Embed Size (px)
Citation preview

Substituted Cyclodextrin as a Model for a Squid Enzyme That Hydrolyzes the Nerve GasSomanAuthor(s): Francis C. G. Hoskin, Diane M. Steeves and John E. WalkerSource: Biological Bulletin, Vol. 197, No. 2, Centennial Issue: October, 1899-1999 (Oct., 1999),pp. 284-285Published by: Marine Biological LaboratoryStable URL: http://www.jstor.org/stable/1542654 .
Accessed: 25/06/2014 03:40
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
Marine Biological Laboratory is collaborating with JSTOR to digitize, preserve and extend access toBiological Bulletin.
http://www.jstor.org
This content downloaded from 195.78.109.12 on Wed, 25 Jun 2014 03:40:08 AMAll use subject to JSTOR Terms and Conditions
REPORTS FROM THE MBL GENERAL SCIENTIFIC MEETINGS REPORTS FROM THE MBL GENERAL SCIENTIFIC MEETINGS
2. Schreiber, D. R., F. J. Millero, and A. S. Gordon. 1990. Mar. Chem. 28: 275-284.
3. Harwood, V. J. and A. S. Gordon. 1994. Appl. Environ. Microbiol. 60: 1794-1753.
4. Battistuzi, G., R. Petrucci, L. Silvagni, F. R. Urbani, and S. Caiola. 1981. Ann. Hum. Genet. 45: 223-229.
5. Duncan, R., M. A. Faggart, A. J. Roger, and N. W. Cornell. Mol. Biol. Evol. 16: 383-396.
6. Fleming, D. E. B., D. R. Chettle, J. G. Wetmur, R. J. Desnick, J-P.
2. Schreiber, D. R., F. J. Millero, and A. S. Gordon. 1990. Mar. Chem. 28: 275-284.
3. Harwood, V. J. and A. S. Gordon. 1994. Appl. Environ. Microbiol. 60: 1794-1753.
4. Battistuzi, G., R. Petrucci, L. Silvagni, F. R. Urbani, and S. Caiola. 1981. Ann. Hum. Genet. 45: 223-229.
5. Duncan, R., M. A. Faggart, A. J. Roger, and N. W. Cornell. Mol. Biol. Evol. 16: 383-396.
6. Fleming, D. E. B., D. R. Chettle, J. G. Wetmur, R. J. Desnick, J-P.
Robin, D. Boulay, N. S. Richard, C. L. Gordon, and C. E. Webber. 1998. Environ. Res. 77: 49-61.
7. Burden, V. M., M. B. Sandheinrich, and C. A. Caldwell. 1998. Environ. Pollut. 101: 285-289.
8. Shine, J. P., R. V. Ika, and T. E. Ford. 1995. Environ. Sci. Technol. 29: 1781-1788.
9. Chauhan, S., and M. R. O'Brian. 1995. J. Biol. Chem. 270: 19823- 19827.
Robin, D. Boulay, N. S. Richard, C. L. Gordon, and C. E. Webber. 1998. Environ. Res. 77: 49-61.
