Embed Size (px)
Citation preview

Eur. J. Biochem. 103, 21-27 (1980)
Characterization of Bovine Anterior-Lens-Capsule Basement-Membrane Collagen 1. Pepsin Susceptibility, Salt Precipitation and Thermal Gelation a Property of Non-collagen Component Integrity
David SCHWARTZ, Trevor CHIN-QUEE, and Arthur VElS
Department of Oral Biology, Northwestern University Dental School
(Received July 9, 1979)
Basement membrane collagen was extracted sequentially from bovine anterior lens capsules by repeated pepsinization at very low enzyme-to-collagen ratios. The earliest extracts, derived from 1 - 7-days pepsinization at 4 "C showed that a substantial fraction of basement membrane collagen material did not enter a 6 % or 3 % sodium dodecylsulfate/polyacrylamide gel. The material that did penetrate the gel ran as discrete bands with molecular weights between 220000 and 140000. The relative proportions of these components could be controlled solely by the duration of pepsinization. Furthermore, as pepsinization was continued or as the quantity of pepsin or the extraction tempera- ture was increased basement membrane collagen peptides with molecular weights between 11 5 000 and 30000 were produced, with little material not entering the gel. The extracts containing collagen chains with M, > 130000 were precipitable from acetic acid solution at NaCl concentrations of 0.3-0.6 M and could undergo a thermal gelation at pH 7.03. At or near the transition to the lower- molecular-weight species (M, < 130000), the precipitability at low NaCl concentrations and thermal gelation properties were lost. Correlation of these properties with amino acid composition and with previous electron microscopic studies suggest that strong attractive interactions between pepsin- susceptible molecular end regions are responsible for the salt precipitability and gelation. These end region interactions may also play a role in inhibiting D-periodic striated fibril formation in basement membrane structures in vivo.
Basement membranes are unique connective tissues in that the collagen within them is not organized into striated filaments, although basement membranes such as the anterior lens capsule may have some organized filamentous character [1]. In previous reports [2,3] we have demonstrated that basement membrane col- lagen molecules from intact or minimally degraded lens capsule preparations are in the form of procolla- gen-like molecules where the individual chains have M, of about 175000 and, similar to type I procollagen, have extensions at both ends of the triple-helical sec- tion of the molecule. The distribution of cross-stria- lions seen in segment-long-spacing crystallites of base- ment membrane collagen is different from that of type I. This raised the possibility [1,2] that it was the different arrangement of potential side-chain inter- acting groups that might prevent the formation of
Enzymes. Pepsin (EC 3.4.23.1); collagenase (EC 3.4.24.3).
typical 70-nm D-periodic fibrils. We also noted, how- ever, that the collagen molecules with end regions intact were highly aggregated, even after reduction and alkylation. Thus, an alternative basis for the difficulty in periodic fibril assembly might be the preferential formation of end-interacted filaments or networks.
The objective of this study was to determine the nature and aggregative properties of the least degraded basement membrane collagen molecules that can be extracted from a formed tissue, and to assess the effect of partial or controlled proteolysis on these properties. Trelstad and Lawley [4] have used the thermal gela- tion of collagen in vitro to separate some basement membrane collagens from other types. They have shown that pepsinized basement membrane collagens remain soluble during concomitant thermal gelation in vitro of type I collagen. However, Comper and Veis [5,6] have shown that, with type I collagen, proteolytic removal of the molecular end regions markedly de-

22 Basement-Membrane Collagen: Pepsinization, Precipitation, Heat Gelation
creases the rate at which thermal gelation will take place, a finding supported by the recent work of Wil- liams et al. [7]. Thus the possibility existed that mini- mally degraded basement membrane collagen might also form thermal gels and we have, therefore, ex- amined the thermal gelation of anterior lens capsule collagen and its salt precipitability as a function of the intactness of the end regions. We have for the most part avoided conditions which produce cleavage in the helical region [2].
