Embed Size (px)
Citation preview

International Journal of Food Microbiology 129 (2009) 150–156
Contents lists available at ScienceDirect
International Journal of Food Microbiology
j ourna l homepage: www.e lsev ie r.com/ locate / i j foodmicro
Characterisation of non-pathogenic Yersinia pseudotuberculosis-like strains isolatedfrom food and environmental samples
T. Niskanen a,⁎, R. Laukkanen a, A. Murros a, J. Björkroth a, M. Skurnik b,H. Korkeala a, M. Fredriksson-Ahomaa c
a Department of Food and Environmental Hygiene, Faculty of Veterinary Medicine, P.O. Box 66, FI-00014 University of Helsinki, Helsinki, Finlandb Department of Bacteriology and Immunology, Haartman Institute, University of Helsinki, and Helsinki University Central Hospital Laboratory Diagnostics, Helsinki, Finlandc Institute of Hygiene and Technology of Food of Animal Origin, Faculty of Veterinary Medicine, Ludwig Maximilian University, Munich, Germany
⁎ Corresponding author.E-mail address: [email protected] (T. Niskanen
0168-1605/$ – see front matter © 2008 Elsevier B.V. Aldoi:10.1016/j.ijfoodmicro.2008.11.015
a b s t r a c t
a r t i c l e i n f oArticle history:
Non-pathogenic Yersinia pse Received 4 June 2008Received in revised form 5 November 2008Accepted 9 November 2008Keywords:Yersinia pseudotuberculosisFoodEnvironmental isolatesPFGERibotypingLPSO-antigen
udotuberculosis-like strains were recovered from Finnish food and environmentalsamples. These strains could not be differentiated from Y. pseudotuberculosis strains using API 20E or otherphenotypical tests. However, all of the strains were inv-, and virF-negative with polymerase chain reaction(PCR), while all Y. pseudotuberculosis strains used as controls were inv-positive and fresh Y. pseudotuberculosisstrains were also virF-positive, indicating that the Y. pseudotuberculosis-like strains were non-pathogenic.Using pulsed-field gel electrophoresis (PFGE) with NotI enzyme and ribotyping with EcoRI and HindIIIenzymes, the Y. pseudotuberculosis-like strains, which grouped genetically together, could be differentiatedfrom true Y. pseudotuberculosis strains and from strains belonging to other sucrose-negative Yersinia species.In addition, the O-antigen gene cluster of one Y. pseudotuberculosis-like strain was characterized, and itdiffered from those of known Y. pseudotuberculosis serotypes. This study demonstrates that identification ofY. pseudotuberculosis from food and environmental sources using solely biochemical reactions can beincorrect, and when a strain cannot be serotyped to known Y. pseudotuberculosis serotypes, the pathogenicpotential of isolates should be determined.
© 2008 Elsevier B.V. All rights reserved.
1. Introduction
Yersinia pseudotuberculosis is a Gram-negative bacterium capableof infecting humans and a wide range of animals. It causes a variety ofintestinal and extra-intestinal infections in humans and animals andsuch post-infectious complications as arthritis and erythema nodosusin humans (Jalava et al., 2006). Although most cases of humaninfections are sporadic (Vincent et al., 2008), several outbreaks haverecently been reported in Finland (Jalava et al., 2004; Rimhanen-Finneet al., 2008). In the scientific opinion on monitoring and identificationof human enteropathogenic Yersinia spp., which has recently beenpresented by European Food Safety Agency (EFSA), all Y. pseudotu-berculosis strains should be considered pathogenic (EFSA, 2007).
Y. pseudotuberculosis can be divided into 15 O-serotypes (O:1–O:15)and into 10 subtypes (O:1a–O:1c, O:2a–O:2c, O:4a–O:4b, O:5a–O:5b)based on variability in the lipopolysaccharide (LPS) O-side chain (O-antigen) (Bogdanovich et al., 2003). Themost commonly isolated strainsin Europe belong to the serotypes O:1–O:3, while serotypes O:4–O:15mainly occur in Asia. Most Y. pseudotuberculosis strains are consideredpathogenic; only strains of the relatively rare serotypes O:7 and O:11–
).
l rights reserved.
O:15 are apparently non-pathogenic (Nagano et al., 1997; Skurnik,2004). The pathogenic strains contain a 70 kb virulence plasmid (pYV),which is essential for all pathogenic Yersinia species, and chromosomalgenes encoding a number of virulence factors. Among the chromosomalfactors, the invasinprotein (Inv) is themost important factor in the earlyphases of intestinal infection (Marra and Isberg, 1997). It promotesbinding and penetration of Y. pseudotuberculosis into cells at the surfaceof the intestinal lumen (Isberg et al., 1987, Simonet and Falkow, 1992).
The API 20E identification kit is widely used for identification ofpresumptive Yersinia isolates; it has been shown to be accurate inidentifying Y. enterocolitica and Y. pseudotuberculosis. API 20E has apositive identification rate of 90% for Y. pseudotuberculosis whenincubated at a temperature between 25 °C and 30 °C, instead of at37 °C (Neubauer et al., 1998). Polymerase chain reaction (PCR) is arapid and specific method for identifying bacterial species. The invgene in the chromosome of Y. pseudotuberculosis has been used toidentify this species (Nakajima et al., 1992; Kaneko et al., 1995). Thegenes responsible for the biosynthesis of the O-antigen are located inthe chromosome of Y. pseudotuberculosis between the hemH and gskgenes (Skurnik, 1999). Multiplex PCR assay based on the geneticpolymorphism of this O-antigen gene cluster is a useful tool togenetically serotype all known Y. pseudotuberculosis serotypes andalso to correct misidentification of strains as Y. pseudotuberculosis(Bogdanovich et al., 2003).

151T. Niskanen et al. / International Journal of Food Microbiology 129 (2009) 150–156
Molecular techniques, like pulsed-field gel electrophoresis (PFGEand ribotyping, are valuable alternatives for characterisation of Y.pseudotuberculosis. PFGE has been shown to have the highestdiscriminatory power for subtyping of Y. pseudotuberculosis strains(Iteman et al., 1995, Niskanen et al., 2002; Nuorti et al., 2004).Ribotyping using EcoRI and EcoRV allows subtyping of strains of thesame serotype (Voskressenskaya et al., 2005). However, ribotypinghas some limitations, e.g. it does not clearly differentiate between Y.pseudotuberculosis and Y. pestis, and the varying bands may exhibitonly slight differences in size, making identification challenging.
