Suv39 ndash connection with Sumoylation Maison et al 2016
bull gt Potential importance of CAF-1
More to learn with this modelhellip
GENERAL CONCLUSIONS
bull The parallel development of novel epigenetic drugs and the discovery of multiple oncogenic dependencies epigenetic antagonisms and synthetic lethal relationships has open a new era in epigenetic targeting of solid tumors (CONTEXT)
bull Therapeutic targeting of epigenetic dysfunction may not be limited to the use of epigenetic drugs the role of epigenetic regulation in orchestrating other essential cellular processes such as DNA repair or immunity and connection with metabolism opens additional therapeutic opportunities (BROADER IMPACT)
bull Connections with suppression of tumor-induced inflammation suggests that approaches targeting the anti-tumor immunity or tumor micro-environment might deserve further exploration
FUTURE CHALLENGES
bull tissue- andor subunit-dependent and epigenetic antagonism or synthetic lethal relationships observed in one system may not be translatable to another gt evaluation in the tumor type of interest
bull epigenetic drugs as monotherapy for oligoclonal diseases where the epigenetic alteration serves as oncogenic driver (mostly pediatric hematologic or selected solid malignancies) in polyclonal tumors where the epigenetic alteration is a favoring permissive or secondary event combinations with conventional chemotherapies targeted therapies or immune therapies may be required
bull inhibiting the catalytic activity of some subunitsmdashsuch as EZH2mdashmight not be sufficient to reach a full anti-tumor activity important to discriminate the contribution of the non-catalytic activity of chromatin remodelers to oncogenic events
bull primary and acquired resistance will need to be studied (notably by re-biopsing and molecularly characterizing patients tumors at the occurrence of resistance) and combinatorial approaches should be considered to address this issue
bull It should be remembered that epigenetic modifications are usually reversible
The Chromatin Dynamics Group Institut Curie Paris
La vie de Chacircteau Retreat Nouan le Fuselier 2016
Institut Curie Hospital
Translational ResearchPaul Cottu
Roman Rouzier
Anne-Vincent Salomon
Sergio Roman-Roman
Elisabetta Marangoni
David Gentien
Cecile Reyes
Collaborators
Vera Pancaldi
Alfonso Valencia
Genevieve Almouzni
Team
Jean-Pierre Quivy
Dominique Ray-Gallet
Christele Maison
Guillaume Orsi
Ekaterina Boyartchuk
Delphine Bailly
Daniel Jeffrey
Yann Louault
Audrey Forest
Camille Clement
David Sitbon
Julia Torne
Katrina Podsypanina
NCI NIH
USA
Yves Pommier
Fabricio G Sousa
EP CHPEECABOO
ACKNOWLEDGMENTS
Sebastian Amigorena
Lucia Pace Rhys Allan
Elina Zueva Heidi Schreiber
Vanessa Masson
Florence Cammas team
Peter Adams
Stanford USALaura Attardi
Univ Munich
GermanyAxel Imhof
CNIO Spain
Fundacion la
scienca di vida
Chile
Alejandra
Loyola
Beatson Inst
Scotland
Sophie Postel-
Vinay amp JC Soria
Gustave Roussy

Zueva Quivy Almouzni and Amigorena 2013
Suv39h1-sustained heterochromatin organization plays a critical role in the expansion and programming of short-lived effector CD8+ T cells (ongoing)
NEXT
Stay tuned The next episodeWith Seb
RNA
Domains
Interacting
partnersMethylated K9 of H3
H3 histone fold
RNADNAChromatin
HP1a HP1 b
p150 CAF-1 KAP1
Suv39
Dnmt1 Dnmt3a Orc1amp 3Lachner et al Nature 2001
Bannister et al Nature 2001
Nielsen et al Mol Cell 2001
Maison et al Nat Genet 2002
Muchardt et al EMBO Rep 2002 Brasher et al EMBO J 2000
Cowieson et al Curr Biol 2000
Murzina et al Mol Cell 1999
