Embed Size (px)
Citation preview

Characterization of Differentially Expressed Genes following BriefCardiac Ischemia
Ralph Knöll, René Zimmermann, Margarete Arras, and Wolfgang Schaper1
Department of Experimental Cardiology, Max-Planck-Institut, Benekestr. 2, 61231 Bad Nauheim, Germany
Received March 7, 1996
For the study of ischemia-related adaptations we employed DDRT-PCR on mRNA obtained from heartregions undergoing brief (109) occlusions/reperfusions. We found and sequenced 52 differentially expressedclones that we further characterized using Northern analysis with RNA from a wide variety of tissues. Weselected only clones (1) with a new sequence, (2) that showed a Northern signal, and (3) that exhibitedorgan-specific (heart, muscle) or (4) situation-specific (ischemia, non-specific stress) expression. We found twonew heart-specific transcripts and three new stress-inducible genes. The transcripts with known sequencesshowed an expression pattern typical for repair processes after ischemia-reperfusion (ubiquitin, ATPases). About20% of clones were truly differentially expressed.© 1996 Academic Press, Inc.
Brief periods of myocardial ischemia followed by reperfusion induce adaptational processes thatresult in delay of cell death for hours (ischemic preconditioning) and weeks and months (hiber-nating myocardium) (11). We have previously shown that brief periods of ischemia are stronginducers of gene expression (1,3,5,7) but a link between increased transcription of a mRNA speciesand adaptational changes was so far not established on the basis of traditional methods of cDNA-RNA hybridization. DDRT-PCR has the advantage that it is free from the bias of probe selectionand that a relatively general working hypothesis (influence of transient ischemia, molecular basisof adaptation) can be tested with a high probability for the detection of new genes.
MATERIAL AND METHODS
Animal preparation.Animals used in this study were handled in accordance with the American Physiological Societyguidelines for animal welfare and the experimental protocol was approved by the Bioethical Committee of the District ofDarmstadt, Federal Republic of Germany. Twenty castrated male mixed breed Landrace-type domestic pigs, weighingbetween 21 and 39 kg, were sedated with azaperone 2 mg/kg i.m. and then anesthetized with pentobarbital 30 mg/kg i.v.After tracheal intubation, the thorax was opened through midline thoracotomy and the left anterior descending coronaryartery (LAD) was occluded for 10 minutes followed by 30 minutes of reperfusion and then occluded for a further 10 minutes(4timepoint 10-30-10). Pigs were sacrificed immediately after the second occlusion, or after 30 and 90 minutes ofreperfusion (4timepoints: (10-30)2 or (10-30)2+60 (see Fig. 1)). Sham animals were sacrificed without occlusion. Experi-mental tissue was removed from the LAD area and control tissue was removed from the region of the left circumflexcoronary artery (Cx) area (1,5).RNA isolation and Northern blotting.Total RNA was isolated by the method of (4). and Northern blots were performed
as described in (7).Reverse transcription and differential display.0.2mg of total RNA, in the presence of 6.25mM of one downstream primer
(4dT11XY, X4ACG, Y4ACGT, twelve different primers), was denatured in 8ml of DEPC water for 10 minutes at 70°Cand placed on ice. 4ml of dNTPs (100mM), 4ml of 5X reverse transcriptase buffer, 2ml of 0.1M DDT and 0.5ml of RNasinwere added. This mix was stored at roomtemperature for 3 minutes before 1.5ml of reverse transcriptase (300U) were added.The reactionmix again was stored at roomtemperature for 8 minutes and incubated at 60°C for 35 minutes. The reaction wasstopped by heating the tube for 5 minutes at 95°C and by the addition of 10ml of ddH2O. The resulted first strand cDNAwas immediately used for PCR or stored at −80°C. For PCR amplification usually mastermixes were prepared and each PCRamplification (20ml final reaction volume) was done in the presence of: 1ml of first strand cDNA, 2.5mM of one downstreamprimer, 0.5mM of one upstream primer (26 different upstream and 12 different downstream primers, described in (2)),10mM Tris/HCl pH8.3, 50mM KCl, 1.5mM of MgCl2, 2mM of dNTPs, 1ml of 10mCi/ml (a-35S) dATP (>1000mCi/mMol
1 Fax: 49 6032 705-419.
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS221,402–407 (1996)ARTICLE NO. 0608
4020006-291X/96 $18.00Copyright © 1996 by Academic Press, Inc.All rights of reproduction in any form reserved.