The aim of this work was to characterise three presumptive Y.pseudotuberculosis strains isolated from food and environmentalsamples using several phenotypical and genotypical tests. In thisreport, we show that the strains initially identified as Y. pseudotu-berculosis by biochemical tests were non-pathogenic and clearlydiffered from Y. pseudotuberculosis strains.
2. Materials and methods
2.1. Sucrose-negative Yersinia strains
In total, 20 sucrose-negative strains were included in the study(Table 1). Three sucrose-negative strains (#1–3), initially identified as Y.pseudotuberculosis, were isolated according to Niskanen et al. (2002)from lettuce, water and soil in 1999 and 2000 during two Y.pseudotuberculosis outbreaks in southern Finland (Nuorti et al., 2004).The identification as Y. pseudotuberculosis was accomplished using API20E (BioMérieux, Marcy l’Etoile, France), which has been shown to bean accurate test for identification of Y. pseudotuberculosis isolates(Neubauer et al.,1998). However, as these strains could not be serotypedby serotypeO:1–O:6-specific antisera (Denka Seikan, Tokyo, Japan), theywere further characterized together with 17 other sucrose-negativestrains (#4–20) belonging to different Yersinia species (Table 1). Allstrains except the reference strains were isolated at the Department ofFood and Environmental Hygiene, Helsinki according to Niskanen et al.(2002). All the20pure cultured strains included in this studywere storesat −70 °C in Cryobank™ tubes (Mast Group, Merseyland, UK ).
All further characterisations except PCR-serotyping and O-antigencharacterisation were done at the Department of Food and Environ-
Table 1Sucrose-negative Yersinia strains characterized in this study
Strain # Species Source Isolation year
1 ÅYV7.1 KOH2a Yersinia spp Water 19992 ÅYV25K1a Yersinia spp Soil 19993 Å125 KOH2a Yersinia spp Lettuce 20004 ÅYV162.1K3a Y. pseudotuberculosis Lettuce 20005 CCGU 5855b Y. pseudotuberculosis6 CIP 55.86b Y. pseudotuberculosis 19557 CIP 55.87b Y. pseudotuberculosis Mink 19558 CIP A119b Y. pseudotuberculosis Pig 19529 LIP91 Y. pseudotuberculosis Bird faeces 199810 YLI16.5K3c Y. pseudotuberculosis Bird faeces 200111 YTSN26.1Pd Y. pseudotuberculosis Pig tonsils 199912 YTSL3.3K2d Y. pseudotuberculosis Pig tonsils 199913 YTPS40.1Kd Y. pseudotuberculosis Pig tonsils 199914 YTPS37.1P1d Y. pseudotuberculosis Pig tonsils 199915 CIP 80.30b Y. kristensenii Human urine 198016 ATCC 33639b Y. kristensenii17 CIP 103162b Y. aldovae Drinking water 198818 DSM 14987b Y. aleksiciae Human faeces19 P102e Y. enterocolitica Pig tonsils 199520 A160e Y. enterocolitica Pig tonsils 1995
a Nuorti et al., 2004.b Reference strain.c Niskanen et al., 2003.d Niskanen et al., 2002.e Fredriksson-Ahomaa et al., 2002.
mental, Helsinki. PCR-serotyping and O-antigen characterisationweredone at the Department of Bacteriology and Immunology, Helsinki.Before characterisation, the pure cultured strains from Cryobank™tubes were plated on tryptic soy agar (TSA, Difco, Detroit, MI, USA) andincubated at 30 °C for 20–24 h.
2.2. Identification using API 20E, biotyping and serotyping
All 20 sucrose-negative strains were characterised using the API20E system. Due to the temperature dependence of the Voges–Proskauer test, the API 20E system was incubated at 25 °C instead of37 °C for 20–24 h. The strains were identified using the databaseprovided by the manufacturer (APILAB V4.0, BioMeriéux).
All strains were biotyped using the reactions for Y. pseudotubercu-losis (raffinose, melibiose, citrate) (Tsubokura & Aleksic, 1995) and forY. enterocolitica (pyrazinamide, esculin, salicin, tween, indol, xylose,trehalose) (Wauters et al., 1987). All tests except melibiose, citrate andindol tests were done according to International Organisation forStandardisation (ISO, 2003). Melibiose, citrate and indol tests wereperformed as part of the API 20E system.
The O-antigens were determined by slide agglutination test usingcommercial O:1–O:6 antisera for Y. pseudotuberculosis and O:3 forY. enterocolitica (Denka Seiken) Multiplex PCR assay, allowing dis-tinction of all 21 known Y. pseudotuberculosis serotypes, was furtherused for typing Y. pseudotuberculosis-like strains according toBogdanovich et al. (2003). Template DNA was prepared directly frombacterial colonies by boiling. Briefly, one colony was suspended in100 µl of water and boiled for 10 min. Two microliters of supernatantafter being centrifuged at 13,000 rpm for 5 min was used for PCR.
2.3. Pathogenicity tests
The pathogenic potential of the sucrose-negative strainswas studiedby pheno- and genotypic methods. The virF gene located on the Yersiniavirulence plasmid, the inv gene in the chromosome of Y. pseudotubercu-losis and the ail gene, in the chromosome of Y. enterocolitica weredetected using PCR (Nakajima et al., 1992). In brief, 2–3 colonies frompure culture on TSA were suspended in 100 µl of water. DNA wasreleasedbyboiling the suspension for 10min. The lysatewas centrifugedfor 3min (13000 rpm). Twomicrolitres of the supernatant were used asa template. The PCR reaction (50 µl) contained 0.5 U of Dynazyme DNApolymerase (Finnzyme, Espoo, Finland), 200 µM of each dNTP(Finnzyme) and 0.3 µM of each primer (Oligomer, Helsinki, Finland).The primers based on the sequences of virF (5′-TCGTGGCAGCTATG-CTGTTC-3′ and 5′-ATACGTCGCTCGCTTATCCA-3′), inv (5′-TAAGGGTAC-TATCGCGGCGGA-3′ and 5′-CGTGAAATTAACCGTCACACT-3′) and ail (5′-ACTCGATGATAACTGGGGAG-3′ and 5′-CCCCCAGTAATCCATAAAGGG-3′)were used to amplify a 492-, 295- and 170-bp fragment, respectively.
