Embed Size (px)
Citation preview

S
Chimeric a2-, frz-Adrenergic Receptors:Delineation of Domains Involved in EffectorCoupling and Ligand Binding Specificity
BRIAN K. KOBILKA, TONG SUN KOBILKA, KIEFER DANIEL,JOHN W. REGAN, MARc G. CARON, ROBERT J. LEFKOWITZ
The a2 and I2 adrenergic receptors, both of which areactivated by epinephrine, but which can be differentiatedby selective drugs, have opposite effects (inhibitory andstimulatory) on the adenylyl cyclase system. The tworeceptors are homologous with each other, rhodopsin,and other receptors coupled to guanine nucleotide regula-tory proteins and they contain seven hydrophobic do-mains, which may represent transmembrane spanningsegments. The function of specific structural domains ofthese receptors was determined after construction andexpression ofa series of chimeric a2-,42-adrenergic recep-tor genes. The specificity for coupling to the stimulatoryguanine nucleotide regulatory protein lies within a regionextending from the amino terminus of the fifth hydro-phobic domain to the carboxyl terminus of the sixth.Major determinants of a2- and P2-adrenergic receptoragonist and antagonist ligand binding specificity are con-tained within the seventh membrane spanning domain.Chimeric receptors should prove useful for elucidatingthe structural basis of receptor fimction.
T t HE ADRENERGIC RECEPTORS (al-, a2-, 1I-, AND P2-),which mediate the physiological effects of catecholamines,belong to the family of plasma membrane receptors that are
coupled to guanine nucleotide regulatory proteins (G proteins) (1).This receptor family also includes rhodopsin and the visual coloropsins, the muscarinic cholinergic receptors, and many other neuro-transmitter receptors and receptors for peptide hormones. A com-mon feature ofG protein-coupled receptors is that agonist occupan-cy of the receptor leads to receptor activation of a G protein, whichin turn modulates the activity of an effector enzyme or ion channel.Several of the G protein-coupled receptors (including the majorsubtypes of adrenergic receptors) have been cloned and found toshare structural features with rhodopsin (2). The most consistentlyconserved of these features is the existence of seven clusters ofhydrophobic amino acids. In addition, there is significant aminoacid sequence similarity among these receptors, which is moststriking in the hydrophobic domains. For bovine rhodopsin, physi-cal and biochemical studies have revealed that these hydrophobicdomains may form seven alpha helices that span the lipid bilayer (3).It has been suggested that these alpha helices form a pocket for thechromophore 1 1-cis-retinal (3). Thus, in an analogous fashion, the
hydrophobic domains ofthe adrenergic receptors may form a pocketin the plasma membrane for binding ligands.
Because so many different hormones, neurotransmitters, and drugreceptors are likely to have structures homologous with the adrener-gic receptors, it is necessary to achieve an understanding of thestructural basis for the various functional properties of these recep-tors, in particular the specificity of ligand binding and effectorcoupling. This has been done heretofore (i) by mutagenesis, espe-cially the deletion of specific peptide sequences (4-6), and (ii)biochemically, where proteases have been used to cleave definedpeptide segments from the digitonin solubilized receptor (7). Thesemethods, although useful in delineating regions of the receptor thatdo not influence its function, suffer from difficulties in that it isdifficult to draw compelling inferences about the role of specificdomains based on loss of functions.
In order to circumvent such problems, and to establish a poten-tially general approach to the study ofG protein-coupled receptorsso that positive inferences can be drawn about functions associatedwith specific receptor domains, we have constructed and expressed aseries of chimeric a2,02-adrenergic receptor genes. All of thesubtypes of adrenergic receptors are activated by epinephrine, butthey differ in their affinity for various subtype selective agonists andantagonists. Furthiermore, the 02-adrenergic receptors (132-AR's)couple to G. (the stimulatory G protein for adenylyl cyclase) whilethe a2-adrenergic receptors (a2-AR's) couple to Gi (the inhibitory Gprotein for adenylyl cyclase). These two receptors therefore, respec-tively, stimulate and inhibit the enzyme. By studying the ligandbinding and adenylyl cyclase activating properties of these chimericreceptors, in which various regions of the a2- and P2-adrenergicreceptors have been interchanged, we have deduced structuraldomains that determine the specificity ofligand binding and effectorcoupling.The a2- and 132-adrenergic receptors. We have described the
cloning of the genes for both the human a2-AR (8) and the human2-AR (9). Both genes have been expressed in Xenopus laevis oocytesby injecting the oocytes with receptor-specific mRNA (8, 10).Receptors expressed in this way can be detected by binding tospecific radioactively labeled ligands. ['25I]Cyanopindolol can beused to detect expressed 132-AR (10). The P2-AR expressed inXenopus oocyte membranes has an affinity for ['25I]cyanopindolol of63 pM and has a typical 2-AR agonist order of potency, with
The authors are at the Howard Hughes Medical Institute, Departments of Medicine,Biochemistry and Physiology, Duke University Medical Center, Durham, NC 27710.
SCIENCE, VOL. 2401310
on
Janu
ary
23, 2
008
www.
scie
ncem
ag.o
rgDo
wnlo
aded
from

isoproterenol (3-AR agonist) being more potent than epinephrine(a2- and 3-AR agonist), which in turn is much more potent than p-aminoclonidine (a2-AR agonist) (Table 1). These agonists, with theexception of p-aminoclonidine, stimulate 2-AR's, expressed inXenopus oocyte membranes, to activate endogeneous adenylyl cy-clase (Table 2).