7. Burden, V. M., M. B. Sandheinrich, and C. A. Caldwell. 1998. Environ. Pollut. 101: 285-289.
8. Shine, J. P., R. V. Ika, and T. E. Ford. 1995. Environ. Sci. Technol. 29: 1781-1788.
9. Chauhan, S., and M. R. O'Brian. 1995. J. Biol. Chem. 270: 19823- 19827.
Reference: Biol. Bull. 197: 284-285. (October 1999)
Substituted Cyclodextrin as a Model for a Squid Enzyme that Hydrolyzes the Nerve Gas Soman Francis C. G. Hoskin (Marine Biological Laboratory, Woods Hole, Massachusetts 02543),
Diane M. Steeves1, and John E. Walker'
Reference: Biol. Bull. 197: 284-285. (October 1999)
Substituted Cyclodextrin as a Model for a Squid Enzyme that Hydrolyzes the Nerve Gas Soman Francis C. G. Hoskin (Marine Biological Laboratory, Woods Hole, Massachusetts 02543),
Diane M. Steeves1, and John E. Walker'
Certain phosphorus-fluorine (P-F) compounds are powerful in- hibitors of the nerve enzyme acetylcholinesterase (AChE), and are thus termed "nerve gases." One such compound is soman, 1,2,2- trimethylpropyl methylphosphonofluoridate. An enzyme that hy- drolyzes, and thus detoxifies soman has been purified from squid nerve and immobilized on agarose resin. However, this enzyme, termed organophosphorus acid anhydrolase (OPAA), hydrolyzes the relatively non-toxic pair of diastereoisomers, C(+)P(+), more
rapidly than the toxic pair C(?)P(-) (1). Cyclodextrins are donut- shaped heptahexose molecules that can also act as P-F hydrolyzing "synzymes" (2, 3); but their rates are much lower than those of the OPAAs. We now report the rapid hydrolysis and detoxication of soman by a substituted cyclodextrin: 2-0-(4-carboxy-3-iodosoben- zoyl)-,3-cyclodextrin (IBA-3CD) (4). Dimebu (3,3-dimethylbutyl methylphosphonofluoridate) (5), an isomer of soman with only one chiral center, P(?), is also hydrolyzed by IBA-f3CD. Information about the catalysis of these reactions by IBA-j3CD contributes to our understanding of the active site of the naturally occurring OPAA.
The hydrolysis of soman or dimebu was monitored with a fluoride-sensitive electrode (1); the loss of AChE inhibitory po- tency, by a modified Ellman reaction (1). A 5-ml solution was made 25 mM in Pipes buffer, 3 mM in soman or dimebu, and 0.03 mM in IBA-,3CD; pH 7, 22?-23?C. After approximately half
hydrolysis, as measured by the fluoride electrode, 0.1 ml of the reaction solution was removed, diluted at ice-water temperature, and tested for AChE inhibitory potency. The results of the AChE determinations, and the fluoride measurements at that point are
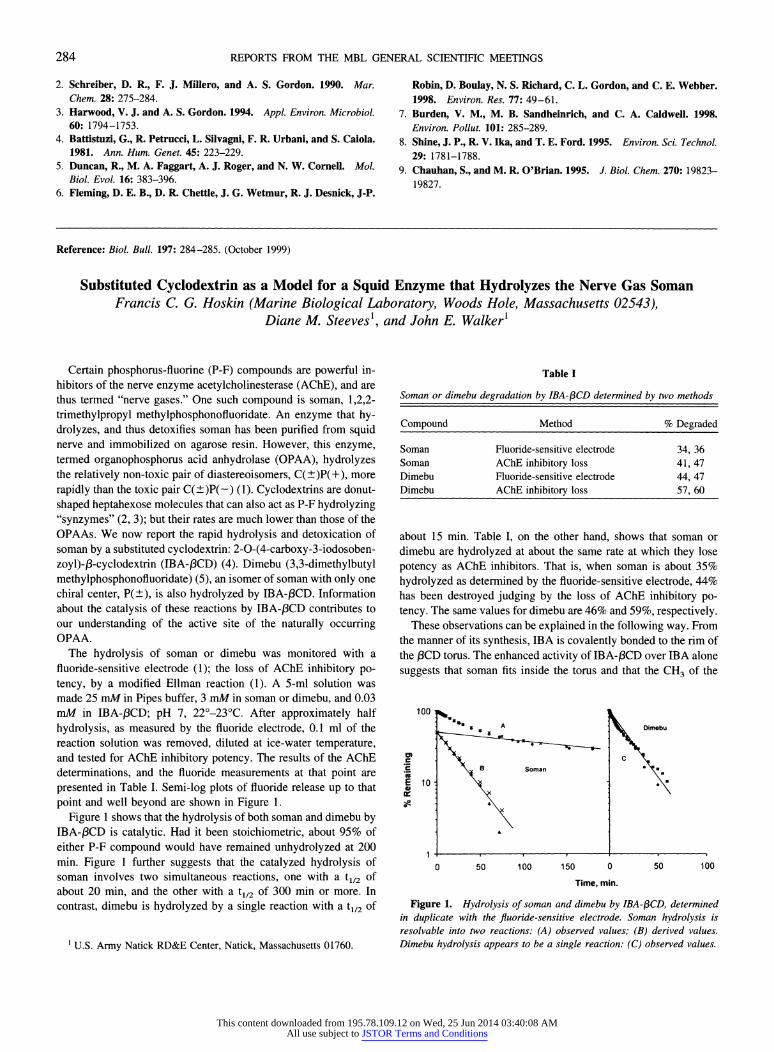
presented in Table I. Semi-log plots of fluoride release up to that point and well beyond are shown in Figure 1.