Pyrazinamidase activity was tested by the method of Kandolo andWauters (1985) and done according to ISO (2003). In brief, somecolonies from pure culture on TSA were inoculated on the slant ofpyrazinamidase agar tubes, which were incubated at 25 °C for 48 h.One millilitre of 1% freshly prepared ferrous ammonium sulphatesolutionwas then flooded on the slant. Non-pathogenic strains show apyrazinamidase-positive reaction, which can be seen as a red colourafter 1–5 min.
Calcium dependence and Congo red uptake at 37 °C were per-formed on Congo red-magnesium oxalate agar (CR-MOX) according toRiley and Toma (1989). The strains were grown on CR-MOX agar platesat 37 °C for 24 h. pYV-positive strains grow as small red colonies at37 °C, while pYV-negative strains grow as large colourless colonies.
2.4. Characterization of the O-antigen gene cluster of strain Å125 KOH2
The Expand™ Long Template PCR System (Roche, Basel, Switzer-land) was used to amplify the hemH-gsk region of strain Å125
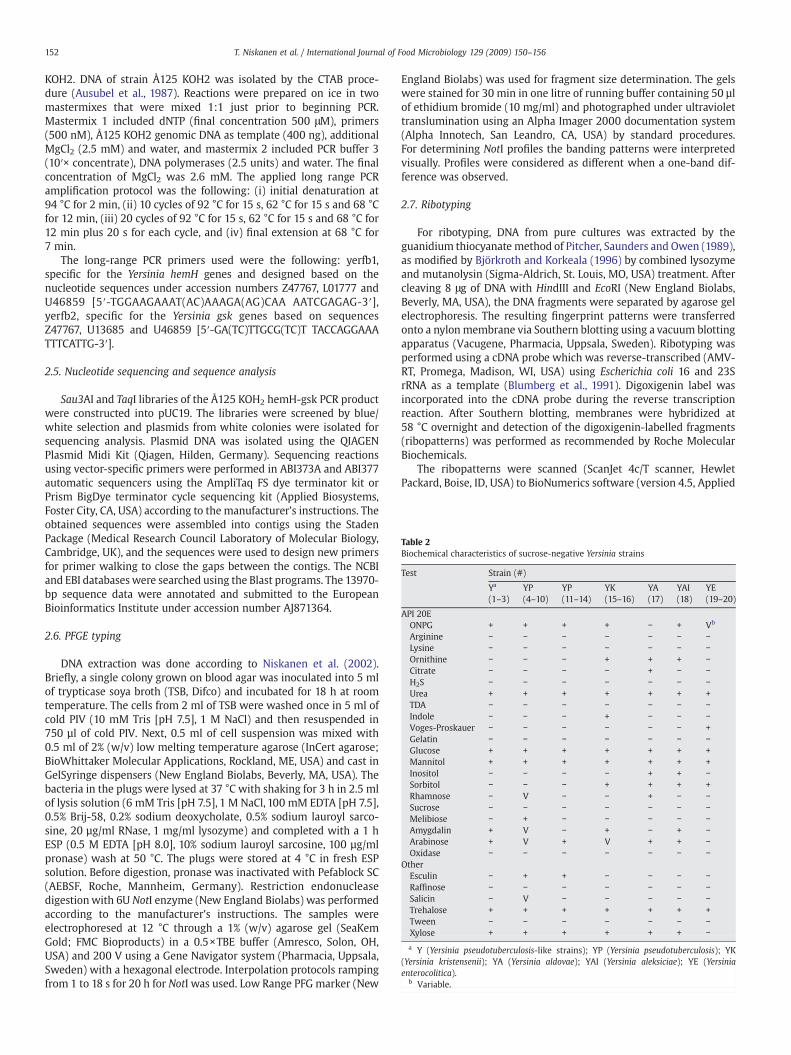
Table 2Biochemical characteristics of sucrose-negative Yersinia strains
Test Strain (#)
Ya
(1–3)YP(4–10)
YP(11–14)
YK(15–16)
YA(17)
YAI(18)
YE(19–20)
API 20EONPG + + + + − + Vb
Arginine − − − − − − −Lysine − − − − − − −Ornithine − − − + + + −Citrate − − − − + − −H2S − − − − − − −Urea + + + + + + +TDA − − − − − − −Indole − − − + − − −Voges-Proskauer − − − − − − +Gelatin − − − − − − −Glucose + + + + + + +Mannitol + + + + + + +Inositol − − − − + + −Sorbitol − − − + + + +Rhamnose − V − − + − −Sucrose − − − − − − −Melibiose − + − − − − −Amygdalin + V − + − + −Arabinose + V + V + + −Oxidase − − − − − − −
OtherEsculin − + + − − − −Raffinose − − − − − − −Salicin − V − − − − −Trehalose + + + + + + +Tween − − − − − − −Xylose + + + + + + −a Y (Yersinia pseudotuberculosis-like strains); YP (Yersinia pseudotuberculosis); YK
(Yersinia kristensenii); YA (Yersinia aldovae); YAI (Yersinia aleksiciae); YE (Yersiniaenterocolitica).
b Variable.
152 T. Niskanen et al. / International Journal of Food Microbiology 129 (2009) 150–156
KOH2. DNA of strain Å125 KOH2 was isolated by the CTAB proce-dure (Ausubel et al., 1987). Reactions were prepared on ice in twomastermixes that were mixed 1:1 just prior to beginning PCR.Mastermix 1 included dNTP (final concentration 500 μM), primers(500 nM), Å125 KOH2 genomic DNA as template (400 ng), additionalMgCl2 (2.5 mM) and water, and mastermix 2 included PCR buffer 3(10′× concentrate), DNA polymerases (2.5 units) and water. The finalconcentration of MgCl2 was 2.6 mM. The applied long range PCRamplification protocol was the following: (i) initial denaturation at94 °C for 2 min, (ii) 10 cycles of 92 °C for 15 s, 62 °C for 15 s and 68 °Cfor 12 min, (iii) 20 cycles of 92 °C for 15 s, 62 °C for 15 s and 68 °C for12 min plus 20 s for each cycle, and (iv) final extension at 68 °C for7 min.
