Embed Size (px)
Citation preview

Characterization of Sac7 as a Negative Regulator of the Rho1-mediated
Clathrin Independent Endocytic Pathway in Saccharomyces cerevisiae
By
Matthew R. Peck
A thesis submitted to The Johns Hopkins University in conformity with the requirements
for the degree of Master of Science in Molecular and Cellular Biology
Baltimore, Maryland
April 2015
©2015 Matthew Richard Peck
All Rights Reserved

Abstract
Although it had, until recently, been thought that clathrin-mediated endocytosis was the
sole mechanism for endocytosis in Saccharomyces cerevisiae, a clathrin-independent pathway
that utilizes the small GTPase, Rho1, was recently discovered. Small GTPases are positively
regulated by GTP exchange factors, or GEFs, and negatively regulated by GTPase activing
proteins, or GAPs. Relatively little is known about Sac7, the primary GAP of this pathway. This
study presents the finding of multiple proteins that physically interact with Sac7 via a yeast two-
hybrid screen and preliminary evidence regarding the potential roles these proteins may play in
the clathrin-independent endocytic pathway.
ii

Acknowledgements
At this time, I would like to thank those who helped me to complete this thesis.
I would like to thank everybody in the Wendland Lab. This would certainly not be
possible without the active support of Dr. Beverly Wendland and Dr. Derek Prosser. I would also
like to thank the other members of the Wendland Lab: Kate Bradford, Kristie Wrasman, Kyle
Hoban, Alex Wade, Sami Lux, Marjon Khairy, Thaddeus Woodard, Josh Katz, Asia Poprawski,
and Lydia Nyasea.
I would like to thank members of the Integrated Imaging Center, for whom I am very
much in debt for their help in microscopy portions of this thesis: J. Michael McCaffery, Bethany
Ackerman, and Erin Pryce.
Finally, I would also like to thank my friends and family, without whose constant support
I would not have been able to attend Johns Hopkins University, let alone complete this thesis.
iii

Table of Contents
Title Page Page i
Abstract Page ii
Acknowledgments Page iii
Table of Contents Page iv
List of Tables Page v
List of Figures Page v
Introduction Pages 1-4
Materials and Methods Pages 5-8
Results Pages 9-16
Discussion Pages 17-20
Literature Cited Pages 21-25
iv

List of Tables
Table 1: Plasmids used in this study. Page 5
Table 2: Strains used in this study. Page 6
Table 3: Genes and corresponding sequences resulting from the Sac7 yeast two-hybrid screen.
Page 13
List of Figures
Figure 1: Confirmation of successful construction of pGBKT7 plasmids expressing Sac7-Gal4
DBD and Lrg1-Gal4 DBD chimeras. Page 10.
Figure 2: Growth suggesting interaction with Sac7 but not with Lrg1. Page 11
Figure 3: Slow grower growth suggesting interaction with Sac7 and often Lrg1. Page 12
Figure 4: Slow grower growth suggesting interaction with Sac7 and often Lrg1. Page 14
Figure 5: Difference between deletion strain and wild type strain were found to be negligible.
Page 16
v

Introduction
Endocytosis is an important evolutionarily conserved process necessary for nutrient
uptake, protein turnover at the membrane, and detection of external stimuli (Toret and Drubin,
2006; Prosser et al. 2011). Clathrin-mediated endocytosis (CME) is the best well characterized
mode of endocytosis in Saccharomyces cerevisiae (Boettner et al., 2011). In fact, until recently,
it was believed that CME was the only endocytic pathway in S. cerevisiae, the budding yeast,
despite the existence of clathrin-independent pathways, like caveolae mediated endocytosis,
micropinocytosis, and phagocytosis, in other organisms (Prosser et al., 2011). However, in yeast
cells lacking clathrin, endocytosis occurs, even if at severely reduced rates, suggesting that
Saccharomyces cerevisiae possessed some clathrin-independent method of performing this
indispensable function (Kaksonen et al., 2005; Newpher and Lemmon, 2006). This observation
contributed to the hypothesis that a clathrin-independent pathway exists in S. cerevisiae.
Clathrin-mediated endocytosis is not possible without cargo-specific adaptors at the
membrane which their cargo of interest and requires to the engage clathrin, cargos, and accessory
proteins. Endocytic adaptor proteins bind these distinct elements together to form a functional
unit. There are four main endocytic adaptor proteins required for endocytosis: the
AP180/PICALM homologues, Yap1801 and Yap1802; and the epsins, Ent1 and Ent2.
Simultaneous deletion of 3 endocytic adaptors does not significantly affect endocytosis, but
simultaneous deletion of all four endocytic adaptor proteins is lethal. However, it is not the
entirety of the endocytic adaptor proteins that is necessary (Maldonado-Baez et al., 2008).
Within Ent1 and Ent2 is an Epsin N-Terminal Homology (ENTH) domain, and within
Yap1801 and Yap1802 is an AP180 N-Terminal Homology (ANTH) domain. The ENTH and
ANTH domains are used to bind the plasma membrane, specifically phosphatidylinositol (4, 5)-
1