Amersham), 0.2ml of Taq DNA Polymerase (1U). PCR conditions were as follows: denaturation for 2 minutes at 94°C,followed by 40 cycles of: 30 seconds 94°C, 60 seconds 40°C, 30 seconds 72°C, 72°C for 5 minutesThe resulting cDNAs were stored at 4° or 3.5ml of PCR product were immediately mixed with 2ml of formamide loading
buffer (495% formamide, 0.09% bromophenol blue, 0.09% xylene cyanol), gelelectrophoresed on a 6% native polyacryl-amide sequencing gel, transferred onto 3 MM paper and autoradiographed. Those bands differing in their expressionbetween the nonischemic and ischemic area were excised and reamplified in 50ml reactions using the same conditions asbefore, except downstream and upstream primer concentration was 2.5mM and dNTP concentration was 100mM. Theresulting fragments were polished withPfu DNA polymerase, cloned into a pCR-Script™ SK(+) cloning vector andsequenced by the use of a ALF automatic sequencer. The sequences were analyzed using the EMBL gene library and testedfor expression by Northern hybridization.Statistical analysis.Significant differences between ischemic (4I) and nonischemic (4N) were obtained by normalizing
the emission data obtained by the phosphorimager [(I-N)/N] and then comparing the expression tcrit 4 mean/SEM with theStudent’s t-distribution. Statistical significance was accepted at p<0.05.
RESULTS
In our study in postischemic hearts we have counted about 4000 identifiable mRNA species(with z10% of all the possible primer combinations) of which about 60 (z1.5%) appeared to bedifferentially expressed. Of these we have sequenced 52. Sixteen of these gene fragments producedhybridization signals in Northern blots, in 6 the sequence showed strong homology to known genes(and 46 sequences were, more or less, unknown): 2 of the gene fragments with known sequences( >70% sequence homology over 200 base pairs) belong to enzymes of the mitochondrial energymetabolism (mitochondrial creatine kinase, ubiquinone), one was a heat shock protein, one wasmyosin, one was G-rich sequence factor-1 and one ab-DNA polymerase. Among the fragmentswithin the bracket of 55 to 70% sequence homology were: two fragments with distinct sequencehomologies to enzymes of the energy metabolism, two to the calcium binding protein genes,apoptosis protein RP-8, FK506 or rapamycin binding protein, 28S ribosomal protein, transferrinreceptor mRNA and retinoblastoma susceptibility gene (see also table 1). The sequenced cDNAswere usually from the 39 untranslated region and their average length was between 200 and 500bp.This limited sequence information might be the reason that some genes could not be identified(assuming the presence in the GenBank computer of cDNAs with unsequenced 39 ends), thereforewe assume that 30% of the genes are known but not identified and probably 50% are reallyunknown, which was also suggested by (8). An important confirmatory finding was the identifi-cation of ubiquitin, known as a member of the stress inducible genes and previously reported byus to be induced following ischemia and reperfusion (1). This result indicates clearly that theapplied technique is useful in order to detect stress/ischemia inducible genes. Northern hybridiza-tion with the cloned cDNAs against RNA from normal and postischemic hearts showed that onlya minority (z20%) exhibited true differential expression. Two of the 3 differentially expressedgenes represent unknown sequences. Since the mRNA levels of gene RKCU12.2 were found to be6 times higher at 10-30-10 (p<0.05) in the control as well as in the ischemic area in comparisonto the values obtained from the control animals as well as compared to the values from the animals
FIG. 1. Our ischemia protocol. The LAD was occluded for 10 minutes, reperfused for 30 minutes and again occludedfor 10 minutes. The heart was excised (4HE) immediately after the second occlusion, following 30 or 60 minutes ofreperfusion.