MATERIALS AND METHODS
Basement Membrane Collagen Extraction
The procedure described previously [2] was utilized with some modification. The anterior lens capsules were dissected from 200 bovine eyes and immediately washed in HzO at 4°C. Repeated washings in water followed by low-speed centrifugation was used to dis- rupt any attached cells [8]. The washed capsules were suspended in 100 ml 0.1 M acetic acid at 7°C with 0.001 mg pepsin/ml (Worthington). After a 24-h digestion the capsules were centrifuged at 20000 x g, the supernatant removed and the capsular material resuspended in a second 100 ml of 0.1 M acetic acid with 0.001 mg pepsin/ml. This procedure was repeated at 48 and 72 h and at 7, 11 and 14 days after initiation of the digestion. After 14 days the remaining capsular material was brought up to 23 "C and 5 mg pepsin was added to the 100 ml 0.1 M acetic acid solution. After a final two days of digestion no insoluble residue remained.
Each of the supernatants collected prior to redi- gestion was immediately adjusted to pH 7 with sodium hydroxide. Sodium chloride was added to 5% (w/v) and the resultant collagen precipitate collected by centrifugation and dissolved in 0.1 M acetic acid. The small amount of precipitate not soluble in 0.1 M acetic acid was removed by centrifugation. The solutions were dialyzed against HzO (7 days, daily changes) to remove sodium chloride.
In a separate set of experiments a number of cap- sules were subjected to a single 24-h pepsin exposure that was followed by repeated acetic acid extractions without any further addition of pepsin or enzyme inhibitors. The acetic acid extractions were made every third day for a period of 25 days at 4 "C.
Sodium Chloride Precipitation
Collagen from the 48-h, 7-day and 16-day extracts was dissolved in 0.1 M acetic acid at 0.5 mg/ml and spun at 20000 x g to remove any particulate material. Four 5-ml aliquots of each extract were placed in plastic centrifuge tubes at 7°C. To each tube 4.0 M
NaCl at 7 "C was added to bring the final NaCl con- centration to 0.3, 0.4, 0.5 or 0.6 M, respectively. The precipitates which formed in each tube after storage overnight in the cold room were separated from the supernatant by centrifugation. Precipitates were de- salted for gel electrophoresis. The 0.6 M NaCl super- natant was subsequently made 2 M in NaCl without appearance of further precipitate. The 2.0 M NaCl solution was desalted by dialysis, lyophilized and the small amount of material which resulted taken for electrophoresis.
Thermal Gelation
Portions of the lens capsule extracts at 0.25 mg/ml were dialyzed against 0.1 M acetic acid, centrifuged to removed insoluble material and dialyzed again overnight at 7 "C against 0.02 M sodium phosphate buffer, pH 7.10. After the overnight dialysis some cloudiness was apparent in the dialysis tubes of the 48-h and 7-day extracts. Half of each sample was placed in a pre-cooled flask at 4 "C and the remainder centri- fuged at 4°C at 20000 rev./min for 1 h in a Sorvall SS34 rotor. The clarified solutions were kept at 4°C.
1-ml aliquots of each collagen solution were trans- ferred to quartz cuvettes, held in a temperature-con- trolled jacket in a recording spectrophotometer. After equilibration at 4 "C and establishment of base line absorbance, the temperature was increased to 23 "C and the absorbance monitored as a function of time. Equilibration at the higher temperature was accom- plished within a 2 - 3-min period. After thermal gela- tion, the precipitated protein was collected for elec- tron microscopy, electrophoresis and amino acid analysis.
Electron Microscopy
Drops of gel suspension to be examined were placed on carbon/formvar-coated grids. Without al- lowing the drop to dry, the grid was immediately washed in phosphate buffer at pH 7. Finally, a few drops of 2 % phosphotungstic acid adjusted to pH 7 were run over the grid. The grid was blotted with filter paper to remove most of the stain and then air dried. This procedure has been shown to eliminate many of the artificial aggregates of collagen molecules which can be produced by the routine handling of collagen solutions for electron microscopy [9].