Aagaard et al EMBO J 1999
Prasanth et al PNAS 2010
Nosawa et al NCB 2010
Ronoblet et al BBRC 2011
Mouse HP1a and partners
HP1a stabilityaccumulation
at pericentric
heterochromatin
go and run
MORE TOLEARN
TAKE HOME MESSAGE letrsquos consider
the heterochromatin axis as an interesting target in
tumorigenesis - immune response and
possible role in mediated HIV latency
bull The examples of Suv39h1 and HP1a today gt more
bull An illustration with the TH2- TH1 reprogrammingndash Notion of stemness
bull Further characterization needed both for in vivo models and target
--------------------------------------------------------
bull gt how is Suv39h1 contributing ndash Novel functions for Suv39 ndash connection with Sumoylation Maison et al 2016
bull gt Potential importance of CAF-1
More to learn with this modelhellip
GENERAL CONCLUSIONS
bull The parallel development of novel epigenetic drugs and the discovery of multiple oncogenic dependencies epigenetic antagonisms and synthetic lethal relationships has open a new era in epigenetic targeting of solid tumors (CONTEXT)
bull Therapeutic targeting of epigenetic dysfunction may not be limited to the use of epigenetic drugs the role of epigenetic regulation in orchestrating other essential cellular processes such as DNA repair or immunity and connection with metabolism opens additional therapeutic opportunities (BROADER IMPACT)
bull Connections with suppression of tumor-induced inflammation suggests that approaches targeting the anti-tumor immunity or tumor micro-environment might deserve further exploration
FUTURE CHALLENGES
bull tissue- andor subunit-dependent and epigenetic antagonism or synthetic lethal relationships observed in one system may not be translatable to another gt evaluation in the tumor type of interest
bull epigenetic drugs as monotherapy for oligoclonal diseases where the epigenetic alteration serves as oncogenic driver (mostly pediatric hematologic or selected solid malignancies) in polyclonal tumors where the epigenetic alteration is a favoring permissive or secondary event combinations with conventional chemotherapies targeted therapies or immune therapies may be required
bull inhibiting the catalytic activity of some subunitsmdashsuch as EZH2mdashmight not be sufficient to reach a full anti-tumor activity important to discriminate the contribution of the non-catalytic activity of chromatin remodelers to oncogenic events
bull primary and acquired resistance will need to be studied (notably by re-biopsing and molecularly characterizing patients tumors at the occurrence of resistance) and combinatorial approaches should be considered to address this issue
bull It should be remembered that epigenetic modifications are usually reversible
The Chromatin Dynamics Group Institut Curie Paris
La vie de Chacircteau Retreat Nouan le Fuselier 2016
Institut Curie Hospital
Translational ResearchPaul Cottu
Roman Rouzier
Anne-Vincent Salomon
Sergio Roman-Roman
Elisabetta Marangoni
David Gentien
Cecile Reyes
Collaborators
Vera Pancaldi
Alfonso Valencia
Genevieve Almouzni
Team
Jean-Pierre Quivy
Dominique Ray-Gallet
Christele Maison
Guillaume Orsi
Ekaterina Boyartchuk
Delphine Bailly
Daniel Jeffrey
Yann Louault
Audrey Forest
Camille Clement
David Sitbon
Julia Torne
Katrina Podsypanina
NCI NIH
USA
Yves Pommier
Fabricio G Sousa
EP CHPEECABOO
ACKNOWLEDGMENTS
Sebastian Amigorena
Lucia Pace Rhys Allan
Elina Zueva Heidi Schreiber
Vanessa Masson
Florence Cammas team
Peter Adams
Stanford USALaura Attardi
Univ Munich
GermanyAxel Imhof
CNIO Spain
Fundacion la
scienca di vida
Chile
