Embed Size (px)
Citation preview

Sandalwood genetics and essential oil biochemistry
Chris Jones
School of Plant Biology, FNAS
Supervisors:
A/Prof Julie A Plummer (UWA)
A/Prof Emilio L Ghisalberti (UWA)
Dr Margaret Byrne (DEC)
Dr Elizabeth L Barbour (FPC)
A/Prof Jörg Bohlmann (UBC)

Outline of presentation
�Botany, distribution and diversity of sandalwood
�Economic significance and the move to plantations
�Key problems affecting the plantation sector
�Experiment 1: Genetic diversity analysis
�Experiment 2: Essential oil composition
�Experiment 3: Biosynthesis of sesquiterpenes in Santalum album
�Conclusions and future work
�Questions

Distribution of Santalum (Radomiljac et. al 1998)
S. album
S. macgregorii
S. spicatum,
S. murrayanum,
S. accuminatum S. lanceolatum
S. insulare
S. austrocaledonicum
S. freycinetianum,
S. ellipticumS. boninense
S. yasi

Economic importance of the sandalwoods
~$100,000 per ton!

Supply and demand…
1800’s 2000’s

The Kununurra collection

The trouble with sandalwood…
�Root parasitism means host selection is vital
�Short, medium and long term hosts (up to 50 yr!) means high maintenance silviculture
�Now achieving satisfactory growth and survival
�…until harvest time

What is causing this variation?
�Genetic variation = more adaptable to change
� Interbreeding populations contain individuals with unique genotypes
�This may include a predisposition to yield more heartwood, produce different compounds in the oil, or generally grow faster.
� If specific genes control heartwood production, how can we exploit this to improve productivity?

Three main hypotheses:
1. The genetic diversity of Australian Santalum album captured in Kununurra is reflective of its evolutionary history and natural distribution
2. That the essential oil yield and composition will reflect this genetic diversity, with regular patterns in similar genotypes
3. That these oils are synthesised in the heartwood by terpene synthase (TPS) enzymes, which are encoded by expressed TPS genes

Experiment 1: Genetic diversity of sandalwood

Genetic diversity analysis: Methods
�Restriction fragment length polymorphism analysis
�Total DNA is cut with restriction enzymes, run on gel electrophoresis and Southern blotted
�32
P-labelled homologous DNA (probe) hybridised to blots, washed and exposed to x-ray film
�Polymorphic DNA appears as bands (1 band = homozygous, 2 = heterozygous)
� 20 probes from a S. spicatum genomic library
�Also used Petunia chloroplast probes to assess phylogenetic history of the Austral Santalum spp.

CSI Crawley…

Genetic diversity analysis: Results
� Santalum album genome is highly homozygous!
� Of the 20 nuclear probes, only 2 were polymorphic
� Recent major restriction of gene flow to the population
0.095
0.391
0.137
0.681
Fit (inbreeding)
Butcher et. al 19980.1310.124Acacia mangium (PNG)
Butcher et. al 20020.4900.480Eucalyptus camaldulensis
Byrne et. al 20030.2100.201Santalum spicatum (WA)
This work0.0850.032Santalum album (Arboretum)
ReferenceHeHo
Byrne, M., MacDonald, B., Brand, J. E., 2003. Regional genetic differentiation in Western Australian sandalwood (Santalum spicatum) as revealed by nuclear RFLP analysis. Theoretical and Applied Genetics 107, 1208-1214.Butcher, P. A., Moran, G. F., Perkins, H. D., 1998. RFLP diversity in the nuclear genome of Acacia mangium. Heredity 81, 205-213.Butcher, P. A., Otero, A., McDonald, M. W., Moran, G. F., 2002. Nuclear RFLP variation in Eucalyptus camaldulensis Dehnh. from northern Australia. Heredity 88, 402-412.

30 length units
Genetic diversity analysis: Results
UPGMA of 275 trees
S. spicatum out-group

Genetic diversity analysis: Results
� Chloroplast RFLPs show major divergence of northern species from southern (S. spicatum)
� Confirms Australian origin of the genus (Harbaugh and Baldwin 2007)
� Recent speciation from major restriction of gene flow: long range dispersal of seed to islands, vicariance of PNG
� S. album from Timor is essentially identical to Indian material
� Dispersal of S. album to India was very recent, most likely by humans in the last 2000 years
� Continued reduction in diversity through over-exploitation, self pollination and vegetative regrowth
Harbaugh, D. T., Baldwin, B. G., 2007. Phylogeny and biogeography of the sandalwoods (Santalum, Santalaceae): Repeated dispersals throughout the Pacific. American Journal of Botany 64, 1028-1040.

