Embed Size (px)
Citation preview

Chapter 9: Population Growth
1

2
Population growth and regulation
• Population sizes in nature fluctuate,
are not constant
– natality and mortality sensitive to changes in
climate and food supply
– degree of variation dependent on
• magnitude of flux in environmental conditions
• inherent stability of the population
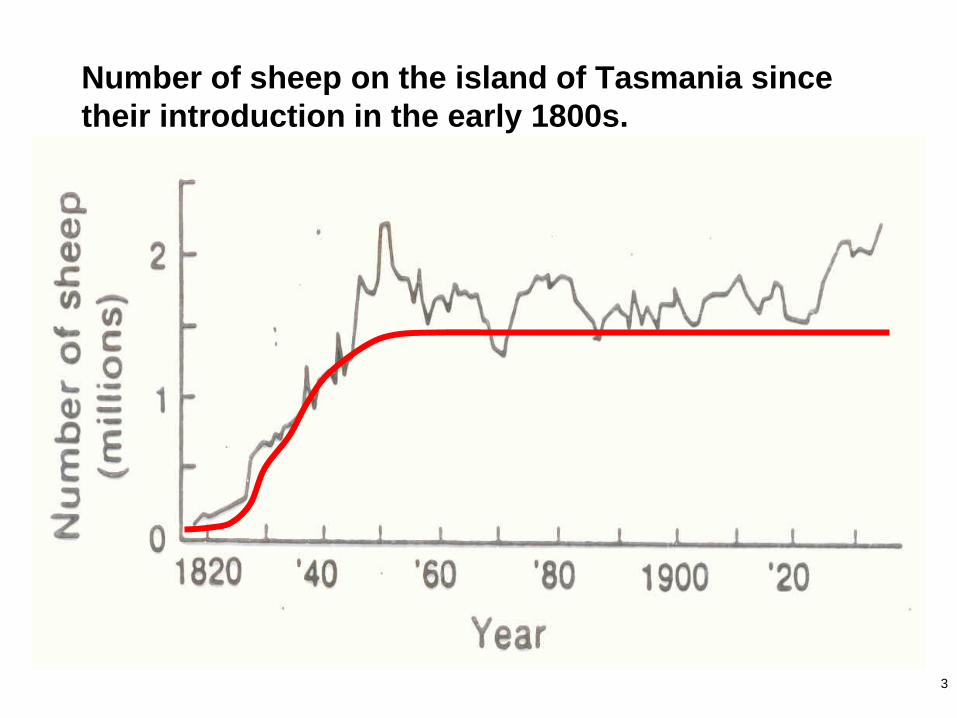
3
Number of sheep on the island of Tasmania since
their introduction in the early 1800s.

4
Population growth and regulation
• Population sizes in nature fluctuate,
are not constant
– short lived organisms are more sensitive to
environmental changes than long-lived
species
– limiting factor usually responsible for
population increases and decreases and is
often seasonal
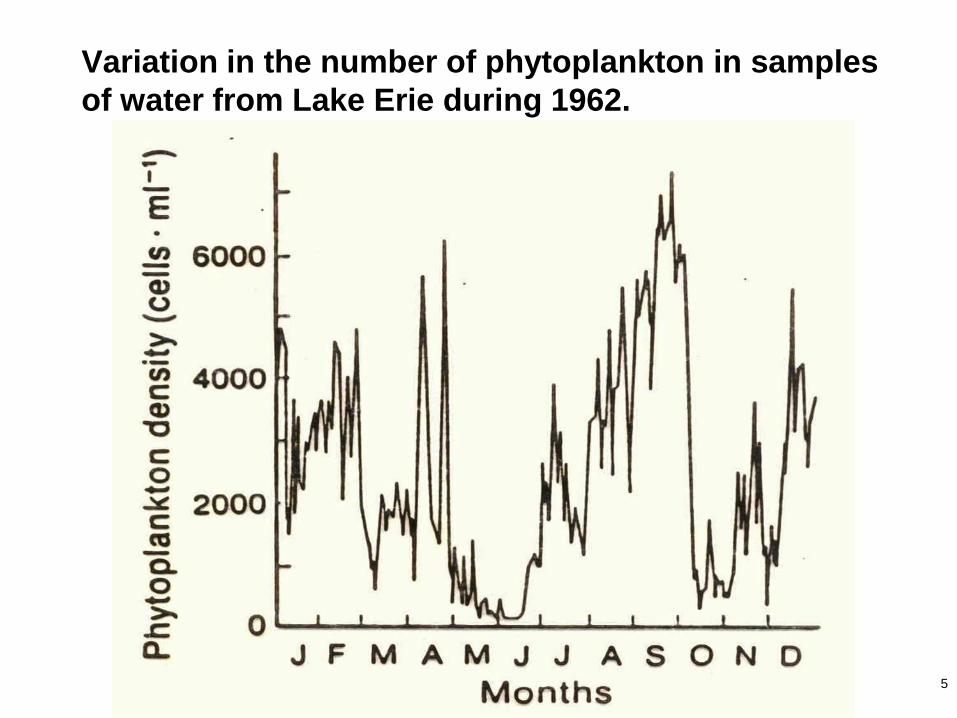
5
Variation in the number of phytoplankton in samples
of water from Lake Erie during 1962.