Alejandra
Loyola
Beatson Inst
Scotland
Sophie Postel-
Vinay amp JC Soria
Gustave Roussy

RNA
Domains
Interacting
partnersMethylated K9 of H3
H3 histone fold
RNADNAChromatin
HP1a HP1 b
p150 CAF-1 KAP1
Suv39
Dnmt1 Dnmt3a Orc1amp 3Lachner et al Nature 2001
Bannister et al Nature 2001
Nielsen et al Mol Cell 2001
Maison et al Nat Genet 2002
Muchardt et al EMBO Rep 2002 Brasher et al EMBO J 2000
Cowieson et al Curr Biol 2000
Murzina et al Mol Cell 1999
Aagaard et al EMBO J 1999
Prasanth et al PNAS 2010
Nosawa et al NCB 2010
Ronoblet et al BBRC 2011
Mouse HP1a and partners
HP1a stabilityaccumulation
at pericentric
heterochromatin
go and run
MORE TOLEARN
TAKE HOME MESSAGE letrsquos consider
the heterochromatin axis as an interesting target in
tumorigenesis - immune response and
possible role in mediated HIV latency
bull The examples of Suv39h1 and HP1a today gt more
bull An illustration with the TH2- TH1 reprogrammingndash Notion of stemness
bull Further characterization needed both for in vivo models and target
--------------------------------------------------------
bull gt how is Suv39h1 contributing ndash Novel functions for Suv39 ndash connection with Sumoylation Maison et al 2016
bull gt Potential importance of CAF-1
More to learn with this modelhellip
GENERAL CONCLUSIONS
bull The parallel development of novel epigenetic drugs and the discovery of multiple oncogenic dependencies epigenetic antagonisms and synthetic lethal relationships has open a new era in epigenetic targeting of solid tumors (CONTEXT)
bull Therapeutic targeting of epigenetic dysfunction may not be limited to the use of epigenetic drugs the role of epigenetic regulation in orchestrating other essential cellular processes such as DNA repair or immunity and connection with metabolism opens additional therapeutic opportunities (BROADER IMPACT)
bull Connections with suppression of tumor-induced inflammation suggests that approaches targeting the anti-tumor immunity or tumor micro-environment might deserve further exploration
FUTURE CHALLENGES
bull tissue- andor subunit-dependent and epigenetic antagonism or synthetic lethal relationships observed in one system may not be translatable to another gt evaluation in the tumor type of interest
bull epigenetic drugs as monotherapy for oligoclonal diseases where the epigenetic alteration serves as oncogenic driver (mostly pediatric hematologic or selected solid malignancies) in polyclonal tumors where the epigenetic alteration is a favoring permissive or secondary event combinations with conventional chemotherapies targeted therapies or immune therapies may be required
bull inhibiting the catalytic activity of some subunitsmdashsuch as EZH2mdashmight not be sufficient to reach a full anti-tumor activity important to discriminate the contribution of the non-catalytic activity of chromatin remodelers to oncogenic events
bull primary and acquired resistance will need to be studied (notably by re-biopsing and molecularly characterizing patients tumors at the occurrence of resistance) and combinatorial approaches should be considered to address this issue
bull It should be remembered that epigenetic modifications are usually reversible
The Chromatin Dynamics Group Institut Curie Paris
La vie de Chacircteau Retreat Nouan le Fuselier 2016
Institut Curie Hospital
Translational ResearchPaul Cottu
Roman Rouzier
Anne-Vincent Salomon
Sergio Roman-Roman
Elisabetta Marangoni
David Gentien
Cecile Reyes
Collaborators
Vera Pancaldi
Alfonso Valencia
Genevieve Almouzni
Team
Jean-Pierre Quivy
Dominique Ray-Gallet