In contrast to the 2-AR, a2-AR expressed in Xenopus oocytescannot be detected with [125I]cyanopindolol, but instead binds[3H]yohimbine (a2-AR antagonist) with high affinity (2.5 nM).Competition binding studies with [3H]yohimbine for a2-AR ex-pressed in Xenopus oocytes show a typical a2-AR agonist order ofpotency, withp-aminoclonidine (a2-AR agonist) being more potentthan epinephrine (a2- and (-AR agonist), which is much morepotent than isoproterenol ((3-AR agonist). These binding studies ona2-AR expressed in Xenopus oocytes (8) are in agreement withstudies on a2-AR expressed in simian COS-7 cells (Table 3). Thus,like the 132-AR, expression of the a2-AR in Xenopu oocyte mem-
Fig. 1. (A) Diagram ofthe wild-type, a2-adrener- Agic receptor (a2-AR) and the wild-type, 12 recep- 1 2tor (2-AR). The hydrophobic domains are a2ARshown as forming a-helices that span the plasma L CY-, YCOfNmembrane. These putative a helices are numbered i AC -1 to 7 from NH2-terminus (extracellular) to theCOOH-terminus (intracellular). (B) Chimeric re-ceptors made from combinations of the wild-typea2-AR and 12-AR. The a2-AR sequence is indi- Bcated by a solid line and 02-AR sequence is CR1indicated by an open line. The a2-AR (a) and 12- 14007E413E)AR (13) amino acid sequences from NH2- to L CYP+. YOH-
iL AC+COOH-terminus are indicated in parentheses be- K AC+>EP>PACside each chimeric receptor. (C) Split receptors. COOHcSR(1-5) represents a truncation of the 2-AR NHafter amino acid 262 while SR(6-7) represents CR2 rthe 2-AR in which amino acids 3 to 261 have 414r17go413) ^been deleted. Beside each receptor is a summary L AC+of the functional characteristics of the receptor i tO>EP >PACexpressed inXenous Iaevis oocytes or COS-7 cells(or both). The functional properties include: (i) CR3the ability to bind the 12-AR antagonist 125I- R3labeled cyanopindolol (CYP) and the a2-AR- L CYP+ YOH-specific antagonist [3H]yohimbine (YOH); (ii) IL O>PAC>ethe ability to couple to G, and activate adenylyl COOHKcyclase (AC) after stimulation by epinephrine; NHand (iii) the relative potency ofthe a2- and 132-AR CR4agonist epinephrine (EPI), the a2-AR receptor- L CYP+) OM )specific agonist p-aminoclonidine (PAC), and the . AC-1-AR-specific agonist isoproterenol (ISO) for r NDthe receptor as determined by ligand binding COOh'studies or adenylyl cyclase activation (or both). N
(ND, not determined.) Chimeric and split recep- 1CR5tor genes were constructed by splicing desired L CYP-- YOH-restriction endonuclease fragments from the wild- i AC-IL NDtype receptor genes with synthetic oligonucleo-tide adapters. The restriction endonuclease frag-ments encoding the desired structural domains of Cthe a2-AR and 12-AR were isolated by prepara- NK
tive agarose gel electrophoresis. DNA sequences SR(1-5)encoding amino acids not encoded by the DNA in WM).these fragments were synthesized (Applied Bio- L AC-systems model 380 B DNA synthesizer), so thatthe restriction fragments from the wild-type re- Fr ;ceptors plus the synthetic oligonucleotide adapt- SR(15)+SR('7)ers together contain all sequences necessary to NACP+YOH-encode a complete chimeric receptor.The oligo- N. 1S>M>PACnucleotides were phosphorylated at the 5' hy-droxyl and annealed before the ligation reaction.The recombinant genes were identified by restriction endonuclease mapping,and the splice junctions were evaluated by dideoxy sequencing with the useofa denatured double-stranded DNA template (14). To ensure uniformity inthe expression of the chimeric receptor, split receptors, and wild-typereceptors, the 3' and 5' untranslated regions of all genes were derived fromthe 12-AR cDNA (9). Receptor genes were expressed in Xenopus laevis
3 JUNE I988
branes can be documented and characterized by ligand binding.However, unlike the 132-AR, a functional interaction of adenylylcyclase with the a2-AR expressed in Xenopus oocyte membranes hasnot been observed. Thus, stimulation of a2-AR in Xenopus oocytemembranes does not lead to inhibition of adenylyl cyclase activity.Chimeric receptors. To determine which structural domains of
these two receptors confer specificity for agonist and antagonistbinding as well as G protein coupling, we constructed ten chimericreceptor genes from the human 132-AR and human platelet a2-ARgenes. These chimeric receptor genes were expressed in Xenopusoocytes and COS-7 cells, and the ability of the chimeric receptors tobind P2-AR- and a2-AR-specific ligands and to activate adenylylcyclase was determined. When the ligand binding properties and Gprotein-coupling specificities of the various chimeric receptors arecorrelated with the a2-AR and P32-AR amino acid sequences ofthesechimeric receptors, it is possible to assign functional properties tospecific structural domains.
3 4 5 6 7 1 2 3 4 5 6 7'N2 NH2
L CYAC.YOH-COOH ACCrL ISO>EPI>PAC
COON00H
~ ~ ~ ~~ CR6
IXI-~~~~~~(1295)a(U7-M5 ))-L CYP-, YOH+L AC+
I L EP4ISO8>PAC
-C
LCYP-. YOH-L AC-
I ~~~~~COOHKN
'2CR8
u(1.173).(1l744A M747)L CYP-, YOH+I AC+& PAC>EPI>ISO
'2
CR9
L CYP-, YOH4.L AC+PAC>EPIBO8
CR10
u(1.173).P(174.262),A(34445O)-L CYP-, YOH-L AC-
IL ND
12
COOH0
coo -~~~CO
SR(6-7)P(1.2).P23.413)L CYP-. YOH-
l AC-
NH2 IiL ND
oocytes by injecting oocytes with mRNA transcribed from the receptor geneligated into pSP65 as described for the a2-AR and 12-AR (8, 10). Expressionof the genes in COS-7 cells was done by transfecting cells with genes clonedinto pBC12MI in the presence of DEAE-dextran (15). Adenylyl cyclase andligand binding assays are described below.
RESEARCH ARTICLES I3I
on
Janu
ary
23, 2
008
www.
scie
ncem
ag.o
rgDo
wnlo
aded
from
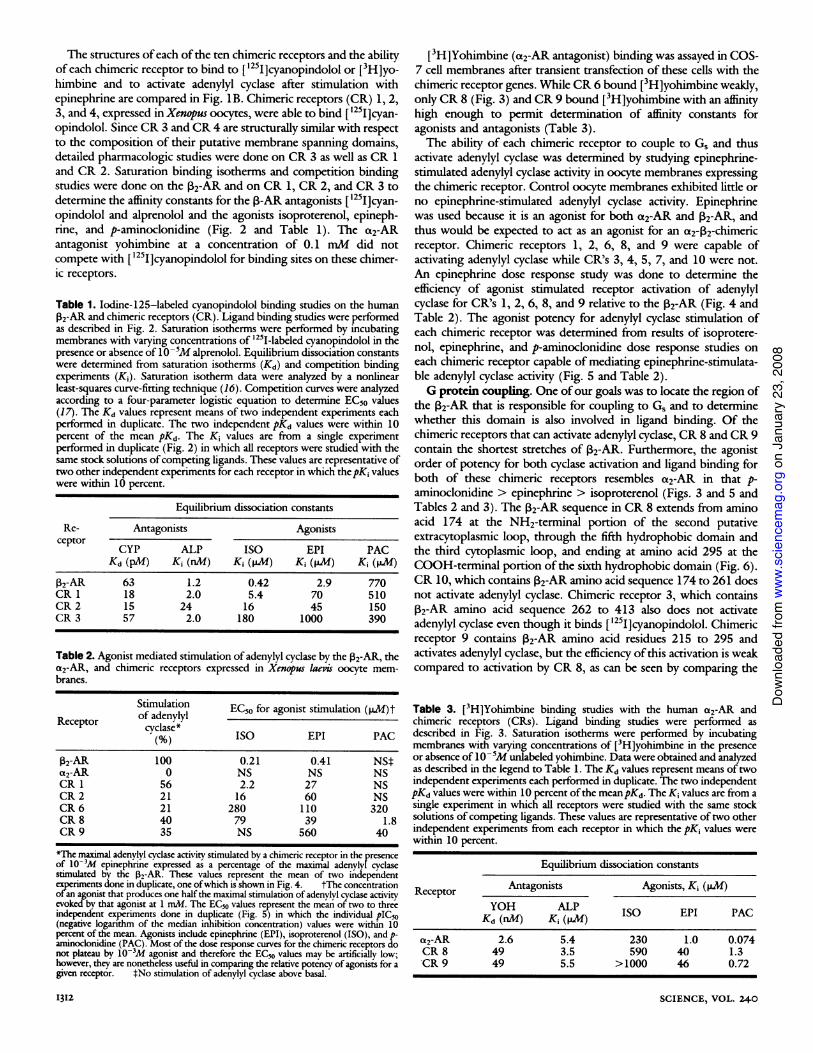
The structures ofeach ofthe ten chimeric receptors and the abilityof each chimeric receptor to bind to ['25I]cyanopindolol or [3H]yo-himbine and to activate adenylyl cyclase after stimulation withepinephrine are compared in Fig. lB. Chimeric receptors (CR) 1, 2,3, and 4, expressed in Xenpus oocytes, were able to bind [ '25I]cyan-opindolol. Since CR 3 and CR 4 are structurally similar with respectto the composition of their putative membrane spanning domains,detailed pharmacologic studies were done on CR 3 as well as CR 1and CR 2. Saturation binding isotherms and competition bindingstudies were done on the P2-AR and on CR 1, CR 2, and CR 3 todetermine the affinity constants for the p-AR antagonists [1251]cyan-opindolol and alprenolol and the agonists isoproterenol, epineph-rine, and p-aminoclonidine (Fig. 2 and Table 1). The a2-ARantagonist yohimbine at a concentration of 0.1 mM did notcompete with ['25I]cyanopindolol for binding sites on these chimer-ic receptors.
