Embed Size (px)
Citation preview

CENTRO DE INVESTIGACIÓN Y DE ESTUDIOS
AVAN ZADOS DEL INSTITUTO POLITÉCNICO
NACIONAL
UNIDAD MÉRIDA
DEPARTAMENTO DE FÍSICA APLICADA
ELECTROFISIOLOGÍA Y TERMOELECTRICIDAD
EN SISTEMAS VEGETALES
Tesis que presenta
KAREN BEATRÍZ GÓMEZ HERNÁNDEZ
para obtener el grado de
MAESTRA EN CIENCIAS
en la especialidad de
FISICOQUÍMICA
Directores de tesis
DR. RODRIGO TARKUS PATIÑO DIAZ
DR. MIGUEL ANGEL MUNGUÍA ROSAS
MÉRIDA, YUCATÁN AGOSTO 2016

Este trabajo de tesis se realizó en tres diferentes Laboratorios: Laboratorio de
Termodinámica Experimental del Departamento de Física Aplicada y Laboratorio de
Ecología Terrestre del Departamento de Ecología Humana, ambos en el Centro de
Investigación y de Estudios Avanzados – Unidad Mérida, y en el Laboratoire
Interdisciplinaire des Energies de Demain (LIED) de la Universidad de París 7 Diderot,
Francia.
Los resultados parciales fueron presentados en la ciudad de Puebla, en el Coloquio
Nacional de Calorimetría, Análisis Térmico y Termoquímica en la Benemérita Universidad
Autónoma de Puebla, del 6 al 8 de abril de 2016.

I
AGRADECIMIENTOS
Primeramente, quiero agradecerle a Dios por bendecirme en cada momento de mi
vida y por darme la fuerza necesaria en todo momento.
A mis padres, por ser siempre el mejor apoyo en mi vida y por dedicarse a motivarme
con su amor incondicional, para ser lo que ahora soy.
A mi esposo, por todo el amor, apoyo y comprensión que siempre me ha otorgado y
por estar conmigo cumpliendo una meta más.
Al Dr. Rodrigo Patiño por su tiempo otorgado, por brindarme la confianza y
transmitirme su conocimiento y experiencia, tanto para un crecimiento personal como
profesional.
Al Dr. Miguel Munguía y a su equipo de trabajo por brindarme la confianza para el
uso de su equipo de Laboratorio y por el asesoramiento durante el proceso. También, por sus
aportaciones para el mejoramiento de este trabajo.
A los asesores, Dr. Luis Díaz y Dr. Geonel Rodríguez por sus correcciones y
aportaciones para el mejoramiento de la tesis.
Al Dr. Christophe Goupil por el apoyo brindado durante la estancia en Paris, Francia.
A los estudiantes de Licenciatura, Karen May y Dylan Peniche por su valioso apoyo
durante la realización de experimentos.
Al centro de Investigación y de Estudios Avanzados – Unidad Mérida por la
formación académica de excelencia y al Consejo Nacional de Ciencia y Tecnología
(CONACYT) por el apoyo económico brindado durante el posgrado.

II
RESUMEN
Este trabajo de investigación tuvo como objetivo explorar la electrofisiología de los
sistemas vegetales y adentrarse en el fenómeno termoeléctrico de estos sistemas. El
fenómeno termoeléctrico o efecto Thomson es una generalización de dos efectos, Seebeck y
Peltier, que conjugan la interacción de la variación de la temperatura con la diferencia del
potencial eléctrico de un material. Aunque estos fenómenos termoeléctricos son ampliamente
conocidos en materiales metálicos o en el estado sólido, existen pocas referencias en
materiales biológicos o sistemas vivos.
Se estudió el fenómeno termoeléctrico en tres especies de plantas: Ruellia nudiflora
(Acanthaceae), Epipremnum aureum (Araceae) y Ficus benjamina (Moraceae) con tres
diferentes metodologías: (1) controlando la temperatura de la raíz en solución acuosa dentro
de un baño térmico; (2) controlando la temperatura ambiental en una cámara climática, con
posibilidad de hacer experimentos simultáneos con seis plantas en suelo como sustrato; y (3)
midiendo este fenómeno en diferentes partes de una planta.
Al someter a las plantas a una diferencia de temperatura entre las raíces y el tallo se
determinaron los valores del coeficiente Seebeck para R. nudiflora con un intervalo de 0.6 a
13 mV/K. En el caso de E. aureum, se encontró que el potencial eléctrico no depende de las
diferencias de temperatura, por lo que el valor del coeficiente Seebeck era muy cercano a
cero, sin embargo, se observó que el resultado es dependiente de la altura del tallo en la que
se hace la medición, así como también de la posición de la planta, Para F. benjamina, se
hicieron experimentos dinámicos para observar la respuesta eléctrica en diferentes partes de
la planta y con base a esto, se obtuvo una correlación de esta respuesta con la temperatura.
Estos resultados podrán dar la pauta para la generación de un modelo que explique el
fenómeno electrofisiológico en los sistemas vegetales y para encontrar posibles aplicaciones.

III
ABSTRACT
This research had the objective of exploring the electrophysiology of the vegetables
systems and deepening into the thermoelectric phenomenon of these systems. The
thermoelectric phenomenon or Thompson effect is a generalization of two effects, Seebeck
and Peltier, which combines the interaction between the variation of the temperature and the
electric potential of a material. Even though these thermoelectric phenomena are widely
known in metallic materials or in solid state, there are few references in biological materials
or living systems.
The thermoelectric phenomenon was studied in three species of plants: Ruellia nudiflora
(Acanthaceae), Epipremnum aureum (Araceae) and Ficus benjamina (Moraceae), and with
three different methodologies : (1) by controlling the temperature of the root in aqueous
solution inside a thermal bath.; (2) by controlling the room temperature in a climatic chamber,
with the possibility of doing simultaneous experiments with six plants on the soil as
substratum; and (3) by measuring this phenomenon at different parts of a plant.
When exposing the plants to variations of temperature between the roots and the stem,
the Seebeck coefficient for R. nudiflora was determined within an interval of values from 0.6
to 13 mV/K. In the case of E. aureum, it was found that the electric potential does not depend
from the temperature variations, which means that the Seebeck coefficient value was very
close to zero, however, it was noticed that the electric potential is dependent on the height of
the stem at which the measurement is done, as well as on the plant position. For F. benjamina,
dynamic experiments were performed to observe the electric answer in different parts from
the plant, and with this foundation, a correlation was obtained from this answer with the
temperature variations. These results could give a guideline for the generalization of a model
that explains the electrophysiology phenomenon in the vegetable systems, as well as to find
possible applications.

IV
INDICE GENERAL
AGRADECIMIENTOS ..................................................................................................................... I
RESUMEN ......................................................................................................................................... II
ABSTRACT ...................................................................................................................................... III
INDICE GENERAL ......................................................................................................................... IV
ÍNDICE DE FIGURAS .................................................................................................................... VI
ÍNDICE DE TABLAS ..................................................................................................................... VII
INTRODUCCIÓN ............................................................................................................................. 1
1.1. Termoelectricidad ............................................................................................................. 1
1.1.1. Efecto Seebeck ............................................................................................................... 2
1.1.2. Efecto Peltier .................................................................................................................. 3
1.2. Fisiología y electrofisiología vegetal ................................................................................. 5
1.3. Especies vegetales .............................................................................................................. 7
1.3.1. Ruellia nudiflora ............................................................................................................ 8
1.3.2. Epipremnum aureum .................................................................................................... 8
1.3.3. Ficus benjamina ............................................................................................................. 9
1.4. Objetivo ............................................................................................................................ 10
2. SECCIÓN EXPERIMENTAL ............................................................................................... 11
2.1. Materiales y Métodos ...................................................................................................... 11
2.1.1. Descripción general de los experimentos ................................................................... 11
2.1.2. Baño térmico ................................................................................................................ 11
2.1.3. Cámara climática ........................................................................................................ 14
2.1.4. Conexiones múltiples ................................................................................................... 16
2.2. Resultados ........................................................................................................................ 19
2.2.1. Ruellia nudiflora .......................................................................................................... 20
2.2.2. Epipremnum aureum .................................................................................................. 25
2.2.3. Ficus benjamina ........................................................................................................... 26
2.3. Discusión .......................................................................................................................... 30
3. CONCLUSIONES Y PERSPECTIVAS ................................................................................ 32
BIBLIOGRAFÍA ............................................................................................................................. 34
APÉNDICE A1 ................................................................................................................................ 36
APÉNDICE A2 ................................................................................................................................ 38

V
APÉNDICE A3 ................................................................................................................................ 40
APÉNDICE A4 ................................................................................................................................ 47
APENDICE A5 ................................................................................................................................ 49

VI
ÍNDICE DE FIGURAS
Figura 1. Diagrama que muestra el efecto Seebeck entre dos metales (color azul y rojo) con uniones
en dos diferentes temperaturas (T1 y T2). La diferencia de temperaturas genera una corriente eléctrica
(figura superior). El potencial eléctrico V asociado a esta corriente se relaciona con (T2-T1) a través
del coeficiente Seebeck α (figura inferior). ......................................................................................... 2
Figura 2. Utilizando una juntura Peltier entre dos metales (color azul y rojo) y suministrando un
potencial eléctrico Vin , una corriente I se hace fluir a través del circuito, generando un aumento de la
temperatura en una de las junturas y una disminución de la temperatura en la otra juntura (T1 y T2). 4
Figura 3. Plantas estudiadas en este trabajo: a) Ruellia nudiflora, b) Epipremnum aureum y c) Ficus
benjamina. ........................................................................................................................................... 7
Figura 4. Vista general del montaje experimental: a) Planta Ruellia nudiflora, b) Baño recirculador,
c) Chasís cDAQ-9172 con módulos NI9203/NI9205 y d) Equipo de cómputo con el software Labview
SignalExpress 2009 instalado............................................................................................................ 12
Figura 5. Montaje experimental con seis diferentes plantas de Ruellia nudiflora dentro de la cámara
climática, acondicionadas con dos niveles de iluminación (100 y 40%). ......................................... 15
Figura 6. Vista general del montaje experimental de conexiones múltiples: a) Especie Ficus
benjamina, b) Módulo de conexiones, c) Multímetro con sistema de adquisición de datos y d) Equipo
de cómputo con el software Spyder Python 2.7 instalado. ................................................................ 16
Figura 7. Conexiones múltiples en una planta de la especie Ficus benjamina. a) Termopar I (sesión
1), b) Termopar I (sesión 2) y c) Termopar II (sesiones 1 y 2). Las letras mayúsculas (A-X) indican
puntos de conexión y las letras en rojo (Vi) significan el potencial eléctrico entre dos puntos de
conexión. ........................................................................................................................................... 17
Figura 8. Mediciones de temperatura realizadas, Tplanta y Traíz con respecto al tiempo para un
experimento típico. ............................................................................................................................ 19
Figura 9. Gráfica mostrando la diferencia de las dos temperaturas (ΔT = Tplanta – Traíz) y la diferencia
de potencial eléctrico (ΔV) obtenidas con respecto al tiempo, con su respectivo valor promedio (líneas
rectas). ............................................................................................................................................... 20
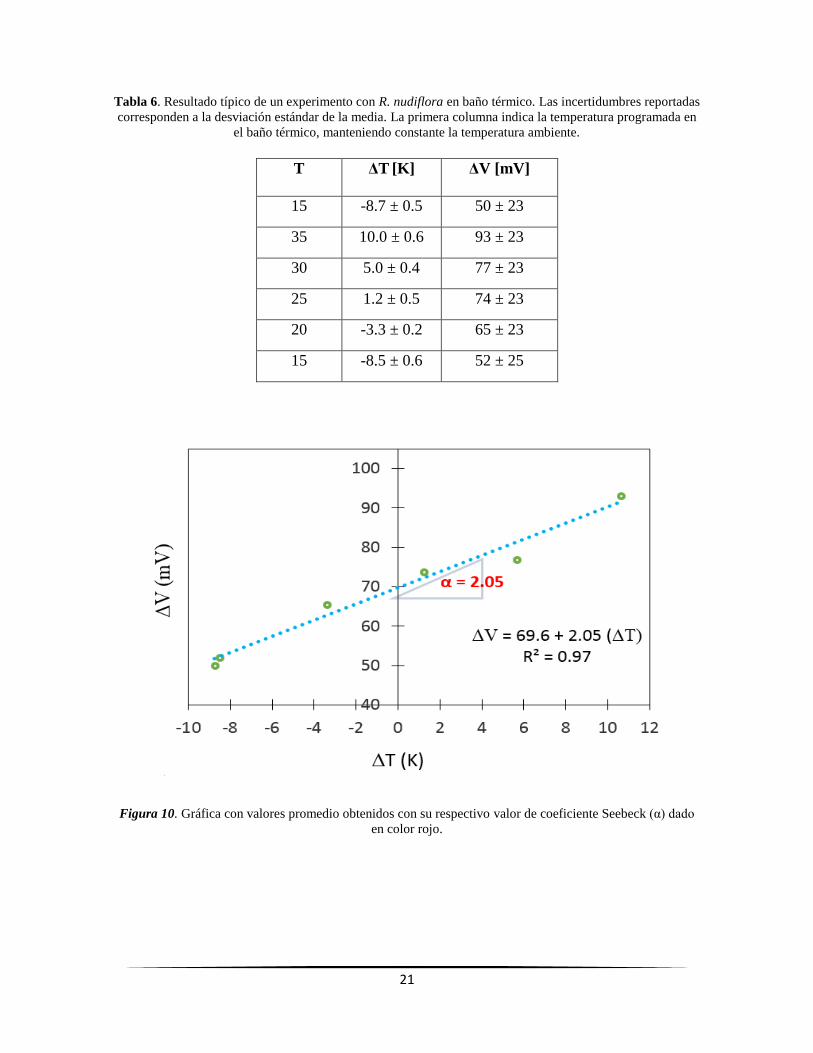
Figura 10. Gráfica con valores promedio obtenidos con su respectivo valor de coeficiente Seebeck
(α) dado en color rojo. ....................................................................................................................... 21
Figura 11. Resultado típico de un experimento con E. aureum en el baño térmico con distintas
distancias de conexión (L1 = 13 cm, L2 = 23 cm, L3 = 29 cm, L4 = 37 cm, L5 = 47 cm). ........... 25
Figura 12.Gráfica que muestra la correlación entre la longitud y el valor promedio de los potenciales
eléctricos medidos para cinco longitudes en tres diferentes posiciones. ........................................... 26
Figura 13. Gráficas obtenidas durante cada experimento: Vista general de las temperaturas medidas
T1 y T2 (superior izquierdo) y de T2-T1 (superior derecho) y de los puntos de conexión de potencial
eléctrico en la planta (V1-V12). ........................................................................................................ 27
Figura 14. Correlación entre el tiempo, la temperatura T2 y la respuesta eléctrica obtenida en la rama
más joven de la planta (mediciones de V1). ...................................................................................... 28
Figura 15. Correlación en el tiempo, de las temperaturas T1, T2 y T2-T1 respectivamente, con la
respuesta eléctrica obtenida. Los círculos rojos muestran regiones en el tiempo donde estas
correlaciones son más notorias. ......................................................................................................... 29

