Embed Size (px)
Citation preview

Biosynthesis of Tyrosine O-Sulfate by Cell Proteoglycan from the Marine Sponge, MicrocionaproliferaAuthor(s): Octavian Popescu, Rey Interior, Gradimir Misevic, Max M. Burger and William J.KuhnsSource: Biological Bulletin, Vol. 197, No. 2, Centennial Issue: October, 1899-1999 (Oct., 1999),pp. 279-281Published by: Marine Biological LaboratoryStable URL: http://www.jstor.org/stable/1542651 .
Accessed: 25/06/2014 01:04
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
Marine Biological Laboratory is collaborating with JSTOR to digitize, preserve and extend access toBiological Bulletin.
http://www.jstor.org
This content downloaded from 188.72.126.47 on Wed, 25 Jun 2014 01:04:19 AMAll use subject to JSTOR Terms and Conditions

COMPARATIVE BIOCHEMISTRY COMPARATIVE BIOCHEMISTRY
both products being combined in equal amounts and incubated as above.
Dot blots were prepared for both experiments by spotting 2 ,u1 of one of the two premixtures on nitrocellulose. Non-specific reactive sites were blocked with 1% bovine serum albumin in
phosphate buffered saline. The spots were then treated with 2 ,ug of biotin-labeled hyaluronic acid binding protein (B-HABP) (Seikagaku No. 400763-1) for one hour, and then washed. This was followed by streptavidin-conjugated peroxidase and color
development with 3'3 diaminobenzidine and hydrogen peroxide. If
binding occurred, a brown coloration would appear at sites where substrates were applied to the nitrocellulose.
In experiment 1, two preparations of MAF gave a strong brown color at the sites of application of B-HABP; but MAF that was incubated with HAase showed no coloration. This result shows that the depolymerization of HA prevents the binding of MAF to B-HABP. In experiment 2, the site that contained MAF pre-treated with synthetic MAF-HABP remained almost colorless after the
application of biotin-labeled HABP. This finding indicated that HA in MAF had combined with the synthetic binding site and was unable to bind to the HABP probe. Taken together, the results of the two experiments demonstrate that the MAF macromolecule contains HA, presumably in polymeric form (Fig. 1).
These results extend our earlier findings that had demonstrated the
binding of biotin-labeled HA to the synthetic MAF-HABP (7). The existence of an HA binding amino acid sequence within the MAF core peptide suggests that its coupling to HA is a likely event. Our belief that HA-HABP coupling derives from a single site on the MAF core is supported by recent studies of Femandez-Busquets (manu- script in preparation). The occurrence of RHAMM, or other HA
both products being combined in equal amounts and incubated as above.
Dot blots were prepared for both experiments by spotting 2 ,u1 of one of the two premixtures on nitrocellulose. Non-specific reactive sites were blocked with 1% bovine serum albumin in
phosphate buffered saline. The spots were then treated with 2 ,ug of biotin-labeled hyaluronic acid binding protein (B-HABP) (Seikagaku No. 400763-1) for one hour, and then washed. This was followed by streptavidin-conjugated peroxidase and color
development with 3'3 diaminobenzidine and hydrogen peroxide. If
binding occurred, a brown coloration would appear at sites where substrates were applied to the nitrocellulose.
In experiment 1, two preparations of MAF gave a strong brown color at the sites of application of B-HABP; but MAF that was incubated with HAase showed no coloration. This result shows that the depolymerization of HA prevents the binding of MAF to B-HABP. In experiment 2, the site that contained MAF pre-treated with synthetic MAF-HABP remained almost colorless after the
application of biotin-labeled HABP. This finding indicated that HA in MAF had combined with the synthetic binding site and was unable to bind to the HABP probe. Taken together, the results of the two experiments demonstrate that the MAF macromolecule contains HA, presumably in polymeric form (Fig. 1).
These results extend our earlier findings that had demonstrated the
binding of biotin-labeled HA to the synthetic MAF-HABP (7). The existence of an HA binding amino acid sequence within the MAF core peptide suggests that its coupling to HA is a likely event. Our belief that HA-HABP coupling derives from a single site on the MAF core is supported by recent studies of Femandez-Busquets (manu- script in preparation). The occurrence of RHAMM, or other HA
receptors on Microciona cell populations may thus provide a means of MAF-cell binding that is calcium independent and distinct from the low molecular weight anionic carbohydrate epitopes previously de- scribed (10, 11). A signalling role for HA and its receptors has also been proposed (7); such linkages may enable Microciona cells to cross-communicate (Fig. 2).