Amino Acid Analysis and Gel Electrophoresis
Samples for amino acid analysis were hydrolyzed at 105 "C for 22 h in 6 M HC1 in sealed nitrogen- flushed tubes. Analyses were carried out using a Jeol 6 AH analyzer with a single-column program. Poly- acrylamide gel electrophoresis in sodium dodecylsul-

D. Schwartz, T. Chin-Quee, and A. Veis 23
Table 1. Amino acid compositions of the sequential lens capsule extracts Each analysis is the average of two independent analyses. Results are given as residues/1000 residues
Amino acid Amount in extract obtained at Type I (rat skin)
24 h 48 h 7 days 11 days 16 days
Lysine Histidine Arginine Hydroxylysine 4-Hydroxyproline 3-Hydroxyproline Aspartic acid Threonine Serine Glutamic acid Proline Glycine A 1 an i n e Half-cystine Valine Methionine Isoleucine Leucine Tyrosine Phenylalanine
10 9
28 45
128 11 55 26 55
100 69
269 41 5
30 10 23 50 7
28
11 9
30 41
127 15 53 24 37 86 70
300 39 8
19 14 22 56 7
29
7 8
25 48
133 18 51 24 35 76 71
294 40 11 17 17 25 63 2
29
7 8
25 41
109 21 62 27 52 88 60
293 38
5 28 14 27 53 10 30
7 8
23 40
109 23 69 33 60 80 66
266 40 16 21 11 27 54 15 28
27 5
42 7
102 3
47 16 36 76
123 329 109
16 7 9
25 4
13
-
fate followed the procedure of Furthmayr and Timpl [lo]. Mercaptoethanol or dithioerythritol were added to 1 concentration in sample and running buffers.
Collagenase Digests
Bacterial collagenase purified by the method of Peterkofsky and Dieglemann [ l l ] was used as de- scribed [2] to determine the collagenase sensitivity of each extract.
RESULTS
Characterization of the Extracted Collagens
The final pepsin digestion at 23 "C ultimately led to the complete solubilization of the capsules but more than 75 % of the recoverable high-molecular-weight material was obtained in the digests carried out at low temperature in the first 7 days of extraction. 30 mg material was extracted after 24 h, and 20 mg each after 48 h and 7 days of digestion. The amino acid analyses of all extracts (Table 1) are comparable to that of a standard basement membrane collagen (kindly pro- vided by Dr N. A. Kefalides) but distinctly different from type 1 collagen (see Table 1). There are notable differences in the content of certain amino acids be- tween the sequential extracts, suggesting that the cap- sular proteins are not being degraded in a uniform fash- ion. For instance, the glycine content of the 48-h extract is higher than that of the 24-h extract and remains
elevated in each extract until the final high-tempera- ture pepsinization. This is also true of 3-hydroxy- proline. In contrast, the 4-hydroxyproline content decreases after 7 days of digestion.
Fig.1 shows the results of gel electrophoresis of the collagens from different extracts, obtained in each case under reducing conditions on the same slab gel. Lanes 1 and 7 are standard type I and show the positions of ctl(I), p11(I) and y(I), used for molecular weight calibration. A large portion of the 24-h base- ment membrane collagen is highly aggregated and does not enter the gel. In addition to a P-like component with M, of 220000 (based on the type I collagen cali- bration), there are four distinct bands which appear in the pro-a region, withM, of 170000,160000,150000 and 140000. As the low-temperature digestion of the capsular residue continues, at low pepsin/collagen ratios, slightly lower-molecular-weight collagens are produced. This is shown by the increase in density of the 140000-M, band and the disappearance of the 170000-Mr component. Over the first 7 days of diges- tion all of the collagen in each extract is in the form of high-molecular-weight components. Even under de- naturing and reducing conditions (as in Fig. 1) a major portion of the collagen is in a highly aggregated state, not entering the running gel until the digestion has been carried on past 7- 11 days. Assuming that the intact molecule has a continuous helical region as in type I, then the pepsin-sensitive helical region [2] begins to be attacked at between 7 - 11 days under our conditions of digestion. In addition to a 1400O0-Mr

24
P
a
a
Basement-Membrane Collagen: Pepsinization, Precipitation, Heat Gelation
Y
P
a
1 2 3 4 5 6 7
Fig. 1. Comparison of 24-h, 48-h, 7-day, 11-day and 16-day pepsin digests of anterior lens collagen with type I collagen, on 6% poly- acrylamide gels in sodium dodecylsulfate, under reducing conditions. All but the 16-day extraction was carried out at 4 'C at low pepsin concentration (see text). Among those components which are frac- tionated there is an obvious shift from higher to lower-molecular- weight species. All bands are eliminated after bacterial collagenase digestion. Lanes 1 and 7, standard type I ; lane 2, 24-h extract; lane 3, 48-h extract; lane 4, 7-day extract; lane 5, 11-day extract; lane 6, 16-day extract
component, the 1 1-day extract contains chain frag- ments with M , of 85000 and 60000. The content of aggregated material is greatly decreased at 11 days (lane 5 , Fig. 1).