VII
ÍNDICE DE TABLAS
Tabla 1. Nombres comunes de Ruellia nudiflora en diferentes Estados de México. ......................... 8
Tabla 2. Experimentos con Ruellia nudiflora a diferentes condiciones en baño térmico. ............... 13
Tabla 3. Posición de la planta E.aureum para cada experimento. .................................................... 14
Tabla 4. Experimentos con la planta Ruellia nudiflora con diferentes condiciones en la cámara
climática, mostrando los diferentes grupos de plantas. ..................................................................... 15
Tabla 5. Descripción de las condiciones de cada experimento para la planta F. benjamina. La
temperatura T corresponde a la del agua añadida. ............................................................................ 18
Tabla 6. Resultado típico de un experimento con R. nudiflora en baño térmico. Las incertidumbres
reportadas corresponden a la desviación estándar de la media. La primera columna indica la
temperatura programada en el baño térmico, manteniendo constante la temperatura ambiente. ...... 21
Tabla 7. Promedios del valor absoluto del coeficiente Seebeck de los experimentos con R. nudiflora
en tres distintas condiciones experimentales. La incertidumbre reportada corresponde a la desviación
estándar calculada por el método de mínimos cuadrados. En la última columna se muestra el promedio
general para los trece experimentos; la incertidumbre reportada corresponde a la desviación estándar
de la media. ....................................................................................................................................... 22
Tabla 8. Promedio de los valores absolutos del coeficiente Seebeck de los experimentos con
R.nudiflora en cámara climática a diferentes condiciones: porcentaje de iluminación y dirección del
cambio de la temperatura ambiente. La incertidumbre reportada corresponde a la desviación estándar
de la media. ....................................................................................................................................... 23
Tabla 9. Promedio de los valores absolutos del coeficiente Seebeck de los experimentos con R.
nudiflora en cámara climática, clasificados por grupos de plantas en series de tiempo, marcado entre
paréntesis en la segunda columna. Todos los experimentos corresponden al control de la temperatura
ambiente absoluta. La incertidumbre reportada corresponde a la desviación estándar de la media. 24

1
INTRODUCCIÓN
Este trabajo de investigación explora la electrofisiología de los sistemas vegetales, es
decir, la respuesta eléctrica proporcionada por las plantas; esta respuesta se puede deber a
varios factores relacionados con su fisiología, tanto en el metabolismo normal de la planta
como frente a algún tipo de estrés. Cuando esta respuesta es causada por una diferencia de
temperatura, se le conoce como efecto Seebeck, el cual es un fenómeno termoeléctrico.
1.1. Termoelectricidad
La termoelectricidad es un fenómeno que fue descubierto en 1821 por Thomas Johann
Seebeck, quien observó una corriente eléctrica dentro de un circuito cerrado hecho de
diferentes metales con sus uniones mantenidas a diferente temperatura. El efecto Seebeck es
la formación de una diferencia de potencial originado por una diferencia de temperaturas.
Este efecto explica el acoplamiento de dos potenciales termodinámicos, el potencial
electroquímico y la temperatura, y es la base del funcionamiento de los termopares.
En 1834, Jean Peltier observó que dos flujos, uno de calor y el otro de una corriente
eléctrica, podrían ser acoplados bajo condiciones isotérmicas. Este efecto, llamado Peltier,
tiene aplicación práctica en dispositivos de refrigeración pequeños, teniendo la ventaja, a
diferencia de los refrigeradores basados en la compresión y descompresión de gases, de no
tener partes móviles que se desgasten [1]. Más tarde, en 1855 William Thompson, futuro
Lord Kelvin, usando argumentos termodinámicos, unificó los efectos Seebeck y Peltier.

2
1.1.1. Efecto Seebeck
El efecto Seebeck es la conversión de diferencias de temperatura directamente a
electricidad. Seebeck descubrió que la aguja de una brújula se desviaba cuando se formaba
un circuito cerrado de dos metales unidos en dos lugares con una diferencia de temperatura
entre las uniones (Figura 1). Esto se debe a que los metales responden diferentemente a la
diferencia de temperatura, creando una corriente de circuito, que a su vez produce un campo
magnético. En ese momento, Seebeck no reconoció allí una corriente eléctrica implicada, así
que llamó al fenómeno “efecto termomagnético”, pensando que los dos metales quedaban
magnéticamente polarizados por el gradiente de temperatura.
Figura 1. Diagrama que muestra el efecto Seebeck entre dos metales (color azul y rojo) con uniones en dos
diferentes temperaturas (T1 y T2). La diferencia de temperaturas genera una corriente eléctrica (figura
superior). El potencial eléctrico V asociado a esta corriente se relaciona con (T2-T1) a través del coeficiente
Seebeck α (figura inferior).

3
El voltaje creado es del orden de varios microVoltios por Kelvin. El voltaje obtenido
puede ser derivado de:
𝑽 = ∫ (𝑺𝑩(𝑻) − 𝑺𝑨(𝑻)) 𝒅𝑻𝑻𝟐
𝑻𝟏
donde SA y SB son los coeficientes Seebeck de los metales A y B en función de la temperatura,
y T1 y T2 son las temperaturas de las dos uniones. Los coeficientes Seebeck no siempre son
lineales en función de la temperatura, y dependen de la temperatura absoluta, el material y la
estructura molecular de los conductores. Si los coeficientes Seebeck son efectivamente
constantes para el rango de temperatura medido, la fórmula anterior puede aproximarse
como:
𝑽 = (𝑺𝑩 − 𝑺𝑨) ∙ (𝑻𝟐 − 𝑻𝟏)
Si la diferencia de temperatura ΔT entre los dos extremos de un material es pequeña,
entonces el coeficiente Seebeck de un material se define (aproximadamente) como [2]:
𝜶 =𝚫𝑽
𝚫𝑻
1.1.2. Efecto Peltier
El efecto Peltier hace referencia a la creación de una diferencia de temperatura debida a
la diferencia de voltaje eléctrico. Sucede cuando una corriente se hace pasar por dos metales
o semiconductores conectados para formar lo que se conoce como junturas de Peltier (Figura
2). La corriente propicia una transferencia de calor de una juntura a la otra: una se enfría en
tanto que otra se calienta [3].

4
Figura 2. Utilizando una juntura Peltier entre dos metales (color azul y rojo) y suministrando un potencial
eléctrico Vin , una corriente I se hace fluir a través del circuito, generando un aumento de la temperatura en
una de las junturas y una disminución de la temperatura en la otra juntura (T1 y T2).
El calor Peltier absorbido por la unión inferior por unidad de tiempo, �̇�, es igual a:
�̇� = 𝝅𝑨𝑩𝑰 = (𝝅𝑩 − 𝝅𝑨)𝑰̇
Donde 𝜋 es el coeficiente Peltier 𝜋𝐴𝐵 de todo el termopar, 𝜋𝐴 y 𝜋𝐵 son los
coeficientes de cada material. Los coeficientes Peltier representan el intercambio de calor por
unidad de carga a través de un material dado.
En 1854 Thomson halló una relación, ahora llamada relación de Thomson o Kelvin,
entre los coeficientes correspondientes. La temperatura absoluta T, el coeficiente de Peltier
𝝅 y el coeficiente Seebeck α se relacionan en la siguiente expresión[1]:
𝝅 = 𝜶 ∙ 𝑻

5
1.2. Fisiología y electrofisiología vegetal
La fisiología vegetal es la ciencia que estudia cómo funcionan las plantas, esto es, qué
ocurre en las plantas que las mantiene vivas. Explica a través de leyes físicas y químicas
cómo las plantas son capaces de utilizar la energía de la luz para, a partir de sustancias
inorgánicas, sintetizar moléculas orgánicas con las cuales se construyen complejas
estructuras que forman el cuerpo de la planta. Pero el aspecto más importante no es el cúmulo
de procesos físicos y químicos que tienen lugar en cada punto concreto de la planta y en cada
momento de su programa de desarrollo, sino cómo se integran dichos procesos en el espacio
y en el tiempo y su modulación por el medio ambiente para llevar a buen término el desarrollo
de la planta[4].
La electrofisiología celular es la rama de fisiología encargada de estudiar la actividad
eléctrica en diferentes estructuras orgánicas y su respuesta ante estímulos eléctricos. La
actividad eléctrica está relacionada estrechamente con los procesos metabólicos en los seres
vivos y los iones, son los encargados de transportarla entre los diferentes elementos
constitutivos de los sistemas biológicos a través de procesos como el de difusión. Los
procesos finales de la transformación de energía se realizan en la célula, la cual está aislada
de su entorno por una membrana semipermeable que controla el paso de las sustancias
eléctricamente cargadas[5].
Un fenómeno que tiene gran impacto en la respuesta fisiológica de las plantas y su
electrofisiología celular es el estrés. El estrés se identifica como una desviación significativa
de las condiciones óptimas para la vida. Dichas condiciones ocasionan cambios en todos los
niveles funcionales de los organismos. Desde un punto de vista biológico, el estrés tiene una

6
connotación más amplia, refiriéndose a los cambios ambientales que alteran al estado
fisiológico de las plantas. El estrés es el conjunto de respuestas bioquímicas o fisiológicas
que definen un estado particular del organismo diferente al observado bajo un rango de
condiciones óptimas. Son manifestaciones fenotípicas de estrés las deformaciones como el
amarillamiento, manchas, necrosis, etcétera. Otras menos obvias requieren técnicas
especiales para su detección, como la baja asimilación enzimática, inducción a transmisión
de genes, cambios en la composición química, etcétera. Múltiples factores ambientales
inducen estados de estrés en plantas. Existen tres tipos de estrés: ambiental (hídrico, por alta
y baja temperatura, por salinidad, etc.), fisiológico (hormonal, tasas de fotorespiración, etc.)
y bioquímico (por factores abióticos, genes de resistencia, etc.)[6].
La detección de potenciales eléctricos en las plantas indica que la señalización eléctrica
es un sistema importante para transmitir información a través de largas distancias entre sus
órganos. La razón por la que las plantas han desarrollado vías para la transmisión de la señal
eléctrica está probablemente relacionada con su necesidad de responder rápidamente a los
factores de estrés ambiental [7]. La excitación electroquímica es causada por flujos iónicos a
través de la membrana plasmática de la célula [8], creando una modificación de carga
eléctrica en la propia membrana, así como una carga diferencial en cada lado. Este trans-
potencial de la membrana es la diferencia de tensión (o diferencia de potencial eléctrico)
entre el interior y exterior de una célula (Vint – Vext).
La termoelectricidad como parte de procesos no isotérmicos en organismos vivos, ya se
ha investigado; sin embargo, a excepción del caso de los insectos como avispas y abejas [9-
11] , para los cuales han sido analizados efectos termoeléctricos, otros estudios simplemente
informaron el registro del aumento de la temperatura en las hojas de Colocasia odora usando

7
un termopar [12], Otro reporte muestra un análisis de piroelectricidad, erróneamente tomado
como termoelectricidad [13].
1.3. Especies vegetales
Las preocupaciones por el deterioro ambiental, el aumento de la demanda de alimentos,
y en general de biomasa, son algunos de los estímulos para el estudio de las funciones de las
plantas, ya que sirven de filtro para la contaminación ambiental, regulan la temperatura,
generan oxígeno, y, por consiguiente, contribuyen en la reducción del calentamiento global.
En los últimos años, este estudio se ha visto favorecido por un gran avance en las técnicas de
investigación, de tal manera que muchas funciones de las plantas se pueden explicar hoy día
con un detalle físico molecular que era inconcebible en la década de 1980 [14].
Para este trabajo de investigación se utilizaron tres especies diferentes (Figura 3): Ruellia
nudiflora, Epipremnum aureum y Ficus benjamina, las características relevantes para este
estudio se explican más a detalle a continuación.
Figura 3. Plantas estudiadas en este trabajo: a) Ruellia nudiflora,
b) Epipremnum aureum y c) Ficus benjamina.