Table 1. Iodine-125-labeled cyanopindolol binding studies on the humanP2-AR and chimeric receptors (CR). Ligand binding studies were performedas described in Fig. 2. Saturation isotherms were performed by incubatingmembranes with varying concentrations of '251-labeled cyanopindolol in thepresence or absence of 10-5M alprenolol. Equilibrium dissociation constantswere determined from saturation isotherms (Kd) and competition bindingexperiments (K1). Saturation isotherm data were analyzed by a nonlinearleast-squares curve-fitting technique (16). Competition curves were analyzedaccording to a four-parameter logistic equation to determine EC50 values(17). The Kd values represent means of two independent experiments eachperformed in duplicate. The two independent pKd values were within 10percent of the mean pKd. The Ki values are from a single experimentperformed in duplicate (Fig. 2) in which all receptors were studied with thesame stock solutions ofcompeting ligands. These values are representative oftwo other independent experiments for each receptor in which thepK, valueswere within 10 percent.
Equilibrium dissociation constants
Re- Antagonists Agonistsceptor CYP ALP ISO EPI PAC
Kd (pM) Ki (nM) Ki (p.M) Ki (IpM) Ki (pM1)VAR 63 1.2 0.42 2.9 770CR 1 18 2.0 5.4 70 510CR 2 15 24 16 45 150CR 3 57 2.0 180 1000 390
Table 2. Agonist mediated stimulation of adenylyl cyclase by the 132-AR, thea2-AR, and chimeric receptors expressed in Xenopus laevis oocyte mem-branes.
Stimu:lation EC50 for agonist stimulation (PM)tReceptor of adenylyl
cyclase*cyc%ase) ISO EPI PAC
P2-AR 100 0.21 0.41 NSta2-AR 0 NS NS NSCR 1 56 2.2 27 NSCR 2 21 16 60 NSCR 6 21 280 110 320CR 8 40 79 39 1.8CR 9 35 NS 560 40
*The maximnal adenylyl cyclase activity stimulated by a chimeric receptor in the presenceof 10-3M epinephrine expressed as a percentage of the maximal adenylyfcyclasestimulated by the 2-R. These values represent the mean of two independentexperiments done in duplicate, one ofwhich is shown in Fig. 4. tThe concentrationofan agonist that produces one half the maximal stimulation of adenylvl cyclase activityevoked by that agonist at 1 mM. The EC50 values represent the mean of two to threeindependent experiments done in duplicate (Fig. 5) in which the individual pIC,o(negative logarithm of the median inhibition concentration) values were within 10percent of the mean. Agonists include epinephrine (EPI), isoproterenol (ISO), and p-aminoclonidine (PAC). Most of the dose response curves for the chimeric receptors donot plateau by 10-3M agonist and therefore the EC50 values mav be artificially low;however, they are nonetheless useful in comparing the relative potency of agonists for agiven receptor. tNo stimulation of adenylyl cydase above basal.
1312
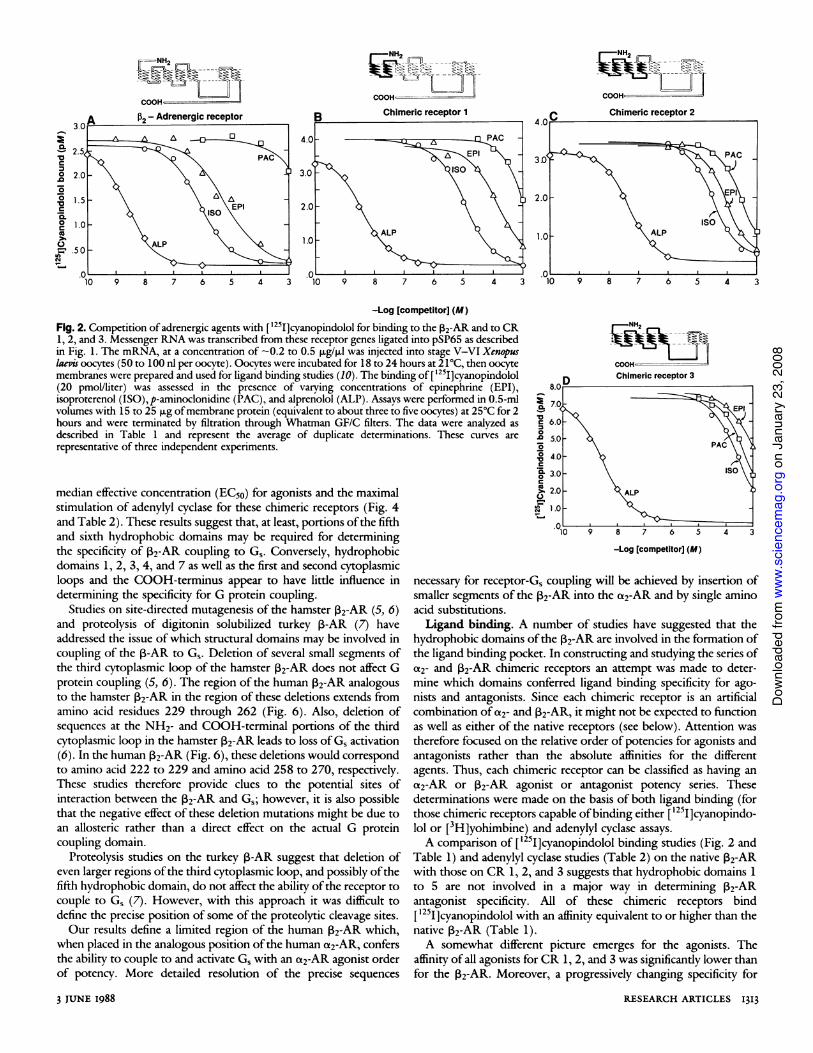
[3H]Yohimbine (a2-AR antagonist) binding was assayed in COS-7 cell membranes after transient transfection of these cells with thechimeric receptor genes. While CR 6 bound [3H]yohimbine weakly,only CR 8 (Fig. 3) and CR 9 bound [3H]yohimbine with an affinityhigh enough to permit determination of affinity constants foragonists and antagonists (Table 3).The ability of each chimeric receptor to couple to G, and thus
activate adenylyl cyclase was determined by studying epinephrine-stimulated adenylyl cyclase activity in oocyte membranes expressingthe chimeric receptor. Control oocyte membranes exhibited little orno epinephrine-stimulated adenylyl cyclase activity. Epinephrinewas used because it is an agonist for both c2-AR and 2-AR, andthus would be expected to act as an agonist for an a2-V2-chimericreceptor. Chimeric receptors 1, 2, 6, 8, and 9 were capable ofactivating adenylyl cyclase while CR's 3, 4, 5, 7, and 10 were not.An epinephrine dose response study was done to determine theefficiency of agonist stimulated receptor activation of adenylylcyclase for CR's 1, 2, 6, 8, and 9 relative to the P2-AR (Fig. 4 andTable 2). The agonist potency for adenylyl cyclase stimulation ofeach chimeric receptor was determined from results of isoprotere-nol, epinephrine, and p-aminoclonidine dose response studies oneach chimeric receptor capable of mediating epinephrine-stimulata-ble adenylyl cyclase activity (Fig. 5 and Table 2).G protein coupling. One ofour goals was to locate the region of