Christele Maison
Guillaume Orsi
Ekaterina Boyartchuk
Delphine Bailly
Daniel Jeffrey
Yann Louault
Audrey Forest
Camille Clement
David Sitbon
Julia Torne
Katrina Podsypanina
NCI NIH
USA
Yves Pommier
Fabricio G Sousa
EP CHPEECABOO
ACKNOWLEDGMENTS
Sebastian Amigorena
Lucia Pace Rhys Allan
Elina Zueva Heidi Schreiber
Vanessa Masson
Florence Cammas team
Peter Adams
Stanford USALaura Attardi
Univ Munich
GermanyAxel Imhof
CNIO Spain
Fundacion la
scienca di vida
Chile
Alejandra
Loyola
Beatson Inst
Scotland
Sophie Postel-
Vinay amp JC Soria
Gustave Roussy

TAKE HOME MESSAGE letrsquos consider
the heterochromatin axis as an interesting target in
tumorigenesis - immune response and
possible role in mediated HIV latency
bull The examples of Suv39h1 and HP1a today gt more
bull An illustration with the TH2- TH1 reprogrammingndash Notion of stemness
bull Further characterization needed both for in vivo models and target
--------------------------------------------------------
bull gt how is Suv39h1 contributing ndash Novel functions for Suv39 ndash connection with Sumoylation Maison et al 2016
bull gt Potential importance of CAF-1
More to learn with this modelhellip
GENERAL CONCLUSIONS
bull The parallel development of novel epigenetic drugs and the discovery of multiple oncogenic dependencies epigenetic antagonisms and synthetic lethal relationships has open a new era in epigenetic targeting of solid tumors (CONTEXT)
bull Therapeutic targeting of epigenetic dysfunction may not be limited to the use of epigenetic drugs the role of epigenetic regulation in orchestrating other essential cellular processes such as DNA repair or immunity and connection with metabolism opens additional therapeutic opportunities (BROADER IMPACT)
bull Connections with suppression of tumor-induced inflammation suggests that approaches targeting the anti-tumor immunity or tumor micro-environment might deserve further exploration
FUTURE CHALLENGES
bull tissue- andor subunit-dependent and epigenetic antagonism or synthetic lethal relationships observed in one system may not be translatable to another gt evaluation in the tumor type of interest
bull epigenetic drugs as monotherapy for oligoclonal diseases where the epigenetic alteration serves as oncogenic driver (mostly pediatric hematologic or selected solid malignancies) in polyclonal tumors where the epigenetic alteration is a favoring permissive or secondary event combinations with conventional chemotherapies targeted therapies or immune therapies may be required
bull inhibiting the catalytic activity of some subunitsmdashsuch as EZH2mdashmight not be sufficient to reach a full anti-tumor activity important to discriminate the contribution of the non-catalytic activity of chromatin remodelers to oncogenic events
bull primary and acquired resistance will need to be studied (notably by re-biopsing and molecularly characterizing patients tumors at the occurrence of resistance) and combinatorial approaches should be considered to address this issue
bull It should be remembered that epigenetic modifications are usually reversible
The Chromatin Dynamics Group Institut Curie Paris
La vie de Chacircteau Retreat Nouan le Fuselier 2016
Institut Curie Hospital
Translational ResearchPaul Cottu
Roman Rouzier
Anne-Vincent Salomon
Sergio Roman-Roman
Elisabetta Marangoni
David Gentien
Cecile Reyes
Collaborators
Vera Pancaldi
Alfonso Valencia
Genevieve Almouzni
Team
Jean-Pierre Quivy
Dominique Ray-Gallet
Christele Maison
Guillaume Orsi
Ekaterina Boyartchuk
Delphine Bailly
Daniel Jeffrey
Yann Louault
Audrey Forest