8
1.3.1. Ruellia nudiflora
Esta especie pertenece a la familia Acanthaceae. Conocida con distintos nombres
comunes (Tabla 1); es una hierba perenne que normalmente mide de 20 a 30 cm de altura. Las
hojas son alargadas de color verde oscuro. Las flores tienen forma de trompeta con
hendiduras, son de color púrpura y forman ramilletes llamativos. Sus frutos son alargados y
pequeños parecidos a vainas. Se distribuye desde Texas hasta el sureste de México y habita
en clima cálido desde el nivel del mar hasta los 160 msnm [15]. Generalmente se encuentra
creciendo sobre todo en áreas perturbadas, y bajo una amplia gama de condiciones climáticas
y del suelo. Tiene una amplia distribución en la Península de Yucatán [16].
Tabla 1. Nombres comunes de Ruellia nudiflora en diferentes Estados de México.
Nayarit jarisha (cora)
Quintana Roo
cheilahsuuki, che'suk, kabalya ' axnik, kabaya ' axnik, pix-
ulum, xkabalyaxnich (maya)
Yucatán
berranco xiw, kabal ya ' ax niik, kaanba ya' ax
niik,kaanban xa' an (maya).
1.3.2. Epipremnum aureum
Esta especie pertenece a la familia Araceae. Conocida con varios nombres comunes
(“teléfono”, "potus", "pothos" o "potos"), es una liana que puede alcanzar 20 m de alto, con
tallos de hasta 4 cm de diámetro. Trepa mediante raíces aéreas que se enganchan a las ramas
de los árboles. Las hojas son perennes, alternas y acorazonadas, de color verde brillante y
con manchas de color crema en las especies variegadas. Las hojas son enteras en las plantas

9
jóvenes, pero irregularmente pinnadas en las maduras y de hasta 1 m de largo por 45 cm de
ancho. En las plantas jóvenes las hojas no superan los 20 cm de largo y usada como planta
de interior el tamaño de las hojas se reduce considerablemente (10 cm). Las flores surgen en
una espata de más de 23 cm de largo [17].
Requieren una exposición luminosa, pero sin sol directo, ya que se pueden quemar las
hojas. En cambio, si hay poca luz, las hojas pierden lustre, y en las especies variegadas
pierden los tonos crema característicos. Es una planta tropical, pero se usa como planta de
interior en los climas templados. La temperatura de crecimiento óptimo está entre 18 y 24ºC;
temperaturas superiores a 26ºC frenan el crecimiento de la planta. Crece preferentemente en
sustratos permeables, no salinos, no alcalinos y ricos en materia orgánica como en micro-
nutrientes.
1.3.3. Ficus benjamina
El género Ficus pertenece a la familia Moraceae; que posee entre 700 y 2000 especies
en todo el mundo [18, 19], muchas de las cuales son utilizadas como ornamentales [20].
Mundialmente se le conoce como “higuera llorona”, aunque en México se le conoce
simplemente como “Ficus” o “Benjamina” [21, 22].
La especie F.benjamina crece de manera natural en países como Australia, Borneo,
China, Filipinas, India, Indonesia y Malasia. Es una especie de distribución tropical, ya que
para su mejor desarrollo requiere temperaturas entre 30 y 33°C, aunque puede sobrevivir en
temperaturas más bajas, siempre y cuando no sean menores a los 4°C durante el invierno
[22].

10
En México y el mundo son utilizadas en ornamentación urbana y en interiores [22]. La
propagación de esta especie se ha realizado principalmente por acodos aéreos; sin embrago,
como resultado del aumento en la demanda de plantas más pequeñas para usos en interiores,
durante los últimos años, esta práctica ha cambiado por la del enraizado de esquejes.
1.4. Objetivo
El objetivo general de este trabajo de tesis fue explorar las respuestas eléctricas de tres
especies vegetales sometidas a diferencias de temperatura entre la raíz y la parte aérea de la
planta, así como evaluar el efecto sobre la respuesta eléctrica de la planta por el tipo de
sustrato, iluminación, edad de la planta y altura en el tallo a la cual se realizó la medición.
Específicamente para R. nudiflora se propuso observar la influencia de las diferentes
condiciones experimentales en el correspondiente valor del coeficiente Seebeck. Para E.
aureum se planteó evaluar la respuesta eléctrica en función de la longitud del tallo.
Finalmente, para F. benjamina se consideró diferenciar la respuesta eléctrica de diferentes
órganos o tejidos de la planta.

11
2. SECCIÓN EXPERIMENTAL
2.1. Materiales y Métodos
2.1.1. Descripción general de los experimentos
Se establecieron tres diferentes metodologías: baño térmico, cámara climática y
conexiones múltiples, dependiendo de la disponibilidad de infraestructura disponible en cada
laboratorio. En los experimentos se utilizaron dos termopares, uno para la medición de la
temperatura de la raíz y el otro para medir la temperatura de la parte aérea de la planta. De
la misma manera se utilizaron dos o más electrodos para la medición del potencial eléctrico.
Cada montaje experimental obtuvo información distinta y complementaria. Las
especificaciones se mencionarán en cada una de las siguientes secciones.
2.1.2. Baño térmico
Esta metodología fue utilizada para estudiar las especies R. nudiflora y E. aureum. El
montaje experimental consistió de un baño recirculador (PolyScience, modelo 9712) que
controlaba la temperatura de la raíz de la planta, que estuvo sumergida dentro de este, en un
vaso de precipitado. Se utilizaron dos termopares tipo J para monitorizar la temperatura de
la raíz y del laboratorio, al igual que dos electrodos que miden el potencial eléctrico generado.
Los electrodos fueron dos cables de cobre tipo UTP (Unshielded Twisted Pair) que se
conectaron a la raíz y al tallo de la planta, respectivamente (Figura 4). Para una mejor
conductividad del electrodo conectado al tallo, se utilizó como interfase un papel absorbente
humedecido constantemente con agua destilada. También se utilizó un chasís que funcionó
como interfaz de los sensores con dos módulos de National Instruments (uno para medición

12
de temperatura y otro para medición del potencial eléctrico) conectados a una computadora
portátil. El programa utilizado para la adquisición de datos fue el Labview Signal Express
(National Instruments) el cual permite mediciones automáticas en tiempo real.
Figura 4. Vista general del montaje experimental: a) Planta Ruellia nudiflora,
b) Baño recirculador, c) Chasís cDAQ-9172 con módulos NI9203/NI9205 y
d) Equipo de cómputo con el software Labview SignalExpress 2009 instalado.
Para R. nudiflora se realizaron un total de trece experimentos con diferentes condiciones
a los que estuvieron expuestas las plantas. Se trabajaron cinco temperaturas (15, 20, 25, 30,
35 °C) para el baño recirculador. En seis experimentos, la variación de temperatura del baño
recirculador fue de forma ascendente y en siete experimentos de forma descendente (ver Tabla
2). En la mayoría de los experimentos se utilizó como sustrato, una solución acuosa 3.6 M
KCl y un medio de cultivo Sueoka, alto en sales (Apéndice A1).
a
b
c
d

13
Tabla 2. Experimentos con Ruellia nudiflora a diferentes condiciones en baño térmico.
Experimento Intervalo de
Temperatura (°C)
Sustrato
1 [15-35]
Solución acuosa
3.6 M KCl
2 [35-15]
3 [35-15]
4 [35-15]
5 [15-35]
6 [35-15]
7 [15-35]
8 [35-15]
9 [15-35]
10 [15-35]
11 [35-15]
12 [15-35] Medio Sueoka
13 [35-15]
Para E. aureum se realizaron un total de seis experimentos (Tabla 3), con un intervalo
de temperatura de 15 a 33°C, con incrementos de 3°C. Con esta planta se tomaron mediciones
de potencial eléctrico a diferentes distancias a lo largo del tallo (13, 23, 29, 37 y 47cm de
altura con respecto al inicio de la raíz) y en tres posiciones distintas: vertical hacia arriba,
horizontal y vertical hacia abajo, para elucidar algún efecto de la fuerza de gravedad en la
respuesta eléctrica de la planta. Los electrodos y termopares para esta planta, fueron los
mismos durante los seis experimentos.

14
Tabla 3. Posición de la planta E.aureum para cada experimento.
Posición
Vertical hacia arriba
Horizontal
Vertical hacia abajo
Experimentos 1 y 4 2 y 5 3 y 6
2.1.3. Cámara climática
Para este montaje experimental se utilizó una cámara climática (KBW 720, BINDER), la
cual controla la temperatura ambiente y tiene la ventaja de que se le puede variar la intensidad
de iluminación. La iluminación está dada por cinco lámparas fluorescentes en cada uno de
los dos niveles de la cámara, siendo posible iluminar cada nivel de manera independiente al
100, 60, 40 y 0 %. Las plantas utilizadas fueron de la especie R. nudiflora y sus raíces
estuvieron siempre dentro de la maceta, es decir, en suelo (Figura 5). Para lograr un
aislamiento parcial de la raíz con respecto de la temperatura de la cámara, se colocaron las
macetas dentro de una nevera de unicel, evitando un cambio muy brusco en la temperatura
de la raíz al hacer el cambio de la temperatura de la cámara. Cabe mencionar, que se utilizaron
treinta plantas diferentes clasificadas en cinco grupos (A, B, C, D y E), cada uno con seis
plantas, que en ocasiones se utilizaron en más de un experimento. Para la adquisición de
datos se siguió la misma metodología que en el montaje de baño químico descrito en la
sección anterior.