The long-range PCR primers used were the following: yerfb1,specific for the Yersinia hemH genes and designed based on thenucleotide sequences under accession numbers Z47767, L01777 andU46859 [5′-TGGAAGAAAT(AC)AAAGA(AG)CAA AATCGAGAG-3′],yerfb2, specific for the Yersinia gsk genes based on sequencesZ47767, U13685 and U46859 [5′-GA(TC)TTGCG(TC)T TACCAGGAAATTTCATTG-3′].
2.5. Nucleotide sequencing and sequence analysis
Sau3AI and TaqI libraries of the Å125 KOH2 hemH-gsk PCR productwere constructed into pUC19. The libraries were screened by blue/white selection and plasmids from white colonies were isolated forsequencing analysis. Plasmid DNA was isolated using the QIAGENPlasmid Midi Kit (Qiagen, Hilden, Germany). Sequencing reactionsusing vector-specific primers were performed in ABI373A and ABI377automatic sequencers using the AmpliTaq FS dye terminator kit orPrism BigDye terminator cycle sequencing kit (Applied Biosystems,Foster City, CA, USA) according to the manufacturer's instructions. Theobtained sequences were assembled into contigs using the StadenPackage (Medical Research Council Laboratory of Molecular Biology,Cambridge, UK), and the sequences were used to design new primersfor primer walking to close the gaps between the contigs. The NCBIand EBI databases were searched using the Blast programs. The 13970-bp sequence data were annotated and submitted to the EuropeanBioinformatics Institute under accession number AJ871364.
2.6. PFGE typing
DNA extraction was done according to Niskanen et al. (2002).Briefly, a single colony grown on blood agar was inoculated into 5 mlof trypticase soya broth (TSB, Difco) and incubated for 18 h at roomtemperature. The cells from 2 ml of TSB were washed once in 5 ml ofcold PIV (10 mM Tris [pH 7.5], 1 M NaCl) and then resuspended in750 µl of cold PIV. Next, 0.5 ml of cell suspension was mixed with0.5 ml of 2% (w/v) low melting temperature agarose (InCert agarose;BioWhittaker Molecular Applications, Rockland, ME, USA) and cast inGelSyringe dispensers (New England Biolabs, Beverly, MA, USA). Thebacteria in the plugs were lysed at 37 °C with shaking for 3 h in 2.5 mlof lysis solution (6 mM Tris [pH 7.5], 1 M NaCl, 100 mM EDTA [pH 7.5],0.5% Brij-58, 0.2% sodium deoxycholate, 0.5% sodium lauroyl sarco-sine, 20 µg/ml RNase, 1 mg/ml lysozyme) and completed with a 1 hESP (0.5 M EDTA [pH 8.0], 10% sodium lauroyl sarcosine, 100 µg/mlpronase) wash at 50 °C. The plugs were stored at 4 °C in fresh ESPsolution. Before digestion, pronase was inactivated with Pefablock SC(AEBSF, Roche, Mannheim, Germany). Restriction endonucleasedigestionwith 6U NotI enzyme (New England Biolabs) was performedaccording to the manufacturer's instructions. The samples wereelectrophoresed at 12 °C through a 1% (w/v) agarose gel (SeaKemGold; FMC Bioproducts) in a 0.5×TBE buffer (Amresco, Solon, OH,USA) and 200 V using a Gene Navigator system (Pharmacia, Uppsala,Sweden) with a hexagonal electrode. Interpolation protocols rampingfrom 1 to 18 s for 20 h for NotI was used. Low Range PFG marker (New
England Biolabs) was used for fragment size determination. The gelswere stained for 30 min in one litre of running buffer containing 50 µlof ethidium bromide (10 mg/ml) and photographed under ultraviolettranslumination using an Alpha Imager 2000 documentation system(Alpha Innotech, San Leandro, CA, USA) by standard procedures.For determining NotI profiles the banding patterns were interpretedvisually. Profiles were considered as different when a one-band dif-ference was observed.
2.7. Ribotyping
For ribotyping, DNA from pure cultures was extracted by theguanidium thiocyanate method of Pitcher, Saunders and Owen (1989),as modified by Björkroth and Korkeala (1996) by combined lysozymeand mutanolysin (Sigma-Aldrich, St. Louis, MO, USA) treatment. Aftercleaving 8 μg of DNA with HindIII and EcoRI (New England Biolabs,Beverly, MA, USA), the DNA fragments were separated by agarose gelelectrophoresis. The resulting fingerprint patterns were transferredonto a nylon membrane via Southern blotting using a vacuum blottingapparatus (Vacugene, Pharmacia, Uppsala, Sweden). Ribotyping wasperformed using a cDNA probe which was reverse-transcribed (AMV-RT, Promega, Madison, WI, USA) using Escherichia coli 16 and 23SrRNA as a template (Blumberg et al., 1991). Digoxigenin label wasincorporated into the cDNA probe during the reverse transcriptionreaction. After Southern blotting, membranes were hybridized at58 °C overnight and detection of the digoxigenin-labelled fragments(ribopatterns) was performed as recommended by Roche MolecularBiochemicals.