the 2-AR that is responsible for coupling to Gs and to determinewhether this domain is also involved in ligand binding. Of thechimeric receptors that can activate adenylyl cyclase, CR 8 and CR 9contain the shortest stretches of 2-AR. Furthermore, the agonistorder of potency for both cyclase activation and ligand binding forboth of these chimeric receptors resembles ct2-AR in that p-aminoclonidine > epinephrine > isoproterenol (Figs. 3 and 5 andTables 2 and 3). The 2-AR sequence in CR 8 extends from aminoacid 174 at the NH2-terminal portion of the second putativeextracytoplasmic loop, through the fifth hydrophobic domain andthe third cytoplasmic loop, and ending at amino acid 295 at theCOOH-terminal portion of the sixth hydrophobic domain (Fig. 6).CR 10, which contains 2-AR amino acid sequence 174 to 261 doesnot activate adenylyl cyclase. Chimeric receptor 3, which contains2-AR amino acid sequence 262 to 413 also does not activateadenylyl cyclase even though it binds [1251]cyanopindolol. Chimericreceptor 9 contains 2-AR amino acid residues 215 to 295 andactivates adenylyl cyclase, but the efficiency of this activation is weakcompared to activation by CR 8, as can be seen by comparing the
Table 3. [3H]Yohimbine binding studies with the human a2-AR andchimeric receptors (CRs). Ligand binding studies were performed asdescribed in Fig. 3. Saturation isotherms were performed by incubatingmembranes with varying concentrations of [3H]yohimbine in the presenceor absence of 105M unlabeled yohimbine. Data were obtained and analyzedas described in the legend to Table 1. The Kd values represent means of twoindependent experiments each performed in duplicate. The two independentpKd values were within 10 percent ofthe meanpKd. The Kj values are from asingle experiment in which all receptors were studied with the same stocksolutions of competing ligands. These values are representative oftwo otherindependent experiments from each receptor in which the pKj values werewithin 10 percent.
Equilibrium dissociation constants
Receptor Antagonists Agonists, Kj (jiM)YOH ALP ISO EPI PACKd (nM) K (p.M) IS EP PA
a2-AR 2.6 5.4 230 1.0 0.074CR 8 49 3.5 590 40 1.3CR 9 49 5.5 >1000 46 0.72
SCIENCE, VOL. 240
on
Janu
ary
23, 2
008
www.
scie
ncem
ag.o
rgDo
wnlo
aded
from
NHA
COOH.
P2- Adrenergic receptor
-NH2
MOOH
Chimeric receptor 1
cooH~~ ~ ~ ~ ~
COOHi
Chimeric receptor 2
-Log [competitor] (M)
Fig. 2. Competition of adrenergic agents with ['25I]cyanopindolol for binding to the P2-AR and to CR1, 2, and 3. Messenger RNA was transcribed from these receptor genes ligated into pSP65 as describedin Fig. 1. The mRNA, at a concentration of -0.2 to 0.5 ,ug/,d was injected into stage V-VI Xenopuslaepis oocytes (50 to 100 nl per oocyte). Oocytes were incubated for 18 to 24 hours at 21°C, then oocytemembranes were prepared and used for ligand binding studies (10). The binding of ['251]cyanopindolol(20 pmollliter) was assessed in the presence of varying concentrations of epinephrine (EPI),isoproterenol (ISO), p-aminoclonidine (PAC), and alprenolol (ALP). Assays were performed in 0.5-milvolumes with 15 to 25 ,g ofmembrane protein (equivalent to about three to five oocytes) at 25°C for 2hours and were terminated by filtration through Whatman GF/C filters. The data were analyzed asdescribed in Table 1 and represent the average of duplicate determinations. These curves arerepresentative of three independent experiments.
median effective concentration (EC5o) for agonists and the maximalstimulation of adenylyl cyclase for these chimeric receptors (Fig. 4and Table 2). These results suggest that, at least, portions ofthe fifthand sixth hydrophobic domains may be required for determiningthe specificity of V32-AR coupling to G,. Conversely, hydrophobicdomains 1, 2, 3, 4, and 7 as well as the first and second cytoplasmicloops and the COOH-terminus appear to have little influence indetermining the specificity for G protein coupling.