Literature Cited
1. Henkart, P., S. Humphreys, and T. Humphreys. 1973. Biochem- istry 12: 3045-3050.
2. McLaurin, J., T. Franklin, W. Kuhns, and P. Fraser. 1999. Amy- loid (In press).
3. Jumblatt, J., V. Schlup, and M. Burger. 1980. Biochemistry 19: 1038-1042.
4. Misevic, G., and M. Burger. 1986. J. Biol. Chem. 261: 2853-2859. 5. Yang, B., L. Zhang, and E. Turley. 1993. J. Biol. Chem. 268:
8617-8623. 6. Fernandez-Busquets, X., R. Kammerer, and M. Burger. 1996.
J. Biol. Chem. 271: 23558-23565. 7. Kuhns, W., X. Fernandez-Busquets, M. Burger, M. Ho, and E.
Turley. 1998. Biol. Bull. 195: 216-218. 8. Misevic, G., J. Finne, and M. Burger. 1987. J. Biol. Chem. 262:
5870-5877. 9. Misevic, G., and M. Burger. 1993. J. Biol. Chem. 268: 4922-4929.
10. Spillmann, D., K. Hard, J. Thomas-Oates, J. Vliegenthart, G. Misevic, M. Burger, and J. Finne. 1993. J. Biol. Chem. 268: 13378-13387.
11. Spillmann, D., J. Thomas-Oates, A. van Kuik, J. Vliegenthart, G. Misevic, M. Burger, and J. Finne. 1995. J. Biol. Chem. 270: 5089-5097.
receptors on Microciona cell populations may thus provide a means of MAF-cell binding that is calcium independent and distinct from the low molecular weight anionic carbohydrate epitopes previously de- scribed (10, 11). A signalling role for HA and its receptors has also been proposed (7); such linkages may enable Microciona cells to cross-communicate (Fig. 2).
Literature Cited
1. Henkart, P., S. Humphreys, and T. Humphreys. 1973. Biochem- istry 12: 3045-3050.
2. McLaurin, J., T. Franklin, W. Kuhns, and P. Fraser. 1999. Amy- loid (In press).
3. Jumblatt, J., V. Schlup, and M. Burger. 1980. Biochemistry 19: 1038-1042.
4. Misevic, G., and M. Burger. 1986. J. Biol. Chem. 261: 2853-2859. 5. Yang, B., L. Zhang, and E. Turley. 1993. J. Biol. Chem. 268:
8617-8623. 6. Fernandez-Busquets, X., R. Kammerer, and M. Burger. 1996.
J. Biol. Chem. 271: 23558-23565. 7. Kuhns, W., X. Fernandez-Busquets, M. Burger, M. Ho, and E.
Turley. 1998. Biol. Bull. 195: 216-218. 8. Misevic, G., J. Finne, and M. Burger. 1987. J. Biol. Chem. 262:
5870-5877. 9. Misevic, G., and M. Burger. 1993. J. Biol. Chem. 268: 4922-4929.
10. Spillmann, D., K. Hard, J. Thomas-Oates, J. Vliegenthart, G. Misevic, M. Burger, and J. Finne. 1993. J. Biol. Chem. 268: 13378-13387.
11. Spillmann, D., J. Thomas-Oates, A. van Kuik, J. Vliegenthart, G. Misevic, M. Burger, and J. Finne. 1995. J. Biol. Chem. 270: 5089-5097.