bisphosphate (Itoh et al., 2001). These domains regulate Cdc42, a protein important in cell
polarity. However, the ENTH and ANTH domains do not include other sites necessary for
clathrin-mediated endocytic function, like the clathrin-binding motif. Therefore, it was predicted
that, by deleting all four endocytic adaptors and adding in an ENTH domain, clathrin-mediated
endocytosis could be eliminated while retaining viability. Although the resulting yeast cells,
hereafter referred to as 4Δ cells, are temperature sensitive, they are viable and have severe
endocytic defects (Maldonado-Baez et al., 2008). These cells were critical in the discovery of a
clathrin-independent endocytic pathway (Prosser et al., 2011).
High-copy suppressor screens find proteins of interest by overexpressing randomized
proteins and then choosing those proteins that suppress a mutant phenotype. Using a high-copy
suppressor screen to test for growth at 37°C in 4Δ yeast cells, a clathrin-independent endocytic
pathway was discovered that relies on the small GTPase, Rho1 (Prosser et al., 2011). The Rho1
endocytic pathway does a relatively small amount of the total endocytosis performed; therefore,
visualization of the pathway only can be done when clathrin-mediated endocytosis is not
functioning. In the study that discovered the Rho1 endocytic pathway, cells with all four
endocytic adaptors deleted but with an ENTH1 domain added were used. Ste3-GFP was used as
the endocytic cargo to visualize this pathway (Prosser et al., 2011). Ste3 is a constitutively
endocytosed receptor expressed in alpha cells used to detect pheromones of the opposite mating
type (Chen and Davis, 2000). This strategy identified Mid2, Rom1, and Rho1 as important
components of this pathway. (Prosser et al. 2011)
However, Rho1, a Ras-like protein whose deletion is fatal for the cell, is involved in a
variety of functions. These functions include establishing cell polarity, regulating protein kinase
C (Qadota, 1996; Drgonova, 1996), regulating Fks1 for 1,3 β-glucan synthesis (Qadota, 1996),
2

and regulating Skn7 for transcription in response to oxidative stress (Helliwell, 1998). When
bound to GTP, Rho1 is active and can perform one or more of its myriad functions but becomes
inactive when the GTP is converted into GDP. In order to control the activity of Rho1, the cell
employs GTPase activating proteins (GAPs) to encourage GTP hydrolysis by Rho1 and guanine
nucleotide exchange factors (GEFs) to encourage the replacement of a GDP with a GTP. Thus,
GEFs promote activation of their cognate GTPases, while GAPs promote their inactivation.
As described above, Rho1 is involved in a large number of cellular processes. Distinct
mechanisms have evolved to use Rho1 in particular circumstances. Both Rho1 GAPs and Rho1
GEFs have been shown to regulate different aspects of Rho1’s function. As an example, the
Rho1 GAPs, Bem2 and Sac7, but not the Rho1 GAP, Bag7, downregulate the Pkc1-Mpk1
pathway (Schmidt et al., 2002). Similarly, Lrg1 is the only Rho1 GAP involved in 1,3- β-glucan
synthesis (Watanabe et al., 2001).
In its function as a Rho1 GAP, Sac7 is involved in Rho1-mediated endocytosis as a
negative regulator of the pathway (Prosser et al., 2011). It was discovered in a screen for proteins
whose mutation suppressed a temperature-sensitive act1 mutant, and was required for growth at
low temperatures (Dunn and Shortle, 1990). It was found later that the cell wall integrity (CWI)
pathway is downregulated by Sac7-mediated inactivation of Rho1 (Bickle et al., 1998; Lockshon
et al., 2012).
There is evidence that Rho1 and Sac7 functions are linked in cell cycle. This evidence
stems from the first study on Sac7, in which yeast with a Sac7 deletion exhibited a sporulation
defect. Sac7 is a negative of Rho1 in the TORC2 signaling pathways, which are important both
for actin organization and for eukaryotic cell growth (Ho et al., 2008; Pracheil et al., 2012). Sac7
is a negative regulator of Slt2, a protein important in cell cycle progression (Arias et al., 2011;
3

Martin et al. 2000). This is in agreement with the finding that SAC7 overexpression results in
slower growth (Akada et al., 1997). In addition, growth defects of mutants in the cell cycle
regulators Cdc34 and Cdc5 are worsened by Sac7 deletion (Varelas et al., 2006). The master
regulator of the meiotic and mitotic cell cycles, Cdk1, phosphorylates Rho1, Bni1, and Sac7
(Ubersax et al., 2003).
Study of the Rho1 endocytic pathway could have applications in humans. A homolog of
Rho1, RhoA, exists in humans, and can even rescue yeast lacking Rho1 (Qadota et al., 1994),
suggesting that RhoA has similar functions to Rho1 (Prosser, 2012). In addition, a RhoA
endocytic pathway exists in mammals (Ellis and Mellor, 2000; Khelfaoui et al., 2009; Kippert et
al., 2007; Wang et al., 2012). RhoA activation increases clathrin-independent endocytosis in
mammalian cells and is an integral participant in Type II phagocytosis (Ellis and Mellor, 2000).
This pathway is critical for oligodendrocyte function (Kippert et al., 2007), making predictable
the finding that inhibition of this pathway recently was proposed as a remedy for mental
retardation in a mouse model (Khelfaoui et al., 2009). Sac7 deletion also reduces the effect of
αS, the toxin responsible for the symptoms of Parkinson’s disease (Wang et al., 2012).
Despite potential applications for understanding Sac7’s role as a negative regulator of
Rho1-mediated endocytosis, Sac7 structure and function are poorly understood (Schmidt et al,
1997). The protein is 655 amino acids long and has only one predicted domain, the Rho1 GAP
domain, which is approximately 230 amino acids in length. Although Sac7 is involved in Rho1
downregulation in a variety of processes, the regulation of Sac7 has not been well characterized.
Physically interacting proteins of Sac7 are candidate regulators of Sac7, and therefore potential
regulators of the Rho1 endocytic pathway through this negative regulator. Therefore, a yeast
two-hybrid screen was performed to find physical interactors of Sac7.
4