Studies on site-directed mutagenesis of the hamster V2-AR (5, 6)and proteolysis of digitonin solubilized turkey ,-AR (7) haveaddressed the issue of which structural domains may be involved incoupling of the ,B-AR to G,. Deletion of several small segments ofthe third cytoplasmic loop of the hamster 2-AR does not affect Gprotein coupling (5, 6). The region of the human P2-AR analogousto the hamster 132-AR in the region of these deletions extends fromamino acid residues 229 through 262 (Fig. 6). Also, deletion ofsequences at the NH2- and COOH-terminal portions of the thirdcytoplasmic loop in the hamster ,B2-AR leads to loss of G, activation(6). In the human 2-AR (Fig. 6), these deletions would correspondto amino acid 222 to 229 and amino acid 258 to 270, respectively.These studies therefore provide clues to the potential sites ofinteraction between the V2-AR and G,; however, it is also possiblethat the negative effect of these deletion mutations might be due toan allosteric rather than a direct effect on the actual G proteincoupling domain.
Proteolysis studies on the turkey 3-AR suggest that deletion ofeven larger regions ofthe third cytoplasmic loop, and possibly ofthefifth hydrophobic domain, do not affect the ability ofthe receptor tocouple to G, (7). However, with this approach it was difficult todefine the precise position of some of the proteolytic cleavage sites.Our results define a limited region of the human ,32-AR which,
when placed in the analogous position of the human a2-AR, confersthe ability to couple to and activate G, with an a2-AR agonist orderof potency. More detailed resolution of the precise sequences
3 JUNE I988
NH2
COOHJChimeric receptor 3
QCL30
0
IC0ccoL)
n
-Log [competitor] (M)
necessary for receptor-Gs coupling will be achieved by insertion ofsmaller segments of the 132-AR into the a2-AR and by single aminoacid substitutions.Ligand binding. A number of studies have suggested that the
hydrophobic domains ofthe 132-AR are involved in the formation ofthe ligand binding pocket. In constructing and studying the series ofa2- and P2-AR chimeric receptors an attempt was made to deter-mine which domains conferred ligand binding specificity for ago-nists and antagonists. Since each chimeric receptor is an artificialcombination of a2- and 2-AR, it might not be expected to functionas well as either of the native receptors (see below). Attention was
therefore focused on the relative order of potencies for agonists andantagonists rather than the absolute affinities for the differentagents. Thus, each chimeric receptor can be classified as having an
a2-AR or j32-AR agonist or antagonist potency series. Thesedeterminations were made on the basis of both ligand binding (forthose chimeric receptors capable ofbinding either ['251]cyanopindo-lol or [3H]yohimbine) and adenylyl cyclase assays.A comparison of ['25I]cyanopindolol binding studies (Fig. 2 and
Table 1) and adenylyl cyclase studies (Table 2) on the native 132-ARwith those on CR 1, 2, and 3 suggests that hydrophobic domains 1
to 5 are not involved in a major way in determining 2-ARantagonist specificity. All of these chimeric receptors bind[1251]cyanopindolol with an affinity equivalent to or higher than thenative ,32-AR (Table 1).A somewhat different picture emerges for the agonists. The
affinity of all agonists for CR 1, 2, and 3 was significantly lower thanfor the 132-AR. Moreover, a progressively changing specificity for
RESEARCH ARTICLES 1313
Q0.v
c
300
io._
Qla0
0C0
C.
04
on
Janu
ary
23, 2
008
www.
scie
ncem
ag.o
rgDo
wnlo
aded
from
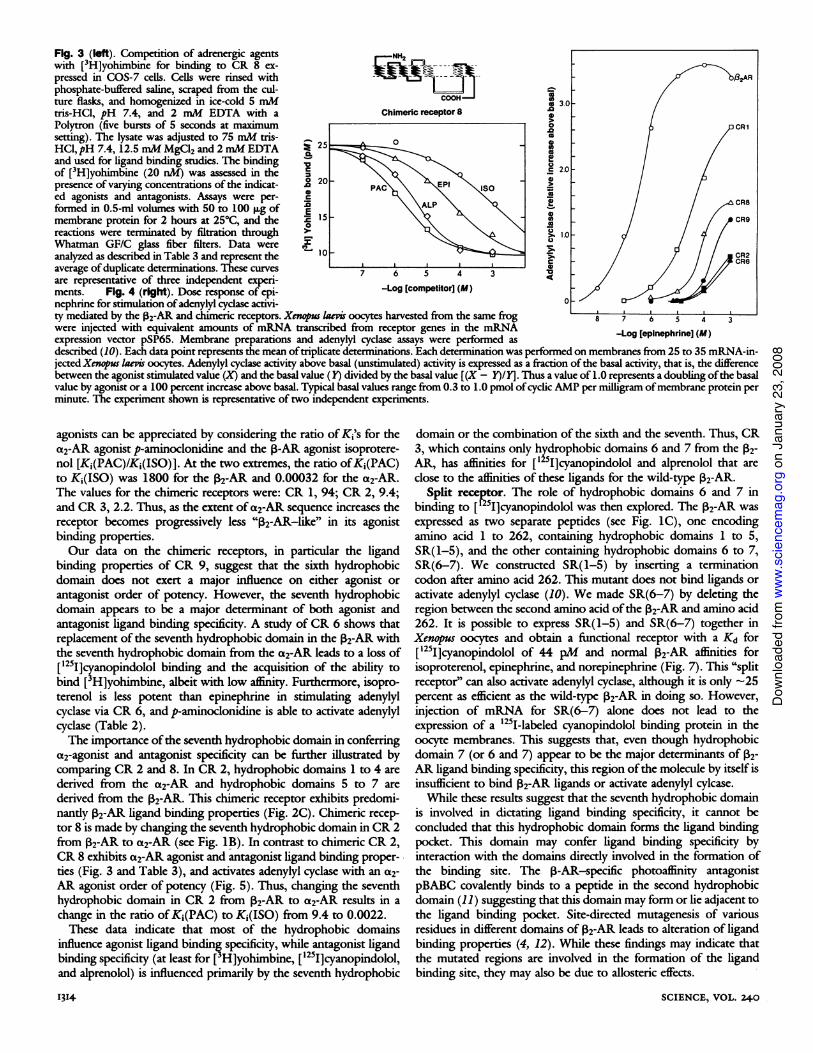
Fig. 3 (left). Competition of adrenergic agents NHwith [3H]yohimbine for binding to CR 8 ex- - -->pressed in COS-7 cells. Cells were rinsed with -2--S-¶ I,ARphosphate-buffered saline, scraped from the cul- I1Jture flasks, and homogenized in ice-cold 5 mM .8 3.0tris-HCI, pH 7.4, and 2 mM EDTA with a Chimeric receptor 8 /Polytron (five bursts of 5 seconds at maximum a CR1setting). The lysate was adjusted to 75 mM tris- 2 XIHCI, pH 7.4,12.5 mM MgCI2 and 2 mM EDTA > 25 =and used for ligand binding studies. The binding j 2.0of [3H]yohimbine (20 nM) was assessed in the 2.0 -
presence of varying concentrations of the indicat- o20 PPied agonists and antagonists. Assays were per- IC CR8formed in 0.5-ml volumes with 50 to 100 Rg of E \Amembrane protein for 2 hours at 250C, and the O 15 _ CR9reactions were terminated by filtration through > lo /Whatman GF/C glass fiber filters. Data were
1\ I
analyzed as described in Table 3 and represent the 0> CR2average of duplicate determinations. These curves 7 6 5 4 3are representative of three independent experi-ments. Fig. 4 (right). Dose response of epi- -Log [competitor] (M)nephrine for stimulation ofadenylyl cydase activi- 0ty mediated by the ,82-AR and chimeric receptors. Xnopus laevis oocytes harvested from the same frog 8 7 6 5 4 3were injected with equivalent amounts of mRNA transcribed from receptor genes in the mRNA -Log [epinephrine] (M)expression vector pSP65. Membrane preparations and adenylyl cyclase assays were performed asdescribed (10). Each data point represents the mean oftriplicate determinations. Each determination was performed on membranes from 25 to 35 mRNA-in-jected Xenps iaevis oocytes. Adenylyl cyclase activity above basal (unstimulated) activity is expressed as a fraction of the basal activity, that is, the differencebetween the agonist stimulated value (X) and the basal value (1Y) divided by the basal value [(X - Y)/Y]. Thus a value of 1.0 represents a doubling ofthe basalvalue by agonist or a 100 percent increase above basal. Typical basal values range from 0.3 to 1.0 pmol ofcyclic AMP per milligram ofmembrane protein perminute. The experiment shown is representative of two independent experiments.