The ribopatterns were scanned (ScanJet 4c/T scanner, HewletPackard, Boise, ID, USA) to BioNumerics software (version 4.5, Applied

Table 3Identification of sucrose-negative Yersinia strains using API 20E and bio- and serotyping
Strain# Species API 20E Bio-typing O-serotype
Code Identification
1 Yersinia spp. 1014103 Y. pseudotuberculosis(98.4%)
2 NTa
2 Yersinia spp. 1014103 Y. pseudotuberculosis(98.4%)
2 NT
3 Yersinia spp. 1014103 Y. pseudotuberculosis(98.4%)
2 NT
4 Y. pseudotuberculosis 1014142 Y. pseudotuberculosis(99.9%)
1 YP O:2a
5 Y. pseudotuberculosis 1014141 Y. pseudotuberculosis(99.7%)
1 NT
6 Y. pseudotuberculosis 1014152 Y. pseudotuberculosis(99.9%)
1 YP O:2
7 Y. pseudotuberculosis 1014152 Y. pseudotuberculosis(99.9%)
1 NT
8 Y. pseudotuberculosis 1014152 Y. pseudotuberculosis(99.9%)
1 NT
9 Y. pseudotuberculosis 1014152 Y. pseudotuberculosis(99.9%)
1 YP O:1
10 Y. pseudotuberculosis 1014152 Y. pseudotuberculosis(99.9%)
1 YP O:2
11 Y. pseudotuberculosis 1014102 Y. pseudotuberculosis(99.4%)
2 YP O:3
12 Y. pseudotuberculosis 1014102 Y. pseudotuberculosis(99.4%)
2 YP O:3
13 Y. pseudotuberculosis 1014102 Y. pseudotuberculosis(99.4%)
2 YP O:3
14 Y. pseudotuberculosis 1014102 Y. pseudotuberculosis(99.4%)
2 YP O:3
15 Y. kristensenii 1154503 Y. kristensenii (99.4%)16 Y. kristensenii 1154501 Y. kristensenii (92.3%)17 Y. aldovae 0314712 Identification not
valid18 Y. aleksiciae 1114703 Y. kristensenii (73.5%)19 Y. enterocolitica 1015502 Y. enterocolitica
(69.8%)4 YE O:3
20 Y. enterocolitica 0015502 Y. enterocolitica(57.9%)
4 YE O:3
a NT = not typeable.
153T. Niskanen et al. / International Journal of Food Microbiology 129 (2009) 150–156
Maths, Sint–Martens–Latem, Belgium) as tiff files. The patterns werenormalized based on the mobility of the standards, and a similaritymatrix was created. The similarity between all pairs was expressedby Dice coefficient correlation, and UPGMA (unweighed pair-groupmethod using arithmetic averages) clustering was used for the con-struction of the dendrogram presented in Fig. 3. Based on the useof internal controls in the database, pattern optimization and bandposition tolerance of 0.6 and 1.8, respectively, were allowed.
Fig. 1. Comparison of the LPS biosynthetic gene clusters of strain Å125 KOH2 and Yersinia en6-dideoxy-D-xylo-hex-4-ulopyranose.
3. Results and discussion
3.1. Biochemical tests
The reactions from the API 20E kit and six additional tests wereused to differentiate the 20 sucrose-negative Yersinia strains, i.e.,raffinose fermentationwas used for biotyping of Y. pseudotuberculosis,and esculin hydrolysis, tween-esterase activity, salicin, trehalose andxylose fermentation, for biotyping of Y. enterocolitica (Table 2). AllY. pseudotuberculosis strains (#4–14)were identified correctlywith API20E when the strips were incubated at 25 °C for 20–24 h. The threepresumptive Y. pseudotuberculosis strains (#1–3) were identified asY. pseudotuberculosis with a likelihood of over 98% (Table 3); how-ever, the likelihood was slightly higher among the true Y. pseudotu-berculosis strains (#4–14). The remaining sucrose-negative Yersiniastrains (#15–20) were not identified as Y. pseudotuberculosis with API20E. Using esculin as an additional test, Y. pseudotuberculosis strains(#4–14) could be differentiated from sucrose-negative strains (#15–20and#1–3) (Table 2). The esculin test,which is included in the biotypingscheme of Y. enterocolitica, is widely used to differentiate potentialpathogenic Y. enterocolitica from non-pathogenic Y. enterocolitica.
3.2. Bio- and serotyping
Of Y. pseudotuberculosis strains (#4–14),most belonged to biotype 1(melibiose+/citrate−/raffinose−). The Y. pseudotuberculosis strains iso-lated from Finnish pigs (#11–14) and one strain from lettuce belongedto biotype 2 (melibiose−/citrate−/raffinose−). Y. pseudotuberculosis-likestrains (#1–3) belonged to biotype 2 (Table 3). Using commercial anti-O:1–6 antisera, strains (#1–3) and Y. pseudotuberculosis referencestrains (#5,7, and 8) could not be serotyped. Serotyping performed byslide agglutination is sometimes unable to type Y. pseudotuberculosisstrains, especially rough ones. An alternative way to identify Y.pseudotuberculosis serotypes is to use O-antigen gene cluster-specificmultiplex PCR, which has been shown to correlate well with classicalserotyping (Bogdanovich et al., 2003). Using this method strains #1–3could not be serotyped, but strain #4 was further subtyped to O:2a.
