Embed Size (px)
Citation preview

Chapter 12: Intracellular sorting

Principles of intracellular sorting

Principles of intracellular sorting
Cells have many distinct compartments (What are they? What do they do?)
Specific mechanisms are required for transport of proteins in and out of each compartment.

Principles of intracellular sorting
Proteins move between compartment in 2 different ways:1) as proteins,
via pores or translocasesmay be folded or unfolded
2) within vesiclesrequiring packaging and membrane fusion

Principles of intracellular sorting
All (well, almost all) proteins begin in the cytosol
2 classes of import:1) Post-translational import:
Proteins are fully translated then sent to the correct location
2) Co-translational import: Proteins are simultaneously translated and
imported.

Principles of intracellular sorting
In the absence of specific signals, proteins remain in the cytosol
Specific signal sequences (=targeting sequences) on the protein direct them to correct place
The signal sequences are often (not always) cleaved by signal peptidases

Principles of intracellular sorting

Nuclear transport
What is the nature of the barrier between the cytosol and the inside of the nucleus (i.e., the nuclear matrix)?
What macromolecules need to get in and out of the nucleus?

Nuclear transportAccess to the nucleus is via a nuclear pore complex
Channel allows:-free diffusion of small particles -active transport of larger proteins

Nuclear transport


Nuclear transport: the proteins
Proteins that need to be transported through nuclear pores have nuclear localization signals (NLS)
NLS are short stretches of amino acids (e.g. KKKRK) found almost anywhere in the polypeptide (Why anywhere?)
NLS can be specific for export (NES) or import (NIS)
Some proteins have both NIS and NES

Nuclear transport: the receptors
Proteins that transport cargo through nuclear pores are called nuclear transport receptors
These proteins bind:-NLS of the cargo proteins -inner lining of nuclear pores
A large gene family encodes Nuclear Import Receptors and Nuclear Export Receptors



Step 1: Translation and recognition
Once a nuclear protein is made within the cytosol, it gets folded
The NIS often extends from the protein as a loop, exposed on the surface
The loop is recognized by a Nuclear Import Receptor

Step 2: Import
The receptor binds to FG-repeats (ie PHE-GLY residues) on proteins that line the nuclear pore
Receptor, with its cargo, walks along the FG-repeats to enter the nucleus
Once inside, the protein is released
The receptor protein is sent back out of the nucleus

Mitochondria and chloroplasts
What is the nature of the barrier between the cytosol and the inside of the mitochondria/ chloroplast?
How many compartments are there in these organelles?
What macromolecules need to get in and out of these organelles?
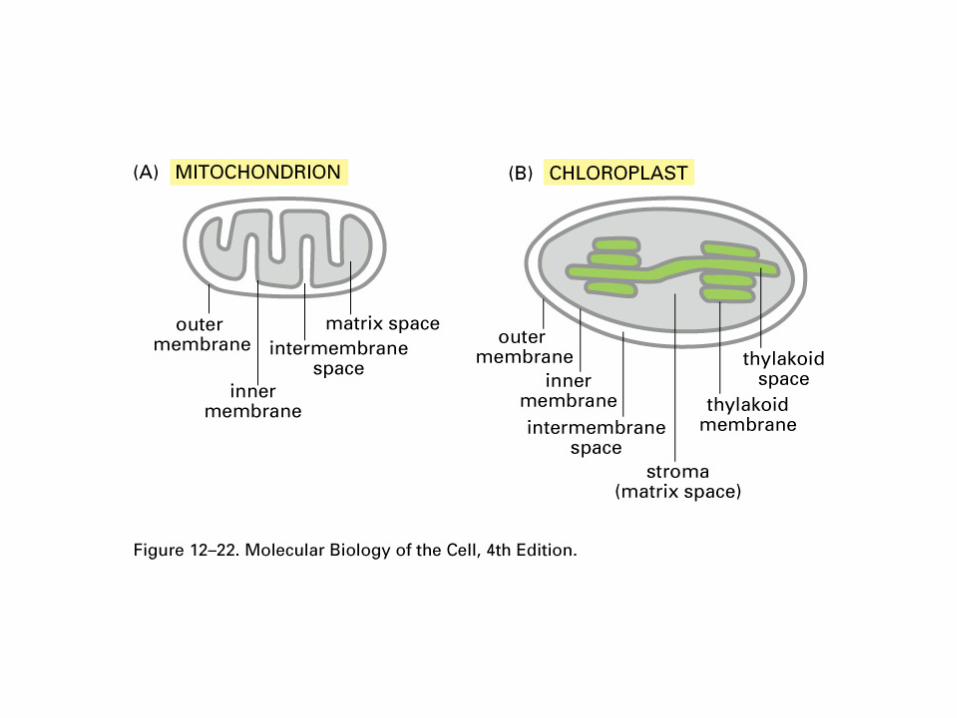