All of the aggregated material in Fig. 1 and all of the bands attributed above to collagen fragments of LY chains are collagenous as demonstrated by their complete removal by collagenase digestion.
The final small portion of the capsular material which required digestion at 23 "C and high pepsin/ collagen ratios for solubilization was slightly different in composition (Table 1) from the 48-h extract. This was especially evident in the increased content of 3-hydroxyproline, half-cystine and tyrosine. These data suggest that the 16-day extract represented selected, pepsin-resistant and cross-linked portions of type IV collagen rather than intact molecules. The gel electrophoresis data lead to the same conclusion in that all fragments were of low molecular weight (Fig. 1, lane 6), with only faint bands appearing at the 1000OO-Mr and 85000-M, positions and the majority appearing at 60000 and smaller.
In the parallel experiment involving only a single 24-h pepsinization followed by repeated acid extrac- tions of the residual capsules over a period of 31 days, there was no shift of high-molecular-weight compo- nents in the extract nor was there any significant appearance of material less than 140000 in molecular
Fig. 2. NaCl precipitation of the 48-h pepsin extract. Salt was added from a 4 M NaCl solution at 4 "C to the extract dissolved in 0.1 M acetic acid also at 4'C. Precipitation began at 0.3 M NaCl and was complete by 0.6 M NaCI. This result was identical for the 7-day extract; however, the 11-day and 16-day extracts showed no salt precipitability
weight. Thus it is unlikely that endogenous protease activity could account for the production of the specific components described above.
Sodium Chloride Precipitation
Chung et al. [I21 separated interstitial collagen from pepsin-digested basement membrane collagen by salt precipitation from acetic acid solutions. At 0.9 M NaCl in 0.5 M acetic type I and I11 collagens were precipitated [13] whereas the basement mem- brane collagen remained in solution [12]. Upon apply- ing this method directly to the various pepsin digest supernatants, with the intention of removing any type I or I11 collagens, we observed (as illustrated in Fig.2) that precipitates were obtained when the sodium chloride concentration was brought to 0.3 M and that precipitation was complete at 0.6 M NaCl with the 48-h extract. Identical results were obtained for the 7-day extract. Components seen in acrylamide gel electrophoresis of the 0.6 M precipitate were identical to those seen in the initial sample, and even under conditions of over-loading, no al(1) or a2 chains could be detected. The amino acid composition of the pre- cipitate was comparable to that given in Table 1, that is, typical of 48-h basement membrane collagen extract. In comparison, the 16-day preparation, ob- tained as described by Chung et al. [12] at a 1/10 pep- sin/collagen ratio rather than the 1 /lo00 pepsin/colla- gen ratio used for our digests, showed no precipitation up to 1.2 M NaCl.