3.3. The O-antigen gene cluster
To elucidate why O-genotyping failed, the locus between the hemHand gsk genes carrying the O-antigen gene cluster in Y. pseudotubercu-losis was sequenced from presumptive Y. pseudotuberculosis strain #2(Å125 KOH2). This strain represented strains #1–3 since all hadindistinguishable EcoRV restriction patterns for the PCR-amplifiedhemH-gsk fragment. Altogether a sequence of 13970 bp was deter-mined, and a summary of the analysis is shown in Fig. 1. We were able
terocolitica O:3 (YeO3) between the hemH and gsk genes. Sugp, UDP-2-acetamido-2,

Table 4Predicted gene products of the hemh-gsk locus of Yersinia strain Å125 KOH2
Protein name Size (aa) Putative function Identity % Organism, gene product Accession number
Wzx 421 Flippase, O-unit translocator 72 Y. enterocolitica O:3, Wzx CAA87698WbcH 286 Glycosyl (putative fucosyl) transferase 51 Y. enterocolitica O:8, WbcH AAC60771WbcG 252 Glycosyltransferase 42 E. coli O86, WbnJ AAV80758Gmd 372 GDP-mannose-4,6-dehydratase 85 Y. mollaretii, Gmd ZP_00826187Fcl 321 Fucosyl synthase 79 Y. mollaretii, Fcl ZP_00826188ManC 468 mannose-1-phosphate guanylyltransferase 74 Y. pseudotuberculosis, ManC YP_069552ManB 458 phosphomannomutase 73 Y. pseudotuberculosis, ManB YP_001721900WbcO 341 UDP-N-acetylmuramyl pentapeptide phosphotransferase 87 Y. enterocolitica O:3, WbcO CAA87703WbcP 637 UDP-N-acetylglucosamine-4,6-reductase 92 Y. enterocolitica O:3, WbcP CAA87704WbcQ 338 N-acetylgalactosaminyltransferase 89 Y. frederiksenii, WbcQ ZP_00829797Gne 337 UDP-N-acetylglucosamine 4-epimerase 90 Y. kristensenii, Gne ABA86989
Table 5Characteristics of sucrose-negative Yersinia strains
Strain#
Species Pathogenicity tests Ribotype PFGE
inv ail virF CRMOX PYZ HindIII/EcoI NotI
1 Yersinia spp − − − − + Yr1 Y12 Yersinia spp − − − − + Yr1 Y13 Yersinia spp − − − − + Yr1 Y24 Y. pseudotuberculosis + − + + − YPr1 YP15 Y. pseudotuberculosis + − − − − YPr2 YP26 Y. pseudotuberculosis + − − − − YPr3 YP37 Y. pseudotuberculosis + − − − − YPr4 YP48 Y. pseudotuberculosis + − − − − YPr5 YP59 Y. pseudotuberculosis + − + + − YPr6 YP610 Y. pseudotuberculosis + − + + − YPr7 YP711 Y. pseudotuberculosis + − + + − YPr8 YP812 Y. pseudotuberculosis + − + + − YPr9 YP813 Y. pseudotuberculosis + − + + − YPr10 YP814 Y. pseudotuberculosis + − + + − YPr11 YP815 Y. kristensenii − − − − + YKr1 YK116 Y. kristensenii − − − − + YKr2 YK117 Y. aldovae − − − − + YAr1 YA118 Y. aleksiciae − − − − + YArI1 YAI119 Y. enterocolitica − + + + − YEr1 YE120 Y. enterocolitica − + − − − YEr1 YE2
154 T. Niskanen et al. / International Journal of Food Microbiology 129 (2009) 150–156
to identify by sequence similarity 11 genes between the hemH and gskgenes and these were named wzx, wbcH, wbcG, gmd, fcl, manC, manB,wbcO,wbcP,wbcQ and gne as illustrated in the Fig.1. Comparison of thenucleotide sequence against the bacterial nucleotide sequences in thedatabases revealed that the outer core gene cluster of Y. enterocoliticaO:3was relatedmost closely to the Å125KOH2 gene cluster, such that acentral gene block (wbcK-wbcN) in the O:3 outer core gene cluster wasreplaced in Å125 KOH2 by the mannose and fucose biosynthetic block(wbcG-manB) (Fig.1). The homologousflanking regions shared 74% and82% sequence identity between theÅ125KOH2 and the Y. enterocoliticaO:3 gene clusters. The gmd-manC block located in the middle of theÅ125 KOH2 gene cluster is very common in Enterobacteriaceae, and theclosest blocks are present in several Yersinia species, including Y. pestisY. pseudotuberculosis and Y. mollaretii, but also in Aeromonas, Escheri-chia and Salmonella. Since we could not identify the O-antigenpolymerase and chain length determinant genes (wzy and wzz,respectively) in the gene cluster, it is possible that the strain expresses,similar to Y. enterocolitica O:3 (Skurnik et al., 1995, 1999), a non-polymerized outer core structure. Indeed, in silver-stained polyacry-lamide gel electrophoresis analysis of the LPS, we could only identify asingle LPS band corresponding to a migration of LPS moleculescontaining lipid A plus full core oligosaccharide; no ladder-like bandscharacteristic for O-polysaccharide were visible (data not shown). Thegenetic setup of the Å125 KOH2 gene cluster suggests that the strainexpresses a tetrasaccharide consisting of Sugp (2-acetamido-2,6-dideoxy-D-xylo-hex-4-ulopyranose), N-acetylgalactosamine, fucoseand mannose. Chemical and structural studies on the LPS are neededto confirm this hypothesis.
The proteins showing closest similarity to Å125 KOH2 proteins arelisted in Table 4. Except for the ca. 70% similarity of the gmd-manBblock to those present in some of the 21 known Y. pseudotuberculosisserotypes (Bogdanovich et al., 2003), no similarities were detected.This explains why the strains were not typeable by the O-antigen-specific multiplex PCR. At protein level, the right flanking blockgenes showed interesting homologues in addition to those listed inTable 4. WbcO homologues with N50% identity were present inY. enterocolitica, Y. frederiksenii, several pseudomonads and E. coli.WbcP homologues with N58% identity were present in Y. enterocoli-tica, Y. frederiksenii, Plesiomonas, Shigella, E. coli, Schewanella, Photo-bacterium, Photorhabdus, Moritella and Aeromonas strains, and, veryinterestingly, also in Neisseriae involved in pilin glycosylation. Onthe other hand, WbcQ homologues were more rare, and those with85–89% identity were present in Y. enterocolitica, Y. frederiksenii andY. kristensenii, a few others were b50% identical. Gene homologueswith N85% identity were present in Y. enterocolitica, Y. frederiksenii andY. kristensenii, and a vast number of homologues with 60–65% identitywere widely distributed among bacteria.