6
Influences on population growth rates
• Stabilizing factors
– maintain population at equilibrium level
– density dependent
• Non-stabilizing factors
– no equilibrium level maintained
– density independent

7
Stabilizing factors
• Maintain population at equilibrium size (K)
• density when growth rate > K
• density when growth rate < K
• termed density dependent factors
• Examples of density dependent factors

8
Hypothetical birth and death rates in a regulated
population.

9
Growth rate of a hypothetical population with respect
to increasing density.

10
Density approaching equilibrium in a hypothetical
population.

11
Fig. 9.1 (p. 144): Inherent growth potential of a
population; comparison of different reproductive rates
(starting N=10).
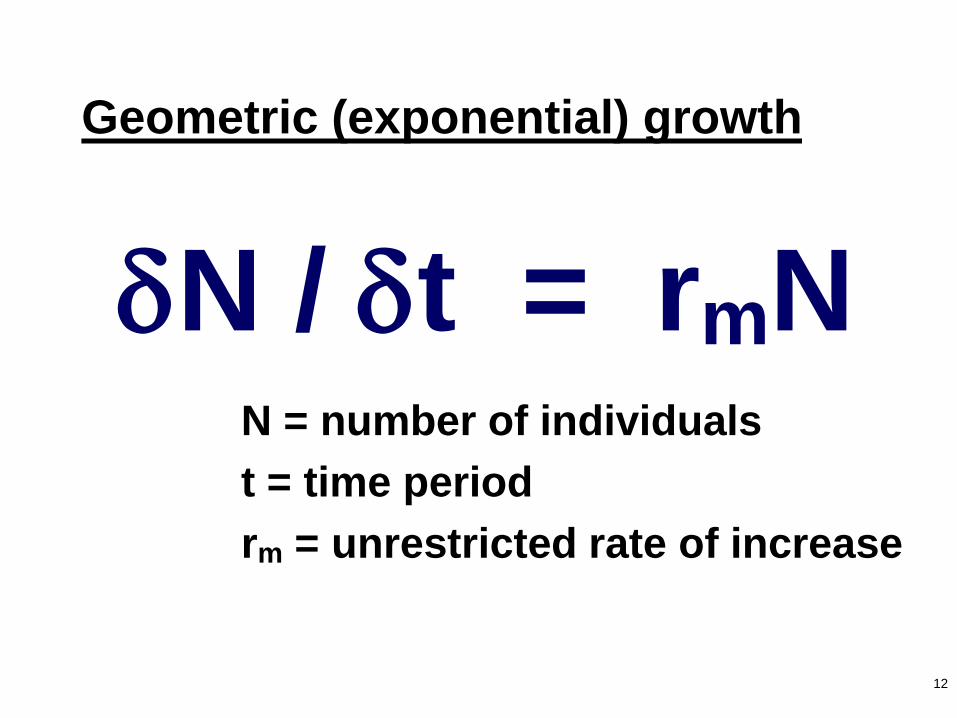
12
Geometric (exponential) growth
N / t = rmNN = number of individuals
t = time period
rm = unrestricted rate of increase

13
Geometric population growth
t N
0 2
1 4
2 8
3 16
4 32
5 64
6 128
7 256
8 512
9 1024
10 2048
11 4096
12 8192
13 16384
14 32768
15 65536
16 131072
17 262144
18 524288
19 1048576
20 2097152
0
500000
1000000
1500000
2000000
2500000
1 3 5 7 9 11 13 15 17 19 21
Nu
mb
er
in p
op
ula
tio
n
Time (Generation)
Plotted on arithmetic scale