Camille Clement
David Sitbon
Julia Torne
Katrina Podsypanina
NCI NIH
USA
Yves Pommier
Fabricio G Sousa
EP CHPEECABOO
ACKNOWLEDGMENTS
Sebastian Amigorena
Lucia Pace Rhys Allan
Elina Zueva Heidi Schreiber
Vanessa Masson
Florence Cammas team
Peter Adams
Stanford USALaura Attardi
Univ Munich
GermanyAxel Imhof
CNIO Spain
Fundacion la
scienca di vida
Chile
Alejandra
Loyola
Beatson Inst
Scotland
Sophie Postel-
Vinay amp JC Soria
Gustave Roussy

GENERAL CONCLUSIONS
bull The parallel development of novel epigenetic drugs and the discovery of multiple oncogenic dependencies epigenetic antagonisms and synthetic lethal relationships has open a new era in epigenetic targeting of solid tumors (CONTEXT)
bull Therapeutic targeting of epigenetic dysfunction may not be limited to the use of epigenetic drugs the role of epigenetic regulation in orchestrating other essential cellular processes such as DNA repair or immunity and connection with metabolism opens additional therapeutic opportunities (BROADER IMPACT)
bull Connections with suppression of tumor-induced inflammation suggests that approaches targeting the anti-tumor immunity or tumor micro-environment might deserve further exploration
FUTURE CHALLENGES
bull tissue- andor subunit-dependent and epigenetic antagonism or synthetic lethal relationships observed in one system may not be translatable to another gt evaluation in the tumor type of interest
bull epigenetic drugs as monotherapy for oligoclonal diseases where the epigenetic alteration serves as oncogenic driver (mostly pediatric hematologic or selected solid malignancies) in polyclonal tumors where the epigenetic alteration is a favoring permissive or secondary event combinations with conventional chemotherapies targeted therapies or immune therapies may be required
bull inhibiting the catalytic activity of some subunitsmdashsuch as EZH2mdashmight not be sufficient to reach a full anti-tumor activity important to discriminate the contribution of the non-catalytic activity of chromatin remodelers to oncogenic events
bull primary and acquired resistance will need to be studied (notably by re-biopsing and molecularly characterizing patients tumors at the occurrence of resistance) and combinatorial approaches should be considered to address this issue
bull It should be remembered that epigenetic modifications are usually reversible
The Chromatin Dynamics Group Institut Curie Paris
La vie de Chacircteau Retreat Nouan le Fuselier 2016
Institut Curie Hospital
Translational ResearchPaul Cottu
Roman Rouzier
Anne-Vincent Salomon
Sergio Roman-Roman
Elisabetta Marangoni
David Gentien
Cecile Reyes
Collaborators
Vera Pancaldi
Alfonso Valencia
Genevieve Almouzni
Team
Jean-Pierre Quivy
Dominique Ray-Gallet
Christele Maison
Guillaume Orsi
Ekaterina Boyartchuk
Delphine Bailly
Daniel Jeffrey
Yann Louault
Audrey Forest
Camille Clement
David Sitbon
Julia Torne
Katrina Podsypanina
NCI NIH
USA
Yves Pommier
Fabricio G Sousa
EP CHPEECABOO
ACKNOWLEDGMENTS
Sebastian Amigorena
Lucia Pace Rhys Allan
Elina Zueva Heidi Schreiber
Vanessa Masson
Florence Cammas team
Peter Adams
Stanford USALaura Attardi
Univ Munich
GermanyAxel Imhof
CNIO Spain
Fundacion la
scienca di vida
Chile
Alejandra
Loyola
Beatson Inst
Scotland
Sophie Postel-
Vinay amp JC Soria
Gustave Roussy

FUTURE CHALLENGES
bull tissue- andor subunit-dependent and epigenetic antagonism or synthetic lethal relationships observed in one system may not be translatable to another gt evaluation in the tumor type of interest