Materials and Methods
Plasmid Construction
In this study, 4 plasmids were produced: pGBKT7-Sac7, pGBKT7-Lrg1, pGBKT7-Bag7,
and pACTII-Rho1. These were constructed using PCR to amplify each gene from the genomic
DNA. Enzymatic digestion at key sites in the base plasmids, pGBKT7 and pACTII, homologous
recombination, and growth on synthetic media selecting for successful homologous
recombination were used to construct the product plasmids. Correct construction was verified by
sequencing, enzymatic digestion, and Western blots. All plasmids used in this study are shown
in Table 1.
Plasmid Details Description Source pBW2053 2µ, URA3 pRS416::ENT2 Prosser et al., 2011 pBW0768 CEN, TRP1 pRS414::ENT1 Laboratory plasmid pBW0778 CEN, TRP1 pRS414::ENTH1 Laboratory plasmid pWB2053 2µ, URA3 YEp24::ROM1 Prosser et al., 2011 pBW2525 2µ, TRP1 pGBKT7.SAC7 This study pBW2526 2µ, LEU2 pGBKT7.LRG1 This study pGBKT7 2µ, TRP1 pGBKT7 Laboratory plasmid pACT2 2µ, LEU2 pACT2 Laboratory plasmid
Yeast Strains
For the yeast two-hybrid screen, the PJ69-4A strain was used. This strain is deficient in
its ability to produce uracil, tryptophan, leucine, histidine, and adenine. Activation at this strain’s
GAL4 promoter will induce the production of histidine and adenine. This was accomplished by
Table 1. Plasmids used in this study.
5

having GAL4 promoters integrated immediately upstream of the HIS3 and ADE2 coding
sequences. All strains used in this study are found in Table 2.
Strain Genotype Source
SEY6210 MATα/a, his3-Δ200 trp1-Δ901 leu2-3, 112 ura3-52 lys2-801 suc2-Δ9 BAR1 Laboratory Strain
BWY2858 MATa/α, Ste3-GFP::KANMX6 Prosser et al., 2010
BWY3399 Matα, ent1::LEU2, ent2::HIS3, yap1801::HIS3, yap1802::LEU2, pBW0768 Prosser et al., 2010
BWY3400 Matα, ent1::LEU2, ent2::HIS3, yap1801::HIS3, yap1802::LEU2, pBW0778 Prosser et al., 2010
BWY6570 MATα, YDL211C-GFP::KANMX6 This Study BWY6572 MATα, Ste3-GFP::KANMX6, ydl211c::NATMX4 This Study BWY6578 MATα, Ste3-GFP::KANMX6, tda7::NATMX4 This Study
BWY6584 MATα, ent1::LEU2, ent2::HIS3, yap1801::HIS3, yap1802::LEU2, Ste3-GFP::KANMX6, ydl211c::NATMX4, pWB0768 This Study
BWY6585 MATα, ent1::LEU2, ent2::HIS3, yap1801::HIS3, yap1802::LEU2, Ste3-GFP::KANMX6, ydl211c::NATMX4, pWB0778 This Study
BWY6586 MATα, ent1::LEU2, ent2::HIS3, yap1801::HIS3, yap1802::LEU2, STE3-GFP::KANMX6, tda7::NATMX4, pWB0768 This Study
BWY6587 MATα, ent1::LEU2, ent2::HIS3, yap1801::HIS3, yap1802::LEU2, STE3-GFP::KANMX6, tda7::NATMX4, pWB0778 This Study
PJ69-4A MATa, trp1-901, leu2-3,112 ura3-52, his3-200, gal4Δ, gal80Δ, lys2::GAL1-HIS3, GAL2-ADE3, met2::GAL7-lacZ James et al., 1996
Yeast Media
The medium used for growing yeast was always either rich medium (yeast peptone
dextrose [YPD]) or, in cases where a particular plasmid needed to be maintained in a strain,
synthetic medium (yeast nitrogen base [YNB]) lacking leucine, tryptophan, leucine, histidine, or
uracil.
Yeast Two-Hybrid Screen
The yeast strain PJ69-4A was transformed with pGBKT7-Sac7 along with plasmids from
the FRYL genomic library. The FRYL library consists of randomized pieces of yeast genomic
Table 2. Strains used in this study. All strains, excluding PJ69-4A, used in this study are isogenic to SEY6210 except at the indicated loci.
6

DNA of mean length 700 base pairs that produce protein chimeras consisting of the activation
domain of Gal4 combined with the protein resulting from the randomized 700 base pairs (Flores
et al. 1999; Fromont-Racine, M et al, 1997). Colonies able to grow on media lacking leucine,
tryptophan, leucine and histidine were selected, and plasmids from these colonies were isolated.
After retransformation into PJ69-4A with pGBKT7-Sac7 to confirm interaction by repeated
growth on selective media, these plasmids were sequenced. Sequences then were put into Base
Local Alignment Search Tools, or BLASTs, that matched these sequences with intervals from
the yeast genome. If a sequence matched a protein through both a nucleotide BLAST and a
protein BLAST, and was expressed as an in-frame fusion with the GAL4 activation domain, the
protein was declared a match.
Deletion and GFP-Tagging
Deletions and GFP-tagging of candidate Sac7 interacting proteins were accomplished
through genomic integration through PCR (Goldstein and McCusker, 1999; Longtine et al.
1998).
Microscopy and Image Analysis
An inverted microscope, the Axiovert 200 (Zeiss), and a 100x, 1.4 NA Plan-Apochromat
objective lens were used to obtain all images in this study. A Sensicam (Cooke) was used to take
pictures while an X-Cite 120 PC fluorescence illumination system was used to excite Ste3-GFP.
All images were taken with cells in YNB medium, and cells were prepared identically.
Testing for Interaction with other Rho1 GAPs
PJ69-4A cells were transformed with a pGBKT7 plasmid expressing the Rho1 GAP
whose interaction was to be tested. Cells containing the desired pGBKT7 plasmid then were
transformed with candidate interactors from the yeast 2 hybrid screen. Growth on selective
7

media indicated interaction. As a positive control, a pACTII-Rho1 plasmid was co-transformed
with both pGBKT7-Lrg1 and pGBKT7-Bag7.
8