15
Figura 5. Montaje experimental con seis diferentes plantas de Ruellia nudiflora dentro de la cámara
climática, acondicionadas con dos niveles de iluminación (100 y 40%).
En total se realizaron catorce experimentos. En la Tabla 4, se describen todos los
experimentos con las diferentes condiciones a las que estuvieron expuestas las plantas. En
cada experimento se trabajaron siete diferentes temperaturas en un intervalo establecido,
manteniendo constante la iluminación en dos niveles (100% y 40%, o 60% y 0%).
Tabla 4. Experimentos con la planta Ruellia nudiflora con diferentes condiciones en la cámara climática,
mostrando los diferentes grupos de plantas.
Experimento Grupo Intervalo de
Temperatura (°C)
Iluminación (%) *
1
A
[10 -40] 100 / 40
2 [10 -40] 100 / 40
3 [10 -34] 100 / 40
4 [35-11] 100 / 40
5 B [35-11] 100 / 40
6 C [10 -34] 100 / 40
7 D
[10 -34] 100 / 40
8 [10 -34] 60 / 0
9 E [35-11] 60 / 0
10 B [35-11] 60 / 0
11 C [10-34] 100 / 40
12
D
[10-34] 100 / 40
13 [35-11] 60 / 0
14 [10-34] 100 / 40
*100% = 5 lámparas

16
2.1.4. Conexiones múltiples
El sistema de medición es presentado en la Figura 6. Se utilizaron dos termopares para la
medición de la temperatura y 24 electrodos que fueron conectados a una planta de la especie
Ficus benjamina. Los sensores fueron conectados a un módulo que a su vez estaba conectado
a un multímetro para la medición del potencial eléctrico. Los datos de medición fueron
registrados de manera automática a través de una computadora utilizando un software
programado especialmente para tal propósito. Se realizaron dos sesiones, la diferencia entre
una y otra fue la posición del termopar I, ya que para la sesión 1, este estuvo midiendo la
temperatura del medio ambiente (aire) y para la sesión 2, se midió la temperatura del suelo.
Figura 6. Vista general del montaje experimental de conexiones múltiples: a) Especie Ficus benjamina, b)
Módulo de conexiones, c) Multímetro con sistema de adquisición de datos y d) Equipo de cómputo con el
software Spyder Python 2.7 instalado.
Fueron utilizados veinticuatro electrodos colocados en toda la planta de esta especie.
A diferencia de los electrodos utilizados en los experimentos de baño térmico y cámara
climática, los cables de cobre utilizados en este montaje experimental para hacer las
conexiones, tienen soldada una pequeña lámina de plata que es conectada a la planta
utilizando un gel especial para electrodos (Signalgel, Parker) con el cual aumenta la
b
c
a
d

17
conductividad eléctrica y se proporciona un contacto apropiado con la superficie de la planta.
También se utilizaron dos termopares, el primero (I) para medir la temperatura del medio
ambiente (T1) en la sesión 1, y durante la sesión 2 fue cambiado de posición colocándolo
directamente en el suelo (T1); y el segundo (II) situado sobre una hoja para medir la
temperatura de la planta (T2). Todas las conexiones fueron establecidas tal y como se muestra
en la Figura 7.
Figura 7. Conexiones múltiples en una planta de la especie Ficus benjamina. a) Termopar I (sesión 1), b)
Termopar I (sesión 2) y c) Termopar II (sesiones 1 y 2). Las letras mayúsculas (A-X) indican puntos de
conexión y las letras en rojo (Vi) significan el potencial eléctrico entre dos puntos de conexión.

18
En total se realizaron ocho experimentos, seis en la sesión 1 y dos correspondientes
a la sesión 2. En los experimentos se agregaron 100 mL de agua a tres diferentes temperaturas
(4, 21 y 40°C) tal y como se muestra en la Tabla 5. La duración de los experimentos fue desde
un mínimo de 24 horas hasta un máximo de 88 horas cada uno.
Tabla 5. Descripción de las condiciones de cada experimento para la planta F. benjamina. La
temperatura T corresponde a la del agua añadida.
Sesión Experimento T(°C)
1
1, 5 4
2, 4 40
3, 6 21
2 7 4
8 40
El software utilizado fue el Spyder (anteriormente Pydee), el cual es un entorno de
desarrollo multiplataforma de código abierto para la programación científica en el lenguaje
Python. En comparación con otros entornos de desarrollo, Spyder [23]tiene un conjunto
único de características, incluye soporte para herramientas interactivas para la inspección de
datos específicos. Se utiliza el lenguaje Python para el aseguramiento de la calidad del código
y la introspección, como pyflakes y Pylint. Se programó un código para hacer la adquisición
automática de datos de medición, tanto para los termopares como para los electrodos. El
código registra en tiempo real los valores de temperatura y potencial eléctrico, con lecturas
cada 30 s. Adicionalmente, el código permite graficar las mediciones en función del tiempo.

19
2.2. Resultados
Para un experimento típico, con cualquiera de las tres metodologías utilizadas, se
obtienen una gráfica (Figura 8) con las dos mediciones de temperatura realizadas, Tplanta y Traíz
con respecto al tiempo. En el baño térmico como en la cámara climática, se hicieron
mediciones estáticas fijando las temperaturas de la raíz o de la planta, en los experimentos
de conexiones múltiples se hicieron mediciones dinámicas, donde la variación de la
temperatura es una función del tiempo. Los experimentos con Ruellia nudiflora y
Epipremnum aureum fueron estáticos; los experimentos con Ficus benjamina fueron
dinámicos. En la Figura 9 se pueden visualizar los resultados para un experimento estático,
con la diferencia de las dos temperaturas (ΔT = Tplanta – Traíz) y la diferencia de potencial
eléctrico (ΔV) con respecto al tiempo, obteniendo un valor promedio de ambos valores
(líneas rectas en la figura).
Figura 8. Mediciones de temperatura realizadas, Tplanta y Traíz
con respecto al tiempo para un experimento típico.
19.9
20.0
20.1
20.2
20.3
20.4
20.5
0 100 200 300 400 500 600 700 800 900
T pla
nta
(°C
)
T raí
z(°
C)
tiempo (s)

20
Figura 9. Gráfica mostrando la diferencia de las dos temperaturas (ΔT = Tplanta – Traíz) y la diferencia de
potencial eléctrico (ΔV) obtenidas con respecto al tiempo, con su respectivo valor promedio (líneas rectas).
2.2.1. Ruellia nudiflora
Para un experimento típico con R. nudiflora en el baño térmico se muestran los resultados
en la Tabla 6: los valores promedio de T y los correspondientes valores promedio de V,
donde la incertidumbre asociada corresponde a la desviación estándar de la media. Al graficar
estos valores promedio, generalmente es posible observar una línea recta cuya pendiente
corresponde al valor del coeficiente Seebeck (Figura 10). Los detalles de estos resultados para
los trece experimentos con baño térmico se muestran en el Apéndice A2. Los valores de los
correspondientes coeficientes Seebeck (α) con sus características específicas se concentran
en la Tabla 7.
-4.5
-4.0
-3.5
-3.0
-2.5
-2.0
-1.5
-1.0
-0.5
0.0
-100
-75
-50
-25
0
25
50
75
100
125
0 100 200 300 400 500 600 700 800 900
ΔT
(K)
V
(m
V)
tiempo (s)
21
Tabla 6. Resultado típico de un experimento con R. nudiflora en baño térmico. Las incertidumbres reportadas
corresponden a la desviación estándar de la media. La primera columna indica la temperatura programada en
el baño térmico, manteniendo constante la temperatura ambiente.
T ΔT [K] ΔV [mV]
15 -8.7 ± 0.5 50 ± 23
35 10.0 ± 0.6 93 ± 23
30 5.0 ± 0.4 77 ± 23
25 1.2 ± 0.5 74 ± 23
20 -3.3 ± 0.2 65 ± 23
15 -8.5 ± 0.6 52 ± 25
Figura 10. Gráfica con valores promedio obtenidos con su respectivo valor de coeficiente Seebeck (α) dado
en color rojo.

22
Tabla 7. Promedios del valor absoluto del coeficiente Seebeck de los experimentos con R. nudiflora en tres
distintas condiciones experimentales. La incertidumbre reportada corresponde a la desviación estándar
calculada por el método de mínimos cuadrados. En la última columna se muestra el promedio general para los
trece experimentos; la incertidumbre reportada corresponde a la desviación estándar de la media.
Experimento Condiciones |𝜶|𝒑𝒓𝒐𝒎 (mV/K) |𝜶|𝒈𝒆𝒏𝒆𝒓𝒂𝒍 (mV/K)
1
Temperatura
ascendente. *
3.0 ± 0.9
2.6 ± 1.0
5
7
9
10
2
Temperatura
descendente. *
2.3 ± 1.0
3
4
6
8
11
12
Medio Sueoka. 2.6 ± 2.1 13
*En solución KCl.
De la Tabla 7, es posible observar que no hay diferencia significativa entre los valores del
coeficiente Seebeck calculados para cada una de las tres condiciones experimentales, es decir
no importa si la temperatura de la raíz se modifica de manera ascendente o descendente, o si
la raíz está sumergida en solución acuosa de KCl o medio Sueoka. De los trece experimentos
en baño térmico se puede calcular un coeficiente Seebeck promedio con su respectiva
desviación estándar: [2.6 ± 1.0] mV/K, con tres órdenes de magnitud mayor que los valores
de coeficiente Seebeck de los materiales en estado sólido [24]. Como se observa la desviación
estándar de la media es grande, esto es debido al ruido en la señal de medición del potencial
eléctrico, tal y como se mostró en la Figura 9 (color naranja) y en la Tabla 6 (ΔV). Estos errores
se propagan en el cálculo del correspondiente coeficiente Seebeck y en el uso de distintas

23
plantas. Además el coeficiente Seebeck encontrado para R.nudiflora es un valor
relativamente pequeño que es afectado considerablemente por la incertidumbre experimental.
Pero al tener este coeficiente tan pequeño, significa que el potencial eléctrico medido no
depende ampliamente de la diferencia de temperatura.
Para los experimentos en la cámara climática, los detalles de los resultados se muestran
en el Apéndice A3, donde se puede observar que de los 84 valores de coeficiente Seebeck
calculados, seis presentaron coeficientes de correlación (R2) menores a 0.29, por lo que no
fueron considerados en los análisis posteriores. Los valores de los correspondientes
coeficientes Seebeck (α) con sus características específicas se concentran en la Tabla 8.
Tabla 8. Promedio de los valores absolutos del coeficiente Seebeck de los experimentos con R.nudiflora en
cámara climática a diferentes condiciones: porcentaje de iluminación y dirección del cambio de la
temperatura ambiente. La incertidumbre reportada corresponde a la desviación estándar de la media.
Experimentos Iluminación (%) Tamb |𝜶|𝒑𝒓𝒐𝒎 (mV/K)
1,2,3,6,7,11,12,14 100 Ascendente 7.0 ± 2.8
4,5 100 Descendente 3.1 ± 2.0
8,9 60 Ascendente 5.0 ± 1.4
10,13 60 Descendente 2.0 ± 1.0
1,2,3,6,7,11,12,14 40 Ascendente 6.9 ± 3.0
4,5 40 Descendente 4.5 ± 2.9
8,9 0 Ascendente 4.0 ± 2.0
10,13 0 Descendente 2.7 ± 1.0
Como se puede observar en la Tabla 8 para todas las condiciones de iluminación probadas,
el valor del coeficiente Seebeck es mayor cuando la temperatura ambiente es ascendente y
menor cuando la temperatura ambiente es descendente. La mayor diferencia entre los valores
del coeficiente Seebeck es de 4.3 mV/K para 100% de iluminación, que sobrepasa los valores
de incertidumbre asociados. Cuando se comparan los experimentos de 100% y 40% de
iluminación (realizados de manera simultánea) puede notarse que no hay una diferencia

24
significativa en los valores del coeficiente Seebeck; lo mismo se observa al comparar las
iluminaciones de 60% y 0%. Las diferencias de los valores del coeficiente Seebeck en todos
los casos es menor que los valores de incertidumbre asociados. En general, se puede observar
que los valores del coeficiente Seebeck son mayores en los experimentos de la cámara
climática que en los del baño térmico. También se pueden analizar los resultados por grupos
de plantas donde se puede observar el efecto de la edad de la planta en el valor del coeficiente
Seebeck (Tabla 9). Tres grupos de plantas (A, C y D) se estudiaron en más de un experimento
con las mismas condiciones de iluminación y control de la temperatura; de esta manera fue
posible comparar el valor del coeficiente Seebeck en función del tiempo o de la edad de la
planta. En general se tiene que el valor del coeficiente Seebeck disminuye con el aumento de
la edad de las plantas, aunque el patrón no es constante en todos los experimentos.
Tabla 9. Promedio de los valores absolutos del coeficiente Seebeck de los experimentos con R. nudiflora en
cámara climática, clasificados por grupos de plantas en series de tiempo, marcado entre paréntesis en la
segunda columna. Todos los experimentos corresponden al control de la temperatura ambiente absoluta. La
incertidumbre reportada corresponde a la desviación estándar de la media.
Grupo Experimentos Iluminación (%) Tamb |𝜶|𝒑𝒓𝒐𝒎 (mV/K)
A
1 (día 0) 100
Ascendente 8.6 ± 3.8
40 9.0 ± 2.1
2 (1 día) 100
Ascendente 7.5 ± 4.5
40 8.5 ± 0.1
3 (5 días) 100
Ascendente 5.7 ± 1.2
40 6.7 ± 3.3
C
6 (día 0) 100
Ascendente 5.8 ± 1.6
40 2.8 ± 0.2
11 (7 días) 100
Ascendente 5.1 ± 3.2
40 5.8 ± 1.6
D
7 (día 0) 100
Ascendente 9.4 ± 2.9
40 10.6 ± 2.3
12 (11 días) 100
Ascendente 7.1 ± 1.0
40 7.2 ± 2.7
14 (14 días) 100
Ascendente 7.1 ± 2.4
40 4.5 ± 1.6

