Embed Size (px)
Citation preview

690
Biochimica et Biophysica Acta, 497 (1977) 690--701 © Elsevier/North-Holland Biomedical Press
BBA 28231
CHANGES IN RAT ADRENAL CYCLIC NUCLEOTIDES DURING NORMAL AND NEOPLASTIC GROWTH
NORMA W. STOWE, THOMAS H. WHITLEY, ROBERT L. NEY and ALTON L. STEINER
Division o f Endocrinology, Department o f Medicine, University o f Norlh Carolina School o f Medicine, Chapel Hill, N.C. 27514 (U.S.A.)
(Received October 19th, 1976)
Summary
In an at tempt to correlate changes in cyclic nucleotide levels with in vivo growth of the rat adrenal gland we have measured adrenal cyclic AMP and cyclic GMP in normal, hyperplastic, and neoplastic rat adrenals. The first group of animals were subject to either unilateral adrenalectomy (ADX) or acute hypophysec tomy 1 h prior to unilateral adrenalectomy (HADX). Cyclic nucleo- tides were measured in the contralateral adrenal post-operatively. In HADX rats cyclic GMP rose steadily throughout the 7 day study period, while ADX rats exhibited significant decreases in adrenal cyclic GMP. Cyclic AMP remained approximately 1.5 pm/mg tissue in HADX rats, while in ADX rats there was significant elevation of adrenal cyclic AMP at all time points. Cyclic GMP/cyclic AMP ratios remained constant in HADX animals; however, the growing adrenals of ADX animals exhibited depressed cyclic GMP/cyclic AMP ratios at all t ime periods.
Adrenal hyperplasia was induced in a second group of animals by a trans- plantable, corticotropin-secreting, pituitary tumor. Adrenals from age-matched animals served as controls. Adrenal cyclic AMP was significantly elevated in tumor-bearers at a time corresponding to the peak accumulation of adrenal weight, protein and DNA in these animals. In contrast, adrenal cyclic GMP in both tumor-bearers and control animals fell steadily throughout the study period. Cyclic GMP/cyclic AMP ratios of control animals decreased from 2 to 3 weeks post-transplant remaining at the 3 week value during the period corre- sponding to rapid adrenal growth in tumor-bearers. The cyclic GMP/cyclic AMP ratio in the hyperplastic adrenals of tumor-bearers decreased steadily through- out their rapid growth period, suggesting a positive correlation between adrenal growth and depression of the cyclic GMP/cyclic AMP ratio.
Cyclic nucleotide levels in neoplastic adrenals of rats bearing the transplant- able adrenocortical carcinoma 494 were compared with cyclic nucleotides in normal rat adrenal glands. Cyclic AMP was not different in the two groups.

691
However, the cyclic GMP content of neoplastic adrenals was significantly lower than that of normal adrenal tissue, causing a suppression of the cyclic GMP/ cyclic AMP ratio in the neoplastic tissue. Thus, measurement of adrenal cyclic nucleotides in both hyperplastic and neoplastic rat adrenal glands suggests that adrenal growth in vivo may be characterized by a depression of the cyclic GMP/ cyclic AMP ratio.
Introduct ion
Cyclic AMP has been demonstrated to be an intracellular mediator of the growth-promoting and steroidogenic effects of cort icotropin on the rat adrenal gland [1--3]. Although the role of cyclic GMP in the rat adrenal still remains obscure, it has been implicated as a regulator of growth in some mammalian tis- sues [4]. We have previously investigated the relationship of these two cyclic nucleotides in the atrophying rat adrenal after acute and chronic hypophysec- tomy. In both acutely and chronically hypophysectomized rats, adrenal cyclic GMP rose while cyclic AMP fell. We further demonstrated that acute physiolo- gic and supraphysiologic doses of cort icotropin were able to suppress the rise in adrenal cyclic GMP in hypophysectomized rats, while producing a 50-fold or greater increase in adrenal cyclic AMP [5].
To investigate further the role of cyclic nucleotides in adrenal growth, we have studied the rat adrenal gland during hyperplastic and neoplastic growth in vivo. In all three groups of experimental animals, the cyclic GMP/cyclic AMP ratio was significantly lower than that of the respective control tissue, sug- gesting that adrenal growth in vivo may be characterized by a depression of the cyclic GMP/cyclic AMP ratio.
Materials and Methods
Materials
Cyclic nucleotides used for radioimmunoassay standards, and beef heart 3',5'-cyclic nucleotide phosphodiesterase were purchased from Sigma Chemical Company, St. Louis, Mo. Iodinated derivatives of the cyclic nucleotides and the antibodies used in the radioimmunoassay were prepared as described elsewhere [6]. Rabbit 7-globulins used as a carrier in the radioimmunoassay were pur- chased from Miles Laboratories, Inc., Miles Research Products Division, Elk- hart, Ind. Refined cort icotropin powder dissolved in 1% bovine serum albumin in 0.9% sodium chloride (final pH 7.0) was obtained from Parke, Davis Com- pany, Detroit, Mich.
Female Fischer rats (Strain 344) used to maintain the MtT/F4 tumor colony as well as age-matched female Fischer rats used as controls were purchased from Charles River Breeding Laboratories, Inc. Wilmington, Mass. Male Sprague-Dawley rats weighing 160--180 g used in unilateral adrenalectomy studies were also purchased from Charles River Breeding Laboratories, Inc.