Vol. 221, No. 2, 1996 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
403

TABLE 1
Clone S.bp Closest homologous sequence (match>70% in 200 bp, 6 gaps)
RK3.1 340 Human (87% in 280bp), rat (80% in 287 bp), chicken (71% in 205bp)mitochondrial creatine kinase (N)
RKDU1E1.6 210 B.taurus mRNA for ubiquinone oxidoreductase complex (85% in 219bp)(N)
RKDU2E2.1 270 Human (97% inz270bp), rat (84% inz270bp) b-DNA polymerase mRNARKD3.4 470 Human G-rich sequence factor-1 (GRSF-1) mRNA (79% in 379bp)RKD4.1 400 z80% homology to human, rat, mouse, bovine ubiquitin mRNA, 39 end (N)RKD8.2 400 z80% homology to human, rat, mouse myosin mRNA, 39 end (N)
Clone S.bp Closest homologous sequence (match <70% in 200 bp, 6 gaps)
RK4.9 260 Human ATP synthase (52% in 188bp)RK8.5 140 Rat transferrin receptor mRNA 39 end (60% in 72bp)RK9.2 280 Homo sapiens cDNA clone HHCPR 62, hippocampus (64% in 274bp)RK10.2 350 Human retinoblastoma susceptibility gene (53% in 248bp)RKU12C1 400 “Apoptosis” protein RP-8 (55% in 161bp)RKU15E1 400 Human profilaggrin gene, calcium binding protein (48% in 216bp)RKCU12310 210 Rat (72% in 120 bp), human (68% in 120 bp) acyl CoA synthetase, long
chain (N)RKDU1E1.1 330 Unknown sequence, (dictyostelium discoideum myosin heavy chain kinase
48,2% in 224bp) (N)RKCU1342 400 Rat (86% in 169bp), human (83% in 175bp) ribosomal protein S28 (N)RKDU1E21 210 Human (54% in 161bp) fragile X mental retardation protein (FMR-1) gene;
A. nidulans (69% in 157bp) nuclear movement protein gene (N)RKDU1E2.2 400 Unknown (N)RKCU12.2 500 Unknown (N)RKD42.2 280 Pig SR Ca2+ pump gene 2–39 end (73% in 175bp) (N)RKD44.8 400 Unknown (N)RKD46.5 200 Unknown (N)RKD5.1.1 300 Cow (67% in 178 bp), human (66% in 178 bp) 25 kD FK506 or rapamycin
binding protein mRNA (N)RKD7.1 210 Unknown (N)
Table 1 shows a list of some genes so far cloned and sequenced. About 30 unknown genes, not detectable by Northernblotting, were not included. (N)4 this gene is detectable by Northern blotting. S.bp4 sequenced basepairs. Percentagevalues in the clamp indicates highest sequence homologies to known genes. All inserts are in the range of about 200–500 bp.
FIG. 2. The timecourse of the induction of gene RKCU12.2, with a significant induction at 10-30-10 (p<0.05).
Vol. 221, No. 2, 1996 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
404
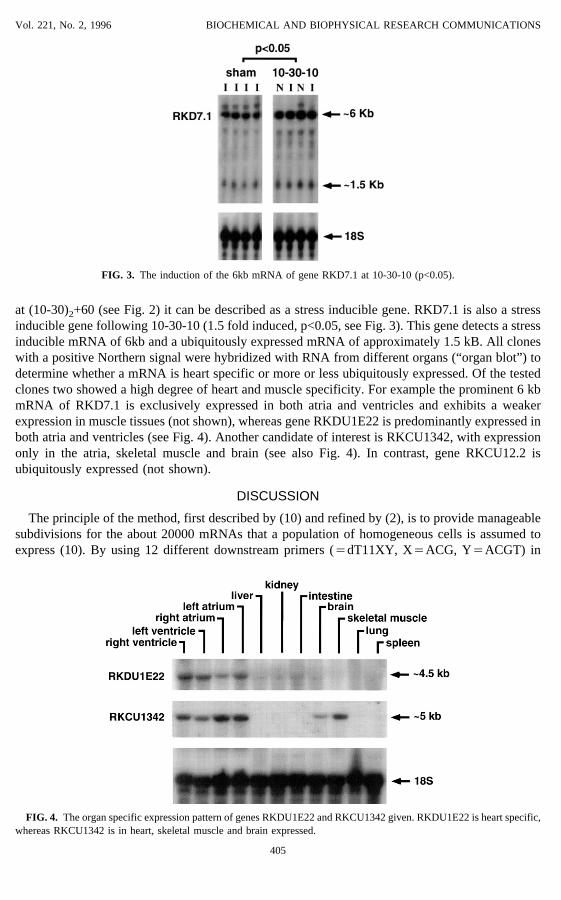
at (10-30)2+60 (see Fig. 2) it can be described as a stress inducible gene. RKD7.1 is also a stressinducible gene following 10-30-10 (1.5 fold induced, p<0.05, see Fig. 3). This gene detects a stressinducible mRNA of 6kb and a ubiquitously expressed mRNA of approximately 1.5 kB. All cloneswith a positive Northern signal were hybridized with RNA from different organs (“organ blot”) todetermine whether a mRNA is heart specific or more or less ubiquitously expressed. Of the testedclones two showed a high degree of heart and muscle specificity. For example the prominent 6 kbmRNA of RKD7.1 is exclusively expressed in both atria and ventricles and exhibits a weakerexpression in muscle tissues (not shown), whereas gene RKDU1E22 is predominantly expressed inboth atria and ventricles (see Fig. 4). Another candidate of interest is RKCU1342, with expressiononly in the atria, skeletal muscle and brain (see also Fig. 4). In contrast, gene RKCU12.2 isubiquitously expressed (not shown).