Reference: Biol. Bull. 197: 279-281. (October 1999)
Biosynthesis of Tyrosine O-Sulfate by Cell Proteoglycan from the Marine Sponge, Microciona prolifera Octavian Popescu1, Rey Interior4, Gradimir Misevic2, Max M. Burger3, and William J. Kuhns4
(Marine Biological Laboratory, Woods Hole, Massachusetts, 02543)
Reference: Biol. Bull. 197: 279-281. (October 1999)
Biosynthesis of Tyrosine O-Sulfate by Cell Proteoglycan from the Marine Sponge, Microciona prolifera Octavian Popescu1, Rey Interior4, Gradimir Misevic2, Max M. Burger3, and William J. Kuhns4
(Marine Biological Laboratory, Woods Hole, Massachusetts, 02543)
Tyrosine sulfation is a post-translational modification of pro- tein that takes place in the trans-Golgi system (1). The inclusion of such a modified tyrosine in their native sequence confers
strong bioactivity to a variety of ligands and peptides involved in motility, secretion, cell binding, and the promotion of com-
plement action and blood coagulation (2-7). Tyrosine sulfa- tion is catalyzed by a specific tyrosylprotein sulfotransferase
(TPST); the enzyme has a particularly strong affinity for peptide substrates with a consensus sequence that features acidic amino acid residues adjacent, or at least close to, tyrosine (8). Se-
1 Institute for Biological Research, Cluj/Napoca, Romania (O.P.). 2 Biocenter, University of Basel, Switzerland. 3Friedrich Miescher Institute, Box 2543, CH4002, Basel, Switzerland. 4 Hospital for Sick Children, 555 University Avenue, Toronto, Ontario,
Canada, M5G 1X8.
Tyrosine sulfation is a post-translational modification of pro- tein that takes place in the trans-Golgi system (1). The inclusion of such a modified tyrosine in their native sequence confers
strong bioactivity to a variety of ligands and peptides involved in motility, secretion, cell binding, and the promotion of com-
plement action and blood coagulation (2-7). Tyrosine sulfa- tion is catalyzed by a specific tyrosylprotein sulfotransferase
(TPST); the enzyme has a particularly strong affinity for peptide substrates with a consensus sequence that features acidic amino acid residues adjacent, or at least close to, tyrosine (8). Se-
1 Institute for Biological Research, Cluj/Napoca, Romania (O.P.). 2 Biocenter, University of Basel, Switzerland. 3Friedrich Miescher Institute, Box 2543, CH4002, Basel, Switzerland. 4 Hospital for Sick Children, 555 University Avenue, Toronto, Ontario,
Canada, M5G 1X8.
quences with these criteria can be identified in translated por- tions of the cDNA encoding the protein core of the Microciona adhesive proteoglycan (MAF) (9). A related earlier finding of ours established that restriction of environmental sulfate in Microciona cell suspensions is accompanied by a diminution of secreted MAF and a loss of cell motility (10). These findings motivated a search for a functional correlate. We now describe, for the first time, the biosynthesis of tyrosine sulfate by cell
preparations from the marine sponge, Microciona prolifera. Cell suspensions (107/ml) were prepared in sulfate-free arti-
ficial seawater (ASW) from whole sponge fragments and ro- tated for 24 h at 16?C. The cells were then pulsed with 50 ,tCi of sulfur-35 (DuPontNEN NEX-041) and 50 ,uCi tyrosine-ring 3,5 3H (NEN) for 8 h. The pellets were washed 3 times: lysates were prepared in 20 mM Tris (pH 7.4) containing 1% Triton- X-100 and protease inhibitors (25 /ug/ml). Several aliquots (20
quences with these criteria can be identified in translated por- tions of the cDNA encoding the protein core of the Microciona adhesive proteoglycan (MAF) (9). A related earlier finding of ours established that restriction of environmental sulfate in Microciona cell suspensions is accompanied by a diminution of secreted MAF and a loss of cell motility (10). These findings motivated a search for a functional correlate. We now describe, for the first time, the biosynthesis of tyrosine sulfate by cell
preparations from the marine sponge, Microciona prolifera. Cell suspensions (107/ml) were prepared in sulfate-free arti-
ficial seawater (ASW) from whole sponge fragments and ro- tated for 24 h at 16?C. The cells were then pulsed with 50 ,tCi of sulfur-35 (DuPontNEN NEX-041) and 50 ,uCi tyrosine-ring 3,5 3H (NEN) for 8 h. The pellets were washed 3 times: lysates were prepared in 20 mM Tris (pH 7.4) containing 1% Triton- X-100 and protease inhibitors (25 /ug/ml). Several aliquots (20
279 279
This content downloaded from 188.72.126.47 on Wed, 25 Jun 2014 01:04:19 AMAll use subject to JSTOR Terms and Conditions
REPORTS FROM THE MBL GENERAL SCIENTIFIC MEETINGS
1 2 3 a -
kDa -220
- 97.4
-66
-46
*.;' - --30
Amino d Acid nmol
Asp 20.9 Glu 16.7 Ser 1.6 Gly 21.8 Arg 0.4 Thr 0.6 Ala 10.5 Pro 5.7 Tyr-S04 0.1 Tyr 2.2 Val 9.3 Met 2.7 lie 3.9 Leu 10.8 Phe 6.3 Lys 0.7 TOTAL 114.26
cpm 600
0 n-.