The transplantable rat adrenocortical carcinoma 494 was maintained in male Sprague-Dawley rats as has been previously described [ 7].

692
Method of unilateral adrenalectomy Male Sprague-Dawley rats weighing 160--180 g were utilized in these experi-
ments. Rats of the same age were divided into two groups, one of which was subjected to unilateral adrenalectomy through a left costovertebral angle inci- sion. The second group of rats was hypophysec tomized by the transaural approach 1 h before the unilateral adrenalectomy was performed. Light ether anesthesia was used for the surgical procedure. Animals were killed by blunt t rauma without re-exposure to ether. At autopsy, rats with retained pituitary remnants or incomplete adrenalectomies were discarded. The right adrenal gland of each group of rats was removed at specified times, t r immed free of adhering fat and frozen immediately in liquid nitrogen. Time elapsed from excision to freezing of adrenal tissue was I min or less. Tissues were stored at --80°C until they could be processed for radioimmunoassay of cyclic nucleo- tides.
Method of tumor transplant and adrenal excision Female Fischer rats bearing the transplantable pituitary tumor MtT/F4 were
killed by blunt trauma, and tumors were immediately dissected away from the body. Tumor tissue was removed from the capsule and cut into approximately 1 mm 3 pieces. Care was taken to avoid hemorrhagic and necrotic sites of the tumor.
The 1 mm 3 pieces of tumor tissue were implanted by t rochar subcutaneously to the mid-dorsal region of female Fischer rats under light ether anesthesia. Tumors became evident approximately 2 weeks post-implantation, and success of transplant was 90% or better. Both tumor-bearing animals and their age- matched controls were maintained on Purina Rat Chow and water ad libitum.
MtT/F4 tumor-bearing animals and their age-matched controls were killed by blunt trauma at 2, 3, 4, 5, 6, and 8 weeks post-transplantation of the tumor (there were no age-matched controls for 8 week tumor bearing rats). All ani- mals were killed between 1 and 3 p.m. on the day of killing; adrenal glands were removed, t r immed free of adhering fat and immediately frozen in liquid nitrogen. In some animals, renal cortical and hepatic tissues were also obtained and frozen in similar fashion. Tissues were stored at --80°C until they could be processed for radioimmunoassay of cyclic nucleotides, and protein and DNA content . In all animals, one adrenal was assayed for cyclic nucleotides and pro- tein content , while the contralateral adrenal was assayed for DNA content . For those animals f rom which liver and renal cor tex samples were obtained, two samples of each tissue were frozen and stored; one for assay of cyclic nucleo- tides and proteins, the other for assay of DNA content .
Rats bearing the transplantable adrenocortical carcinoma 494 and intact nor- mal rats, whose adrenals served as controls, were killed by blunt trauma. Tumor tissue and normal adrenals were excised and processed for assay of cyclic nucle- otides as described below.
Assay for cyclic GMP and cyclic AMP Frozen tissue samples were extracted with trichloracetic acid [5], and ali-
quots used in the radioimmunoassay as described by Steiner et al. [8]. Confir-

693
matory tests that the assay reflected valid measurements for both nucleotides were performed as previously described [5].
Trichloroacetic acid precipitates were dissolved in 2 M NaOH and assayed for protein content as described by Lowry et al. [9].
Assay o f DNA Frozen tissue samples were weighed and homogenized in 1.0 ml of ice-cold
Krebs-Ringer bicarbonate solution with 200 mg% glucose. Aliquots were removed and diluted to 1.0 ml with buffer. 0.5 ml of 0.05 M Tris buffer (pH 7.4 with MgCI~) was added, and the resulting solution extracted with 0.5 ml of 2 M perchloric acid. Following centrifugation at 2400 X g for 10 min, the supernatant was discarded and the precipitate redissolved in 0.4 ml of 1 M NaOH. After heating in a boiling water bath for 15 min, the solution was brought to 5.0 ml with distilled water and the pH adjusted to 3.5. This solution was heated in a boiling water bath for an additional 10 min and 1.5 ml ali- quot was assayed for DNA by the diphenylamine method as described by Bur- ton [10].
Statistical analysis Data as expressed in graphs and tables represents mean + S.E. unless other-
wise noted. Statistical evaluation of the data was performed using either one-way anal-
ysis of variance and an f distribution of the experimental groups, or by the Mann-Whitney U-Test of Ranks. All values designated as statistically significant are within 95% confidence limits.