Results
Over 1.3 million interactions were tested using the yeast two-hybrid screen with the
negative regulator, Sac7, as the bait. Figure 1 displays the confirmation of the successful
construction of the pGBKT7-Sac7 plasmid via plasmid sequencing and enzymatic digestion. A
Western Blot was used to show correct expression. In the initial screen, growth of 26 colonies
suggested interaction of prey proteins with Sac7. Of these, 12 colonies’ interaction with Sac7
could not be reproduced upon retransformation of the bait and prey plasmids; therefore, these
colonies were not pursued further. The plasmids of 14 colonies reproduced growth on selective
media after retransformation of the plasmids, which indicated interaction with Sac7. Of these 14,
5 colonies produced what will be referred to as a slow-grower phenotype. Figure 2 shows the
consistent growth of the 9 normal growing colonies and Figure 3 shows the growth of the slow-
grower colonies.
9

Figure 1. Confirmation of successful construction of pGBKT7 plasmids expressing Sac7-Gal4 and Lrg1-Gal4 DBD chimeras. (a) Constructed Sac7 and Lrg1 plasmids were digested using EcoR1, yielding a fragment of the correct size of a little more than 1.5 kilobases. (b) A Western Blot confirmed protein was being expressed at the correct size, 94 kiloDaltons. The primary antibody used was anti-Gal4 DNA Binding Domain. (c) Sequencing of the plasmids confirmed that both chimeras were being expressed.
10

Slow-grower colonies consistently show growth on YNB lacking tryptophan, leucine,
histidine, and adenine (YNB-4), which is indicative of protein interaction with Sac7 in a yeast
two-hybrid screen. However, these colonies grow much more slowly than normal growing
colonies. Growth on selective media varies among slow-grower colonies. However, an average
slow-grower colony produces as much growth in 2 weeks as a normal colony would produce in 2
days.
Plasmids from these colonies were isolated and sequenced. The results from this
sequencing are shown in Table 3. Colonies with normal growth produced sequences that
corresponded to proteins in the yeast genome; all slow-grower colonies produced sequences that
did not correspond to known proteins. The proteins implicated by the normal growers in this
screen to be physical interactors of Sac7 are: Ydl211c, a protein of unknown function that
Figure 2. Growth suggesting interaction with Sac7 but with Lrg1. (a) Diagram of streaking pattern used to test for Sac7 and Lrg1 interaction with proteins of interest (POI) (b) Growth after 2 days on conditions selecting for interaction. All four plasmids showing interaction with Ydl211c are included, each with a number corresponding to order of discovery.
11

localizes to the vacuole; Tda7, a cell-cycle regulated protein of unknown function and the
paralog of Ydl211c; Yat2, a carnitine acetyltransferase important in the G1/S transition; Zds2, a
regulatory protein involved in mitosis; Npr3, a component of the SEA, NPR 2/3, and Im1p
complexes; and Flc3, a putative FAD transporter.
In contrast to the strong effect that the Rho1 GAP Sac7 has on the Rho1-endocytic
pathway, the Rho1 GAP, Lrg1, essentially has no effect on this pathway, despite strongly
influencing Rho1 in other functions (Prosser, unpublished results). Therefore, Lrg1 interaction
was used as a negative control to tentatively test for proteins that are specifically involved in the
Rho1 endocytic pathway. Lrg1 was inserted into a pGBKT7 plasmid, and its correct insertion
was confirmed using enzymatic digestion and sequencing. Correct expression was supported by
a Western Blot, as is shown in Figure 1. None of the nine plasmids that yielded sequences that
corresponded to proteins in the yeast genome produced growth indicative of interaction with
Figure 3. Slow grower growth suggesting interaction with Sac7 and often Lrg1. (a) Diagram of streaking pattern used to test for Sac6 and LRg1 interaction with proteins of interest. (b) Growth after two weeks on conditions selecting for interaction.
12

Lrg1. However, of the five slow-grower colonies, four produced growth that indicated
interaction with Lrg1. Thus, the chimeras expressed by the colonies were likely promiscuous in
their physical interaction.
The interacting regions of the Sac7 physically interacting proteins were also identified.
Given the amino acid sequences that resulted from plasmid sequences, it was straightforward to
line these up with the known sequence in the genome. These results are shown in Table 3. NPR3,
had its predicted Sac7 binding region found in a predicted non-cytoplasmic region. Because Sac7
is a cytoplasmic protein, NPR3 was discarded as a potential regulator of the Rho1 endocytic
pathway through Sac7. Ydl211c, which was contained on four candidate plasmids, suggesting
physical interaction Sac7. All four sequences derived from these colonies overlapped with a
region begins with amino acid 330 and ends with the final amino acid at the C-terminal end.
The first four colonies’ plasmid sequences all corresponded to the protein Ydl211c.
Pictures taken of cells expressing GFP-tagged Ydl211c are shown in Figure 4. Previous studies
suggested that Ydl211c localizes to the vacuole (Huh et al., 2003). In my hands, Ydl211c
Table 3. Genes and corresponding sequences resulting from the yeast two hybrid screen.
13

localized to the cell surface, suggesting that it localizes both to the vacuole and to the cell
surface.
Among the proteins found in the screen were Ydl211c and its paralog, TDA7. These two
proteins are related to mucins, which are proteins involved in the production of gels, notably
mucus. Mucins often have a transmembrane domain, a highly glycosylated extracellular domain,
and a cytosolic domain. Both TDA7 and Ydl211c are predicted to have a transmembrane
segment, a cytosolic domain, and an extracellular domain, which one could predict to be
glycosylated based on the proteins’ relation to mucins.
Figure 4: GFP-Tagged Ydl211c suggest localization to the membrane and vacuoles. Saccharomyces cerevisiae cells were subjected to genomic integration via targeting of the endogenous YDL211C gene by which GFP was added to the C-terminal end of Ydl211c and visualized using a fluorescent microscope.
14