3.4. Pathogenicity markers
The pathogenicity of the sucrose-negative strains was studied bygenotypic and phenotypic tests (Table 5). All control Y. pseudotuber-
culosis strains were shown to be inv-positive, while the rest of thesucrose-negative strains, including presumptive Y. pseudotuberculosisstrains (#1–3), were inv-negative. Interestingly, all inv-positive strainswere esculin-positive, whereas all inv-negative strains were esculin-negative. All inv-positive strains and both of the sucrose-negativeY. enterocolitica strains of pathogenic biotype 4 were pyrazinamidase-negative. The pyrazinamidase activity test is included in the biotypingscheme of Y. enterocolitica to differentiate between non-pathogenicand potential pathogenic strains. Negative pyrazinamidase activityhas been shown to correlate well with potential pathogenicity ofY. pseudotuberculosis and Y. enterocolitica. For full pathogenicity,pathogenic Yersinia species need the virulence plasmid, pYV. Allwild-type Y. pseudotuberculosis strains (strains #4, and 9–14) werevirF-positive and showed calcium-dependence and Congo-red uptakewith CR-MOX agar while Y. pseudotuberculosis reference strains #5–8and presumptive Y. pseudotuberculosis strains (#1–3) were virF- andCR-MOX-negative (Table 5). It must be noted, however, that thereference strains may have lost the pYV during repeated cultures inthe laboratory.
3.5. Genotyping
The PFGE patterns of the Y. pseudotuberculosis strains obtainedwithNotI-digested DNA showed low similarity to each other (Fig. 2). This isin accordance with the results of Iteman et al. (1995), who reported ahigh polymorphism in Y. pseudotuberculosis. The three esculin- andinv-negative presumptive Y. pseudotuberculosis strains (#1–3) showedhigh similarity to each other, but differed clearly from the othersucrose-negative strains (#4–20). Fig. 3 presents the results of the

Fig. 2. PFGE profiles with NotI enzyme of 17 sucrose-negative Yersinia strains. Yersinia pseudotuberculosis-like strains (1–3); Y. pseudotuberculosis (4–11); Yersinia kristensenii (12–13);Yersinia aldovae (14); Yersinia aleksiciae (15); Yersinia enterocolitica (16–17); M, low range PFGE marker 1.
155T. Niskanen et al. / International Journal of Food Microbiology 129 (2009) 150–156
numerical analysis of the sucrose-negative strains based on bothEcoRI- and HindIII-deduced ribopatterns. Presumptive Y. pseudotuber-culosis strains (#1–3) shared the same ribopatterns, differing clearlyfrom the other sucrose-negative strains (#4–20) and especially fromY. pseudotuberculosis strains (#4–14) and pathogenic sucrose-negativeY. enterocolitica strains (#19 and 20). Environmental Y. pseudotubercu-losis strain #4 formed its own ribotype and clustered into Y. pseudo-tuberculosis ribogroup. Because the presumptive Y. pseudotuberculosis
Fig. 3. Numerical analysis based on both EcoRI and HindIII deduced ribopatterns presentedb = Collection de l'Institut Pasteur, c = Deutsche Sammlung von Mikroorganismen und Ze
strains (#1–3) were inv- and virF-negative, and their PFGE andribotyping patterns formed own cluster they were tentatively namednon-pathogenic Y. pseudotuberculosis-like strains.
4. Conclusions
This study shows that identification of environmental presumptiveY. pseudotuberculosis strains can be incorrect with biochemical tests.
as a dendrogram. Superscripts: T = type strain, a = American Type Culture Collection,llkulturen GmbH.

156 T. Niskanen et al. / International Journal of Food Microbiology 129 (2009) 150–156
Especially strains that cannot be serotyped should be furthercharacterized to correctly identify them and to determine theirpathogenic potential. One easy and rapid way is to use PCR targetingthe inv gene in the chromosome of Y. pseudotuberculosis and the virFgene on the virulence plasmid.
Acknowledgements
Johanna Björkroth is grateful to the Academy of Finland ELVIRAprogramme (project no. 117921) for financial support. This researchwas partly conducted at the Finnish Centre of Excellence, MicrobialFood Safety Research.
References
Ausubel, F.M., Brent, R., Kingston, R.E., Moore, O.D., Seidman, J.G., Smith, J.A., Struhl, K.,1987. (Eds.) Current protocols in molecular biology. John Wiley and Sons, Inc.:New York.
Björkroth, J., Korkeala, H., 1996. Evaluation of Lactobacillus sake contamination in vacuum-packed sliced cooked meat products by ribotyping. Journal of Food Protection 59,398–401.
Blumberg, H.M., Kiehlbauch, J.A., Wachsmuth, I.K., 1991. Molecular epidemiology of Yersi-nia enterocoliticaO:3 infections: use of chromosomal DNA restriction fragment lengthpolymorphisms of rRNA genes. Journal of Clinical Microbiology 29, 2368–2374.
Bogdanovich, T., Carniel, E., Fukushima, H., Skurnik, M., 2003. Use of O-antigen genecluster-specific PCRs for the identification and O-genotyping of Yersinia pseudotu-berculosis and Yersinia pestis. Journal of Clinical Microbiology 41, 5103–5112.
EFSA, 2007. Monitoring and identification of human enteropathogenic Yersinia spp. TheEFSA Journal, 595, 1–30.
Fredriksson-Ahomaa, M., Niskanen, T., Neubauer, H., Laukkanen, R., Korkeala, H., 2002.Characterisation of sucrose-negative Yersinia enterocolitica 4/O:3 isolates recoveredfrom pig tonsils. International Journal of Food Microbiology 75, 19–25.
Isberg, R.R., Voorhis, D.L., Falkow, S., 1987. Identification of invasin: a protein that allowsenteric bacteria to penetrate cultured mammalian cells. Cell 50, 769–778.
ISO, 2003. Horizontal method for detection of presumptive pathogenic Yersiniaenterocolitica. ISO 10273. International Organisation for Standardisation, Geneva,Switzerland.
Iteman, I., Najdenski, H., Carniel, E., 1995. High genomic polymorphism in Yersiniapseudotuberculosis. In: Ravagnan, G., Chiesa, C. (Eds.), Yersiniosis: present andfuture, vol. 13. Karger, Basel, Switzerland, pp. 106–111.
Jalava, K., Hakkinen, M., Valkonen, M., Nakari, U.M., Palo, T., Hallanvuo, S., Ollgren, J.,Siitonen, A., Nuorti, P., 2006. An outbreak of gastrointestinal illness and erythemanodosum from grated carrots contaminated with Yersinia pseudotuberculosis.Journal of Infectious Diseases 194, 1209–1216.