14
Geometric population growth
t N
0 2
1 4
2 8
3 16
4 32
5 64
6 128
7 256
8 512
9 1024
10 2048
11 4096
12 8192
13 16384
14 32768
15 65536
16 131072
17 262144
18 524288
19 1048576
20 2097152
1
10
100
1000
10000
100000
1000000
10000000
1 3 5 7 9 11 13 15 17 19 21
Nu
mb
er
in p
op
ula
tio
n
Time (Generation)
Plotted on log scale

15
Non-stabilizing factors
• Act upon individuals in the population
without regard to population density
• Do not maintain population at particular
equilibrium size
• termed density independent factors
• Examples of density independent factors

16
Model for density dependent
population growth
• For any finite resource, there is an upper
limit to the number of individuals that can
utilize the resource
• Density dependent population growth is
based on
– geometric limitations
– availability of energy supply
– behavior (territoriality)

17
Logistic equation
• Pierre-François Verhulst (c 1835)
• Logistic (sigmoidal) growth curve
– population growth decreases as the
population size approaches the carrying
capacity (K) of the environment
• food, nutrients
• space

18
Fig. 9.4 (p. 146): Geometric (unlimited resources) versus
logistic (limited resources) growth of a population.

19
Logistic equation for growth of a
single population
N / t = rN [(K-N)/K]N = number of individuals
t = time period
r = unrestricted rate of increase
K = carrying capacity (upper asymptote)
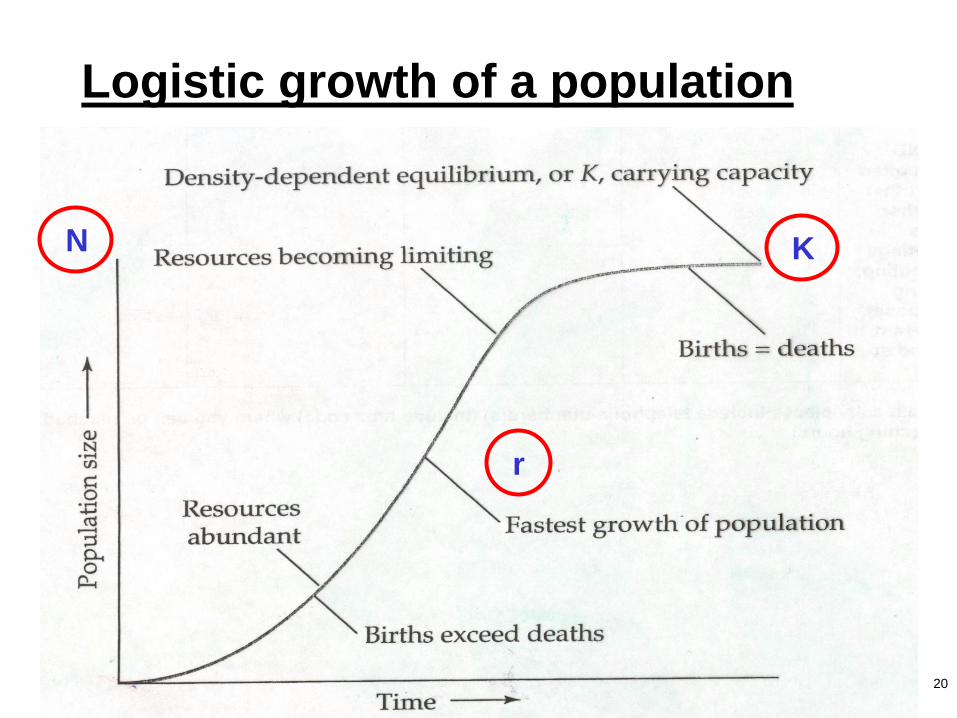
20
Logistic growth of a population
N
r
K

21
(K-N)/K– K: upper asymptote or carrying capacity
– K > N: allows for further population growth
– K = N: no growth; population at equilibrium
size
– K < N: results in decreased growth rate
Logistic equation for growth of a
single population

22
• Population increases determined by
– N: number of individuals present
– r: innate capacity for increase
– (K-N)/K: proportion of K not yet realized
Logistic equation for growth of a
single population
K = 100, N = 2: (K-N)/K = 98/100 = 0.98
δN/ δt = rN(0.98)
K = 100, N = 98: (K-N)/K = 2/100 = 0.02
δN/ δt = rN(0.02)

23
Logistic growth of a population
N
r
K

24

25
• (K-N)/K acts as negative feedback to
population growth
– when N is small
• (K-N)/K ~ 1
• r is maximal
• → exponential growth in population
Logistic equation for growth of a
single population