Neither TDA7 nor Ydl211c were shown to play a significant role in Rho1 mediated
endocytic activity, as is demonstrated in Figure 5. A strain lacking deletions to Sac7 interacting
proteins shows high endocytic activity for the WT + Vector and 4Δ + Ent1 + Vector cells, poor
endocytic activity for the 4Δ + ENTH1 + vector cells, and improved endocytosis for the 4Δ +
ENTH1 + Rom1 cells. If Tda7 or Ydl211c act as negative regulators of the pathway via Sac7, in
a strain with the protein of interest deleted, the 4Δ + ENTH1 + vector cells would show
improved endocytosis. Similarly, these proteins would be identified as positive regulators should
the 4Δ + ENTH1 + Rom1 cells show poor endocytic activity. However, as Figure 5 shows, this
was not the case.
As was expected, all samples of cells in the far right column, WT +Vector, showed strong
endocytosis, as is demonstrated in the small amount of Ste3-GFP found at the cytoplasmic
membrane. Similarly, the column second from right, showing the 4Δ + Ent1 + vector cells, has
universally strong endocytosis. This was expected because Ent1 restores Clathrin Mediated
Endocytosis. The third column is the first column in which endocytosis was dependent on the
Rho1 endocytic pathway. In all three backgrounds, WT, ydl211cΔ, and tda7Δ, endocytosis was
weak, with much of Ste3-GFP remaining at the membrane. The addition of Rom1 in the fourth
column rescued cells of all three backgrounds to the same comparatively strong endocytic state.
There was no appreciable difference between the three backgrounds, so this experiment failed to
show involvement by Ydl211c or Tda7 in the Rho1 endocytic pathway.
15
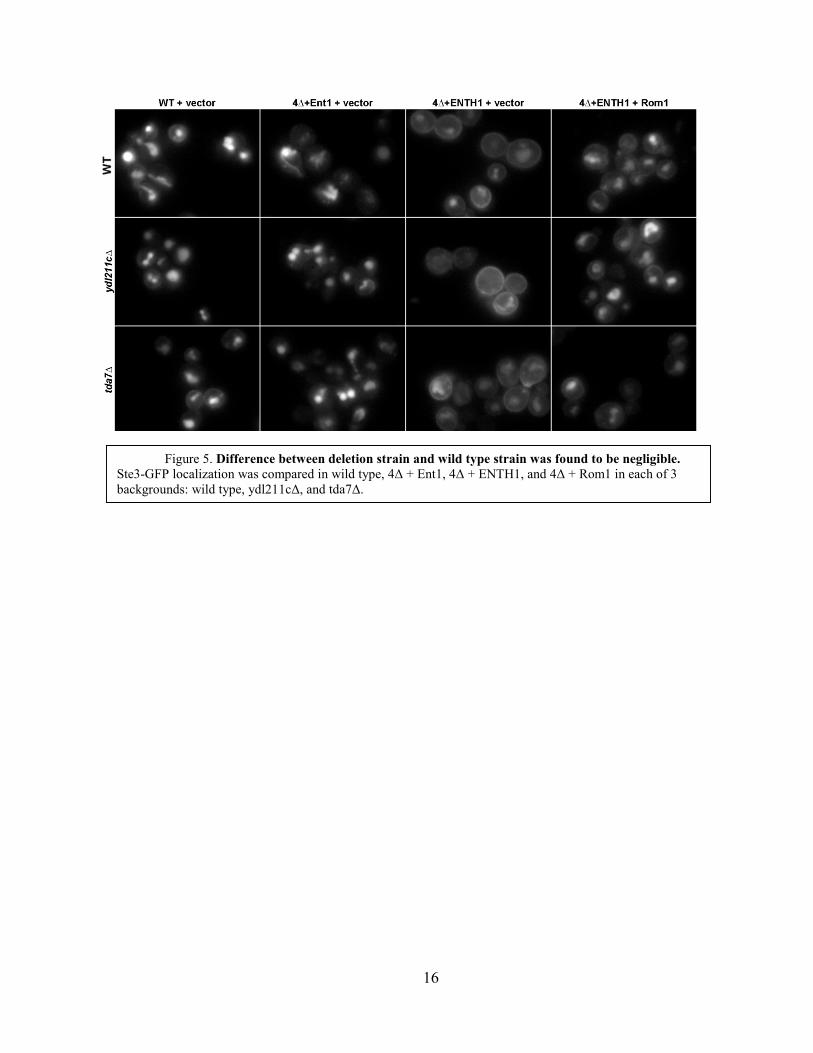
Figure 5. Difference between deletion strain and wild type strain was found to be negligible. Ste3-GFP localization was compared in wild type, 4Δ + Ent1, 4Δ + ENTH1, and 4Δ + Rom1 in each of 3 backgrounds: wild type, ydl211cΔ, and tda7Δ.
16