Thermal Gelation
Trelstad and Lawley [4] utilized thermal gelation as a means for separating type IV collagen from the

D. Schwartz, T. Chin-Quee, and A. Veis 25
0.2
0 0.1 8
T
0
not centrifuged 7
I H
0 15 30 45 60 Time (min)
Fig. 3. Thermal gelation oj48-h pepsin extrucl ut 23°C in phosphate hull& p H 7.03. Upper curve: solution was not centrifuged to remove aggregated material which had formed after overnight dialysis of the extract against the phosphate buffer at 4°C. The lower curve shows the introduction of a short but definite lag phase if the dialysis-induced aggregate is removed by centrifugation before the temperature is increased from 4 "C to 23 "C. Inset is a comparison of the electrophoresis patterns of the final precipitate and type I collagen. There is no indication of type I a-chain components. All the precipitated a chains are in the high-molecular-weight range
Fig. 4. Electron microgruph of the pre-centrqiged thermully precipitated material in Fig. 3. The material was prepared under conditions designed to minimize artificial aggregation during preparation for microscopy (see text). The precipitate is composed of slender 'fibrils', 3-10 nm in diameter, randomly oriented and without any indication of periodicity, similar in structure to the filaments of the anterior lens capsule in vivo
interstitial collagens. The type IV collagen remained as supernatant after centrifugation of the type I and I11 collagen precipitates from kidney, lung and spleen pepsin-solubilized tissue homogenates. As we and others have shown, the anterior lens capsule is almost entirely collagen of the type IV variety. When 48-h
pepsin-digested collagen was dialyzed into 0.02 M sodium phosphate, pH 7.1 at 4 "C, in preparation for thermal gelation, the solutions became turbid. When this turbid solution was transferred to a spectropho- tometer curvette and adjusted to 23 "C precipitation was immediate (Fig. 3 , upper curve). If the solution
26
not centrifuged
cycled
centrifuged P
0
Basemer
0 15 30 45 60 Time (rnin)
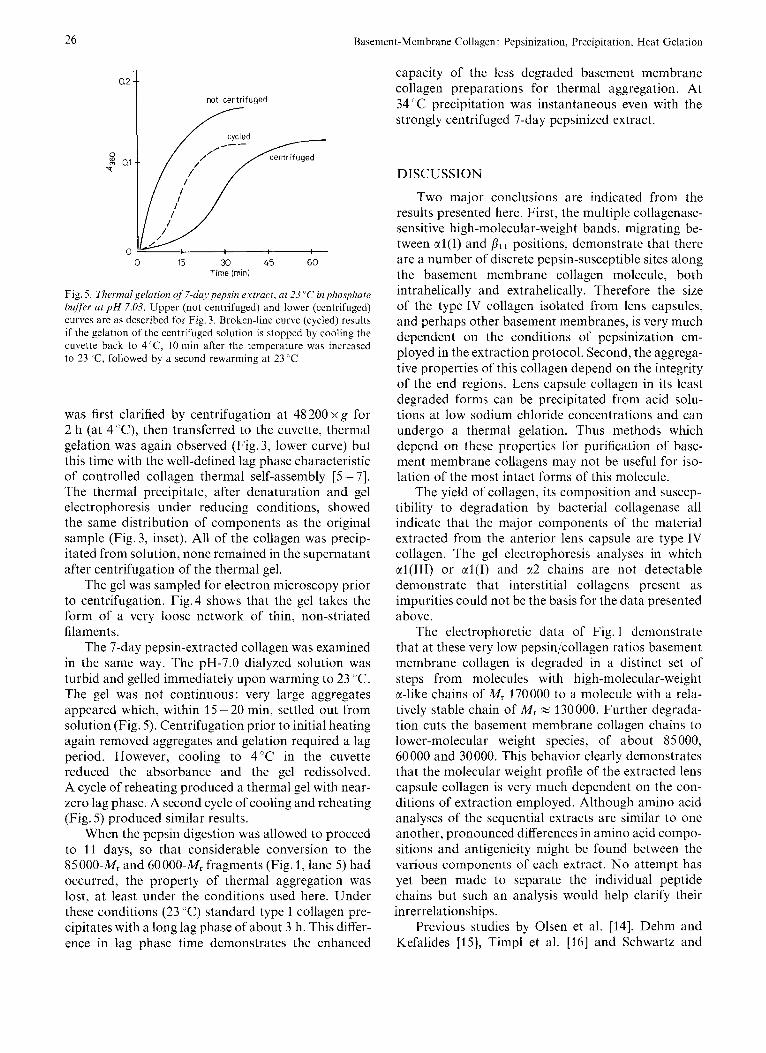
Fig. 5 . Tlierrnalgelution of 7-day pepsin extract, at 23 'L' in phosphate buffer ut p H 7.03. Upper (not centrifuged) and lower (centrifuged) curves are as described for Fig. 3. Broken-line curve (cycled) results if the gelation of the centrifuged solution is stopped by cooling the cuvette back to 4'C, 10 min after the temperature was increased to 23 'C, followed by a second rewarming at 23 ' C
was first clarified by centrifugation at 48 200 x g for 2 h (at 4 rC), then transferred to the cuvette, thermal gelation was again observed (Fig. 3, lower curve) but this time with the well-defined lag phase characteristic of controlled collagen thermal self-assembly [5 - 71. The thermal precipitate, after denaturation and gel electrophoresis under reducing conditions, showed the same distribution of components as the original sample (Fig. 3, inset). All of the collagen was precip- itated from solution, none remained in the supernatant after centrifugation of the thermal gel.