agonists can be appreciated by considering the ratio of Ki's for thea2-AR agonist p-aminoclonidine and the 3-AR agonist isoprotere-nol [Ki(PAC)/K1(ISO)]. At the two extremes, the ratio ofKi(PAC)to K,(ISO) was 1800 for the 132-AR and 0.00032 for the a2-AR.The values for the chimeric receptors were: CR 1, 94; CR 2, 9.4;and CR 3, 2.2. Thus, as the extent ofa2-AR sequence increases thereceptor becomes progressively less "B2-AR-like" in its agonistbinding properties.Our data on the chimeric receptors, in particular the ligand
binding properties of CR 9, suggest that the sixth hydrophobicdomain does not exert a major influence on either agonist or
antagonist order of potency. However, the seventh hydrophobicdomain appears to be a major determinant of both agonist andantagonist ligand binding specificity. A study of CR 6 shows thatreplacement of the seventh hydrophobic domain in the 132-AR withthe seventh hydrophobic domain from the a2-AR leads to a loss of[125I]cyanopindolol binding and the acquisition of the ability tobind [3H]yohimbine, albeit with low affinity. Furthermore, isopro-terenol is less potent than epinephrine in stimulating adenylylcyclase via CR 6, and p-aminoclonidine is able to activate adenylylcyclase (Table 2).The importance ofthe seventh hydrophobic domain in conferring
a2-agonist and antagonist specificity can be further illustrated bycomparing CR 2 and 8. In CR 2, hydrophobic domains 1 to 4 are
derived from the a2-AR and hydrophobic domains 5 to 7 are
derived from the 132-AR. This chimeric receptor exhibits predomi-nantly 12-AR ligand binding properties (Fig. 2C). Chimeric recep-tor 8 is made by changing the seventh hydrophobic domain in CR 2from 2-AR to a2-AR (see Fig. 1B). In contrast to chimeric CR 2,CR 8 exhibits a2-AR agonist and antagonist ligand binding proper-ties (Fig. 3 and Table 3), and activates adenylyl cyclase with an a2-AR agonist order of potency (Fig. 5). Thus, changing the seventhhydrophobic domain in CR 2 from P2-AR to a2-AR results in a
change in the ratio of Ki(PAC) to K1(ISO) from 9.4 to 0.0022.These data indicate that most of the hydrophobic domains
influence agonist ligand binding specificity, while antagonist ligandbinding specificity (at least for [ H]yohimbine, ['25I]cyanopindolol,and alprenolol) is influenced primarily by the seventh hydrophobic
I3I4
domain or the combination of the sixth and the seventh. Thus, CR3, which contains only hydrophobic domains 6 and 7 from the 12-AR, has affinities for [1 5I]cyanopindolol and alprenolol that areclose to the affinities of these ligands for the wild-type 132-AR.
Split receptor. The role of hydrophobic domains 6 and 7 inbinding to [125I]cyanopindolol was then explored. The 132-AR wasexpressed as two separate peptides (see Fig. 1C), one encodingamino acid 1 to 262, containing hydrophobic domains 1 to 5,SR(1-5), and the other containing hydrophobic domains 6 to 7,SR(6-7). We constructed SR(1-5) by inserting a terminationcodon after amino acid 262. This mutant does not bind ligands oractivate adenylyl cyclase (10). We made SR(6-7) by deleting theregion between the second amino acid ofthe 32-AR and amino acid262. It is possible to express SR(1-5) and SR(6-7) together inXenopus oocytes and obtain a functional receptor with a Kd for[1251]cyanopindolol of 44 pM and normal 2-AR affinities forisoproterenol, epinephrine, and norepinephrine (Fig. 7). This "splitreceptor" can also activate adenylyl cyclase, although it is only -25percent as efficient as the wild-type P2-AR in doing so. However,injection of mRNA for SR(6-7) alone does not lead to theexpression of a '251-labeled cyanopindolol binding protein in theoocyte membranes. This suggests that, even though hydrophobicdomain 7 (or 6 and 7) appear to be the major determinants of 132-AR ligand binding specificity, this region ofthe molecule by itself isinsufficient to bind P2-AR ligands or activate adenylyl cylcase.While these results suggest that the seventh hydrophobic domain
is involved in dictating ligand binding specificity, it cannot beconcluded that this hydrophobic domain forms the ligand bindingpocket. This domain may confer ligand binding specificity byinteraction with the domains directly involved in the formation ofthe binding site. The 1-AR-specific photoaffinity antagonistpBABC covalently binds to a peptide in the second hydrophobicdomain (11) suggesting that this domain may form or lie adjacent tothe ligand binding pocket. Site-directed mutagenesis of variousresidues in different domains of 2-R leads to alteration of ligandbinding properties (4, 12). While these findings may indicate thatthe mutated regions are involved in the formation of the ligandbinding site, they may also be due to allosteric effects.