Discussion
Although slow-grower colonies initially were regarded as candidate Sac7 interactors, no
useful sequences were derived from them. Nine normal growing colonies and five slow-grower
colonies showed reproducible growth. All nine normal growing colonies yielded sequences that
corresponded to genes in the yeast genome and none of the five slow-grower colonies produced
such sequences. In addition, 4 of the 5 slow-grower colonies showed growth indicative of
interaction with Lrg1, suggesting that the interaction lacked protein/pathway specificity.
Therefore, we suggest that colonies of this slow-grower phenotype be disregarded in future yeast
two-hybrid screens.
TDA7 and YDL211C are paralogs, and their expression is regulated by the cell cycle.
Tda7 is known to physically interact with Rvs167, Lsb3, and Sla1, which are all involved in
clathrin-mediated endocytosis (Tonikian et al., 2009). Clathrin-mediated endocytosis consists of
three phases: the early immobile phase, the mid/late immobile phase, and the mobile phase
(Boettner et al., 2012). During the immobile phase, Ede1 and clathrin are recruited to the site of
endocytosis (Toret and Drubin, 2006). During the mid/late immobile phase, Las17p is recruited
to the site about 2 minutes after clathrin arrives, and recruits the Arp2/3 complex, which is
required for actin assembly (Boettner et al., 2012; Toret and Drubin, 2006). Notably, Lsb3, one
of the proteins that binds to Tda7, binds Las17. At the same time that Las17 is being recruited to
these sites, the Pan1 complex, which includes Sla1, a second protein known to bind with Tda7,
also is being recruited. Next, Vrp1 and Myo5 are recruited to the site and the process of actin
assembly begins. Internalization of the coat then begins, thus starting the mobile phase. The
forming vesicle must be excised from the membrane, and this is accomplished by Rvs161 and
Rvs167 (Boettner et al., 2012; Toret and Drubin, 2006), a third protein found to bind with Tda7.
17

Thus, Tda7 is known to bind with proteins from the early stages of the clathrin-mediated
endocytic pathway to the end stages. Because Sac7 is a negative regulator of the Rho1 endocytic
pathway, it is tentatively hypothesized that Tda7 plays a role in segregating of the two endocytic
processes. It is possible that Ydl211c, its paralog, could have a similar function.
Npr3 also was found to physically interact with Sac7. Npr3 acts as a part of three
different complexes: the Npr2/3 complex (Neklesa and Davis, 2009), the SEA complex
(Dokudovskaya et al., 2011), and the Iml1p complex (Wu and Tu, 2011). Interaction with Sac7
could, therefore, be relevant for any of these three complexes. The Npr2/3 complex mediates
cellular response to amino acid starvation through the TORC1 pathway (Neklesa and Davis,
2009). Amino acid starvation certainly could have effects on cellular decision making regarding
endocytic function and cell wall maintenance, both of which Sac7 negatively regulates. This
interaction could result in reduced Rho1-mediated endocytosis and less emphasis on cell wall
maintenance in response to amino acid starvation. The SEA complex associates with the vacuole
and is involved in intracellular trafficking (Dokudovskaya et al., 2011). It is possible that an
endosome produced by Rho1 endocytosis could use the SEA complex to attract Sac7 to itself and
thereby downregulate any remnant endocytic machinery on the endosome. The Iml1p complex
acts to regulate non-starvation induced autophagy in the cell (Wu and Tu, 2011). Sac7’s role in
the CWI pathway could be involved here. When the Iml1p complex is acting to induce
autophagy in the cell, actions of the CWI pathway upon the object of the autophagy would be
non-constructive. Therefore, it is conceivable that the Sac7 could act with the Iml1p complex to
downregulate the CWI pathway when autophagy is being performed.
Sac7’s interaction with Yat2 also could have biological significance. Yat2 is a carnitine
acetyltransferase that is found in the mitochondria (Swiegers et al., 2001). Carnitine
18

acetyltransferases are important for the transition from the S phase to the G1 phase in the cell
cycle (Brunner et al., 1997). During the cell cycle, the cell wall undergoes substantial
reconfiguration in order to produce the final product of two independent cells. Therefore, there
must be mechanisms to coordinate the CWI pathway with the progress of the cycle. Yat2’s
physical interaction with Sac7 could be one method of doing this. Lastly, a Yat2-Sac7 interaction
could stop the CWI or Rho1 endocytic pathways from occurring at the mitochondria, or it could
be unrelated to endocytosis.
Flc3 interacting physically with Sac7 potentially is meaningful as well. Flc3 is found in
the endoplasmic reticulum (Sickmann et al., 2003; Protchenko et al., 2006), where it acts as a
flavin adenine dinucleotide transporter (Protchenko et al., 2006) similar to its paralog Flc1
(Byrne and Wolfe, 2005). The Flc3-Sac7 physical interaction could simply help ensure that the
CWI and Rho1 endocytic pathways are not active at the ER. However, deletion of FLC genes
results in poor assembly of the cell wall, thus displaying a role of FLC3 in the cell wall
(Protchenko et al., 2006). Given that Flc3 acts to support cell wall assembly and Sac7 acts to
suppress the CWI pathway, the physical interaction between the two ensures that the processes
of cell wall assembly and the CWI pathway are segregated from each other and therefore do not
interfere with each other.
Of the 6 proteins derived from this screen, two of them, Tda7 and Ydl211c, are paralogs.
Therefore, paralogs of other proteins found in the Sac7 yeast two-hybrid screen may be strong
candidates for physical interaction with Sac7. Of the remaining 4 proteins, two, Flc3 and Zds2,
have paralogs, Flc1 and Zds1, respectively. Future tests for physical interaction with Sac7 would
be remiss to not include testing for the potential interaction between Flc1 and Zds1 with Sac7.
19

Bag7 is also a Rho1 GAP. It had been shown previously that Bag7 and Sac7 act together
in some functions, like cold-sensitivity suppression, but not in others. (Schmidt et al., 2002)
Additionally, Sac7 and Bag7 are paralogs, and could share other functions. Therefore, we predict
that some of the Sac7 interacting proteins identified in this study would be found to interact with
Bag7, so future studies of these proteins would be wise to include study on interaction with
Bag7.
Although 5 proteins were found in this study to interact with Sac7, only two, Ydl11c and
Tda7, were tested for involvement in the Rho1 endocytic pathway. Due to time constraints, the
potential for Flc3, Yat2, or Zds2 involvement in the pathway was not tested. Future studies
should investigate these three proteins as potential regulators of the pathway through Sac7.
The functions of the various domains of Sac7 are not well characterized. Although this
current study successfully found 6 proteins that physically interact with Sac7, characterization of
Sac7 did not determine which domains of Sac7 were binding these proteins. Therefore, future
work could concern which parts of Sac7 were involved in binding the proteins found in the
study. Because the only known domain of Sac7 is the Rho1 GAP domain (Schmidt et al, 2002),
preliminary study likely would break Sac7 into the pre-Rho1 GAP domain, the Rho1 GAP
domain, and the post Rho1 GAP domain. Future studies will continue to elucidate more about
the regulation of the Rho1 endocytic pathway, allowing for clearer understanding of endocytosis
in yeast, endocytic processes in general, and perhaps allowing for applications in higher order
animals, including humans.
20