26
• (K-N)/K acts as negative feedback to
population growth
– as N increases
• (K-N)/K decreases
• ↓ actual rate of increase (ra)
Logistic equation for growth of a
single population

27
• (K-N)/K acts as negative feedback to
population growth
– as N approaches K
• (K-N)/K close to zero
• ra is minimal
• greatly decreased rate of population
growth
Logistic equation for growth of a
single population

28
Fig. 9.2 (p. 144): Net reproductive rate (r0) as a linear
function of population density (N) at time t.

29
Laboratory study of the logistic model
• Carlson (1913): Saccharomyces cervisiae
• Gause (1934): Paramecium caudatum
• Pearl (1927): Drosophila melanogaster
• Park et al (1964): Tribolium castaneum
• Slobodkin (1954): Daphnia obtusa
• Ayala (1968): Drosophila melanogaster
• Gause (1931): Moina macrocopa
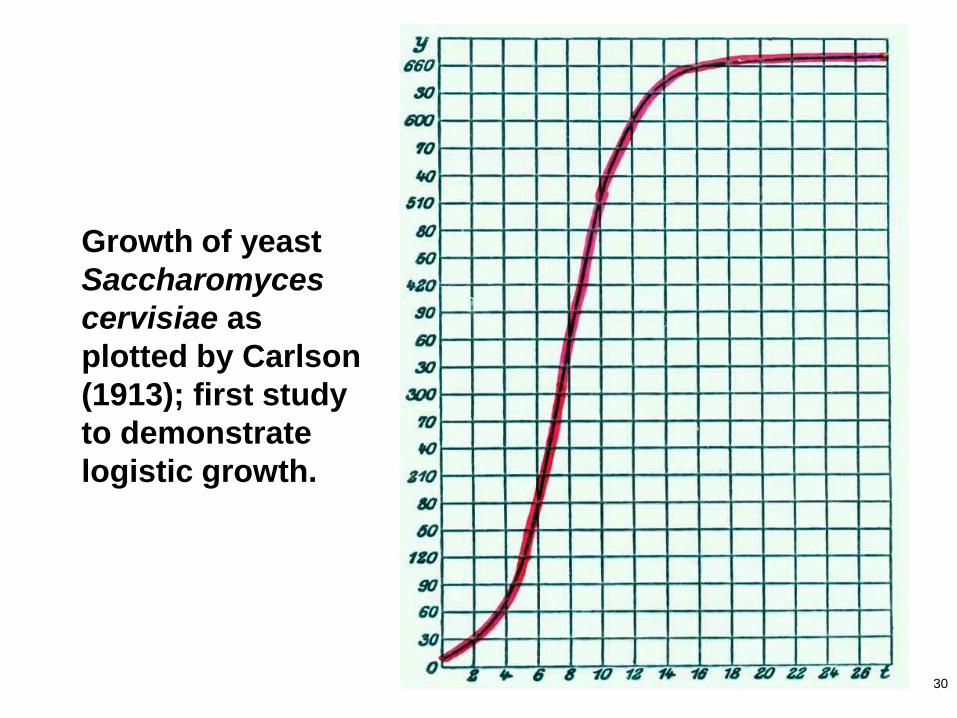
30
Growth of yeast
Saccharomyces
cervisiae as
plotted by Carlson
(1913); first study
to demonstrate
logistic growth.

31
Fig. 9.5 (p. 148): Population growth in the protozoans
Paramecium caudatum and P. aurelia (Gause 1934)

32
Fig. 9.6 (p. 149): Population growth of Drosophila
melanogaster under laboratory conditions (Pearl 1927).

Fig. 9.7 (p. 149): Population growth of two genetic
strains of the flour beetle Tribolium castaneum (Park et
al. 1964).
33

34
Relationship between K and food level in Daphnia
obtusa (Slobodkin 1954).

35
How to increase K
• Drosophila experiment by Ayala (1968)
– increase food 50% proportional increase in
young fruit flies
– population as a whole increased only 17%
– overcrowding of adult fruit flies caused
decrease in life expectancy, i.e., increased
mortality rate