The gel was sampled for electron microscopy prior to centrifugation. Fig.4 shows that the gel takes the form of a very loose network of thin, non-striated filaments.
The 7-day pepsin-extracted collagen was examined in the same way. The pH-7.0 dialyzed solution was turbid and gelled immediately upon warming to 23 "C. The gel was not continuous: very large aggregates appeared which, within 15 - 20 min, settled out from solution (Fig. 5). Centrifugation prior to initial heating again removed aggregates and gelation required a lag period. However, cooling to 4'C in the cuvette reduced the absorbance and the gel redissolved. A cycle of reheating produced a thermal gel with near- zero lag phase. A second cycle of cooling and reheating (Fig. 5 ) produced similar results.
When the pepsin digestion was allowed to proceed to 11 days, so that considerable conversion to the 85 000-M, and 60 000-M, fragments (Fig. 1, lane 5) had occurred, the property of thermal aggregation was lost, at least under the conditions used here. Under these conditions (23 "C) standard type I collagen pre- cipitates with a long lag phase of about 3 h. This differ- ence in lag phase time demonstrates the enhanced
it-Membrane Collagen : Pepsinization, Precipitation, Heat Gelation
capacity of the less degraded basement membrane collagen preparations for thermal aggregation. At 34 "C precipitation was instantaneous even with the strongly centrifuged 7-day pepsinized extract.
DISCUSSION
Two major conclusions are indicated from the results presented here. First, the multiple collagenase- sensitive high-molecular-weight bands, migrating be- tween al(1) and ,&I positions, demonstrate that there are a number of discrete pepsin-susceptible sites along the basement membrane collagen molecule, both intrahelically and extrahelically. Therefore the size of the type IV collagen isolated from lens capsules, and perhaps other basement membranes, is very much dependent on the conditions of pepsinization em- ployed in the extraction protocol. Second, the aggrega- tive properties of this collagen depend on the integrity of the end regions. Lens capsule collagen in its least degraded forms can be precipitated from acid solu- tions at low sodium chloride concentrations and can undergo a thermal gelation. Thus methods which depend on these properties for purification of base- ment membrane collagens may not be useful for iso- lation of the most intact forms of this molecule.
The yield of collagen, its composition and suscep- tibility to degradation by bacterial collagenase all indicate that the major components of the material extracted from the anterior lens capsule are type IV collagen. The gel electrophoresis analyses in which al(II1) or al(1) and a2 chains are not detectable demonstrate that interstitial collagens present as impurities could not be the basis for the data presented above.
The electrophoretic data of Fig. 1 demonstrate that at these very low pepsin/collagen ratios basement membrane collagen is degraded in a distinct set of steps from molecules with high-molecular-weight a-like chains of M , 170000 to a molecule with a rela- tively stable chain of M, = 130 000. Further degrada- tion cuts the basement membrane collagen chains to lower-molecular weight species, of about 85 000, 60000 and 30000. This behavior clearly demonstrates that the molecular weight profile of the extracted lens capsule collagen is very much dependent on the con- ditions of extraction employed. Although amino acid analyses of the sequential extracts are similar to one another, pronounced differences in amino acid compo- sitions and antigenicity might be found between the various components of each extract. No attempt has yet been made to separate the individual peptide chains but such an analysis would help clarify their inrerrelationships.