25
2.2.2. Epipremnum aureum
El resultado de un experimento típico con E. aureum en el baño térmico se muestran en
la Figura 11: los valores promedio de T y los correspondientes valores promedio de V de
las cinco longitudes a lo largo del tallo. En general se observa que el potencial eléctrico no
cambió de manera importante con los diferentes valores de T a lo largo del tallo, por lo que
en realidad el valor del coeficiente Seebeck se puede considerar como cero. En consecuencia,
por cada potencial eléctrico determinado a una longitud específica se obtuvo un promedio
para los valores obtenidos en cada T. Los detalles de estos resultados para los seis
experimentos con baño térmico se muestran en el Apéndice A4.
Figura 11. Resultado típico de un experimento con E. aureum en el baño térmico con distintas distancias de
conexión (L1 = 13 cm, L2 = 23 cm, L3 = 29 cm, L4 = 37 cm, L5 = 47 cm).
En general, se encuentra que el potencial eléctrico promedio a todas las temperaturas
aumenta con la longitud del tallo de la planta y también puede depender de la posición del
tallo. En ocasiones se encuentra una longitud característica por encima de la cual el potencial
eléctrico ya no varía significativamente (Figura 12).
-250
-200
-150
-100
-50
0
-10 -5 0 5 10 15
ΔV
(m
V)
ΔT (K)
L1
L2
L3
L4
L5

26
Figura 12.Gráfica que muestra la correlación entre la longitud y el valor promedio de los potenciales
eléctricos medidos para cinco longitudes en tres diferentes posiciones.
2.2.3. Ficus benjamina
En la Figura 13 se muestra un esquema general de los resultados típicos para un
experimento, con los cambios de valores de temperatura y potencial eléctrico en función del
tiempo, de manera global, y también clasificados por los diferentes tejidos u órganos de la
especie: tallo principal (V2, V5, V7 y V9), ramas (V1, V3, V4, V6, V8 y V11) y raíz con
tallo principal (V10 y V12). También se grafican los acercamientos del momento en que se
agrega agua. Los detalles de estos resultados para los ocho experimentos se muestran en el
Apéndice A5.
-250
-200
-150
-100
-50
0
10 20 30 40 50
ΔV
pro
med
io(m
V)
Longitud (cm)
Vertical hacia arriba
Horizontal
Vertical hacia abajo

27
Figura 13. Gráficas obtenidas durante cada experimento: Vista general de las temperaturas medidas T1 y T2
(superior izquierdo) y de T2-T1 (superior derecho) y de los puntos de conexión
de potencial eléctrico en la planta (V1-V12).
----- T1 ----- T2

28
Con los primeros experimentos se observó una respuesta electrofisiológica muy
importante para una parte en específico de la planta, que es su rama más joven (V1). En esta
parte se pudo percibir que en la mayoría de los experimentos había una correlación entre la
temperatura T2 y la respuesta eléctrica obtenida. En la Figura 14 se puede visualizar esta
respuesta. En menor grado, la respuesta del tallo en su altura intermedia (V7), también
presenta una correlación con el cambio de temperatura para la mayoría de los experimentos.
Figura 14. Correlación entre el tiempo, la temperatura T2 y la respuesta eléctrica obtenida en la rama más
joven de la planta (mediciones de V1).
Para la sesión 2, la correlación con la temperatura T2 continuaba en algunos intervalos
para V1 y V7; pero también pudieron encontrarse correlaciones en algunos intervalos con las
mediciones de la temperatura T1, al igual que para la diferencia de estas temperaturas (T2-
T1), como se observa en la Figura 15. Para las ramas más jóvenes (V1 y V3), así como para
los segmentos más altos del tallo (V2, V5 y V7), en general se observa una correlación parcial
de la respuesta eléctrica con la diferencia de temperatura.
tiempo (s)
tiempo (s)
V1
(m
V)
T2 (
°C)

29
Figura 15. Correlación en el tiempo, de las temperaturas T1, T2 y T2-T1 respectivamente,
con la respuesta eléctrica obtenida. Los círculos rojos muestran
regiones en el tiempo donde estas correlaciones son más notorias.
Las respuestas eléctricas de la planta ante los estímulos al agregar un volumen de
agua fueron generalmente más notorias en las partes más cercanas a la raíz (desde V8 hasta
V12). Para las conexiones V8, V10 y V12, los estímulos fueron observados casi siempre; sin
embargo, para las conexiones V9 y V11, éstos no siempre fueron detectados.
tiempo (s)
tiempo (s)
tiempo (s)
tiempo (s)
tiempo (s)
tiempo (s)
V1
(m
V)
V1
(mV
) V
1 (
mV
) T1
(°C
)
T2 (
°C)
T2 –
T1
(°C
)

30
2.3. Discusión
Durante décadas se ha tenido conocimiento de la existencia de una diferencia de potencial
eléctrico en partes de árboles, incluyendo floema [25], xilema [26], y las hoja [27], y la
relación entre esas partes con el suelo adyacente, aunque el origen de este potencial se ha
mantenido en controversia y es objeto de un considerable debate [28-30]. Estas diferencias
de potencial se han utilizado como intentos de supervisión de actividad eléctrica en la planta,
para lo cual se han planteado varias hipótesis, resaltando principalmente la que menciona el
mecanismo de “transmisión de potencial” [30], que está relacionado con la transpiración y el
flujo de savia. Para este trabajo de investigación se ha planteado la hipótesis de que la
diferencia de potencial eléctrico es debida a una diferencia de temperaturas, específicamente,
entre el tallo y la raíz de la planta, teniendo un efecto termoeléctrico. Aunque se pudo
observar que este efecto se presentó para R. nudiflora y F. benjamina, para E. aureum no fue
así. En lugar de ello, se encontró una relación con la longitud del tallo y con diferentes
órganos o tejidos de la planta, dando así, una respuesta eléctrica no necesariamente por la
diferencia de temperaturas, lo que conlleva a pensar que esta respuesta se origina en otros
lugares de la planta. Adicionalmente, es necesario recalcar que se encontraron correlaciones
de los potenciales eléctricos con las distintas condiciones experimentales y los valores
encontrados de coeficiente Seebeck que presentan una magnitud del orden de mV/K, es decir,
unas mil veces mayor que en un sistema inerte (del orden de µV/K) [24].
Por otra parte, se encuentra la presencia de un estrés como un factor externo a la planta
provocado por el medio ambiente cambiante, que ejerce una influencia que puede ser
importante. Es un concepto relativo y muy subjetivo, ya que una determinada situación
medioambiental puede resultar estresante para una especie y no para otras [6]. La inmovilidad

31
ha podido influir en que las plantas hayan adquirido y perfeccionado a lo largo de la evolución
mecanismos de autodefensa, por lo que, a lo largo de su ciclo vital, están expuestas a un gran
número de condiciones o factores estresantes. Por tal motivo, se estudió el efecto de la
incorporación de agua a distintas temperaturas, el cambio de suelo y la iluminación
proporcionada. Para R. nudiflora no se observó algún cambio relevante al someterla a un
estrés ambiental, causado en este caso por el cambio de sustrato, ya que no se encontraron
diferencias significantes en el valor del coeficiente Seebeck, esto es posiblemente debido a
que esta planta tiene una alta adaptabilidad al estrés y una alta tolerancia a la heterogeneidad
del medio ambiente [31]. Por otra parte, según los resultados obtenidos con respecto a la
variación de temperatura (para este caso, intervalos de temperatura pequeños) indican, tal y
como se reporta en la literatura [32], que esta planta tiene una mayor tolerancia a los cambios
de temperatura. También se observó que la iluminación era independiente del resultado
obtenido durante los experimentos, esto queda confirmado ya que esta planta tiene una
capacidad de crecer en diferentes condiciones de luz lo cual es una peculiaridad que ayuda a
determinar el éxito de su crecimiento [33]. Para F. benjamina, no se encontró una respuesta
eléctrica importante ante el estrés térmico sometido (intervalo de temperatura: 4 a 40°C), esto
coincide con estudios de otros autores donde han encontrado que es una especie de clima
cálido, ya que para su mejor desarrollo requiere temperaturas entre 30 y 33°C, aunque puede
sobrevivir en temperaturas bajas, siempre y cuando no sean menores a los 4°C [34, 35].

32
3. CONCLUSIONES Y PERSPECTIVAS
Todas las plantas estudiadas en este trabajo, presentaron una respuesta eléctrica
característica de cada especie y de las condiciones específicas de cada experimento. Para R.
nudiflora se encontró que el valor del Coeficiente Seebeck permanece constante en las
diferentes condiciones experimentales cuando se variaba la temperatura de la raíz en solución
acuosa. Por otro lado, al variar la temperatura ambiente cuando la raíz de la planta estaba en
suelo, el valor del coeficiente Seebeck dependía de la dirección del cambio de temperatura,
pero era independiente de la intensidad de la iluminación. En este caso también se encontró
que la edad de la planta influía en el valor del coeficiente Seebeck.
Para E. aureum, los cambios de temperatura no influyeron significativamente en los
potenciales eléctricos medidos, por lo que el valor de coeficiente Seebeck para esta especie
es cercano a cero en el intervalo de temperatura estudiado. Sin embargo, estos potenciales
eléctricos pueden depender de la longitud del tallo donde se hizo la medición, así como de la
posición de la planta.
Para F. benjamina se encuentran señales eléctricas distintas dependiendo de la parte de
la planta donde se hacen las mediciones. Para las mediciones en el tallo principal y la rama
más joven se encontraron mejores correlaciones de la temperatura de la planta con respecto
del potencial eléctrico, aunque no para todo el intervalo de tiempo del experimento. No se
pudo encontrar una respuesta importante de la planta por estrés térmico.
Con base en el análisis anterior, es posible sugerir un mejoramiento en la metodología
mediante el uso de sistemas de medición más precisos, conexiones eléctricas más estables
con la planta y un sistema de adquisición automática de datos más robusta. La posibilidad de

33
realizar experimentos dinámicos permite obtener mayor información en tiempo real.
También se pueden explorar diferentes condiciones experimentales como el grado de
humedad del ambiente en el que se encuentra la planta, o el tipo y la cantidad de nutrientes
accesibles. Finalmente, se sugiere analizar la respuesta eléctrica de otras especies vegetales
que pudieran ser de interés. Todo esto podrá dar la pauta para la generación de un modelo
que explique la respuesta electrofisiológica en los sistemas vegetales y para encontrar
posibles aplicaciones de este fenómeno.

34
BIBLIOGRAFÍA
1. Goldsmid, H.J., Introduction to thermoelectricity. 2010, Berlin: Springer. 2. Mahan, G., L. Lindsay, and D. Broido, The Seebeck coefficient and phonon drag in silicon.
Journal of Applied Physics, 2014. 3. Grenier, C., A. Georges, and C. Kollath, Peltier cooling of fermionic quantum gases. Physical
review letters, 2014. 4. Azcón-Bieto, J., Fundamentos de fisiología vegetal. 2008: McGraw-Hill Interamericana de
España S.L. 5. Chaparro, J., Instrumentación en electrofisiología celular vegetal: actividad eléctrica en
plantas. 2001. 6. Taiz, L. and E. Zeiger, Fisiologia vegetal. Fisiologia vegetal. Vol. 2. 2009: McGraw-Hill
Interamericana de España S.L. 7. Fromm, J. and S. Lautner, Electrical signals and their physiological significance in plants.
Plant, Cell & Environment, 2007. 8. Sten-Knudsen, Biological membranes: theory of transport, potentials and electric impulses.
. 2002. 9. Ishay, J.S., et al., Natural thermoelectric heat pump in social wasps. Physical review letters,
2003. 10. Galushko, D., et al., Electrical, thermoelectric and thermophysical properties of hornet
cuticle. Semiconductor science and technology, 2005. 11. Volynchik, S., et al., Presence of a thermoregulatory hot spot in the prothorax of the large
carpenter bee and the bumble bee. Microscopy research and technique, 2006. 12. Van Beek, A.B., C., Observations thermo-électriques sur l'élévation de température des fleurs
de Colocasia odora: Avec une planche lithographiée. 1838. 13. Muller, A.W., Thermoelectric energy conversion could be an energy source of living
organisms. Physics Letters A, 1983: p. 319-321. 14. Coll, J.B., Fisiología vegetal. 2005: Ediciones Pirámide. 15. Abdala-Roberts, L., et al., Noncorrelated effects of seed predation and pollination on the
perennial herb Ruellia nudiflora remain spatially consistent. Biological Journal of the Linnean Society, 2009.
16. Moreira, X., et al., Latitudinal variation in herbivory: influences of climatic drivers, herbivore identity and natural enemies. Oikos, 2015: p. 1444-1452.
17. Qu, L., et al., Thidiazuron promotes adventitious shoot regeneration from pothos (Epipremnum aureum) leaf and petiole explants. In Vitro Cellular & Developmental Biology-Plant, 2002: p. 268-271.
18. Condit, I.J., Ficus: the exotic species. 1 ed. 1969: University of California, Division of Agricultural Sciences.
19. Ibarra-Manríquez, G., et al., El género Ficus L.(Moraceae) en México. Botanical Sciences, 2012: p. 389-452.
20. Salim, R. and S. Khalaf, A role for decaying leaves in mitigating the harmful effects of acid rain. Journal of Environmental Science & Health Part A, 1994: p. 115-127.
21. Morton, J.F., 500 plants of south Florida. 1974, U.S.A.: Seeman. 22. Conover, C. and R. Poole, Relationships of culture and shipping temperature on interior
quality of Ficus benjamina. III International Symposium on Postharvest Physiology of Ornamentals, 1985: p. 245-250.