36
How to increase K
• Drosophila experiment by Ayala (1968)
– increasing space increased overall population
density
– can increase K by increasing food and
space
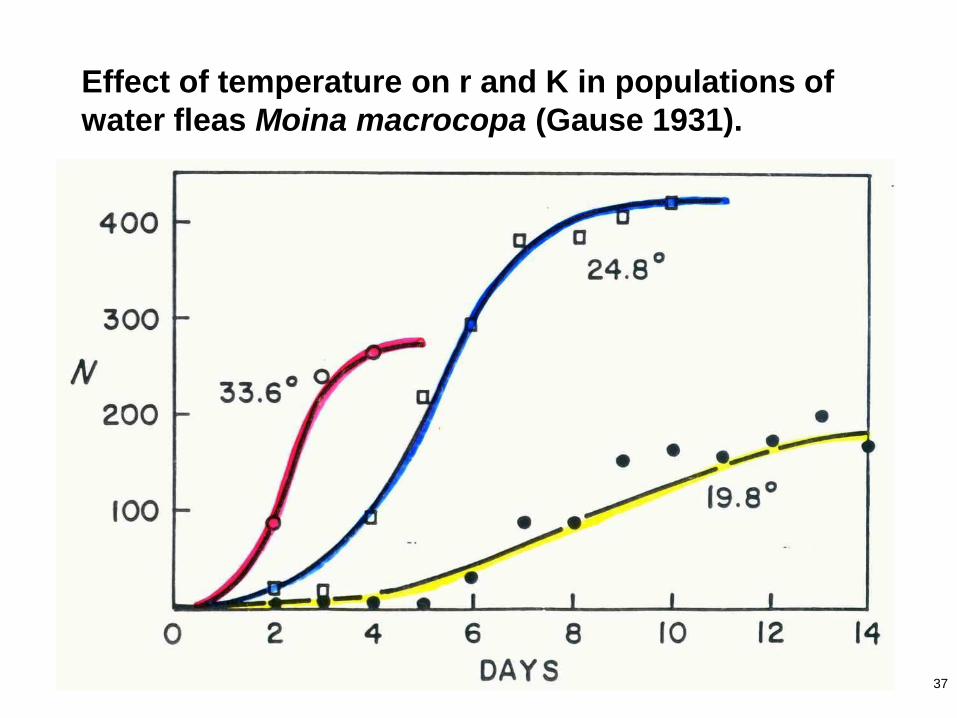
37
Effect of temperature on r and K in populations of
water fleas Moina macrocopa (Gause 1931).

38
Field study of the logistic model
• Ridgway et al. (2006): double-crested
cormorants on Lake Huron
• Saether et al. (2002): ibex in Switzerland
• Johns (2005): whooping cranes at Aransas,
Texas
• Scheffer (1951): reindeer on Pribilof Islands
• Walters et al. (1990): cladocerans in British
Columbia

39
Fig. 9.8 (p. 150): Growth in three colonies of double-
crested cormorants (Phalacrocorax auritus) on Lake
Huron, 1978-2003 (Ridgway et al. 2006).
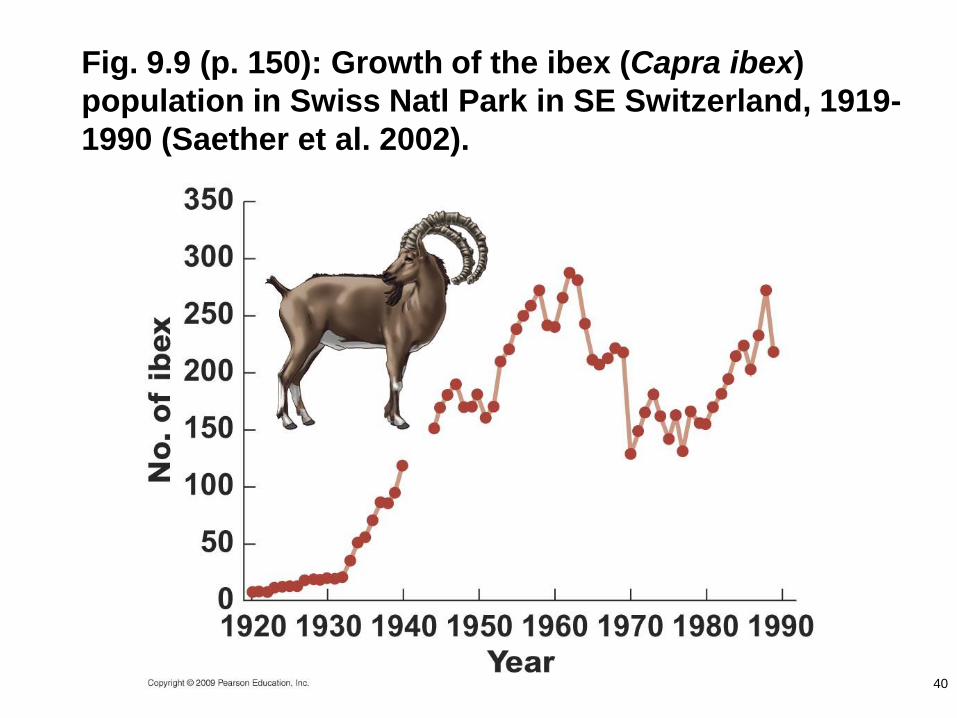
Fig. 9.9 (p. 150): Growth of the ibex (Capra ibex)
population in Swiss Natl Park in SE Switzerland, 1919-
1990 (Saether et al. 2002).
40