Literature Cited
Akada R, Yamamoto J, Yamashita I. Screening and identification of yeast sequences
cause growth inhibition when overexpressed. Mol Gen Genet 1997;254:267–274. doi:
10.1007/s004380050415.
Arias P, et al. (2011) Genome-wide survey of yeast mutations leading to activation of the
yeast cell integrity MAPK pathway: novel insights into diverse MAPK outcomes. BMC
Genomics 12(1):390 PMID:21810245.
Bickle M, et al. (1998) Cell wall integrity modulates RHO1 activity via the exchange
factor ROM2. EMBO J 17(8):2235-2245 PMID:9545237.
Boettner DR, Chi RJ, Lemmon SK. Lessons from yeast for Clathrin-mediated
Endocytosis. Nature Cell Biology 14.1 (2011):2-10. Web.
Brunner S, Kramer K, Denhardt DT, Hofbauer R. Cloning and characterization of murine
carnitine Acetyl. Biochemistry Journal 322 (1997):403-410. Print.
Byrne KP, Wolfe KW (2005) The yeast gene order browser: combining curated
homology and synthetic context reveals gene fate in polyploidy species. Genome Res
15(10):1456-1461.
Chen L, Davis NG (2000) Recycling of the yeast a-factor receptor. J Cell Biol
151(3):731-738.
Dokudovskaya S, Waharte F, Schlessinger A, Pieper U, Devos DP, Cristea IM, Williams
R, Salamero J, Chait BT, Sali A, Field MC, Rout MP, Dargemont C. A conserved coatomer-
related complex containing Sec13 and Seh1 dynamically associates with the vacuole in
Saccharomyces cerevisiae. Mol Cell Proteomics 2011;10:M110.006478.
21

Drgonova J, et al. (1996) Rho1p, a yeast protein at the interface between cell polarization
and morphogenesis. Science 272(5259):277-279.
Dunn TM, Shortle D (1990) Null alleles of SAC7 suppress temperature-sensitive actin
mutations in Saccharomyces cerevisiae. Mol Cell Biol 10(5):2308-2314 PMID:2183030.
Ellis S, Mellor H. Regulation of endocytic traffic by rho family GTPases. Trends Cell
Biol 2000;10(3):85–88.
Fitch PG, Gammie AE, Lee DJ, de Candal VB, Rose MD. (2004) Lrg1p is a Rho1
GTPase-activating protein required for efficient cell fusion in yeast. Genetics 168:733–746.
Flores A, Briand JF, Gadal O, Andrau JC, Rubbi L, Van Mullem V, Boschiero C,
Goussot M, Marck C, Carles C, et al. A protein-protein interaction map of yeast RNA
polymerase III. Proc. Natl. Acad. Sci. U.S.A. 1999;96:7815–7820.
Fromont-Racine M, Rain JC, Legrain P. Toward a functional analysis of the yeast
genome through exhaustive two-hybrid screens. Nat Genet 1997;16:277–282.
Goldstein AL, McCusker JH. (1999). Three new dominant drug resistance cassettes for
gene disruption in Saccharomyces cerevisiae. Yeast. 15:1541–1553.
Helliwell SB, Schmidt A, Ohya Y, Hall MN. The Rho1 effector Pkc1, but not Bni1,
mediates signalling from Tor2 to the actin cytoskeleton. Current Biology 8.22 (1998):1211-S2.
Web.
Ho HL, et al. (2008) Involvement of Saccharomyces cerevisiae Avo3p/Tsc11p in
maintaining TOR complex 2 integrity and coupling to downstream signaling. Eukaryot Cell
7(8):1328-1343 PMID:18552287.
Huh WK, et al. (2003) Global analysis of protein localization in budding yeast. Nature
425(6959):686-691 PMID:14562095.
22

Itoh T, Koshiba S, Kigawa T, Kikuchi A, Yokoyama S, Takenawa T. (2001). Role of the
ENTH domain in phosphatidylinositol-4,5-bisphosphate binding and endocytosis. Science
291:1047–1051.
James, P., J. Halladay, and E.A. Craig. 1996. Genomic libraries and a host strain
designed for highly efficient two-hybrid selection in yeast. Genetics. 144:1425–1436.
Kaksonen M, Toret CP, Drubin DG. 2005. A modular design for the clathrin- and actin-
mediated endocytosis machinery. Cell 123:305–320.
Khelfaoui M, Alice P, Powell AD, Valnegri P, Cheong KW, et al. (2009) Inhibition of
RhoA pathway rescues the endocytosis defects in Oligophrenin1 mouse model of mental
retardation. Human Molecular Genetics 18:2575–2583.
Kippert A, Trajkovic K, Rajendran L, Ries J, Simons M. (2007). Rho regulates
membrane transport in the endocytic pathway to control plasma membrane specialization in
oligodendroglial cells. J Neurosci 27:3560–3570.
Lockshon D, et al. (2012) Rho signaling participates in membrane fluidity homeostasis.
PLoS One 7(10):e45049.
Longtine MS, McKenzie III A, Demarini DJ, Shah NG, Wach A, Brachat A, Philippsen
P, Pringle JR. (1998) Additional modules for versatile and economical PCR-based gene deletion
and modification in Saccharomyces cerevisiae. Yeast 14:953–961.
Lorberg A, et al. (2001) Lrg1p functions as a putative GTPase-activating protein in the
Pkc1p-mediated cell integrity pathway in Saccharomyces cerevisiae. Mol Genet Genomics
266(3):514-526.
23