Results
Compensatory hyperplasia in response to unilateral adrenalectomy Effects o f hypophysectomy plus unilateral adrenalectomy (HADX) and uni-
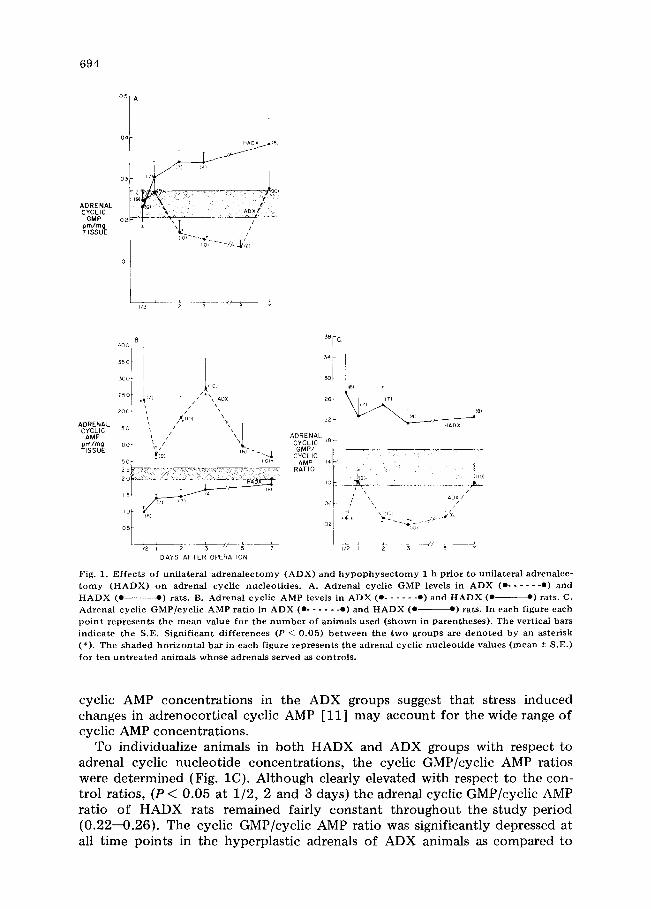
lateral adrenalectomy alone (ADX) on adrenal cyclic GMP, cyclic AMP and adrenal cyclic GMP/cyclic AMP ratio. Rat adrenal cyclic GMP and cyclic AMP were measured over a 7 day period in the right adrenal of animals unilaterally adrenalectomized (ADX) and in the right adrenal of animals acutely hypophy- sectomized 1 h prior to unilateral adrenalectomy (HADX). Cyclic nucleotide levels from ten untreated animals served as control values. In HADX rats, adre- nal cyclic GMP rises gradually over the 7 day study period, and by 2 days post- operatively is clearly elevated over b o t h control ( P < 0.05) and ADX ( P < 0.01) values (Fig. 1A). This observation is similar to our previous findings that rat adrenal cyclic GMP rises following acute and/or chronic hypophysectomy [ 5]. In contrast, the hyperplastic adrenals from ADX animals exhibited signifi- cant decreases in cyclic GMP at 2, 3 and 5 days postoperatively when compared to control (P < 0.025) and HADX (P < 0.01) values.
The cyclic AMP content of ADX and HADX adrenals is shown is Fig. lB. In HADX rats, adrenal cyclic AMP levels are clearly lower than control values from 1/2 to 5 days and are significantly different from the cyclic AMP levels of ADX rats at all time points (P < 0.05). Conversely, ADX adrenal cyclic AMP is elevated by comparison with control (P < 0.05) values. Analysis of individual
694
ADRENAL CYCLIC
GMP pm/mg TISSUE
HADX ~(8)
03 fT)
o2 ~ "\ T #
(IO)~--'~_ l
,~ I ~ ~ ~F -~
ADRENAL CYCLIC AMP
pm/mg TISSUE
B
°°°I25o I 350 T
.~O0 ,(io)
200~ / s \ I ~io) \ i
150 / "/4 i //1 \\x ioo t /
50 * ~io1.
15
0 5
DAYS AFTER OPERATION
ADRENAL CYCLIC GMP/
CYCLIC AMP
RATIO
38Fc
34~ !
26 i(7) [
(4) 22 ~ HADX
~8
14
o . . . . . _ . . . . . . . . . . . . .
06 L \\X & -~- ,A x /
A :i #
Fig . 1. E f f e c t s o f u n i l a t e r a l a d r e n a l e c t o m y ( A D X ) a n d h y p o p h y s e c t o m y 1 h p r i o r t o u n i l a t e r a l a d r c n a l e c - t o m y ( H A D X ) o n a d r e n a l c y c l i c n u c l e o t i d e s . A . A d r e n a l c y c l i c G M P l e v e l s in A D X ( e . . . . . . e ) a n d H A D X ( e - e ) rats . B. A d r e n a l c y c l i c A M P l e v e l s in A D X ( e . . . . . . e ) a n d H A D X ( e ~ ) rats . C. A d r e n a l c y c l i c G M P / c y c l i c A M P r a t i o in A D X ( e . . . . . . e ) a n d H A D X ( e - - e ) rats . In e a c h f i g u r e e a c h p o i n t r e p r e s e n t s t h e m e a n v a l u e f o r t h e n u m b e r o f a n i m a l s u s e d ( s h o w n in p a r e n t h e s e s ) . T h e v e r t i c a l bars i n d i c a t e t h e S . E . S i g n i f i c a n t d i f f e r e n c e s (P ~ 0 . 0 5 ) b e t w e e n t h e t w o g r o u p s arc d e n o t e d b y an a s t e r i s k ( * ) . T h e s h a d e d h o r i z o n t a l b a r in e a c h f i g u r e r e p r e s e n t s t h e a d r e n a l c y c l i c n u c l e o t i d e v a l u e s ( m e a n + S . E . ) f o r t e n u n t r e a t e d a n i m a l s w h o s e a d r e n a l s s e r v e d as c o n t r o l s .
cyclic AMP concentrations in the ADX groups suggest that stress induced changes in adrenocortical cyclic AMP [11] may account for the wide range of cyclic AMP concentrations.