Figure 1 shows that the hydrolysis of both soman and dimebu by IBA-/3CD is catalytic. Had it been stoichiometric, about 95% of either P-F compound would have remained unhydrolyzed at 200 min. Figure 1 further suggests that the catalyzed hydrolysis of soman involves two simultaneous reactions, one with a tl/2 of about 20 min, and the other with a t1/2 of 300 min or more. In contrast, dimebu is hydrolyzed by a single reaction with a tl12 of
Certain phosphorus-fluorine (P-F) compounds are powerful in- hibitors of the nerve enzyme acetylcholinesterase (AChE), and are thus termed "nerve gases." One such compound is soman, 1,2,2- trimethylpropyl methylphosphonofluoridate. An enzyme that hy- drolyzes, and thus detoxifies soman has been purified from squid nerve and immobilized on agarose resin. However, this enzyme, termed organophosphorus acid anhydrolase (OPAA), hydrolyzes the relatively non-toxic pair of diastereoisomers, C(+)P(+), more
rapidly than the toxic pair C(?)P(-) (1). Cyclodextrins are donut- shaped heptahexose molecules that can also act as P-F hydrolyzing "synzymes" (2, 3); but their rates are much lower than those of the OPAAs. We now report the rapid hydrolysis and detoxication of soman by a substituted cyclodextrin: 2-0-(4-carboxy-3-iodosoben- zoyl)-,3-cyclodextrin (IBA-3CD) (4). Dimebu (3,3-dimethylbutyl methylphosphonofluoridate) (5), an isomer of soman with only one chiral center, P(?), is also hydrolyzed by IBA-f3CD. Information about the catalysis of these reactions by IBA-j3CD contributes to our understanding of the active site of the naturally occurring OPAA.
The hydrolysis of soman or dimebu was monitored with a fluoride-sensitive electrode (1); the loss of AChE inhibitory po- tency, by a modified Ellman reaction (1). A 5-ml solution was made 25 mM in Pipes buffer, 3 mM in soman or dimebu, and 0.03 mM in IBA-,3CD; pH 7, 22?-23?C. After approximately half
hydrolysis, as measured by the fluoride electrode, 0.1 ml of the reaction solution was removed, diluted at ice-water temperature, and tested for AChE inhibitory potency. The results of the AChE determinations, and the fluoride measurements at that point are
presented in Table I. Semi-log plots of fluoride release up to that point and well beyond are shown in Figure 1.
Figure 1 shows that the hydrolysis of both soman and dimebu by IBA-/3CD is catalytic. Had it been stoichiometric, about 95% of either P-F compound would have remained unhydrolyzed at 200 min. Figure 1 further suggests that the catalyzed hydrolysis of soman involves two simultaneous reactions, one with a tl/2 of about 20 min, and the other with a t1/2 of 300 min or more. In contrast, dimebu is hydrolyzed by a single reaction with a tl12 of
U.S. Army Natick RD&E Center, Natick, Massachusetts 01760. U.S. Army Natick RD&E Center, Natick, Massachusetts 01760.
Table I Table I
Soman or dimebu degradation by IBA-43CD determined by two methods
Compound Method % Degraded
Soman Fluoride-sensitive electrode 34, 36 Soman AChE inhibitory loss 41, 47 Dimebu Fluoride-sensitive electrode 44, 47 Dimebu AChE inhibitory loss 57, 60
Soman or dimebu degradation by IBA-43CD determined by two methods
Compound Method % Degraded
Soman Fluoride-sensitive electrode 34, 36 Soman AChE inhibitory loss 41, 47 Dimebu Fluoride-sensitive electrode 44, 47 Dimebu AChE inhibitory loss 57, 60
about 15 min. Table I, on the other hand, shows that soman or dimebu are hydrolyzed at about the same rate at which they lose potency as AChE inhibitors. That is, when soman is about 35% hydrolyzed as determined by the fluoride-sensitive electrode, 44% has been destroyed judging by the loss of AChE inhibitory po- tency. The same values for dimebu are 46% and 59%, respectively.
These observations can be explained in the following way. From the manner of its synthesis, IBA is covalently bonded to the rim of the 13CD torus. The enhanced activity of IBA-f3CD over IBA alone suggests that soman fits inside the torus and that the CH3 of the
about 15 min. Table I, on the other hand, shows that soman or dimebu are hydrolyzed at about the same rate at which they lose potency as AChE inhibitors. That is, when soman is about 35% hydrolyzed as determined by the fluoride-sensitive electrode, 44% has been destroyed judging by the loss of AChE inhibitory po- tency. The same values for dimebu are 46% and 59%, respectively.