Maldonado-Baez L, Dores M R, Perkins EM, Drivas TG, Hicke L, et al. (2008)
Interaction between Epsin/Yap180 adaptors and the scaffolds Ede1/Pan1 is required for
endocytosis. Mol Biol Cell 19:2936–2948.
Martin H, Rodriguez-Pachon JM, Ruiz C, Nombela C, Molina M. Regulatory
mechanisms for modulation of signaling through the cell integrity Slt2-mediated pathway in
Saccharomyces cerevisiae. J Biol Chem 2000;275:1511-1519.
Neklesa TK, Davis RW. (2009) A genome-wide screen for regulators of TORC1 in
response to amino acid starvation reveals a conserved Npr2/3 complex. PLoS Genet 5, e1000515.
Newpher TM, Lemmon SK. (2006). Clathrin is important for normal actin dynamics and
progression of Sla2p-containing patches during endocytosis in yeast. Traffic 7:574–588.
Pracheil T, et al. (2012) TORC2 signaling is antagonized by protein phosphatase 2A and
the far complex in Saccharomyces cerevisiae. Genetics 190(4):1325-1339 PMID:22298706.
Prosser DC, Drivas TG, Maldonado-Baez L, Wendland B. (2011). Existence of a novel
clathrin-independent endocytic pathway in yeast that depends on Rho1 and formin. J Cell Biol
195, 657–671. 10.1083/jcb.201104045.
Prosser DC, Wendland B. 2012. Conserved roles for yeast Rho1 and mammalian RhoA
GTPases in clathrin-independent endocytosis. Small GTPases. 3:229–235Prosser unpublished
work
Protchenko O, et al. (2006) A screen of heme uptake identifies the FLC family required
for import of FAD into the endoplasmic reticulum. Journal of Biological Chemistry
281(30):21445-21457.
Qadota H, Anraku Y, Botstein D, Ohya Y. Conditional lethality of a yeast strain
expressing human RHOA in place of RHO1. Proc Natl Acad Sci U S A. 1994;91:9317–9321.
24

Qadota H, et al. (1996) Identification of yeast Rho1p GTPase as a regulatory subunit of
1,3-beta-glucan synthase. Science 272(5259):279-281.
Schmidt A, et al. (1997) The yeast phosphatidylinositol kinase homolog TOR2 activates
RHO1 and RHO2 via the exchange factor ROM2. Cell 88(4):531-542 PMID:9038344.
Sickmann A, et al. (2003) The proteome of Saccharomyces cerevisiae mitochondria. Proc
Natl Acad Sci U S A., 100 (23):13207-13212.
Swiegers JH, et al. (2001) Carnitine-dependent metabolic activities in Saccharomyces
cerevisiae: three carnitine acetyltransferases are essential in a carnitine-dependent strain. Yeast
18(7):585-595.
Ubersax JA, Woodbury EL, Quang PN, Paraz M, Blethrow JD, Shah K, Shokat KM,
Morgan DO. Targets of the cyclin-dependent kinase Cdk1. Nature. 2003;425:859–864.
Tonikian R, et al. (2009) Bayesian modeling of the yeast SH3 domain interactome
predicts spatiotemporal dynamics of endocytosis proteins. PLoS Biol 7(10).
Toret, CP, Drubin DG. The budding yeast endocytic pathway. Journal of Cell Science
119.22 (2006):4585-4587. Web.
Watanabe D, et al. (2001) Yeast Lrg1p acts as a specialized RhoGAP regulating 1,3-beta-
glucan synthesis. Yeast 18(10):943-51.
Wu X, Benjamin PT. Selective regulation of autophagy by the Iml1-Npr2-Npr3 Complex
in the absence of nitrogen starvation. Molecular Biology of the Cell 1 Nov. 2011:4124-4133.
Print.
25

Curriculum Vitae
Matthew Richard Peck
3027 Decatur Street, West Lafayette, Indiana 47906
[email protected]; (765) 714-9806
EDUCATION
• Honors Core 40 Diploma with Academic Honors, West Lafayette High School, West Lafayette, Indiana, 2011. GPA: 4.0
• Bachelor of Science, Arts and Sciences, Biology, The Johns Hopkins University, Baltimore, Maryland, 2014 GPA: 3.7
• Master of Science, Arts and Sciences, Biology, The Johns Hopkins University, Baltimore, Maryland, 2015
AWARDS
• Dean’s List: Fall 2011, Spring 2012, Fall 2012, Spring 2013, Fall 2014 • Beta Beta Beta Honor Society, Associate Member and President: April 2013
RESEARCH EXPERIENCE
• Research Assistant, Wendland Lab, Department of Biology, The Johns Hopkins University, Baltimore, Maryland. August 2012 to present.
POSTER PRESENTATIONS • Peck Matthew, Prosser Derek, Wendland Beverly. Preliminary Characterization
of Sac7 as a Negative Regulator of the Rho1-mediated Clathrin Independent Endocytic Pathway in Saccharomyces cerevisiae. Poster presented at Beta Beta Beta poster session, The Johns Hopkins University, April 2014, Baltimore, Maryland.
26










![Onconase cytotoxicity relies on the distribution of its ...raineslab.com/sites/default/files/labs/raines/pdfs/Turcotte2009.pdfdependent endocytic pathway [50]. The cytotoxicity of](https://img.dokumen.tips/doc/110x75/608cfcd816e42b72ba23352c/onconase-cytotoxicity-relies-on-the-distribution-of-its-dependent-endocytic.jpg)


