To individualize animals in both HADX and ADX groups with respect to adrenal cyclic nucleotide concentrations, the cyclic GMP/cyclic AMP ratios were determined (Fig. 1C). Although clearly elevated with respect to the con- trol ratios, (P < 0.05 at 1/2, 2 and 3 days) the adrenal cyclic GMP/cyclic AMP ratio of HADX rats remained fairly constant throughout the study period (0.22--0.26). The cyclic GMP/cyclic AMP ratio was significantly depressed at all time points in the hyperplastic adrenals of ADX animals as compared to

695
HADX animals (P < 0.05). At 1/2, 2, 3, and 5 days post-operatively the adrenal cyclic GMP/cyclic AMP ratio of ADX animals was clearly lower than that of the control groups (P < 0.01). The transient rise in the ADX ratio at 1 day may well be an artifact at tr ibutable to the large individual variation in cyclic AMP values at 1/2 and 1 day.
Adrenal weights, protein and DNA/adrenal in ADX and HADX rats. Adrenal weights and total protein per adrenal of rats either unilaterally adrenalecto- mized (ADX) or hypophysectomized prior to unilateral adrenalectomy (HADX) throughout 7 days are shown in Figs. 2A and B, respectively. Adrenal weight had increased significantly by two days in ADX animals ( P < 0.01) when compared to 1/2 day ADX animals. Total protein per adrenal was signifi- cantly greater at five days (P < 0.03) when compared to 1/2 day ADX animals. In contrast, adrenal weight and total protein per adrenal fell steadily with time in HADX animals over the seven day period (P < 0.01), and from 2 to 7 days post-operatively both weight and total protein/adrenal in HADX animals were significantly different from those of ADX animals.
The DNA content of single adrenals from eight intact rats increased from 21.75 + 2.59 pg DNA-P/adrenal to 36.76 + 2.79 pg DNA-P/adrenal at 3 days following unilateral adrenalectomy (P < 0.04) and to 45.06 + 4.55 pg DNA-P/ adrenal at 7 days after unilateral adrenalectomy (P < 0.01).
The increases in adrenal weight, protein and DNA content in ADX animals are indicative of compensatory hyperplasia of the contralateral gland following unilateral adrenalectomy and parallel the depressed cyclic GMP concentrations and low cyclic GMP/cyclic AMP ratios in ADX animals as shown in Figs. 1A and C.
Adrenal hyperplasia induced by a corticotropin-secreting pituitary tumor Rat adrenal weight, protein, and DNA content during the latent and rapid
growth phases of the transplantable pituitary tumor, MtT/F4. Adrenal weight, protein, and DNA content were measured in female Fischer rats bearing the transplantable pituitary tumor, MtT/F4 at 2, 3, 4, 5, 6 and 8 weeks post-im-
30 r A. 50
I WEIGHT 20 PROTE IN 30
mg/ADRENAL mg/ADRENAL
1.0
I I I I I I I I 1/21 2 3 4 5 6 7
DAYS POST OPERATION
'B.
T z,-';Dx
J e I I ! I I
I,,'zl 2 3 4 5 6 7 DAYS POST OPERATION
Fig . 2. A d r e n a l w e i g h t a n d p r o t e i n c o n t e n t in u n i l a t e r a l l y a d r e n a l e c t o m i z e d ( A D X ) r a t s a n d r a t s s u b j e c t t o h y p o p h y s e c t o m y 1 h p r i o r t o u n i l a t e r a l a d r e n a l e c t o m y ( H A D X ) . A. A d r e n a l w e i g h t i n A D X ( e . . . . . . e ) a n d H A D X (e "-) r a t s . B. A d r e n a l p r o t e i n c o n t e n t i n A D X (e . . . . . . e ) a n d H A D X ( e --) r a t s . E a c h p o i n t r e p r e s e n t s t h e m e a n v a l u e f r o m f o u r o r m o r e a n i m a l s . V e r t i c a l b a r s r e p r e s e n t t h e S .E . S i g n i f i c a n t d i f f e r e n c e s b e t w e e n t h e t w o g r o u p s a re d e n o t e d b y * (P < 0 . 0 1 ) a n d ** (P < 0 . 0 5 ) .

696
plantation (Fig. 3). Adrenal DNA content increased at 3 weeks post-transplan- tation, while adrenal weight and protein showed significant increases at 4 weeks post-transplantation. This lag in adrenal growth in tumor-bearing animals is in agreement with the latent growth period of the MtT/F4 as described by Mil- kovic and Bates [12]. During the time period previously described by Milkovic and Bates as the rapid growth phase of the tumor, adrenal DNA content rose almost six-fold from 3 weeks to 5 weeks post-transplant, followed by a conco- mitant increase in adrenal weight and the expected rise of adrenal protein during this time period. At 5--8 weeks post-transplant, adrenal weight, protein, and DNA content plateaued in accord with previous observations that tumor- bearing animals exhibit a plateau of tumor and adrenal growth known as the mature phase [ 12 ].
Age-matched control animals showed no significant changes in either adrenal protein (mg/adrenal) or adrenal DNA content (pg DNA-P/mg tissue) from 2 to 6 weeks post-implantation. Adrenal weight, however showed a significant (P < 0.01) increase from 22.85 + 1.25 mg/adrenal at 2 weeks post-transplant to 30.78 + 1.67 mg/adrenal at 6 weeks post-transplant in control animals.