Jalava, K., Hallanvuo, S., Nakari, U.M., Ruutu, P., Kela, E., Heinasmaki, T., Siitonen, A.,Nuorti, J.P., 2004. Multiple outbreaks of Yersinia pseudotuberculosis infections inFinland. Journal of Clinical Microbiology 42, 2789–2791.
Kandolo, K., Wauters, G., 1985. Pyrazinamidase activity in Yersinia enterocolitica andrelated organisms. Journal of Clinical Microbiology 21, 980–982.
Kaneko, S., Ishizaki, N., Kokubo, Y., 1995. Detection of pathogenic Yersinia enterocoliticaand Yersinia pseudotuberculosis from pork using the polymerase chain reaction.Contributions to Microbiology and Immunology 13, 153–155.
Marra, A., Isberg, R.R., 1997. Invasin-dependent and invasin-independent pathways fortranslocation of Yersinia pseudotuberculosis across the Peyer's patch intestinalepithelium. Infection and Immunity 65, 3412–3421.
Nagano, T., Kiyohara, T., Suzuki, K., Tsubokura, M., Otsuki, K., 1997. Identification ofpathogenic strains within serogroups of Yersinia pseudotuberculosis and thepresence of non-pathogenic strains isolated from animals and the environment.The Journal of Veterinary Medical Science 59, 153–158.
Nakajima, H., Inoue, M., Mori, T., Itoh, K.I., Arakawa, E., Watanabe, H., 1992. Detectionand identification of Yersinia pseudotuberculosis and pathogenic Yersinia enteroco-litica by an improved polymerase chain reaction method. Journal of ClinicalMicrobiology 30, 2484–2486.
Neubauer, H., Sauer, T., Becker, H., Aleksic, S., Meyer, H., 1998. Comparison of systems foridentification and differentiation of species within the genus Yersinia. Journal ofClinical Microbiology 36, 3366–3368.
Niskanen, T., Fredriksson-Ahomaa, M., Korkeala, H., 2002. Yersinia pseudotuberculosiswith limited genetic diversity is a common finding in tonsils of fattening pigs.Journal of Food Protection 65, 540–545.
Niskanen, T.,Waldenström, J., Fredriksson-Ahomaa,M., Olsen, B., Korkeala, H., 2003. VirFpositive Yersinia enterocolitica and Yersinia pseudotuberculosis found in migratorybirds in Sweden. Applied and Environmental Microbiology 8, 4670–4675.
Nuorti, J.P., Niskanen, T., Hallanvuo, S., Mikkola, J., Kela, E., Hatakka, M., Fredriksson-Ahomaa, M., Lyytikainen, O., Siitonen, A., Korkeala, H., Ruutu, P., 2004. Awidespreadoutbreak of Yersinia pseudotuberculosis O:3 infection from iceberg lettuce. TheJournal of Infectious Disease 189, 766–774.
Pitcher, D., Saunders, N., Owen, R., 1989. Rapid extraction of bacterial genomic DNAwithguenidium thiocyanate. Letter in Applied Microbiology 8, 151–156.
Riley, G., Toma, S., 1989. Detection of pathogenic Yersinia enterocolitica by using Congored-magnesium oxalate agar medium. Journal of Clinical Microbiology 27, 213–214.
Rimhanen-Finne, R., Niskanen, T.,Hallanvuo, S.,Makary, P.,Haukka, K., Pajunen, S., Siitonen,A., Ristolainen, R., Pöyry, H., Ollgren, J., Kuusi, M., 2008. Yersinia pseudotuberculosiscausing a large outbreak associated with carrots in Finland, 2006. Epidemiology andInfection 4, 1–6.
Simonet, M., Falkow, S., 1992. Invasin expression in Yersinia pseudotuberculosis. Infectionand Immunity 60, 4414–4417.
Skurnik, M., 1999. Molecular genetics of Yersinia lipopolysaccharide. In: Goldberg, J. (Ed.),Genetics of Bacterial Polysaccharides. CRC Press, Boca Raton, FL, pp. 23–51.
Skurnik, M., 2004. Lipopolysaccharides of Yersinia. In: Carniel, E., Hinnebusch, B.J.(Eds.), Yersinia: molecular and cellular biology. Horizon Bioscience,Wymondham,pp. 215–241.
Skurnik, M., Venho, R., Toivanen, P., Al-Hendy, A., 1995. A novel locus of Yersiniaenterocolitica serotype O:3 involved in lipopolysaccharide outer core biosynthesis.Molecular Microbiology 17, 575–594.
Skurnik, M., Venho, R., Bengoechea, J.A., Moriyón, I., 1999. The lipopolysaccharide outercore of Yersinia enterocolitica serotype O:3 is required for virulence and plays a rolein outer membrane integrity. Molecular Microbiology 31, 1443–1462.
Tsubokura, M., Aleksic, S., 1995. A simplified antigenic scheme for serotyping of Yersiniapseudotuberculosis: phenotypic characterization of reference strains and prepara-tion of O and H factor sera. Contributions to Microbiology and Immunology 13,99–105.
Vincent, P., Leclercq, A., Martin, L., Duez, J.M., Simonet, M., Carniel, E., 2008. Suddenonset of pseudotuberculosis in Humans, France, 2004–05. Emerging InfectiousDiseases 14, 1119–1122.
Voskressenskaya, E., Leclercq, A., Tseneva, G., Carniel, E., 2005. Evaluation of ribotypingas a tool for molecular typing of Yersinia pseudotuberculosis strains of worldwideorigin. Journal of Clinical Microbiology 43, 6155–6160.
Wauters, G., Kandolo, K., Janssens, M., 1987. Revised biogrouping scheme of Yersiniaenterocolitica. Contributions to Microbiology and Immunology 9, 14–21.





















![Quantitative Trait Locus (QTL) Mapping Reveals a Role for …summit.sfu.ca/system/files/iritems1/13352/journal.pone... · 2020. 9. 5. · Yersinia pseudotuberculosis [29]. In addition](https://img.dokumen.tips/doc/110x75/6068f314f94ad21c506e92d2/quantitative-trait-locus-qtl-mapping-reveals-a-role-for-2020-9-5-yersinia.jpg)