I I
I'
I
! _ _ I I I 1
I I I53 35 4 45 5
i I 10 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6 6.5 7 7.5 8 8.5 9 9.5 10
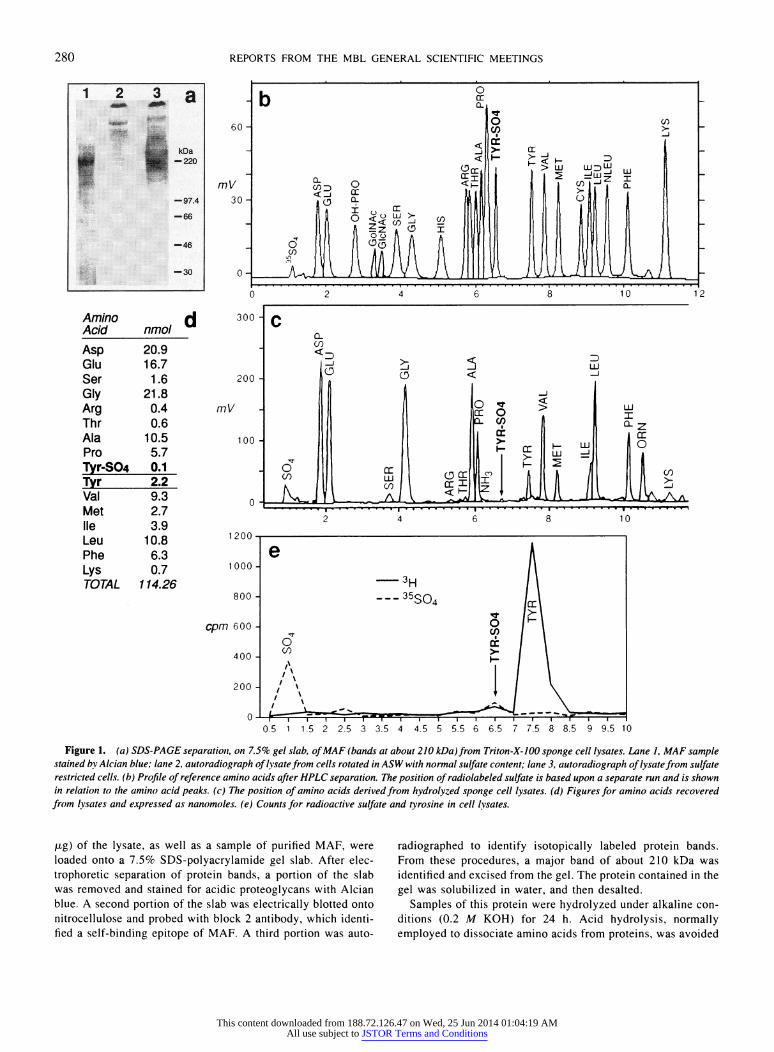
Figure 1. (a) SDS-PAGE separatiot, on 7.5% gel slab, ofMAF (bands at about 210 kDa)from Triton-X-100 sponge cell lysates. Lane I, MAF sample stained by Alcian blue; lane 2, autoradiograph of lysate from cells rotated in ASW with normal sulfate content; lane 3, autoradiograph of lysatefrom sulfate restricted cells. (b) Profile of reference amino acids after HPLC separation. The position of radiolabeled sulfate is based upon a separate run and is shown in relation to the amino acid peaks. (c) The position of amino acids derived from hydrolyzed sponge cell lysates. (d) Figures for amino acids recovered from lvsates and expressed as nanomoles. (e) Counts for radioactive sulfate and tyrosine in cell lysates.
,jg) of the lysate, as well as a sample of purified MAF, were loaded onto a 7.5% SDS-polyacrylamide gel slab. After elec- trophoretic separation of protein bands, a portion of the slab was removed and stained for acidic proteoglycans with Alcian blue. A second portion of the slab was electrically blotted onto nitrocellulose and probed with block 2 antibody, which identi- fied a self-binding epitope of MAF. A third portion was auto-
radiographed to identify isotopically labeled protein bands. From these procedures, a major band of about 210 kDa was identified and excised from the gel. The protein contained in the gel was solubilized in water, and then desalted.