Genetic diversity analysis: Results

Prehistoric dispersal of Santalum from Australia
Lowest sea level during the Pleistocene -120m
250,000 years before present
S. album
Polynesian species
S. macgregorii
India
Timor Is.
Repeated westerly
dispersal by humans
over ~2000 y
Hinduism in Bali?
Chinese trade?
© Field Museum 2000
China
Bali

Experiment 2: Essential oil composition
0.1 mm

10.0 12.5 15.0 17.5 20.0 22.5 25.0 27.5 30.0 32.5 35.0 37.5 40.0 min
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
1.1
1.2
1.3
uV(x1,000)
Chromatogram
1
2
5
3
6
8
74
Chromatogram of S. album oil extract
Essential oil analysis and variation

Essential oil analysis: Methods
� 100 trees aged between 8 and 17 years were cored (non-destructive sampling)
� 2 cores per tree; 30 cm and 1 m above ground
�Dried, ground, extracted in EtOH
�Samples were analysed using GC-FID and CG-MS
�Quantified over 65 compounds
�Chemotype profiles compiled and compared
�Yield and composition patterns explored
�Fit these to genotypes discovered earlier

0
10
20
30
40
50
60
0-10 10-20 20-30 30-40 40-50 More
mg/g oil
Frequency
Essential oil analysis: Results
�Most trees 8-10 yrs at time of core
�Oldest trees ~17 yrs
�Highest core oil yield = 6%

Essential oil analysis: Results
�No distinct chemotypes-identical chromatograms
�No obvious genotype-oil yield correlation (not all genetic!)
�Strong co-occurrence patterns between compounds implies shared intermediates
Jones CG, Ghisalberti EL, Plummer JA, Barbour EL (2006) Phytochemistry 67, 2463–2468

gDNA
(ribosome)
Transcription
Terminator
mRNA
Amino acids Enzyme
Promoter: controls transcription level
UTR-AAAAA 3’5’UTR---ATG
substrate
product
Intron: “junk” DNA Exon: actual gene
ATG
Experiment 3. Sandalwood oil biosynthesis