These observations can be explained in the following way. From the manner of its synthesis, IBA is covalently bonded to the rim of the 13CD torus. The enhanced activity of IBA-f3CD over IBA alone suggests that soman fits inside the torus and that the CH3 of the
100 100
r C
E W o
r C
E W o
Soma
i B Soman
Soma
i B Soman
Dimebu
C a
Dimebu
C a
10 10
0 50 100 150 0 50 100
Time, min.
Figure 1. Hydrolysis of soman and dimebu by IBA-13CD, determined in duplicate with the fluoride-sensitive electrode. Soman hydrolysis is resolvable into two reactions: (A) observed values; (B) derived values. Dimebu hydrolysis appears to be a single reaction: (C) observed values.
0 50 100 150 0 50 100
Time, min.
Figure 1. Hydrolysis of soman and dimebu by IBA-13CD, determined in duplicate with the fluoride-sensitive electrode. Soman hydrolysis is resolvable into two reactions: (A) observed values; (B) derived values. Dimebu hydrolysis appears to be a single reaction: (C) observed values.
284 284
I I I I I I I I 1 1
This content downloaded from 195.78.109.12 on Wed, 25 Jun 2014 03:40:08 AMAll use subject to JSTOR Terms and Conditions

COMPARATIVE BIOCHEMISTRY COMPARATIVE BIOCHEMISTRY
methylphosphono part of soman is oriented away from the IBA. No matter whether the P=O or the P-F is to one side or the other of the IBA, about equal hydrolysis of the AChE inhibitory P(-)F isomers and the relatively non-inhibitory P(+)F pair will result. However, the chiral C-CH3 at the 1-propyl position of soman will, in one configuration, allow a close approach of the P-F to the IBA, but in the other configuration will increase the distance of the P-F from the IBA by 1-2A. The increased distance would cause a much slower hydrolysis rate for half of the racemic soman. The same reasoning applies to dimebu, but since there is no chiral
C-CH3 on the 3,3-dimethylbutyl portion, dimebu is hydrolyzed at a single fast rate.
Both AChE, which is inhibited by soman, and OPAA, the
enzyme that hydrolyzes soman, seem to be indifferent to the
configuration around the C chiral center (1). In contrast, IBA-,3CD, in its catalytic hydrolysis of soman, shows the reverse stereospec- ificity with respect to the two chiral centers. This conclusion is
supported by the use of dimebu, in which there is no C-chiral center. These findings and speculations have important implica-
methylphosphono part of soman is oriented away from the IBA. No matter whether the P=O or the P-F is to one side or the other of the IBA, about equal hydrolysis of the AChE inhibitory P(-)F isomers and the relatively non-inhibitory P(+)F pair will result. However, the chiral C-CH3 at the 1-propyl position of soman will, in one configuration, allow a close approach of the P-F to the IBA, but in the other configuration will increase the distance of the P-F from the IBA by 1-2A. The increased distance would cause a much slower hydrolysis rate for half of the racemic soman. The same reasoning applies to dimebu, but since there is no chiral
C-CH3 on the 3,3-dimethylbutyl portion, dimebu is hydrolyzed at a single fast rate.
Both AChE, which is inhibited by soman, and OPAA, the
enzyme that hydrolyzes soman, seem to be indifferent to the
configuration around the C chiral center (1). In contrast, IBA-,3CD, in its catalytic hydrolysis of soman, shows the reverse stereospec- ificity with respect to the two chiral centers. This conclusion is
supported by the use of dimebu, in which there is no C-chiral center. These findings and speculations have important implica-
tions for protection against, and disposal of, the acid anhydride type nerve gases, of which soman is an important example.
Literature Cited
1. Hoskin, F. C. G. 1990. Pp. 469-480 in Squid as Experimental Animals, D. L. Gilbert, W. J. Adelman, Jr., and J. M. Arnold, eds. Plenum Press, New York.
2. Klotz, I. M., G. P. Royer, and I. S. Scarpa. 1971. Proc. Natl. Acad. Sci. USA 68: 263-264.
3. Breslow, R., and S. D. Dong. 1998. Chem. Rev. 98: 1997-2012. 4. Seltzman, H. H. 1992. Pp. 24-29 in Final Report of Contract No.