Previous studies by Olsen et al. [14], Dehm and Kefalides [15], Timpl et al. [16] and Schwartz and

D. Schwartz, T. Chin-Quee, and A. Veis 21
Veis [2,3] all show that basement membrane collagens contain helical collagenous and non-helical non-colla- genous regions, although the distribution and arrange- ment of these regions in the intact basement membrane structure is not completely known [2]. All of these investigations have shown that the non-collagenous proteins are susceptible to proteases and we have demonstrated that there is at least one pepsin-sensitive region within the triple helix itself [2]. Olsen et al. [14] showed that long filamentous structures, 300 nm in length, could be produced from solubilized lens cap- sule collagen, and Dehm and Kefalides [15] demon- strated that a-chain-size peptide chains could be pro- duced from these and similar molecules after reduc- tion and alkylation and a second pepsinization. All of these data, and our electron microscopic observa- tions, suggest that the four bands observed in lanes 2-5 on the gel of Fig.1, in the 170000-130000-M, region, are single chains with procollagen-like exten- sions. The 220000-M, band, which decreases as pep- sinization time lengthens, may be a two-chain aggre- gate stabilized by cross-linkages not reducible with thiol reagents. It should be noted that all of the molec- ular weight assignments are based on the migration of ctl(1) chains and aggregates and will have to be adjusted when definitive species are isolated and mea- sured.
The salt-precipitation experiments indicate the proclivity of the high-molecular-weight molecules to aggregate. The rather sharp demarcation in sensitivity to salt-precipitability between the 7-day and 11 -day treated samples suggest that the non-collagenous por- tions of this molecule are very important. In particular, those molecules greater than 130000 in relative weight may provide the specific non-collagen regions for interaction and aggregation. The preparations of Chung et al. [12] contain some molecules comprised of a chains with M, of 110 000, mixed with molecules or fragments with M, 55000. These preparations, similar to our 1 1 -day and 16-day pepsin-treated prep-
arations do not precipitate at 0.9 M NaCl concen- trations.
The'thermal gelation properties of the extracts are entirely in accord with the very marked aggregation tendency related to the presence of the high-molec- ular-weight a chains. The network structures and the fine filament diameters seen in the gels support the idea that end-region interactions are crucial and perhaps do prevent helical region interactions in vivo, and the formation of D-periodic fibrils.
This work was supported by a grant from the National Institute of Arthritis, Metabolism and Digestive Diseases AM13921 (to A.V.) and the General Research Support from United States Public Health ServiceiNational Institutes of Health RR 0531 1 (to D.S.).
REFERENCES
1.
2. 3. 4.
5. 6. 7.
8.
9. 10. 11.
12.
13.
14.
15.
16.
Jakus, M. A. (1974) in Oculur Fine Structure, pp. 172-184,
Schwartz, D. & Veis, A. (1978) FEBS Lett. 85, 326-332. Schwartz, D. & Veis, A. (1979) Biopolymers, 18, 2363-2367. Trelstad, R. L. & Lawley, K. K. (1977) Biochem. Biophys. Res.
Comper, W. & Veis, A. (1977) Biopolymers, 16, 2113-2131. Comper, W. & Veis, A. (1977) Biopolymers, 16, 2133-2142. Williams, B. R., Gelman, R. A, , Poppke, D. C. 8 Piez, K. A.
Meezan, E., Hjelle, J. T. & Brendel, K . (1976) Lije Sci. 17,
Schwartz, D. & Veis, A. (1978) Connect. Tin. Res. 6, 185- 190. Furthmayr,H. & Timpl, R. (1971) Anal. Biochem. 41,510-516. Peterkofsky, B. & Dieglemann, R. (1971) Biochemistry, 10,
Chung, E., Rhodes, R. & Miller, E. (1976) Biochem. Biophys.
Chung, E. & Miller, 1:. (1974) Science (Wash. D.C.) 183,
Olsen, B. R., Alper, R. & Kefalides, N. A. (1973) Eur. J . Bio-
Dehm, P. & Kefalides, N. A. (1978) J . B i d Chrm. 253, 6680-
Timpl, R., Martin, G., Brucker, P., Wick, G. & Weideman, H.
Little, Brown, Inc. Boston.
Commun. 76, 376- 384.
(1978) J . Bid. Chem. 253, 6578 - 6585.
1721 - 1732.
988 - 994.
Kcs. Commun. 71, 1167-1174.
1200 - 1201.
chem. 38,220 - 228.
6686.
(1978) Eur. J . Biochem. 84, 43 - 52.
D. Schwartz, T. Chin-Quee, and A. Veis. Department of Oral Biology. Northwestern University Dental School 303 East Chicago Avenue, Chicago, Illmois, U.S.A. 60611





