Mitochondria and chloroplastsIn contrast to the nucleus, mitochondria have:
–Extra membrane compartments–Many membrane proteins–Highly impermeable membranes
Proteins are fully synthesized in the cytosol
Targeted to mitochondria via N-terminal signal sequence
Transported into mitochondria

Step 1. Targeting proteins to mitochondria
Proteins are fully synthesized in the cytosol, but kept unfolded by chaperones (Hsp70 members)
Mitochondrial proteins have an N-terminal signal sequence that is recognized by transport machinery on the mitochondrial outer membrane
TOM (Transport across Outer Membrane) complexes include receptors and translocation channels


Step 2 Crossing the mitochondrial membranes
Outer and inner mitochondrial membranes join at contact sites, where TOM complex interacts with TIM complex
Unfolded proteins are threaded through the channels of TOM and TIM, and released into the mitochondrial matrix
Signal sequence often cleaved

Step 3 Other mitochondrial locations
Many proteins have 2nd signals that result in transport to other mitochondrial locations (e.g. inner mitochondrial membrane, inter-membrane space)


Energy requirementsATP hydrolysis: needed for action of cytosolic chaperones and mitochondrial chaperones (both hsp70 members)
Electrochemical gradient: protein import into mitochondria also requires a membrane potential

Endoplasmic Reticulum
What is the nature of the barrier between the cytosol and the inside of the ER (the ER lumen)?
What macromolecules need to get in and out of the ER?