DISCUSSION
The principle of the method, first described by (10) and refined by (2), is to provide manageablesubdivisions for the about 20000 mRNAs that a population of homogeneous cells is assumed toexpress (10). By using 12 different downstream primers (4dT11XY, X4ACG, Y4ACGT) in
FIG. 3. The induction of the 6kb mRNA of gene RKD7.1 at 10-30-10 (p<0.05).
FIG. 4. The organ specific expression pattern of genes RKDU1E22 and RKCU1342 given. RKDU1E22 is heart specific,whereas RKCU1342 is in heart, skeletal muscle and brain expressed.
Vol. 221, No. 2, 1996 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
405

combination with 26 different upstream oligodecamers the expected mRNA population is dividedinto about 312 groups with about 64 different mRNA species per group. Since one primer pairamplifies more than this number of fragments, we and others (9) obtained about 100 different bandsper lane, which is in good agreement with the theoretical considerations and the results obtainedby (2). An advantage of the method is that RNA from a treated population can be visuallycontrasted on the sequencing gel with that from control tissue and allows therefore selection intotwo directions. To minimize the possibility of false positive clones, each PCR reaction wasperformed in duplicate and both PCR reactions (two control as well as two experimental reactions)were electrophoresed in neighbouring lanes. Specific bands were cut out only when they showeddifferential expression in both lanes. In order to be sure that clone selection was not random, wecompared the sequences of presumed differentially expressed clones with those picked at randomfrom a cDNA library constructed from normal pig heart (Dreier and Zimmermann, unpublished).Comparison with these sequences showed a totally different distribution: the most common se-quences in the library are desmin, titin and myosin which did not appear at all in the differentialdisplay (except one myosin-clone) which indicates that a non-random selection must have beenachieved by the experimental intervention. Most of the fragments differing in expression betweennonischemic and ischemic/reperfused areas could be reamplified and cloned. For practical purposesand since genes with abundant mRNAs might have an important function, only those cDNAs withpositive signals by Northern hybridization underwent further examination. However, only about20% of the clones positive by Northern analysis were truly differentially expressed. Several reasonsmight be responsible for this phenomenon. One reason could be that the identified cDNAs belongto a differentially spliced transcript, probably differentially expressed, but not detectable by North-ern hybridization. The main gene product of those genes may not be differentially expressed.Another reason could be that some cDNAs belong to a gene family and one fragment mayrecognize by Northern blotting several other family-members, probably not differentially ex-pressed. Some fragments showed a tendency to be differentially expressed, but this was notstatistically significant. Another fact is that some cDNAs are likely to be ischemia inducible, butthey may be even more sensitive to the surgical and traumatic stress that the animals undergoduring the experiments despite deep anesthesia (for example RKCU12.2, RKD7.1), a feature thatwe have earlier described for IGF-2(6). The single most important source of error may be the PCRmethod itself which after 40 cycles amplifies small erros out of proportion and may lead to falsepositives. Another problem is the fact that sometimes one differentially expressed band consists oftwo or more cDNAs, as verified by reamplification. Only one of them might exhibit true differ-ential expression. Other authors have also struggled with the problem of false positives and haveadopted other technical solutions. Using higher hybridization temperatures and “hot start” modi-fications the percentage of false positives was reduced to 50% but the overall number of expressedclones decreased significantly, i.e., increased specificity was only reached at the expense of sen-sitivity. We conclude from our experience with