Rat adrenal cyclic nucleotide levels during the latent and rapid growth phases of the MtT/F4. Adrenal cyclic nucleotide levels were measured in tumor-bearing animals and their age-matched controls from 2 to 8 weeks post- implantation. During the latent period of tumor growth (2--3 weeks post-trans-
35O
300 / ~ /~//J
25o ~.. ~ ~ ~ " "
)ugmlADRENAL // /
AND 150 i/I 120 [ / 25 0
mglADRENAL I00 DNA// 20 0
,, / /i/ / /PROTEIN 15 0 PROTEIN 60 mglADRENAL
40 / ~ _ _ _ _ ~ / ~ ~ . _ ~ I O0
5O 2O
I I
WEEKS POST TRANSPLANT Fig. 3. A d r e n a l w e i g h t , p r o t e i n , a n d D N A c o n t e n t i n M t T / F 4 t u m o r beare r s . A d r e n a l D N A (e . . . . . . e) and a d r e n a l w e i g h t ( e -~) are s h o w n o n t h e l e f t v e r t i c a l axis . A d r e n a l p r o t e i n (A A) is s h o w n o n
t he r i g h t v e r t i c a l axis . E a c h p o i n t r e p r e s e n t s t he m e a n v a l u e s f r o m f o u r o r m o r e a n i m a l s . V e r t i c a l ba r s rep- r e s e n t t h e S .E.

697
plant) there were no significant changes in adrenal cyclic AMP (Fig. 4A). From 3 weeks to 6 weeks, however adrenal cyclic AMP levels rose in tumor bearing animals reaching a level at 6 weeks that was 10 times greater than the cyclic AMP concentration at 3 weeks. This increase in cyclic AMP is concomitant with the observed accumulation of adrenal DNA in the tumor bearers.
In marked contrast to adrenal cyclic AMP concentrations, adrenal cyclic GMP levels in tumor-bearing animals exhibited a significant fall (P < 0.01) from 2 weeks to 4 weeks post-transplantation (Fig. 4B). This early decline of adrenal cyclic GMP in tumor bearers precedes any significant accumulation of adrenal weight, protein or DNA. The decrease in adrenal cyclic GMP in tumor-bearing animals was maintained at 5 weeks post-transplant but had returned to the range of the 2 weeks levels of adrenal cyclic GMP at 6 and 8 weeks. The re-es- tablishment of the earlier levels of adrenal cyclic GMP at 6 and 8 weeks post- transplant coincides with the trend toward a steady state in adrenal weight, protein, and DNA in the tumor bearers.
Age-matched control animals fol lowed the same adrenal cyclic GMP pattern as their tumor-bearing counterparts, and there is no statistically significant dif- ference between the two groups. Age-matched control animals at the 2 to 4 weeks post-transplant time period were 60- -84 days old, an age corresponding to an approximately 50% increase in body weight in female Fischer rats (un- published data, Charles River Breeding Laboratories Inc.). Although there were no changes in adrenal cyclic AMP levels in these animals, cyclic GMP levels were significantly depressed during this time period. Bearing in mind the significant
zoo- A
15o
ioo 70
ADRENAL CYCLIC
AMP 60 pm/mg TISSUE
50
40
30
20
IC
TUM~T_/F~RE e
~ ~ ~', //CONTROLS
500
400
300
ADRENAL CYCLIC
GMP pm/mg TISSUE
200
~00
~ T MtT/F4 UMOR-BEARERS
\ /
WEEKS POST TRANSPLANT WEEKS POST TRANSPLANT
Fig . 4. A d r e n a l c y c l i c n u c l c o t i d e levels in M t T / F 4 t u m o r bearers a n d the ir a g e - m a t c h e d c o n t r o l s . A. Adre - nal c y c l i c A M P levels in M t T / F 4 t u m o r bearers ( e e ) and a g e - m a t c h e d c o n t r o l an imals ( e . . . . . . e ) . B . A d r e n a l c y c l i c G M P in M t T / T 4 t u m o r bearers ( e e ) and a g e - m a t c h e d c o n t r o l s ( e . . . . . . e ) . Each p o i n t represent s the m e a n va lue f r o m four or m o r e animals . Vert ica l bars r e p r e s e n t the S.E. S ign i f i cant d i f f e r e n c e s ( P < 0 . 0 5 ) b e t w e e n the t w o groups are r e p r e s e n t e d b y an aster isk ( * ) .
698
increase in adrenal weight in control animals reported above, this may be con- strued as a further indication of a positive correlation between rapid growth and a decline in the level of cyclic GMP in the adrenal gland. Furthermore, once the animal's body weight and organ weight reaches a steady state, it would appear that the cyclic GMP concentration of the adrenal gland returns to the earlier value.