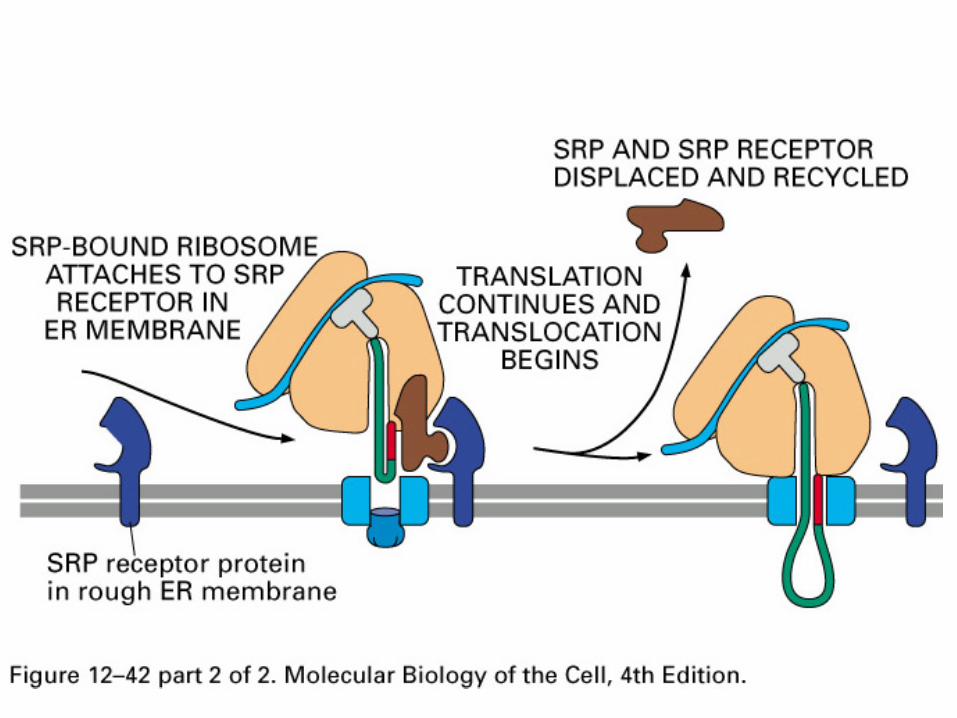

Endoplasmic Reticulum
Proteins destined for the ER are partially translated in the cytosol on free ribosomes
Once the N-terminal signal peptide is made, it is recognized by a complex of RNA and protein (signal recognition particle or SRP)
Once the SRP binds the nascent polypeptide, the complex associates with a specific receptor on the ER (SRP receptor)

Endoplasmic Reticulum
Translation continues, and polypeptide is fed into the ER as it is synthesized (co-translational import)
Signal sequence is usually cleaved by signal peptidase
Another Hsp70 member, BiP lives in the ER lumen and helps pull proteins through the channel, and prevents premature folding

Endoplasmic Reticulum
It is at this point that pathways diverge to give rise to: -ER lumen proteins (soluble)-ER membrane proteins that:
-orient N-terminus in or out of the lumen-cross once or multiple times

ER lumen proteins

Single pass transmembrane proteins
Some proteins cross the ER membrane once
These have stop transfer sequences at specific sites on the proteins, typically a stretch of amino acids that give rise to a hydrophobic a-helix
These sequences can cause the protein to be oriented N-terminal in or out
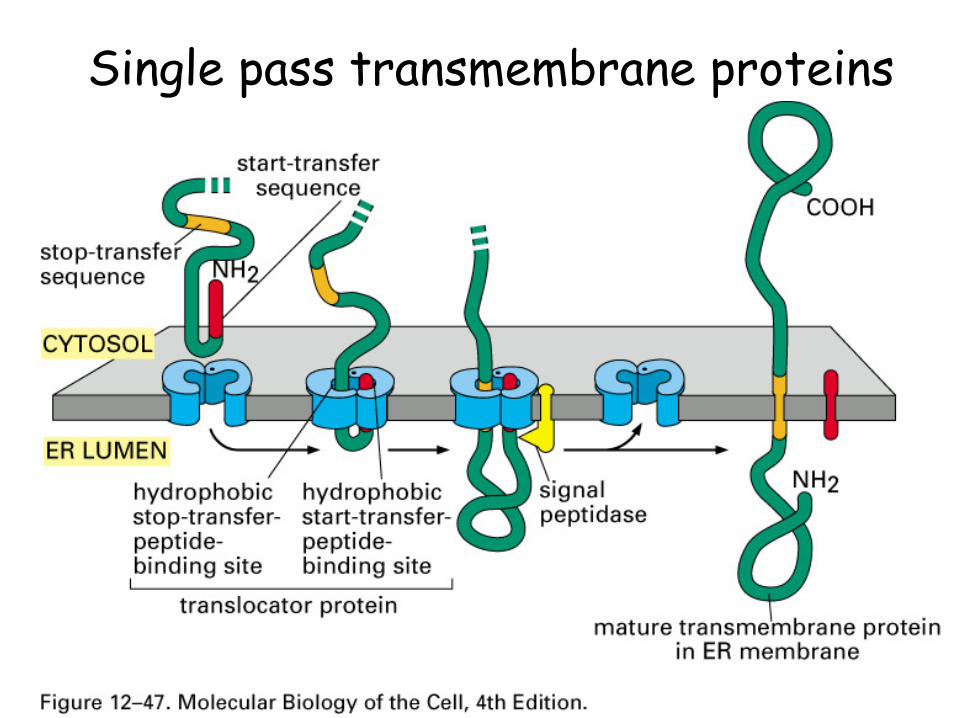
Single pass transmembrane proteins

Single pass transmembrane proteins

Single pass transmembrane proteins

Multi-pass transmembrane proteins





