SCIENCE, VOL. 240
on
Janu
ary
23, 2
008
www.
scie
ncem
ag.o
rgDo
wnlo
aded
from
Chimeric receptor 8
1.50
as
.0
0
a
0. a000 c0-
00
00
C
0
7 6 5 4 3
-Log [agonist] (M)
Fig. 5 (left). Adenylyl cyclase activity stimulated via CR 8. Xenopus oocytemembranes containing CR 8 were assayed for adenylyl cyclase at varyingconcentrations of the indicated agonists. The methods are described in Fig.4. Each data point represents the mean of two independent experiments inwhich determinations were done in duplicate. Each determination wasperformed with membranes from 25 to 35 mRNA-injected Xenopus laevisoocytes. Adenylyl cyclase is expressed as described in Fig. 4. Fig. 6(center). 02-AR amino acid sequence of CR 8. The diagram of CR 8 isshown at the top. The open line represents ,82-AR sequence. The amino acidsderived from the P2-AR are shown in the larger diagram as circles. Blackcircles with white letters represent amino acid identities between the a2-AR
Possible arrangement of hydrophobic domains. The study ofthis set of chimeric receptors has provided insight into the functionof various structural domains. However, these molecules may alsoprovide clues about the arrangement of the various hydrophobicdomains within the parent molecules. These hydrophobic domainsmay form a-helices that span the plasma membrane, as has beensuggested by electron diffraction studies on bacteriorhodopsin (13).For the following discussion we therefore refer to the hydrophobicdomains as membrane spanning a-helices. The arrangement ofthesea-helices with respect to each other might be dictated by theinteractions of various charged, noncharged polar, and nonpolaramino acids as well as the possible formation ofcovalent bonds (thatis, disulfide bridges). The less hydrophobic amino acids of these ahelices are likely to project toward the interior ofthe molecule, whilethe more hydrophobic residues may form a boundary with theplasma membrane. The a helices that lie adjacent to each other havepresumably evolved in such a way as to minimize steric andelectrostatic repulsive forces between each other.
In the process of making chimeric receptors these favorablemolecular interactions may be lost. For example, in making CR 1, a-helix 2 from the a2-AR and a-helix 3 from the 2-AR are forced tolie adjacent to each other. Furthermore, other a helices that maynormally cluster around a-helices 1 and 2 from the V2-AR may beless compatible with a-helices 1 and 2 from the a2-AR. Thesemolecular incompatibilities might be expected to destabilize themolecule, and thus make it less efficient or even nonfunctional. Thisis consistent with the finding that all chimeric receptors in this seriesare less efficient than either parent molecule in binding agonists, andless efficient than the 2-AR in activating G,. Furthermore, chimericreceptors containing two molecular splice junctions such as CR 7, 8,9, and 10 (Fig. 1), might be expected to function even less well thanchimeric receptors with only one splice junction. For example, whileCR 1 and CR 6 are both able to couple to and activate G,, CR 7,
3 JUNE I988
SpNtreeporSR1-)H S(NH
Split receptor SR(1-5) + SR(6-7)
7 6 5-Log [competitor] (M)
and 02-AR. The numbered amino acid residues are discussed in thetext. Fig. 7 (right). Agonist competition binding studies on splitreceptors. SR(1-5) and SR(6-7) were expressed together in Xenpus Iaeisoocytes. Messenger RNA transcribed from SR(1-5) and SR(6-7) genes inthe mRNA expression vector pSP65 was mixed and injected into Xenpuslaevis oocytes. After 24 hours, membranes were prepared and competition-binding was performed as described in Fig. 2. Varying concentrations ofisoproterenol (ISO), epinephrine (EPI), and norepinephrine (NOREPI)competed for binding sites with 75 pM '25I-labeled cyanopindolol. Datawere analyzed as described in Table 1.
which contains both molecular splice junctions found in CR 1 andCR 6, is nonfunctional even though it contains the essentialelements of P2-AR necessary to activate Gs.
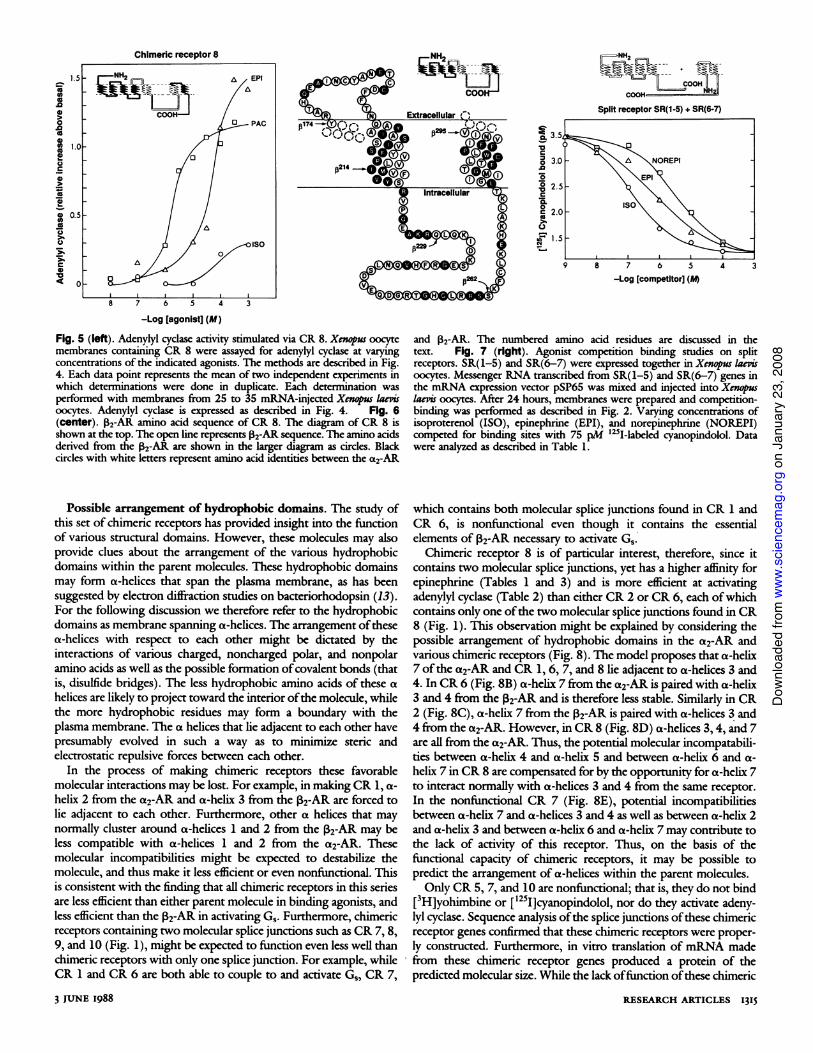
Chimeric receptor 8 is of particular interest, therefore, since itcontains two molecular splice junctions, yet has a higher affinity forepinephrine (Tables 1 and 3) and is more efficient at activatingadenylyl cyclase (Table 2) than either CR 2 or CR 6, each ofwhichcontains only one ofthe two molecular splice junctions found in CR8 (Fig. 1). This observation might be explained by considering thepossible arrangement of hydrophobic domains in the a2-AR andvarious chimeric receptors (Fig. 8). The model proposes that a-helix7 of the a2-AR and CR 1, 6, 7, and 8 lie adjacent to a-helices 3 and4. In CR 6 (Fig. 8B) a-helix 7 from the a2-AR is paired with a-helix3 and 4 from the 2-AR and is therefore less stable. Similarly in CR2 (Fig. 8C), a-helix 7 from the 2-AR is paired with a-helices 3 and4 from the a2-AR. However, in CR 8 (Fig. 8D) a-helices 3,4, and 7are all from the a2-AR. Thus, the potential molecular incompatabili-ties between a-helix 4 and a-helix 5 and between a-helix 6 and a-helix 7 in CR 8 are compensated for by the opportunity for a-helix 7to interact normally with a-helices 3 and 4 from the same receptor.In the nonfunctional CR 7 (Fig. 8E), potential incompatibilitiesbetween a-helix 7 and a-helices 3 and 4 as well as between a-helix 2and a-helix 3 and between a-helix 6 and a-helix 7 may contribute tothe lack of activity of this receptor. Thus, on the basis of thefunctional capacity of chimeric receptors, it may be possible topredict the arrangement of a-helices within the parent molecules.Only CR 5, 7, and 10 are nonfunctional; that is, they do not bind
[3H]yohimbine or [1251]cyanopindolol, nor do they activate adeny-lyl cyclase. Sequence analysis ofthe splice junctions ofthese chimericreceptor genes confirmed that these chimeric receptors were proper-ly constructed. Furthermore, in vitro translation of mRNA madefrom these chimeric receptor genes produced a protein of thepredicted molecular size. While the lack offunction of these chimeric
RESEARCH ARTICLES 13I5
5
vID
NH2
coo
NH2
H
on
Janu
ary
23, 2
008
www.