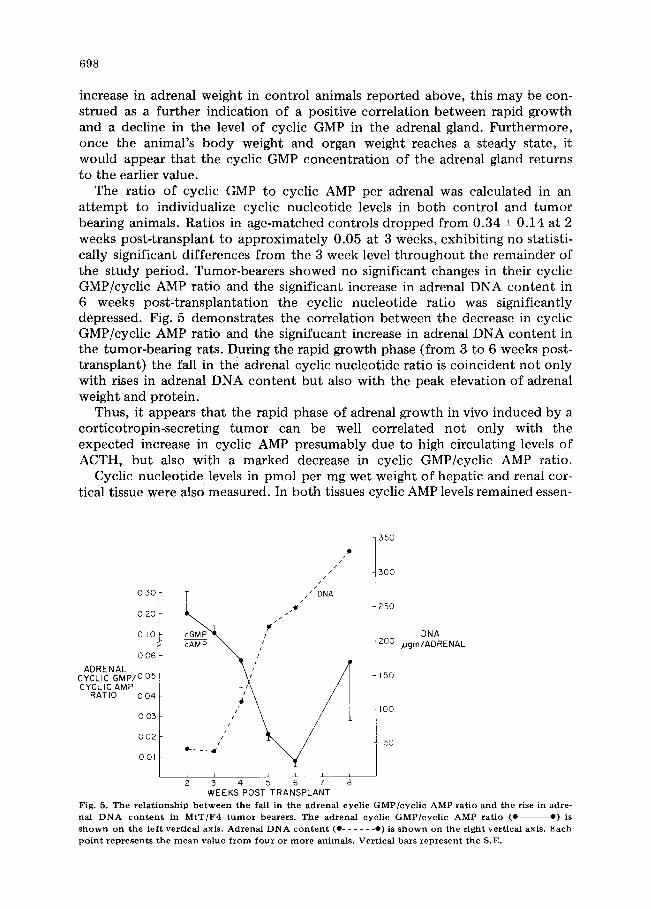
The ratio of cyclic GMP to cyclic AMP per adrenal was calculated in an attempt to individualize cyclic nucleotide levels in both control and tumor bearing animals. Ratios in age-matched controls dropped from 0.34 -+ 0.14 at 2 weeks post-transplant to approximately 0.05 at 3 weeks, exhibiting no statisti- cally significant differences from the 3 week level throughout the remainder of the study period. Tumor-bearers showed no significant changes in their cyclic GMP/cyclic AMP ratio and the significant increase in adrenal DNA content in 6 weeks post-transplantation the cyclic nucleotide ratio was significantly depressed. Fig. 5 demonstrates the correlation between the decrease in cyclic GMP/cyclic AMP ratio and the signifucant increase in adrenal DNA content in the tumor-bearing rats. During the rapid growth phase (from 3 to 6 weeks post- transplant) the fall in the adrenal cyclic nucleotide ratio is coincident not only with rises in adrenal DNA content but also with the peak elevation of adrenal weight and protein.
Thus, it appears that the rapid phase of adrenal growth in vivo induced by a corticotropin-secreting tumor can be well correlated not only with the expected increase in cyclic AMP presumably due to high circulating levels of ACTH, but also with a marked decrease in cyclic GMP/cyclic AMP ratio.
Cyclic nucleotide levels in pmol per mg wet weight of hepatic and renal cor- tical tissue were also measured. In both tissues cyclic AMP levels remained essen-
030
0 20
0 1 0
0.06
ADRENAL CYCLIC GMP/005 CYCLIC AMP RATIO OO4
0 03
0O2
001
/ /
/ /
//DNA
cGM~P i ~j~j
/!
jH e . . . - _ _ 4 • t ' ' '
L I I I I I / 2 3 4 5 6 7 8
WEEKS POST T R A N S P L A N T
350
3 0 0
250
DNA 200 ~ g m / A D R E N A L
150
100
50
Fig. 5. T h e r e l a t i o n s h i p b e t w e e n t he fall in t he a d r e n a l c y c l i c G M P / c y c l i c A M P ra t io a n d t h e rise in adre - nal DNA c o n t e n t in M t T / F 4 t u m o r b e a r e r s . T h e a d r e n a l c y c l i c G M P ] c y c l i c A M P ra t io (0 o) is s h o w n o n the l e f t ver t i ca l ax i s . A d r e n a l D N A c o n t e n t (0 . . . . . . o) is s h o w n o n the r i g h t ver t i ca l ax i s . E a c h
p o i n t r e p r e s e n t s t h e m e a n v a l u e f r o m f o u r o r m o r e a n i m a l s . V e r t i c a l b a r s r e p r e s e n t the S.E.

6 9 9
T A B L E I
C O N C E N T R A T I O N S OF CYCLIC AMP, CYCLIC GMP A N D T H E CYCLIC G M P / C Y C L I C AMP R A T I O IN N O R M A L R A T A D R E N A L S A N D IN A N A D R E N O C O R T I C A L R A T C A N C E R
Values given are the m e a n s +- S.E. The n u m b e r in pa ren theses indicates the n u m b e r of rats.
Tissue Cyclic AMP Cyclic GMP Cyclic GMP/cyc l ic AMP (pro/ rag t issue) ( p m / m g tissue) ra t io
Adrena ls 2 .479 +- 0 . 525 0 .237 -+ 0 .032 0 .130 + 0 .0 3 5 (i0) (10) (i0)
Tumor 2.290 -+ 0.517 0.069 -+ 0.011 0.035 -+ 0.005
(rat adrena l cancer) (11) (11) (11)
P value > 0 . 0 5 < 0 . 0 0 1 < 0 . 0 5
tially constant over the 8 week time period in both tumor bearing and age- matched control animals; and there were no significant differences between the two groups at any time point. In sharp contrast, the cyclic GMP levels in both hepatic and renal cortical tissues decreased 3- to 5-fold at 3 and 5 weeks post- transplantation. Cyclic GMP levels in control animals were not significantly dif- ferent. In both tissues, cyclic GMP/cyclic AMP ratios are decreased at 3 and 5 weeks with no significant differences between control animals and tumor bear- ers, suggesting that the observed changes in hepatic and renal cortical cyclic GMP and the cyclic GMP/cyclic AMP ratio are related to the animals' develop- mental growth.