bull epigenetic drugs as monotherapy for oligoclonal diseases where the epigenetic alteration serves as oncogenic driver (mostly pediatric hematologic or selected solid malignancies) in polyclonal tumors where the epigenetic alteration is a favoring permissive or secondary event combinations with conventional chemotherapies targeted therapies or immune therapies may be required
bull inhibiting the catalytic activity of some subunitsmdashsuch as EZH2mdashmight not be sufficient to reach a full anti-tumor activity important to discriminate the contribution of the non-catalytic activity of chromatin remodelers to oncogenic events
bull primary and acquired resistance will need to be studied (notably by re-biopsing and molecularly characterizing patients tumors at the occurrence of resistance) and combinatorial approaches should be considered to address this issue
bull It should be remembered that epigenetic modifications are usually reversible
The Chromatin Dynamics Group Institut Curie Paris
La vie de Chacircteau Retreat Nouan le Fuselier 2016
Institut Curie Hospital
Translational ResearchPaul Cottu
Roman Rouzier
Anne-Vincent Salomon
Sergio Roman-Roman
Elisabetta Marangoni
David Gentien
Cecile Reyes
Collaborators
Vera Pancaldi
Alfonso Valencia
Genevieve Almouzni
Team
Jean-Pierre Quivy
Dominique Ray-Gallet
Christele Maison
Guillaume Orsi
Ekaterina Boyartchuk
Delphine Bailly
Daniel Jeffrey
Yann Louault
Audrey Forest
Camille Clement
David Sitbon
Julia Torne
Katrina Podsypanina
NCI NIH
USA
Yves Pommier
Fabricio G Sousa
EP CHPEECABOO
ACKNOWLEDGMENTS
Sebastian Amigorena
Lucia Pace Rhys Allan
Elina Zueva Heidi Schreiber
Vanessa Masson
Florence Cammas team
Peter Adams
Stanford USALaura Attardi
Univ Munich
GermanyAxel Imhof
CNIO Spain
Fundacion la
scienca di vida
Chile
Alejandra
Loyola
Beatson Inst
Scotland
Sophie Postel-
Vinay amp JC Soria
Gustave Roussy

The Chromatin Dynamics Group Institut Curie Paris
La vie de Chacircteau Retreat Nouan le Fuselier 2016
Institut Curie Hospital
Translational ResearchPaul Cottu
Roman Rouzier
Anne-Vincent Salomon
Sergio Roman-Roman
Elisabetta Marangoni
David Gentien
Cecile Reyes
Collaborators
Vera Pancaldi
Alfonso Valencia
Genevieve Almouzni
Team
Jean-Pierre Quivy
Dominique Ray-Gallet
Christele Maison
Guillaume Orsi
Ekaterina Boyartchuk
Delphine Bailly
Daniel Jeffrey
Yann Louault
Audrey Forest
Camille Clement
David Sitbon
Julia Torne
Katrina Podsypanina
NCI NIH
USA
Yves Pommier
Fabricio G Sousa
EP CHPEECABOO
ACKNOWLEDGMENTS
Sebastian Amigorena
Lucia Pace Rhys Allan
Elina Zueva Heidi Schreiber
Vanessa Masson
Florence Cammas team
Peter Adams
Stanford USALaura Attardi
Univ Munich
GermanyAxel Imhof
CNIO Spain
Fundacion la
scienca di vida
Chile
Alejandra
Loyola
Beatson Inst
Scotland
Sophie Postel-
Vinay amp JC Soria
Gustave Roussy

Institut Curie Hospital
Translational ResearchPaul Cottu
Roman Rouzier
Anne-Vincent Salomon
Sergio Roman-Roman
Elisabetta Marangoni
David Gentien
Cecile Reyes
Collaborators
Vera Pancaldi
Alfonso Valencia
Genevieve Almouzni
Team
Jean-Pierre Quivy
Dominique Ray-Gallet
Christele Maison
Guillaume Orsi
Ekaterina Boyartchuk
Delphine Bailly
Daniel Jeffrey
Yann Louault
Audrey Forest
Camille Clement
David Sitbon
Julia Torne
Katrina Podsypanina
NCI NIH
USA
Yves Pommier
Fabricio G Sousa
EP CHPEECABOO
ACKNOWLEDGMENTS
Sebastian Amigorena
Lucia Pace Rhys Allan
Elina Zueva Heidi Schreiber
Vanessa Masson
Florence Cammas team
Peter Adams
Stanford USALaura Attardi
Univ Munich
GermanyAxel Imhof
CNIO Spain
Fundacion la
scienca di vida
Chile
Alejandra
Loyola
Beatson Inst
Scotland
Sophie Postel-
Vinay amp JC Soria
Gustave Roussy





