35
23. Kishimoto, K., M. Tsukamoto, and T. Koyanagi, Temperature dependence of the Seebeck coefficient and the potential barrier scattering of n-type PbTe films prepared on heated glass substrates by rf sputtering. Journal of applied physics, 2002: p. 5331-5339.
24. Zhang, Q., et al., Organic thermoelectric materials: Emerging green energy materials converting heat to electricity directly and efficiently. Advanced Materials, 2014. 26(40): p. 6829-6851.
25. Wright, J.P. and D.B. Fisher, Measurement of the sieve tube membrane potential. Plant Physiology, 1981: p. 845-848.
26. Wegner, L.H. and U. Zimmermann, Simultaneous recording of xylem pressure and trans‐root potential in roots of intact glycophytes using a novel xylem pressure probe technique. Plant, Cell & Environment, 1998: p. 849-865.
27. Ksenzhek, O., S. Petrova, and M. Kolodyazhny, Electrical properties of plant tissues: resistance of a maize leaf. Bulgarian Journal of Plant Physiology, 2004: p. 61-67.
28. Gibert, D., et al., Sap flow and daily electric potential variations in a tree trunk. Plant Science, 2006: p. 572-584.
29. Gensler, W. and T.L. Yan, Investigation of the causative reactant of the apoplast electropotentials of plants. Journal of The Electrochemical Society, 1988: p. 2991-2995.
30. Labady, A., et al., Plant bioelectrochemistry: effects of CCCP on electrical signaling in soybean. Bioelectrochemistry, 2002: p. 47-53.
31. Sakai, A.K., et al., The population biology of invasive specie. Annual review of ecology and systematics, 2001: p. 305-332.
32. Cervera, J.C., et al., Microhabitats, germination, and establishment for Mammillaria gaumeri (Cactaceae), a rare species from Yucatan. International Journal of Plant Sciences, 2006: p. 311-319.
33. Pons, T., Induction of dark dormancy in seeds: its importance for the seed bank in the soil. Functional ecology, 1991: p. 669-675.
34. Hummel, R. and C. Johnson, Freezing tolerance in the genus Ficus. HortScience (USA), 1985. 35. Mortensen, L. and G. Larsen, Effects of temperature on growth of six foliage plants. Scientia
horticulturae, 1989: p. 149-159. 36. Sueoka, N., K. Chiang, and J. Kates, Deoxyribonucleic acid replication in meiosis of
Chlamydomonas reinhardi: I. Isotopic transfer experiments with a strain producing eight zoospores. Journal of molecular biology, 1967: p. 47-66.

36
APÉNDICE A1
MEDIO SUEOKA (ALTO EN SALES) [36]
Hacer las siguientes soluciones stock:
1) Solución Beijirinck’s
Pesar 100 g de cloruro de amonio (NH4Cl), 4 g de sulfato de magnesio heptahidratado
(MgSO4 . 7H2O) y 2 g de cloruro de calcio dihidratado (CaCl2 . 2H2O). Diluir estos
tres compuestos y aforar a 1000 ml con agua destilada.
2) Solución de fosfatos
Pesar 144 g de fosfato monobásico de potasio (KH2PO4) y 288 g de fosfato dibásico
de potasio (K2HPO4 anhidro) . Diluir ambos compuestos y aforar a un 1000 ml con
agua destilada.
3) Elementos traza
Pesar los siguientes compuestos y diluir en el volumen especificado:
Sal EDTA disódica 50 g en 250 ml 𝐶𝑜𝐶𝑙2 ∙ 6𝐻2𝑂 1.61 g en 50 ml
𝑍𝑛𝑆𝑂4 ∙ 7𝐻2𝑂 22 g en 100 ml 𝐶𝑢𝑆𝑂4 ∙ 5𝐻2𝑂 1.57 g en 50 ml
𝐻3𝐵𝑂3 11.4 g en 200 ml (𝑁𝐻4)6𝑀𝑜7𝑂24 ∙ 4𝐻2𝑂 1.10 g en 50 ml
𝑀𝑛𝐶𝑙2 ∙ 4𝐻2𝑂 5.06 g en 50 ml 𝐹𝑒𝑆𝑂4 ∙ 7𝐻2𝑂 4.99 g en 50 ml
Recomendaciones: el EDTA debe ser diluirse en agua hirviendo y el sulfato ferroso
debe ser preparado de último para evitar la oxidación.
Modo de preparación: para 1000 ml de mezcla final de elementos traza mezcle bien
todas las soluciones (excepto el EDTA). Lleve a ebullición la mezcla anterior y
agregue la solución EDTA, observando que la mezcla tome un color verde. Diluida

37
perfectamente esta solución, separe de la placa de calentamiento y enfríe a 70 °C.
Mantener esta temperatura y agregar 85 ml de una solución caliente de KOH al 20%
(20 g de KOH en 100 ml de volumen final). No use NaOH para ajustar el pH.
Aforar la solución a 1000 ml, observando que inicialmente, ésta tenga un color verde
claro. Tape el recipiente con un algodón, déjelo de esta manera por 1 o 2 semanas y
agitar cada día. La solución puede tornarse eventualmente púrpura o rosa y queda un
precipitado café o de color semejante que puede ser removido por filtración
empleando papel filtro Whatman 1. Si es necesario, repetir la filtración hasta que la
solución quede clara. Guarde en refrigeración en alícuotas convenientes.
Nota: algunas personas disminuyen el tiempo para la formación del precipitado
haciendo burbujear la solución con aire filtrado. Mientras no tenga precipitados, la
solución sigue siendo utilizable.
Por cada litro de medio a preparar usar: 5 ml de Beijerinck’s, 5 ml de fosfatos y 1 ml de
elementos traza y diluir con agua desionizada o destilada.

38
APÉNDICE A2
RESULTADOS DE EXPERIMENTOS CON
Ruellia nudiflora EN BAÑO TÉRMICO
Experimento
1
T ΔT [K] ΔV [mV] Correlación *
35 10.6 ± 0.6 93± 23
𝜶 = 2.0 ± 0.2
R2=0.96
30 5.7 ± 0.4 77 ± 23
25 1.2 ± 0.5 74 ± 24
20 -3.4 ± 0.2 65 ± 23
15 -8.5 ± 0.7 52 ± 25
Experimento
2
35 8.5 ± 0.7 77 ± 30
𝜶 = 3.4 ± 1.3
R2=0.69
30 3.0 ± 0.5 78 ± 26
25 -1.8 ± 0.4 37 ± 29
20 -6.0 ± 0.2 8 ± 24
15 -10.9 ± 0.5 31 ± 24
Experimento
3
35 9.9 ± 0.6 96 ± 29
𝜶 =2.3 ± 0.9
R2= 0.68
30 5.8 ± 0.5 94 ± 33
25 0.9 ±0.4 58 ± 32
20 -3.8 ± 0.4 55 ± 27
15 -8.6 ± 0.4 63 ± 27
Experimento
4
35 11.2 ± 0.4 164 ± 28
𝜶 =3.3 ± 0.8
R2=0.89
30 6.4 ± 0.6 -----
25 1.6 ± 0.5 152 ± 27
20 -3.5 ± 0.5 122 ± 26
15 -8.0 ± 0.4 99 ± 27
Experimento
5
35 8.4 ± 0.5 -----
𝜶 =3.0 ± 0.5
R2=0.95
30 4.1 ± 0.6 36 ± 30
25 -0.4 ± 0.9 30 ± 28
20 -4.9 ± 0.7 9 ± 32
15 -8.6 ± 0.5 -0.2 ± 31
Experimento
6
35 10.3 ± 1.0 -3 ± 27
𝜶 =- 2.2 ± 1.5
R2= 0.42
30 5.6 ± 0.6 37 ± 25
25 0.5 ± 0.7 64 ± 30
20 -3.6 ± 0.7 34 ±31
15 -8.3 ± 0.6 48 ± 30
*Las unidades de los valores del coeficiente Seebeck son [mV/K]

39
Experimento
7
T ΔT [K] ΔV [mV] Correlación *
35 10.0 ± 0.4 85 ± 23
𝜶 = -2.9 ± 1.0
R2= 0.71
30 4.6 ± 2.0 118 ± 24
25 1.1 ± 0.5 109 ± 22
20 -3.9 ± 0.4 150 ± 22
15 -8.1 ± 0.4 133 ± 23
Experimento
8
35 9.9 ± 0.5 61 ± 26
𝜶 = -0.6 ± 0.4
R2= 0.52
30 5.6 ± 0.4 66 ± 27
25 1.4 ± 0.6 -----
20 -3.9 ± 0.6 78 ± 23
15 -8.9 ± 0.4 69 ± 16
Experimento
9
35 10.1 ± 0.5 9 ± 33
𝜶 = -4.5 ± 1.0
R2= 0.85
30 5.8 ± 0.5 51 ± 35
25 1.5 ± 0.5 72 ± 32
20 -2.9 ± 0.4 95 ± 39
15 -8.0 ± 0.4 91 ± 36
Experimento
10
35 11.3 ± 0.5 -0.5 ± 33
𝜶 = -2.5 ± 0.8
R2= 0.75
30 6.8 ± 0.5 9 ± 35
25 1.6 ± 0.4 18 ± 35
20 -1.9 ± 0.2 51 ± 35
15 -7.3 ± 0.5 37 ±37
Experimento
11
35 10.9 ± 0.4 -63 ± 29
𝜶 = -2.0 ± 1.0
R2= 0.67
30 6.5 ± 0.4 -53 ± 31
25 1.9 ± 0.6 -24 ± 32
20 -2.5 ± 0.5 -----
15 -7.3 ± 0.5 -29 ± 27
Experimento
12
35 10.6 ± 0.5 -46 ± 34
𝜶 = -4.1 ± 0.8
R2= 0.89
30 5.8 ± 0.5 -14 ± 33
25 0.8 ± 0.4 -14 ± 34
20 -3.6 ± 0.6 3 ± 37
15 -8.1 ± 0.5 43 ± 36
Experimento
13
35 10.6 ± 0.5 -606 ± 34
𝜶 = 1.1 ± 0.9
R2= 0.34
30 6.2 ± 0.5 -584 ± 33
25 -0.0 ± 0.7 -605 ± 41
20 -2.7 ± 0.1 -621 ± 32
15 -7.2 ± 0.5
-613 ± 32
*Las unidades de los valores del coeficiente Seebeck son [mV/K]