Adrenal neoplasia Concentrations o f cyclic AMP, cyclic GMP and cyclic GMP/cyclic AMP ratio
in normal rat adrenals and an adrenocortical rat cancer. Table I shows the con- centrations of cyclic AMP, cyclic GMP and cyclic GMP/cyclic AMP ratio in nor- mal rat adrenals and those of an adrenocortical rat carcinoma. There is no sig- nificant difference in the cyclic AMP levels in normal adrenals and that of the tumor, an observation previously noted [7]. In contrast to cyclic AMP, the tumor cyclic GMP concentration is significantly lower than that of normal rat adrenals ( P < 0.001) and the cyclic GMP/cyclic AMP is similarly depressed (P < 0.05). Thus, the adrenocortical rat cancer maintains cyclic AMP levels not different from normal adrenals while the cyclic GMP concentration of the tumor is markedly suppressed, demonstrating a positive correlation between rapid adrenal growth and decreased levels of adrenal cyclic GMP.
Discussion
In each of the in vivo growth models studied, we found a depressed cyclic GMP/cyclic AMP ratio associated with rapid adrenal growth. The reason for the depression of the cyclic GMP/cyclic AMP ratio in the growing adrenal gland is not entirely clear. The stress induced response of the pituitary-adrenal axis is well established; therefore, the possibility exists that the observed changes in

700
cyclic nucleotides in hyperplastic adrenal tissues may be a result of transient increases in corticotropin during killing of non-hypophysectomized rats by blunt trauma. Values from normal intact rats indicate that this method of killing results in rat adrenal cyclic nucleotide levels quite different from those observed in hyperplastic adrenals. Thus, it appears that the reported changes in both cyclic AMP and cyclic GMP in the growing adrenal gland are a reflection of the growth process itself. Observations of a depressed cyclic GMP/cyclic AMP ratio in the hyperplastic adrenal tissues might be explained on the basis of elevated levels of cyclic AMP due to the high circulating levels of corticotropin presumed to occur during both compensatory hyperplasia [14] and the rapid growth phase of the MtT/F4 [12]. However, we observed significant decreases in cyclic GMP as well as increases in cyclic AMP in the hyperplastic growth models; and while the neoplastic adrenal of the adrenocortical carcinoma 494, which is presumably subject to low circulating levels of corticotropin [15], exhibited no change in cyclic AMP, the cyclic GMP levels were markedly sup- pressed. It is therefore unlikely that the known action of corticotropin on the adrenal gland is the sole reason for the correlation between a depressed cyclic GMP/cyclic AMP ratio and the growing adrenal gland.
While we observed a depressed cyclic GMP/cyclic AMP ratio in growing adre- nal, hepatic and renal tissues, others have reported data that are at some vari- ance with our findings. For example, Schlondorff and Weber [16] reported an increase in cyclic GMP levels at a time of rapid growth in developing rat (Sprague-Dawley) kidney. Criss et al. [17] have reported elevated leve~s of cyclic GMP in rat renal cortical tumors which may be explained by the increased levels of particulate guanylate cyclase in these neoplasias. Wise et al. [18], however, were unable to show any changes in either cyclic AMP or cyclic GMP is unilaterally nephrectomized Wistar rats. De Rubertis found the cyclic GMP/cyclic AMP ratio to be increased in human adenocarcinoma of the colon when compared to normal colonic mucosa [19]. In liver growth after partial hepatectomy, cyclic GMP levels remain constant [20] and cyclic AMP is ele- vated at a time of rapid DNA synthesis. These changes result in a depressed cyclic GMP/cyclic AMP ratio because of this rise in cyclic AMP, whereas our own observation of a depressed cyclic GMP/cyclic AMP ratio in liver was due to a fall in the hepatic cyclic GMP.
It appears that no simple conclusions can be made from these studies. It seems likely that the role of the cyclic nucleotides in growth regulation in dif- ferent tissues is variable. Furthermore, it is unclear as to whether or not cyclic nucleotides exert a primary influence in growth processes. It has been postu- lated that the cyclic nucleotides help to regulate calcium concentrations at key intracellular sites, and tha~ this divalent cation is an important modulator of growth [21]. It seems likely that individual cells utilize calcium and the cyclic nucleotides in different ways in hormonally-directed events. In addition, whole tissue levels may not be a sensitive index of cyclic nucleotide regulation at spe- cific intracellular sites. The use of more discriminating techniques such as isola- tion of individual cell types, cell fractionation and cyclic nucleotide immuno- cytochemical localization in conjunction with quantitative cyclic nucleotide measurements would further the understanding of the role of the cyclic nucleo- tides in growth regulation in t~s,~ues.

701
Acknowledgements
The authors wish to thank Dr. Arthur E. Bogden of the Mason Research In- stitute, Worcester, Mass., for his generous gift of five MtT/F4 tumor-bearing rats with which we began our own tumor colony. This study was supported by U.S. Public Health Service Grant AM17438.