41
Fig. 9.10 (p. 151): Population growth in whooping cranes
(Aransas, TX) from near extinction in 1941 (Johns 2005).

42
Reindeer population growth on two Pribilof islands
following their introduction in 1911 (Scheffer 1951).

43
Fig. 9.11 (p. 151):
Population
fluctuations of
Daphnia rosea in
two British
Columbia lakes,
1980-1983
(Walters et al.
1990).
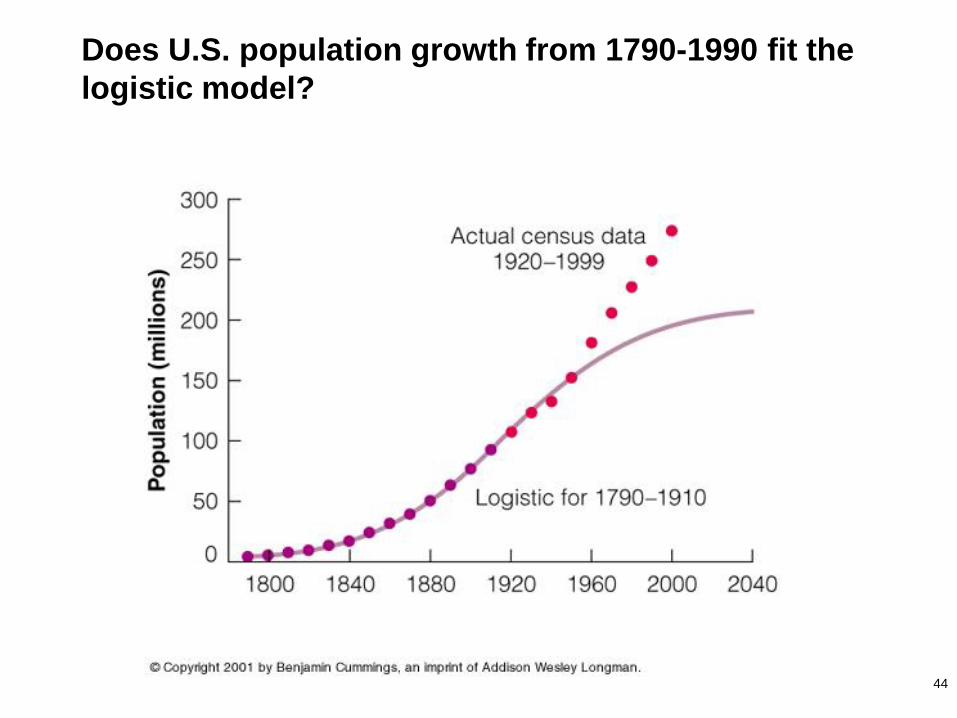
44
Does U.S. population growth from 1790-1990 fit the
logistic model?

45
Assumptions for the logistic model
• Population has a stable age distribution
– population characteristic
– stable age distribution: relative proportion of
individuals in each age class remains constant
over time
– rate of population change per age class = rate
of overall population change
– important assumption, usually not violated

46
Assumptions for the logistic model
• All individuals in the population are
equal
– all individuals use the same amount of
resources
– therefore, all individuals in the population are
assumed to be adults
– constantly violated

47
Assumptions for the logistic model
• r and K are constant and are real,
meaningful attributes of the population
– often violated
– few, if any, environments are constant over a
long period of time
– what is actually observed is some probability
distribution around K oscillations around the
asymptote driven by environmental changes