scie
ncem
ag.o
rgDo
wnlo
aded
from

Fig. 8. Possible arrangement of hy-drophobic domains for the a2-ARand various chimeric receptors. Thehydrophobic domains are depictedas a-helices as they would appearfrom above the plasma membrane(A). Alpha helices from the a2-ARare shown as black circles withwhite numbers while those from02-AR are shown as white circleswith black numbers. Potential de-stabilizing interactions between a-helices from different receptors areindicated by arrows. The modelattempts to explain the observationthat CR 8 (D) functions better thanCR 6 (B), CR 2 (C), or CR 7 (E)as assessed by affinity for epineph-rine (Tables 1 and 3) and by activa-tion of adenylyl cyclase (Table 2).
A oX2AR
B CR6
D CR8
* a2AR nant of both agonist and antagonist ligand binding specificity.O ,e AR Finally, several ofthe first five hydrophobic domains may contribute
to agonist binding specificity. The strength of this approach for thestudy ofthese functionally complex molecules is that conclusions canbe drawn from qualitative changes in receptor function, or from theacquisition of new functions that can be correlated with specificprotein sequences. This is in contrast with more standard mutagene-sis approaches where the end point is the loss offinction resultingfrom amino acid deletions or substitutions. Our results provide an
C CR2 initial structural map for understanding the various functions oftwomodel G protein-coupled receptors. The map requires furtherrefinement and testing of its generality for understanding receptorsof this class.
E CR7
receptors might be explained by molecular incompatibilities be-tween oa-helices as discussed above, it is also possible that thesechimeric receptors failed to insert properly in the plasma membraneas a result of specific amino acid sequences created at the splicejunctions. This may be particularly important for CR 5 and CR 10which have a splice junction in the putative third cytoplasmic loop ina region where the a2-AR and 32-AR share little amino acidsequence similarity.The results from our study of chimeric receptors made from the
2-AR and a2-AR have provided new insights into the functionalrole of several structural domains. The fifth and sixth hydrophobicdomains and the third cytoplasmic loop are capable of conferringspecificity for G, coupling to the P2-AR. The seventh hydrophobicdomain of the a2- and P2-adrenergic receptors is a major determi-
REFERENCES AND NOTES
1. A. G. Gilman,Annu. Rev. Biochem. 56, 615 (1987).2. H. G. Dohlman, M. G. Caron, R. J. Lefkowit, Biochematy 26, 2657 (1987).3. J. B. C. Findlay and D. J. C. Pappin, Biochem. J. 238, 625 (1987).4. R. A. F. Dixon et al., EMBOJ. 6 (11), 3269 (1987).5. R. A. F. Dixon et al., Nature 326, 73 (1987).6. C. D. Strader et al., J. Biol. Chem. 262, 16439 (1987).7. R. C. Rubenstein, S. K.-F. Wong, E. M. Ross, ibid., p. 16655.8. B. K. Kobilka et al., Scinc 238, 650 (1987).9. B. K. Kobilka ct al., Proc. Natl. Acad. Sai. U.S.A. 84, 46 (1987).
10. B. K. Kobilka at al., J. Biol. Chem. 262, 15796 (1987).11. H. G. Dohlman, M. G. Caron, C. D. Strader, N. Amlaiky, R. J. Lefkowitz,
Biochemisty 27, 1813 (1988).12. C. D. Strader ect al., Proc. Nati. Acad. Sci. U.S.A. 84, 4384 (1987).13. R. Henderson and P. N. T. Unwin, Naturc 257, 28 (1975).14. F. Sanger, S. Nicklen, A. R. Coulson, Proc. Natl. Acad. Sci. U.S.A. 74, 5463
(1977).15. B. Cullen, Methods in Enzymlo. 152, 684 (1987).16. A. DeLean, A. A. Hancock, R. J. Lcfkowitz, Mol. Pharmacol. 21, 5 (1982).17. A. DeLean, P. T. Munson, A. Rodbard, Am. J. Physiol. 235, 97 (1978).18. We thank M. Holben for preparation of the manuscript, and 0. Finn for
introducing us to the Xenpus oocyte expression system. Supported in part by NIHgrant HL 16037.25 February 1988; accepted 28 April 1988
1988
AAAS Philip Hauge Abelson PrizeTo Be Awarded to a Public Servant or Scientist
The AAAS Philip Hauge Abelson Prize of $2,500 and acommemorative plaque, established by the AAAS Board ofDirectors in 1985, is awarded annually either to:
(a) a public servant, in recognition of sustained exceptionalcontributions to advanced science, or
(b) a scientist whose career has been distinguished both forscientific achievement and for other notable services to thescientific communitv.AAAS members are invited to submit nominations now for
the 1988 prize, to be awarded at the 1989 Annual Meetingin San Francisco. Each nomination must be seconded by atleast two other AAAS members. The prize recipient will be
selected by a seven-member panel appointed by the Board. Therecipient's travel and hotel expenses incurred in attending theaward presentation will be reimbursed.Nominations should be typed and should include the follow-
ing information: nominee's name, institutional affiliation andtide, address, and brief biographical resume; statement of justifi-cation for nomination; and names, identification, and signaturesof the three or more AAAS member sponsors.
Eight copies of the complete nomination should be submittedto the AAAS Executive Office, 1333 H Street, N.W., Washing-ton, D.C. 20005, for receipt on or before 1 August 1988.
SCIENCE, VOL. 240
LI3I6
on
Janu
ary
23, 2
008
www.
scie
ncem
ag.o
rgDo
wnlo
aded
from





