Sandalwood oil biosynthesis: Methods
�Terpene synthases (TPS) biosynthesise terpenes in all organisms. GPP is turned into monoterpenes, FPP into sesquiterpenes
�Degenerate (wildcard) primers amplified TPS fragment from gDNA.
�Enabled gene-specific primers for Rapid Amplification of cDNA Ends (RACE)
�Sequenced full length gene, clone into bacteria
�Heterologous expression of protein enables functional characterisation.
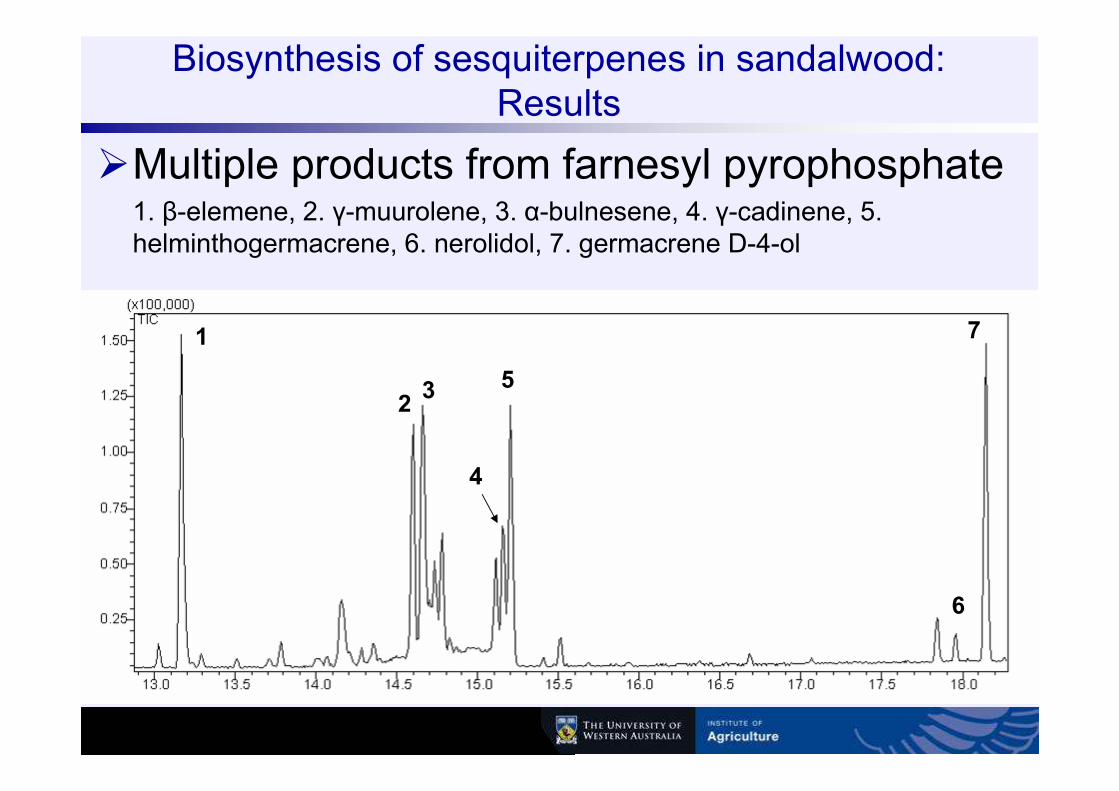
Biosynthesis of sesquiterpenes in sandalwood: Results
1 7
23
4
5
6
�Multiple products from farnesyl pyrophosphate1. β-elemene, 2. γ-muurolene, 3. α-bulnesene, 4. γ-cadinene, 5. helminthogermacrene, 6. nerolidol, 7. germacrene D-4-ol
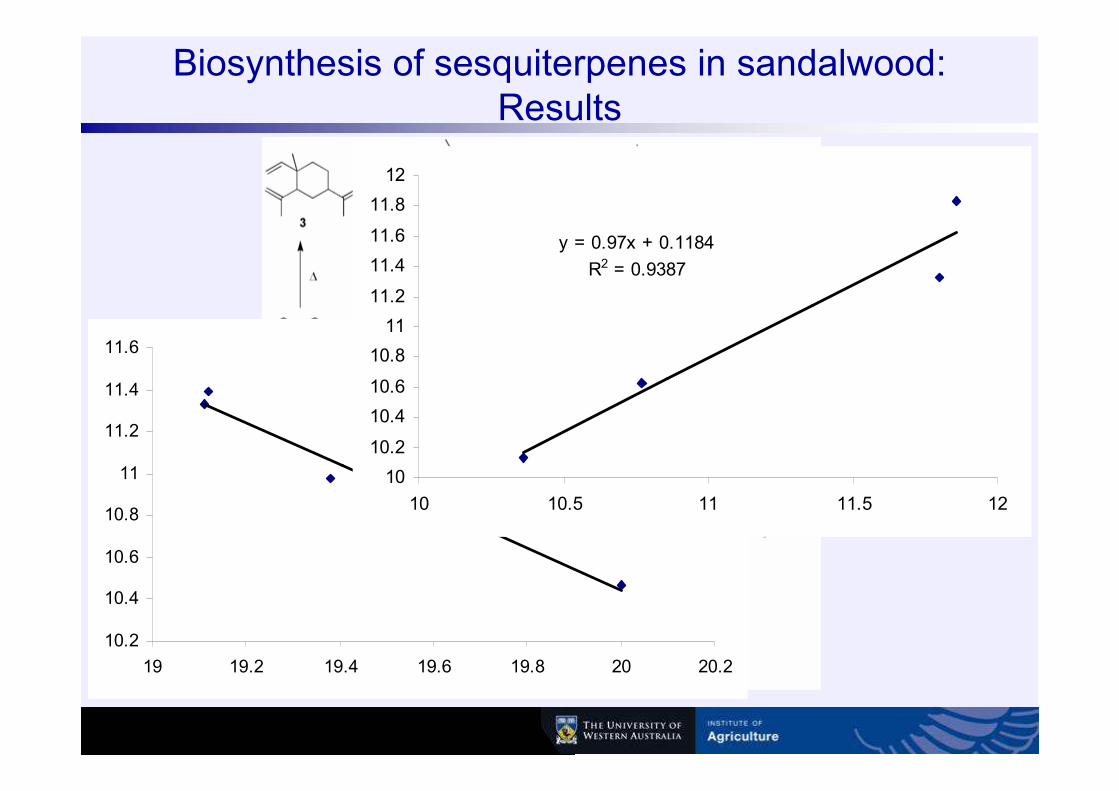
Biosynthesis of sesquiterpenes in sandalwood: Results
y = -1.0001x + 30.447
R2 = 0.9774
10.2
10.4
10.6
10.8
11
11.2
11.4
11.6
19 19.2 19.4 19.6 19.8 20 20.2
y = 0.97x + 0.1184
R2 = 0.9387
10
10.2
10.4
10.6
10.8
11
11.2
11.4
11.6
11.8
12
10 10.5 11 11.5 12

Conclusions
� Lack of genetic diversity in S. album is due to restrictions to gene flow; mostly biogeographic.
� Limited essential oil diversity may also be explained by high homozygousity
�Heartwood oil yield is still highly variable; genetic control may be real, but not detected with the methods used here
�Terpene synthases are active in sandalwood, and are capable of producing multiple sesquiterpene products
�Co-occurrence patterns help explain chemical mechanism responsible for oil biosynthesis.

Future work
�Establish trials of S. album under different growing conditions to explore oil initiation factors
�Explore whether soil rhizobia influences oil production
�Understand physiology of heartwood formation
� Isolate, functionally characterise all of the TPS genes in sandalwood
�Compare gene structure and promoter / regulatory mechanisms of the TPS pathway among species
�Create and validate genetic markers for increased oil production

Acknowledgements
Julie Plummer, Emilio Ghisalberti, Liz Barbour, Margaret Byrne
The Bohlmann lab at UBC
Funding:
Australian Research Council, Forest Products Commission, DAFF Science and Innovation Awards,
Mike Carroll travelling fellowship, Postgraduate travel award, Mary Janet Lindsay travel award

Thankyou!





