48
Fig. 9.15 (p.
154):
Population
growth in
Daphnia
magna in 50
ml of pond
water at (a)
18C and (b)
25C.

49
Assumptions for the logistic model
• A linear relationship exists between r
and N
– often violated
– when N is very small, individuals have difficulty
finding each other for mating
– r expected to increase with increasing N until
some optimal N for mating is achieved
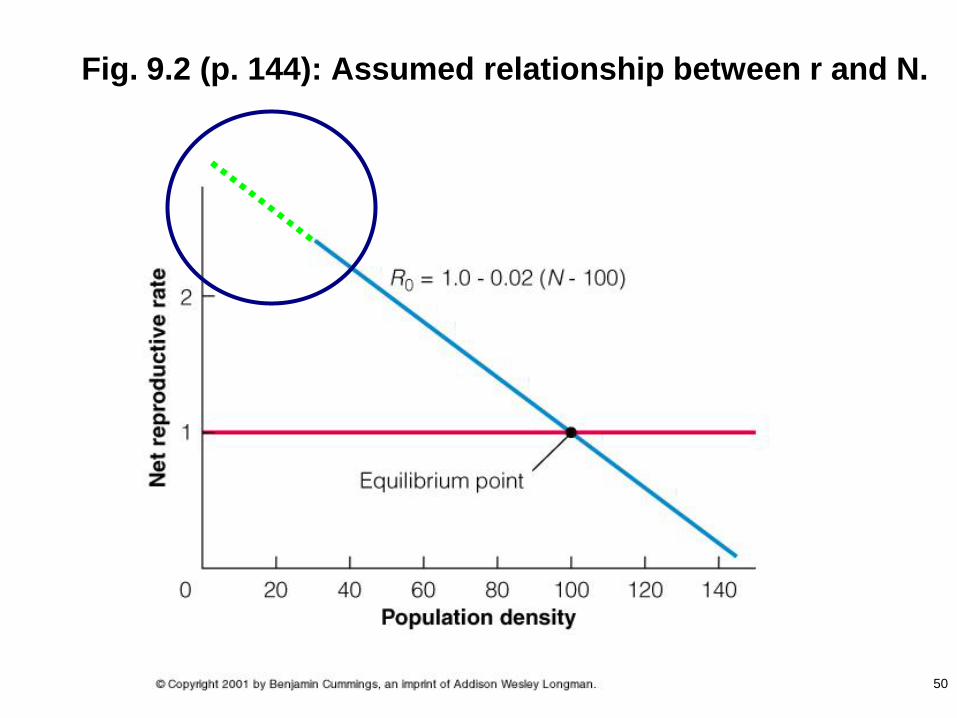
50
Fig. 9.2 (p. 144): Assumed relationship between r and N.

51
Fig. 9.2 (p. 144): More realistic view of the relationship
between r and N.

52
Assumptions for the logistic model
• No time lag is involved
– individuals coming into the population are
assumed to be immediately able to reproduce
– often violated
– necessary to figure a time lag unit (tau ) into
the logistic equation
• when (r)() is low (short lag), normal logistic curve
• when (r)() is high (long lag) transitory population
size > K oscillations
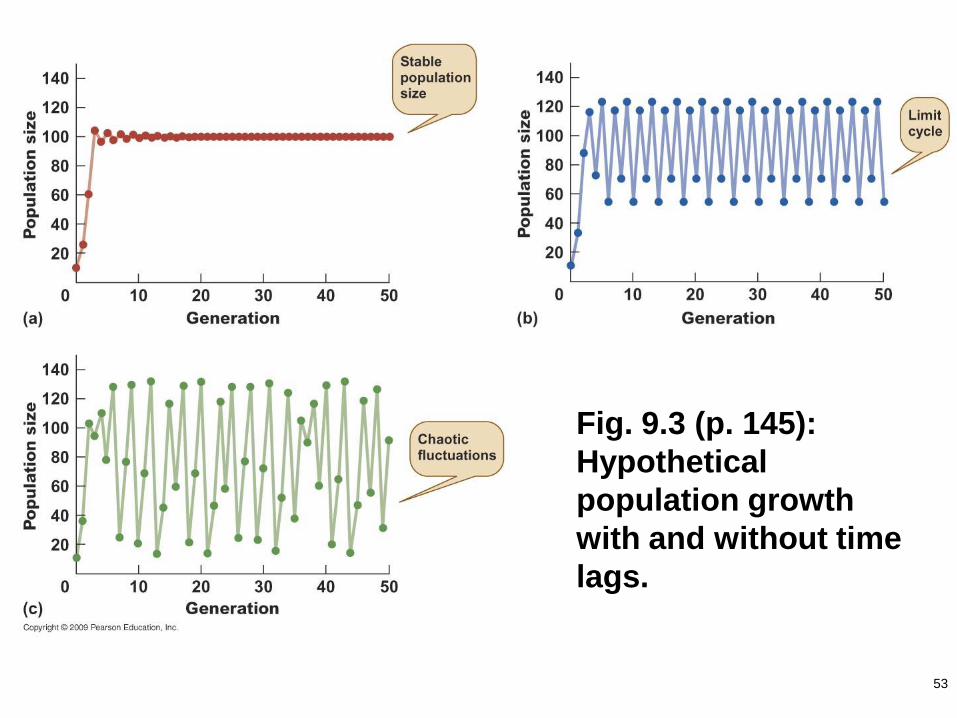
53
Fig. 9.3 (p. 145):
Hypothetical
population growth
with and without time
lags.