DAMD 17-89-C-9012. Synthesis of Soman Scavengers. Requests for this document should be addressed to: Commander, U.S. Army Medical Research and Development Command, Attention SGRD-RM1-S, Fort Detrick, MD 21702 USA.
5. Chettur, G., J. J. DeFrank, B. J. Gallo, F. C. G. Hoskin, S. Mainer, F. M. Robbins, K. E. Steinmann, and J. E. Walker. 1988. Fundam. Appl. Toxicol. 11: 373-380.
tions for protection against, and disposal of, the acid anhydride type nerve gases, of which soman is an important example.
Literature Cited
1. Hoskin, F. C. G. 1990. Pp. 469-480 in Squid as Experimental Animals, D. L. Gilbert, W. J. Adelman, Jr., and J. M. Arnold, eds. Plenum Press, New York.
2. Klotz, I. M., G. P. Royer, and I. S. Scarpa. 1971. Proc. Natl. Acad. Sci. USA 68: 263-264.
3. Breslow, R., and S. D. Dong. 1998. Chem. Rev. 98: 1997-2012. 4. Seltzman, H. H. 1992. Pp. 24-29 in Final Report of Contract No.
DAMD 17-89-C-9012. Synthesis of Soman Scavengers. Requests for this document should be addressed to: Commander, U.S. Army Medical Research and Development Command, Attention SGRD-RM1-S, Fort Detrick, MD 21702 USA.
5. Chettur, G., J. J. DeFrank, B. J. Gallo, F. C. G. Hoskin, S. Mainer, F. M. Robbins, K. E. Steinmann, and J. E. Walker. 1988. Fundam. Appl. Toxicol. 11: 373-380.
Reference: Biol. Bull. 197: 285-286. (October 1999)
Effects of Green Tea Polyphenols on Lens Photooxidative Stress Seymour Zigman, Nancy S. Rafferty, Keen A. Rafferty, and Nathaniel Lewis (Eye Research Laboratory,
Department of Ophthalmology, Boston University School of Medicine, and Marine Biological Laboratory, Woods Hole, Massachusetts 02543)
Reference: Biol. Bull. 197: 285-286. (October 1999)
Effects of Green Tea Polyphenols on Lens Photooxidative Stress Seymour Zigman, Nancy S. Rafferty, Keen A. Rafferty, and Nathaniel Lewis (Eye Research Laboratory,
Department of Ophthalmology, Boston University School of Medicine, and Marine Biological Laboratory, Woods Hole, Massachusetts 02543)
Our purpose was to determine whether tea polyphenols such as
epigallocatechin gallate (EGCG) would protect certain functions of rabbit and dogfish lenses in vitro against the photooxidative stress of UVA irradiation. We have asked specifically whether catalase activity was protected.
New Zealand white rabbit eyes were obtained from PelFreeze
Biologicals within 20 hours of death. Dogfish (Mustelus canis) lenses were obtained fresh (under conditions approved by the MBL Institutional Animal Use Committee), and kept on ice for 5 h before use. The media used to maintain the lenses were Tyrode's solution (Sigma) for rabbit tissues and elasmobranch Ringer's medium (Marine Biological Laboratory) for dogfish tissues.
The HPLC-purified EGCG (one elution peak) was supplied in solid form by the Lipton Company (Englewood Cliffs, NJ), and the solid material was dissolved in Tyrode's or Ringer's solutions to make 50 jug/ml. Whole eyes were placed with their anterior surfaces up in small glass beakers on gauze soaked with medium.
Only the corneas were protruding into the air, and the beakers were filled with Tyrode's or Ringer's.
Whole eyes were exposed to the EGCG solutions as follows. Filled plastic bottle reservoirs were placed on a shelf above the beakers of eyes so that the solutions were delivered through plastic tubing connected to shortened micro-pipette tips at about 12 drops per min for 4 h. For comparison, solutions not containing EGCG were dropped onto control eyes at the same rate.
Cornea epithelia and lens capsule epithelia were dissected, washed in their respective media, and homogenized in glass ho-
Our purpose was to determine whether tea polyphenols such as
epigallocatechin gallate (EGCG) would protect certain functions of rabbit and dogfish lenses in vitro against the photooxidative stress of UVA irradiation. We have asked specifically whether catalase activity was protected.