References
1 H a y n e s , J r . , R .C. ( 1 9 5 8 ) J . Biol. C h e m . 233 , 1 2 2 0 - - 1 2 2 2 2 H a y n e s , J r . , R .C. , K o r i t z , S.B. a n d Pe ron , F .G. ( 1 9 5 9 ) J . Biol. C h e m . 234 , 1 4 2 1 - - 1 4 2 3 3 G r a h a m e - S m i t h , D .G. , B u t c h e r , R.W., Ney , R.L. a n d S u t h e r l a n d , E.W. ( 1 9 6 7 ) J . Biol. C h e m . 2 4 2 ,
5 5 3 5 - - 5 5 4 1 4 G o l d b e r g , N .D. , O ' D e a , R . F . , a n d H a d d o x , M.K. ( 1 9 7 3 ) in A d v a n c e s in Cyc l ic N u c l e o t i d e R e s e a r c h
( G r e e n g a r d , P. a n d R o b i s o n , G .A . , eds.) , Vol . 3, pp . 1 5 5 - - 2 2 3 , R a v e n Press, N e w Y o r k 5 Whi t l ey , T .H. , S t o w e , N.W., Ong , S .H. , N e y , R .L . a n d S te ine r , A .L . ( 1 9 7 5 ) J. Clin. Inves t . 5 6 , 1 4 6 - -
1 5 4 6 S te ine r , A .L . , Pa rke r , C.W. a n d Kipn i s , D.M. ( 1 9 7 2 ) J . Biol. C h e m . 247 , 1 1 0 6 - - 1 1 1 3 7 N e y , R .L . , Hoehe l l a , N . J . , G r a h a m e - S m l t h , D .G. , D e x t e r , R .N. a n d B u t c h e r , R.W. ( 1 9 6 9 ) J. Clin. In-
vest . 48 , 1 7 3 3 - - 1 7 3 9
8 S te ine r , A.L. , Pagl ia ra , A.S. , Chase , L .R . a n d Kipn i s , D.M. ( 1 9 7 2 ) J . Biol. C h e m . 247 , 1 1 1 4 - - 1 1 2 0 9 L o w r y , O .H. , R o s e b r o u g h , N.J . , Fa r r , A.L. a n d R a n d a l l , R . J . ( 1 9 5 1 ) J . Biol. C h e m . 193 , 2 6 5 - - 2 7 5
1 0 B u r t o n , K. ( 1 9 5 6 ) B i o c h e m . J. 62 , 3 1 5 - - 3 2 3 11 Paul , M.I. , K u e t n a n s k y , R. , C r a m e r , H. , Sf lberge ld , S. a n d K o p i n , I .J . ( 1 9 7 1 ) E n d o c r i n o l o g y 88 , 3 3 8 - -
3 4 4 1 2 Milkovic , S. a n d Bates , R.W. ( 1 9 6 4 ) E n d o c r i n o l o g y 7 4 , 6 1 7 - - 6 2 6 13 R i c h m a n , R. , D o b b i n s , C., V o i n a , S., U n d e r w o o d , L. , M a h a f f e e , D. , G i t e l m a n , H.J . V a n Wyk , J . a n d
N e y , R .L . ( 1 9 7 3 ) J . Clin. Inves t . 52, 2 0 0 7 - - 2 0 1 5 1 4 Saye r s , G. a n d Saye r s , M. ( 1 9 4 8 ) R e c e n t Prog . H o r m o n e Res . 2, 8 1 - - 1 1 5 1 5 J o h n s o n , D .F . , Snel l , K.C. , F r a n c o i s , D. a n d H e f t m a n n , E. ( 1 9 6 1 ) A e t a E n d o c r i n o l . 37 , 3 2 9 - - 3 3 5 16 S c h l o n d o r f f , D. a n d Weber , H. ( 1 9 7 6 ) Proc . Nat l . A c a d . Sci. U.S. 73 , 5 2 4 - - 5 2 8 17 Criss, W.E. , M u r a d , F. a n d K i m u r a , H. ( 1 9 7 6 ) J . Cycl ic N u c l e o t i d e Res. 2, 1 1 - - 1 9 1 8 Wise, P .M. , R a t n e r , A. , S o l o m o n , S. a n d S a n b o r n , C .R. ( 1 9 7 6 ) Fed . P roc . 35 , 2 9 5 ( abs t r . ) 19 D e R u b e r t i s , F . R . , C h a y o t h , R. a n d Fie ld , J .B. ( 1 9 7 6 ) J . Clin. Inves t . 5 7 , 6 4 1 - - - 6 4 9 2 0 M a c M a n u s , J .P . , Whl t f i e ld , J . F . , B o y n t o n , A .L . a n d R ix in , R .H . ( 1 9 7 5 ) in A d v a n c e s in Cycl ic Nuc l eo -
t ide R e s e a r c h ( D r u m m o n d , G.I . , G r e e n g a r d , P. a n d R o b i s o n , G .A. , eds.) , Vol . 5, pp . 7 1 9 - - 7 3 4 , R a v e n Press, N e w Y o r k
21 Ber r idge , M.J . ( 1 9 7 5 ) J . Cyc l ic N u c l e o t i d e Res. 1 , 3 0 5 - - 3 2 0





