40
APÉNDICE A3
RESULTADOS DE EXPERIMENTOS CON Ruellia nudiflora EN CÁMARA CLIMÁTICA
E
X
P
E
R
I
M
E
N
T
O
1
Iluminación
100%
T ΔT [K] ΔV [mv]
Planta a Planta b Planta c
40 11.4 ± 0.3 -566 ± 39 -101 ± 26 46 ± 63
35 10.6 ± 0.4 -545 ± 292 -64 ± 55 48 ±49
30 9.6 ± 0.3 -485 ± 275 -50 ± 48 73 ± 30
25 8.0 ± 0.2 -439 ± 267 -32 ± 43 85 ± 37
20 5.5 ± 0.1 -367 ± 35 -20 ± 16 98 ± 53
15 2.0 ± 0.3 -400 ± 264 2 ± 46 120 ± 62
10 -2.8 ± 0.8 -370 ± 289 30 ± 31 135 ± 135
|𝜶| -13.0 ± 3.8 -6.7 ± 0.5 -6.2 ± 0.8
R2 0.69 0.97 0.92
Iluminación
40%
T ΔT [K] ΔV [mv]
Planta d Planta e Planta f
40 12.8 ± 0.4 -73 ± 328 -74 ± 53 -0.3 ± 74
35 12.0 ± 0.3 -49 ± 40 -32 ± 28 38 ± 33
30 10.9 ± 0.3 -21 ± 37 -14 ± 18 73 ± 25
25 9.0 ± 0.1 9 ± 36 -4 ± 11 87 ± 26
20 6.1 ± 0.07 46 ± 267 8 ± 44 111 ± 35
15 2.1 ± 0.05 86 ± 33 27 ± 14 163 ± 54
10 -3.1 ± 0.1 113 ± 31 73 ± 14 136 ± 112
|𝜶| -11.5 ± 1.3 -7.5 ± 1.1 -8.2 ± 2.1
R2 0.93 0.89 0.73
E
X
P
E
R
I
M
E
N
T
O
2
Iluminación
100%
T ΔT [K] ΔV [mv]
Planta a Planta b Planta c
40 11.4 ± 0.4 -36 ± 4 -70 ± 64 -120 ± 19
35 10.8 ± 0.4 -31 ± 3 -57 ± 66 -104 ± 12
30 9.5 ± 0.3 -15 ± 3 -48± 65 -102 ±11
25 7.6 ± 0.2 -5 ± 3 -46 ± 66 -56 ± 11
20 5.2 ± 0.2 -16 ± 16 -37 ± 64 8 ±12
15 1.5 ± 0.01 23 ± 20 -20 ± 67 23 ±10
10 -3.7 ± 0.1 73 ± 8 -14 ± 65 63 ±10
|𝛼| -6.7 ± 0.5 -3.4 ± 0.5 -12.4 ± 0.9
R2 0.97 0.91 0.97
Iluminación
40%
T ΔT [K] ΔV [mv]
Planta d Planta e Planta f
40 13 ±0.3 -123 ±44 -27 ±63 -125 ±20
35 12 ±0.4 -102 ±45 -39 ±68 -109 ±20
30 10 ±0.3 -99 ±44 -56 ±68 -89 ±15
25 8 ±0.2 -82 ±46 -62 ± 62 -69 ±15
20 6 ±0.1 -58 ±44 -50 ±64 -54 ±13
15 2 ±0.04 -18 ±40 -17 ±8 -25 ±14
10 -2 ±0.04 3 ±39 -33 ±8 70 ±37
|𝜶| -8.8 ± 0.5 -0.9 ± 1.3 -9.0 ±0.7
R2 0.98 0.09 0.97

41
E
X
P
E
R
I
M
E
N
T
O
3
Iluminación
100%
T ΔT [K] ΔV [mv]
Planta a Planta b Planta c
34 6.6±0.3 -19±9 -29±86 -70±7
30 5.8±0.3 -1±9 -21±81 -58±8
26 4.7±0.4 14±9 -16±82 -37±9
22 3.3±0.1 16±10 -7±81 -13±12
18 1.0±0.5 21±11 -4±81 6±7
14 -2.1±0.5 38±10 22±81 7±7
10 -5.8±0.3 42±10 36±81 18±7
|𝜶| -5.0 ± 0.9 -5.0 ± 0.3 -7.1 ± 1.5
R2 0.88 0.98 0.82
Iluminación
40%
T ΔT [K] ΔV [mv]
Planta d Planta e Planta f
34 9.4±0.6 -11±71 -43±45 -81±20
30 8.1±0.6 -5±68 -30±44 -84±20
26 5.8±0.5 7±68 -8±43 -65±21
22 3.3±0.6 20±67 22±43 -40±20
18 0.3±0.8 29±64 64±43 -17±22
14 -4.1±0.6 53±63 100±49 -12±25
10 -9.6±0.5 71±57 246±69 5±27
|𝜶| -4.5 ± 0.2 -10.6 ± 0.6 -5.1 ± 0.7
R2 0.99 0.98 0.92
E
X
P
E
R
I
M
E
N
T
O
4
Iluminación
100%
T ΔT [K] ΔV [mv]
Planta a Planta b Planta c
34 5.0±0.1 15±11 3±87 -87±10
30 2.4±0.4 8±9 0±87 -84±9
26 0.2±0.1 20±12 9±84 -80±13
22 -1.5±0.1 43±9 22±81 -42±11
18 -2.9±0.1 56±8 24±78 -14±9
15 -3.9±0.2 67±9 18±63 -8±9
11 -11.8±0.3 78±9 37±78 -3±9
|𝛼| -4.6 ± 1.0 -2.3 ± 0.4 -6.0 ± 1.6
R2 0.81 0.86 0.73
Iluminación
40%
T ΔT [K] ΔV [mv]
Planta d Planta e Planta f
7.5 7.5±38 17±0.2 -316±67 -104±45
3.1 3.1±39 19±0.4 -321±70 -109±47
-0.6 -0.6±37 33±0.1 -326±69 -102±47
-3.2 -3.2±37 43±0.1 -316±69 -73±48
-5.1 -5.1±37 63±0.2 -315±66 -53±48
-6.6 -6.6±38 72±0.3 ----- -46±49
-7.6 -7.6±38 86±0.3 -295±63 -37±48
|𝜶| -4.6 ± 0.8 -1.1 ± 0.8 -5.0 ± 1.0
R2 0.88 0.34 0.81

42
E
X
P
E
R
I
M
E
N
T
O
5
Iluminación
100%
T ΔT [K] ΔV [mv]
Planta a Planta b Planta c
35 7.1±0.4 -24±10 -42±236 -55±17
31 3.1±0.4 -24±29 -63±234 -29±32
27 0.1±0.2 -42±7 -61±225 -50±17
23 -2.3±0.1 -59±8 -81±224 -58±18
19 -4.2±0.1 -46±8 -51±222 -57±15
15 -5.7±0.1 -33±7 -42±217 -52±15
11 -6.9±0.2 -42±8 -70±217 -73±16
|𝜶| 1.5 ± 0.9 0.7 ± 1.2 1.4 ± 1.0
R2 0.34 0.06 0.29
Iluminación
40%
T ΔT [K] ΔV [mv]
Planta d Planta e Planta f
5.1 52±0.2 -180±36 4±301 5.1±12
1.2 60±0.2 -199±41 -12±304 1.2±13
-2.0 2±0.1 -240±31 -15±304 -2.0±13
-4.7 -29±0.1 -178±32 -29±298 -4.7±12
-6.8 -40±0.2 -122±29 -24±296 -6.8±12
-8.8 -39±0.2 -114±28 -15±294 -8.8±12
-10.3 -71±0.3 -117±30 -20±304 -10.3±12
|𝜶| 8.6 ± 1.2 -6.4 ± 3.1 1.4 ±0.6
R2 0.9 0.5 0.54
E
X
P
E
R
I
M
E
N
T
O
6
Iluminación
100%
T ΔT [K] ΔV [mv]
Planta a Planta b Planta c
34 8.1±0.1 -39±10 5±99 7±17
30 6.7±0.1 -51±9 -8±99 -3±17
26 4.9±0.1 -54±10 1±98 -17±18
22 2.7±0.1 -54±10 11±100 -40±18
18 -0.1±0.2 -53±11 15±99 -61±16
14 -3.6±0.3 -88±12 -85±106 -93±14
10 -7.1±0.1 -104±10 -90±104 -88±14
|𝜶| 4.0 ± 0.4 6.5 ± 2.2 7.0 ± 0.8
R2 0.96 0.63 0.94
Iluminación
40%
T ΔT [K] ΔV [mv]
Planta d Planta e Planta f
34 10.6±0.3 -22±56 -19±266 -38±13
30 8.3±0.3 -35±58 -55±268 -50±13
26 5.7±0.1 -33±50 -55±274 -52±12
22 2.6±0.1 -57±60 -72±275 -59±13
18 -1.1±0.1 -54±58 -76±267 -73±13
14 -5.5±0.3 -38±42 -136±274 -111±14
10 -10.2±0.2 -50±50 -86±260 -98±14
|𝜶| 3.0 ± 0.8 2.6 ± 0.7 2.8 ± 0.2
R2 0.82 0.86 0.98

43
E
X
P
E
R
I
M
E
N
T
O
7
Iluminación
100%
T ΔT [K] ΔV [mv]
Planta a Planta b Planta c
34 9.8±0.3 -51±24 -51±150 -71±44
30 8.2v0.2 -66±25 -62±149 -88±45
26 6.0±0.1 -85±25 -93±146 -120±45
22 3.2±0.0 -104±24 -128±144 -151±44
18 -0.2±0.1 -115±24 -144±139 -191±43
14 -4.5±0.3 -112±24 -191v138 -190±41
10 -9.5±0.3 -111±24 -192±143 -179±40
|𝜶| 6.4 ± 0.8 9.8 ± 0.6 12.1 ± 0.6
R2 0.95 0.98 0.99
Iluminación
40%
T ΔT [K] ΔV [mv]
Planta d Planta e Planta f
34 9.8±0.3 -73±24 -37±351 -34±27
30 8.0±0.2 -85±27 -51±357 -23±27
26 5.6±0.2 -121±24 -81±356 -42±26
22 2.8±0.1 -151±25 -111±356 -42±26
18 -0.4±0.1 -201±29 -148±355 -123±26
14 -4.1±0.2 -199±28 -171±338 -130±25
10 -8.5±0.2 -173±27 -161±346 -108±26
|𝜶| 12.7 ± 0.8 11.1 ± 0.3 8.1 ± 3.1
R2 0.98 0.99 0.68
E
X
P
E
R
I
M
E
N
T
O
8
Iluminación
60%
T ΔT [K] ΔV [mv]
Planta a Planta b Planta c
34 9.9±0.2 -32±25 -7±134 -58±28
30 7.6±0.2 -42±24 -14±137 -69±295
26 4.9±0.1 -61±24 -16±137 -48±455
22 1.7±0.1 -102±24 -54±135 -97±456
18 -2.1±0.2 -112±24 -67±137 -98±418
14 -7.0±0.3 -126±23 -122±133 -114±526
10 -11.6±0.2 -118±24 -128±135 -37±523
|𝜶| 6.0 ± 0.8 6.3 ± 0.6 3.6 ± 1.0
R2 0.93 0.95 0.76
Iluminación
0%
T ΔT [K] ΔV [mv]
Planta d Planta e Planta f
34 8.9±0.2 -47±12 18±59 16±12
30 6.8±0.3 -55±20 -1±124 9±13
26 4.3±0.2 -42±66 15±72 4±14
22 1.6±0.1 -93±74 -7±59 -10±14
18 -1.5±0.1 -94±73 -21±60 -5±14
14 -5.4±0.2 -104±75 -96±58 -48±15
10 -9.4±0.2 ----- -101±58 -51±16
|𝜶| 4.6 ± 1.2 7.1 ± 1.2 4.0 ± 0.9
R2 0.70 0.86 0.84

44
E
X
P
E
R
I
M
E
N
T
O
9
Iluminación
60%
T ΔT [K] ΔV [mv]
Planta a Planta b Planta c
35 9.1±0.1 ----- -134±121
31 4.6±0.5 -58±11 -77±105 -97±80
27 0.4±0.3 -61±28 -82±110 -98±83
23 -2.6±0.1 -121±8 -119±121 -133±27
19 -4.7±0.2 -110±7 -104±119 -125±29
15 -6.3±0.2 -84±7 -66±117 -152±25
11 -7.5±0.3 -91±5 -79±115 -170±24
|𝜶| 3.3 ± 2.2 -2.0 ± 1.6 5.8 ± 1.3
R2 0.30 0.25 0.80
Iluminación
0%
T ΔT [K] ΔV [mv]
Planta d Planta e Planta f
35 8.8±0.1 -149±9 -113±125 -----
31 4.7±0.4 -111±14 -35±57 -21±27
27 0.8±0.2 -102±19 0±58 -8±49
23 -2.3±0.1 -131±5 -34±56 -42±10
19 -4.7±0.2 -118±7 -53±91 -55±11
15 -6.6±0.2 -86±4 -131±168 -35±16
11 -8.3±0.3 -91±4 -191±169 -37±13
|𝜶| -2.5 ± 1.1 4.4 ± 4.4 2.0 ± 1.4
R2 0.5 0.16 0.38
E
X
P
E
R
I
M
E
N
T
O
10
Iluminación
60%
T ΔT [K] ΔV [mv]
Planta a Planta b Planta c
35 8.0±0.0 -86±26 -118±108 19±10
31 4.5±0.3 -76±26 -86±110 29±9
27 1.2±0.2 -78±27 ----- 36±9
23 -1.8±0.2 -88±29 -98±109 42±9
19 -4.5±0.1 -91±28 -98±106 42±9
15 -6.5±0.1 -101±29 -87±107 36±9
11 -8.4±0.2 -99±29 -80±109 -----
|𝛼| 1.2 ± 0.5 -1.4 ± 0.8 -1.3 ± 0.7
R2 0.61 0.47 0.71
Iluminación
0%
T ΔT [K] ΔV [mv]
Planta d Planta e Planta f
35 7.8±0.1 ----- -51±9
31 3.9±0.3 -44±14 -60±123 -42±9
27 0.1±0.1 -31±5 -33±122 -24±9
23 -3.1±0.1 -10±6 -61±140 -9±9
19 -5.8±0.1 -7±6 -72±38 4±5
15 -8.0±0.2 -18±6 -84±37 -3±6
11 -9.9±0.2 -18±5 -80±47 2±6
|𝜶| -3.2 ± 0.8 2.6 ± 1.2 -3.3 ± 0.4
R2 0.74 0.55 0.92