DDRT-PCR that its strength is its potential for thediscovery of previously unidentified genes, tissue specific (ie heart) and situation-specific (ie,ischemia). Our success in identifying four new genes with potential functional and structural rolestestifies to that. In this aspect DDRT-PCR is superior to other differential strategies of molecularcloning. One word of caution seems appropriate with regard to the word usage “gene expression”:we have shown earlier that in the heart changes in the tissue mRNA concentration are not alwayscaused by changes in transcription. mRNA stability changes in a significant number of genes inpostischemic heart. Ischemia, because of its inhibitory influence on translation, may thereforeincrease mRNA transitorily. Ischemia can act like cycloheximide and can hence “superinduce”certain genes. We have avoided this potential source of error by studying mRNA only followingshort term ischemia. We found that episodes of brief coronary occlusion-reperfusion studied withDDRT-PCR increased the expression of a surprisingly high number of heretofore unidentified
Vol. 221, No. 2, 1996 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
406

genes (z80%). Further studies showed that four of these were of interest because Northernanalysis, in particular the organ distribution of RNA, revealed that they were either heart or musclespecific (2) or were truly situation-specific (overexpressed only during ischemia), or showed abehavior like immediate early genes, or reacted to the general trauma of the experimental condi-tions like stress genes. The early and high expression of genes RKCU12.2 and RKD7.1 followingischemia shows kinetics similar to that observed by immediate early genes and they may havesomething to do with early ischemic reactions like preconditioning. In addition to the identifiedgenes we obtained interesting organspecific expression patterns, the 6kb mRNA of gene RKD7.1and gene RKDU1E22 is heart specific. In addition gene RKCU1342 is expressed in muscle as wellas in brain tissue. Thus far we do not know whether the 6kb mRNA, only observed in muscletissues, and the 1.5kb mRNA were transcribed only from one gene and therefore may representdifferentially spliced variants or whether they correspond to two different genes. In contrastRKCU12.2 is more or less ubiquitously expressed, except in kidney and liver. Therefore it seemsto function not only in terminally differentiated but also in proliferating cells and tissues.We are currently in the process of doing 59 RACE and screening a heart derived cDNA library
in order to get the full length clones for RKD7.1 and RKCU12.2 as well as for genes withpredominant expression in the heart.
ACKNOWLEDGMENTS
The first author thanks Mrs. A. Möbs and Mr. G. Stämmler for their help in photographic and computer work, respec-tively. Dr. B. Winkler is acknowledged for the computer sequence search.
REFERENCES
1. Andres, J., Sharma, H., Knöll, R., Stahl, J., Sassen, L. M. A., Verdouw, P., and Schaper, W. (1993)Cardiovasc Res27,1421–1429.
2. Bauer, D., Müller, H., Reich, J., Riedel, H., Ahrenkiel, V., Warthoe, P., and Strauss, M. (1993)Nucleic Acids Res21,4272–4280.
3. Brand, T., Sharma, H., Fleischmann, K., Duncker, D. J., McFalls, E. O., Verdouw, P. D., and Schaper, W. (1992)CircRes71, 1351–1360.
4. Chomzynski, P., and Sacchi, N. (1987)Anal Biochem162,156–159.5. Frass, O., Sharma, H. S., Knöll, R., Duncker, D. J., McFalls, E. O., Verdouw, P. D., and Schaper, W. (1993)Cardiovasc
Res27, 2037–2043.6. Kluge, A., Zimmermann, R., Münkel, B., Verdouw, P., Schaper, J., and Schaper, W. (1995)Cardiovasc Res29,
708–716.7. Knöll, R., Arras, M., Zimmermann, R., Schaper, J., and Schaper, W. (1994)Cardiovasc Res28, 1062–1069.8. Kozian, H. K., and Augustin, H. G. (1995)Biochem Biophys Res Commun209,1068–1075.9. Liang, P., Averboukh, L., and Pardee, A. B. (1993)Nucleic Acids Res21, 3269–3275.10. Liang, P., and Pardee, A. B. (1992)Science257,967–971.11. Murry, C. E., Richard, V., Reimer, K. A., and Jennings, R. B. (1990)Circ Res66, 913–931.
Vol. 221, No. 2, 1996 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
407





