54
Conclusions regarding logistic
• Logistic equation fails to fit a lot of data from
the field
• Logistic model persists in ecology
because
– assumptions are well understood
– departures from the logistic model can be
attributed to violations of the assumptions
– logistic model is simple, easy to use
– no better model exists
55
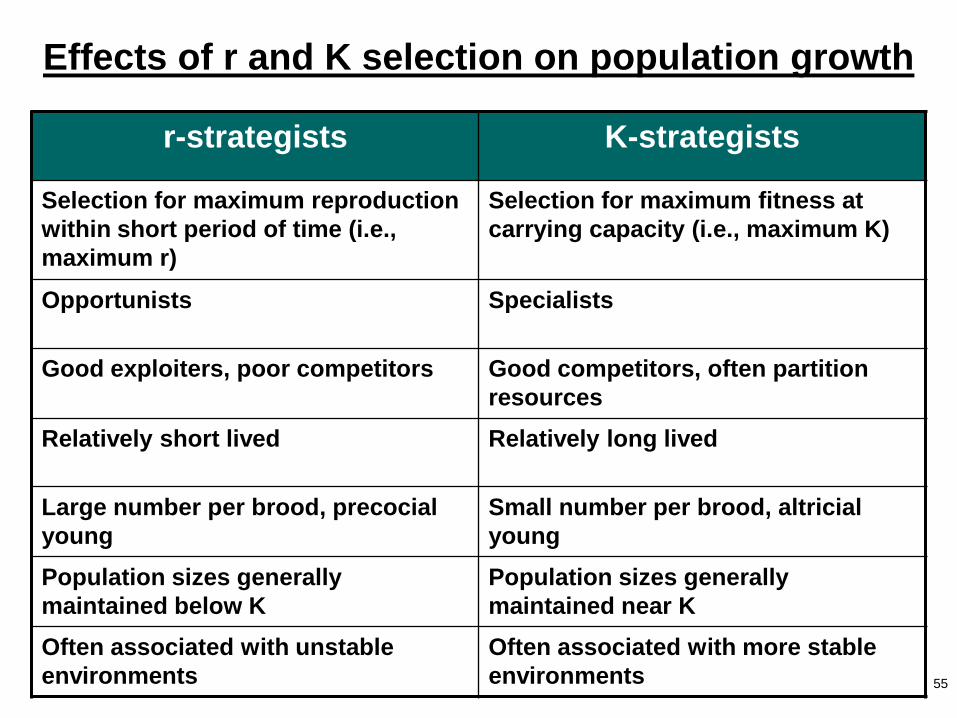
Effects of r and K selection on population growth
r-strategists K-strategists
Selection for maximum reproduction
within short period of time (i.e.,
maximum r)
Selection for maximum fitness at
carrying capacity (i.e., maximum K)
Opportunists Specialists
Good exploiters, poor competitors Good competitors, often partition
resources
Relatively short lived Relatively long lived
Large number per brood, precocial
young
Small number per brood, altricial
young
Population sizes generally
maintained below K
Population sizes generally
maintained near K
Often associated with unstable
environments
Often associated with more stable
environments


![Library System - Case Study[1]sce.uhcl.edu/helm/SWTOOLS/SSI_TAMU/Library_casestudy.pdf · Library System - Address Register - Microsoft Internet Explorer Toois Help Favorites html!](https://img.dokumen.tips/doc/110x75/5b6a5ed77f8b9af6098c003b/library-system-case-study1sceuhcleduhelmswtoolsssitamulibrary-library.jpg)
















![[PPT]PowerPoint Presentation - University of Houston–Clear …sce.uhcl.edu/yang/teaching/csci5333Fall04/XMLdatabase.ppt · Web viewFIGURE 26.7 Subset of the UNIVERSITY database](https://img.dokumen.tips/doc/110x75/5ace63957f8b9a8b1e8b7514/pptpowerpoint-presentation-university-of-houstonclear-sceuhcleduyangteachingcsci5333fall04.jpg)