45
E
X
P
E
R
I
M
E
N
T
O
11
Iluminación
100%
T ΔT [K] ΔV [mv]
Planta a Planta b Planta c
34 8.7±0.2 2±17 61±186 99±22
30 7.1±0.2 -4±17 54±166 94±22
26 5.4±0.1 -32±18 44±168 90±21
22 3.3±0.0 -34±17 41±164 81v21
18 0.7±0.1 -63±20 35±162 82±17
14 -2.3±0.3 -100±19 25±165 60±18
10 -5.6±0.1 -117±18 18±168 45±16
|𝜶| 8.8 ± 0.5 2.9 ± 0.2 3.7 ± 0.4
R2 0.98 0.97 0.94
Iluminación
40%
T ΔT [K] ΔV [mv]
Planta d Planta e Planta f
34 11.5±0.2 114±7 45±239 35±29
30 9.4±0.2 91±8 20±242 20±31
26 6.7±0.2 52±9 -8±242 6±31
22 3.5±0.0 29±6 -17±240 4±31
18 -0.6±0.1 5±7 -41±246 -29±33
14 -4.9±0.3 -8±6 -55±238 -35±32
10 -9.5±0.2 -9±6 -56±242 -41±33
|𝜶| 7.5 ± 1.0 5.8 ± 0.7 4.2 ± 0.5
R2 0.93 0.94 0.94
E
X
P
E
R
I
M
E
N
T
O
12
Iluminación
100%
T ΔT [K] ΔV [mv]
Planta a Planta b Planta c
34 11.3±0.3 55±15 70±97 35±155
30 9.3±0.2 41±16 43±96 22±151
26 6.9±0.1 16±17 18±94 33±152
22 3.8±0.1 -8±15 -27±95 2±145
18 0.0±0.3 -30±16 -76±95 -38±144
14 -4.7±0.5 -60±14 -74±94 -63±140
10 -9.8±0.3 -121±12 -82±89 -79±126
|𝛼| 7.9 ± 0.4 7.6 ±1.4 6.0 ± 0.6
R2 0.98 0.86 0.94
Iluminación
40%
T ΔT [K] ΔV [mv]
Planta d Planta e Planta f
34 10.1±0.3 -45±12 76±76 14±13
30 8.2±0.3 -57±12 61±77 -3±14
26 6.2±0.2 -81±15 29±75 -24±14
22 3.8±0.1 -106±12 -13±74 -53±14
18 0.8±0.1 -114±13 -52±73 -69±14
14 -2.8±0.2 -111±12 -64±73 -61±13
10 -6.8±0.3 -157±12 -89±71 -81±14
|𝜶| 6.0 ± 0.8 10.3 ±1.1 5.4 ± 1.0
R2 0.91 0.94 0.84

46
E
X
P
E
R
I
M
E
N
T
O
13
Iluminación
60%
T ΔT [K] ΔV [mv]
Planta a Planta b Planta c
35 7.0±0.3 11±23 ----- -97±26
31 3.2±0.4 12±24 -34±112 -72±29
27 -0.2±0.2 7±22 -38±111 -59±29
23 -2.1±0.0 6±22 -48±110 -52±29
19 -4.8±0.1 2±22 -61±110 -54±29
15 -6.5±0.2 13±21 -64±106 -59±28
11 -7.7±0.2 21±21 -68±104 -49±28
|𝜶| -0.2 ± 0.5 3.4 ± 0.4 -2.7 ± 0.7
R2 0.04 0.95 0.77
Iluminación
0%
T ΔT [K] ΔV [mv]
Planta d Planta e Planta f
35 7.2±0.2 -100±4 20±54 -33±15
31 3.0±0.4 -65±7 15±54 -42±16
27 -0.2±0.2 -58±5 ----- -47±13
23 -2.0±0.1 -51±4 2±52 -63±10
19 -4.5±0.1 -52±4 5±50 -56±12
15 -6.0±0.2 -48±4 7±48 -30±12
11 -7.1±0.3 -47±4 10±46 -25±19
|𝜶| -3.3 ± 0.7 1.0 ± 0.4 -0.1 ± 1.2
R2 0.82 0.58 0.002
E
X
P
E
R
I
M
E
N
T
O
14
Iluminación
100%
T ΔT [K] ΔV [mv]
Planta a Planta b Planta c
34 9.2±0.2 24±28 -22±94 39±20
30 7.2v0.2 5±28 -46±96 38±14
26 5.0±0.1 -21±28 -71±93 7±15
22 2.4±0.1 -35±27 -92±93 -27±13
18 -0.9±0.2 -72±28 -125±93 -52±16
14 -5.0±0.3 -100±27 -73±13 -130±92
10 -9.7±0.3 -108±28 -131±93 -108±12
|𝛼| 7.3 ± 0.7 4.7 ±1.6 9.4 ±1.3
R2 0.95 0.63 0.9
Iluminación
40%
T ΔT [K] ΔV [mv]
Planta d Planta e Planta f
34 9.3±0.3 15±9 35±77 -9±15
30 7.2±0.3 15±7 32±75 -19±15
26 5.2±0.2 5±7 27±75 -51±17
22 2.6±0.1 -34±7 10±76 -68±14
18 -0.6±0.0 -57±7 4±75 -67±14
14 -4.6±0.3 -68±8 -3±75 -71±14
10 -9.4±0.3 -83±8 -14±76 -109±15
|𝜶| 6.0 ± 0.8 2.8 ± 0.3 4.7 ± 0.87
R2 0.92 0.95 0.87

47
APÉNDICE A4
RESULTADOS DE EXPERIMENTOS CON Epipremnum aureum
EN BAÑO TÉRMICO
Experimento 1
T ΔTprom[K] L1= 13cm L2= 23 cm L3=29cm L4=37 cm L5= 47 cm
15 -8.7±0.2 -132±16 -128±23 -101±24 -60±24 -47±25
18 -5.8±0.0 -158±17 -126±21 -132±25 -87±27 -62±27
21 -2.7±0.3 -176±17 -119±22 -77±32 -78±27 -59±28
24 -1.3±0.2 -170±17 -134v22 -74±23 -75±27 -64±28
27 1.4±0.1 -164±20 -123±22 -71±24 -63±27 -59±28
30 4.8±0.3 -175±19 -133±23 -84±27 -51±31 -64±28
33 8.5±0.1 -183±17 -125±21 -98±24 -57±30 -68±28
Promedio -165 -127 -91 -67 -61
Desviación 17 5 21 13 7
Experimento 2
15 -8.0±0.6 -216±16 -148±20 -87±21 -81±24 -74±27
18 -4.9±0.1 -217±15 -141±18 -100±22 -87±27 -78±24
21 -2.4±0.3 -223±15 -156±19 -124±23 -106±23 -94±24
24 0.8±0.2 -218±15 -153±18 -107±22 -100±24 -85±24
27 3.9±0.2 -214±16 -186±21 -112±22 -104±24 -89±24
30 7.1±0.1 -215±14 -213±19 -126±23 -106±23 -102±23
33 10.4±0.0 -225±16 -217±21 -121±23 -117±25 -104±27
Promedio -218 -173 -111 -100 -90
Desviación 4 32 14 12 12
Experimento 3
15 -9.0±0.2 -90±15 -107±21 -88±26 -20±26 -50±26
18 -6.3±0.2 -116±16 -104±27 -91±27 -37±26 -46±29
21 -2.4±0.1 -137±14 -101±23 -101±23 -48±24 -50±26
24 0.7±0.1 -134±14 -84±26 -117±26 -49±26 -51±26
27 3.8±0.1 -140±14 -109±31 -93±31 -47±26 -69±30
30 6.6±0.3 -160±15 -150±37 -130±37 -53±30 -45±30
33 9.1±0.1 -147±19 -151±32 -156±32 --- -97±57
Promedio -132 -115 -111 -42 -58
Desviación 23 25 25 12 19
Experimento 4
15 -8.3±0.3 -73±18 -83±23 -128±34 -51±25 -50±26
18 -5.5±0.1 -80±14 -67±22 -137±25 -57±25 -58±26
21 -2.4±0.4 --- --- -124±23 -106±22 -94±24
24 -0.3±0.1 -119±16 -88±23 -109±24 -67±25 -75±30
27 2.6±0.3 -156±18 -100±23 -136±26 -79±28 -88±30
30 4.7±0.1 -130±16 -94±22 -111±24 -77±27 -75±30
33 7.7±0.1 -139±17 -99±23 -131±26 -101±30 -79±30
Promedio -116 -89 -125 -77 -74
Desviación 33 12 11 21 15

48
Experimento 5
T ΔTprom[K] L1= 13cm L2= 23 cm L3=29cm L4=37 cm L5= 47 cm
15 -8.0±0.6 --- -148±20 -87±21 -81±24 -74±27
18 -6.4±0.2 -135±19 -139±23 -115±25 -96±28 -89±30
21 -3.1±0.1 -142±19 -143±23 -130±25 -98±27 -96±30
24 0.1±0.1 -163±21 -166±25 -138±27 -104±28 -100±28
27 3.5±0.1 -161±19 -123±24 -124±25 -82±32 -83±30
30 6.9±0.1 -162±19 -123±25 -134±25 -95±29 -82±29
33 9.6±0.0 -162±23 -122±24 -130±25 -88±33 -88±29
Promedio -154 -138 -122 -92 -87
Desviación 12 16 17 9 9
Experimento 6
15 -9.0±0.2 91±19 110±23 88±28 44±29 112±32
18 -5.5±0.1 91±16 144±25 131±28 72±29 125±31
21 -3.6±0.7 103±17 126±24 98±27 86±30 120±33
24 -0.5±0.1 118±18 118±26 113±27 86±32 116±33
27 3.3±0.2 148±20 141±28 133±30 75±36 113±35
30 6.3±0.2 162±27 140±27 125±28 73±30 111±35
33 8.8±0.1 113±18 101±24 117±26 68±30 107±31
Promedio 118 126 115 72 115
Desviación 27 17 17 14 6

49
APENDICE A5
RESULTADOS DE EXPERIMENTOS CON Ficus Benjamina
PARA LA METODOLOGIA DE CONEXIONES MULTIPLES
EXPERIMENTO 1 (t = 19 h, T=4°C) *
*El tiempo (t) se refiere al momento del experimento en el cual se agregó agua a la raíz de la
planta, y la temperatura (T) se refiere a la del agua agregada.
----- T1 ----- T2

50
EXPERIMENTO 2 (t = 2 h, T=40°C) *
*El tiempo (t) se refiere al momento del experimento en el cual se agregó agua a la raíz de la
planta, y la temperatura (T) se refiere a la del agua agregada.
----- T1 ----- T2

51
EXPERIMENTO 3 (t = 7h, T=21°C) *
*El tiempo (t) se refiere al momento del experimento en el cual se agregó agua a la raíz de la
planta, y la temperatura (T) se refiere a la del agua agregada.
----- T1 ----- T2

52
EXPERIMENTO 4 (t = 3 h, T=40°C) *
*El tiempo (t) se refiere al momento del experimento en el cual se agregó agua a la raíz de la
planta, y la temperatura (T) se refiere a la del agua agregada.
----- T1 ----- T2

53
EXPERIMENTO 5 (t = 4 h, T=4°C) *
*El tiempo (t) se refiere al momento del experimento en el cual se agregó agua a la raíz de la
planta, y la temperatura (T) se refiere a la del agua agregada.
----- T1 ----- T2
54
EXPERIMENTO 6 (t = 4 h, T= 21°C) *
*El tiempo (t) se refiere al momento del experimento en el cual se agregó agua a la raíz de la
planta, y la temperatura (T) se refiere a la del agua agregada.
----- T1 ----- T2

55
EXPERIMENTO 7 (t = 5 h, T=4°C) *
*El tiempo (t) se refiere al momento del experimento en el cual se agregó agua a la raíz de la
planta, y la temperatura (T) se refiere a la del agua agregada.
----- T1 ----- T2

56
EXPERIMENTO 8 (t = 5 h, T=40°C) *
*El tiempo (t) se refiere al momento del experimento en el cual se agregó agua a la raíz de la
planta, y la temperatura (T) se refiere a la del agua agregada.
----- T1 ----- T2





