New Zealand white rabbit eyes were obtained from PelFreeze
Biologicals within 20 hours of death. Dogfish (Mustelus canis) lenses were obtained fresh (under conditions approved by the MBL Institutional Animal Use Committee), and kept on ice for 5 h before use. The media used to maintain the lenses were Tyrode's solution (Sigma) for rabbit tissues and elasmobranch Ringer's medium (Marine Biological Laboratory) for dogfish tissues.
The HPLC-purified EGCG (one elution peak) was supplied in solid form by the Lipton Company (Englewood Cliffs, NJ), and the solid material was dissolved in Tyrode's or Ringer's solutions to make 50 jug/ml. Whole eyes were placed with their anterior surfaces up in small glass beakers on gauze soaked with medium.
Only the corneas were protruding into the air, and the beakers were filled with Tyrode's or Ringer's.
Whole eyes were exposed to the EGCG solutions as follows. Filled plastic bottle reservoirs were placed on a shelf above the beakers of eyes so that the solutions were delivered through plastic tubing connected to shortened micro-pipette tips at about 12 drops per min for 4 h. For comparison, solutions not containing EGCG were dropped onto control eyes at the same rate.
Cornea epithelia and lens capsule epithelia were dissected, washed in their respective media, and homogenized in glass ho-
mogenizers in 1.5 ml of medium. Homogenates were cleared of the insoluble residues by precipitation with an Eppendorf centrifuge at 12,000 rpm for 10 min. The presence of EGCG (50 /ig/ml) was detected in the cornea and lens, which were found to exhibit the same fluorescent and absorptive spectral qualities as the pure EGCG (excited at 490 nm, emission at 540 nm). We therefore attribute the fluorescences of these ocular tissues to the presence of EGCG.
Extracted lenses were pre-incubated in Tyrode's (rabbit) or elasmobranch Ringer's (dogfish) solutions containing 5 jLg/ml of EGCG. After the presoaking period, three lenses per group were
placed in beakers containing 15 ml of fresh Tyrode's or Ringer's solutions without EGCG. The beakers containing lenses with an- terior surface pointing upward were placed on a rack in our UVA
exposure chamber. The UVA radiation was provided by 15 Osram Sylvania 15 W
BL (maximum emission at 355 nm) lamps. Total exposure was 20 J/cm2. Lenses were kept on ice until examined.
When the experimental procedures were complete, the lenses were blotted, weighed analytically, and assayed for catalase activ-
ity (i.e., 02 production from H202) with an 02 meter and probe (Microelectrodes) connected to an (XY) LKB recorder. The slope of 02 generation per minute was used as a measure of catalase
activity. Controls for the EGCG-treated lenses and for UV-ex-
posed lenses were included in every experiment. The enzyme catalase is quite sensitive to inactivation due to
UVA exposure (1, 2). This was shown to occur in lenses of
mogenizers in 1.5 ml of medium. Homogenates were cleared of the insoluble residues by precipitation with an Eppendorf centrifuge at 12,000 rpm for 10 min. The presence of EGCG (50 /ig/ml) was detected in the cornea and lens, which were found to exhibit the same fluorescent and absorptive spectral qualities as the pure EGCG (excited at 490 nm, emission at 540 nm). We therefore attribute the fluorescences of these ocular tissues to the presence of EGCG.
Extracted lenses were pre-incubated in Tyrode's (rabbit) or elasmobranch Ringer's (dogfish) solutions containing 5 jLg/ml of EGCG. After the presoaking period, three lenses per group were
placed in beakers containing 15 ml of fresh Tyrode's or Ringer's solutions without EGCG. The beakers containing lenses with an- terior surface pointing upward were placed on a rack in our UVA
exposure chamber. The UVA radiation was provided by 15 Osram Sylvania 15 W
BL (maximum emission at 355 nm) lamps. Total exposure was 20 J/cm2. Lenses were kept on ice until examined.
When the experimental procedures were complete, the lenses were blotted, weighed analytically, and assayed for catalase activ-
ity (i.e., 02 production from H202) with an 02 meter and probe (Microelectrodes) connected to an (XY) LKB recorder. The slope of 02 generation per minute was used as a measure of catalase
activity. Controls for the EGCG-treated lenses and for UV-ex-
posed lenses were included in every experiment. The enzyme catalase is quite sensitive to inactivation due to
UVA exposure (1, 2). This was shown to occur in lenses of
285 285
This content downloaded from 195.78.109.12 on Wed, 25 Jun 2014 03:40:08 AMAll use subject to JSTOR Terms and Conditions





