Samples of this protein were hydrolyzed under alkaline con- ditions (0.2 M KOH) for 24 h. Acid hydrolysis, normally employed to dissociate amino acids from proteins, was avoided
60 -
mV 30 -
0-
0
300
200
mV
100
0
280
This content downloaded from 188.72.126.47 on Wed, 25 Jun 2014 01:04:19 AMAll use subject to JSTOR Terms and Conditions

COMPARATIVE BIOCHEMISTRY COMPARATIVE BIOCHEMISTRY
in this instance because of the acid lability of tyrosine sulfate. The recovered amino acids were derivatized and assayed with a Model ALC 204 liquid chromatograph equipped with a Pico- Tag analysis column (11). A mixture of known and calibrated amino acids was similarly derivatized for reference standards. After sample injection, the column was eluted with an acetoni- trile/water (AN/W) gradient consisting of 10% AN increasing to 51% AN in 12 min. A bolus of 10 /l containing 12 jLg of test sample was injected for each run. Recovery data for the stan- dards was established by converting areas for individual peaks to nanomoles. Test samples of the radio-labeled protein were injected separately into the same machine for radioactive counts (1 ml/assay), and 0.05 ml aliquots were removed at 0.5 min intervals for 10 min, and counted. The figures represent the average of duplicate counts.
The results are presented in Figure la-e. In the first panel (a) (Fig la, lane 3), a radioactive band at about 210 kDa is visualized upon autoradiography, and it corresponds in size to an Alcian blue band shown by MAF (lane 1). On a Western blot this protein gave a positive signal when reacted with block 2 antibody (not shown). Lysates from 26 mM sulfate ASW (i.e., not desulfated), showed very little incorporation of isotope (lane 2). The recoveries and analysis of amino acids are shown for the standards and test samples in Figure lb and c, respectively. Tyrosine sulfate is characterized by a peak at 6.6-6.7 min; this peak is very small in the test sample. The areas of these peaks were scanned and converted to nanomoles (Fig. Id). Recoveries of those amino acids present in large amounts (larger peaks) gave values within 10- 15% on duplicate runs, but the reproducibility of yields from very small peaks was poor. As an example, the nanomolar recovery ratio of tyrosine sulfate to tyrosine was 0.1/2.2 in one run as shown, and 0.03/2.5 in a second run. 35S and 3H radioactivity peaked at 1 min and 7.5 min, respectively (Fig. le). The sample collected at 6.5 min had counts for both isotopes, close to the position at which cold tyrosine sulfate was demonstrated. The results indicate that a small portion of the tyrosine had become sulfated.
The data presented here demonstrate that cell-derived MAF (defined by molecular size and by co-precipitation with specific anti-MAF block 2 antibody)-and perhaps other sponge pro- teins-possess tyrosine sulfate residues, but in small amounts
in this instance because of the acid lability of tyrosine sulfate. The recovered amino acids were derivatized and assayed with a Model ALC 204 liquid chromatograph equipped with a Pico- Tag analysis column (11). A mixture of known and calibrated amino acids was similarly derivatized for reference standards. After sample injection, the column was eluted with an acetoni- trile/water (AN/W) gradient consisting of 10% AN increasing to 51% AN in 12 min. A bolus of 10 /l containing 12 jLg of test sample was injected for each run. Recovery data for the stan- dards was established by converting areas for individual peaks to nanomoles. Test samples of the radio-labeled protein were injected separately into the same machine for radioactive counts (1 ml/assay), and 0.05 ml aliquots were removed at 0.5 min intervals for 10 min, and counted. The figures represent the average of duplicate counts.
The results are presented in Figure la-e. In the first panel (a) (Fig la, lane 3), a radioactive band at about 210 kDa is visualized upon autoradiography, and it corresponds in size to an Alcian blue band shown by MAF (lane 1). On a Western blot this protein gave a positive signal when reacted with block 2 antibody (not shown). Lysates from 26 mM sulfate ASW (i.e., not desulfated), showed very little incorporation of isotope (lane 2). The recoveries and analysis of amino acids are shown for the standards and test samples in Figure lb and c, respectively. Tyrosine sulfate is characterized by a peak at 6.6-6.7 min; this peak is very small in the test sample. The areas of these peaks were scanned and converted to nanomoles (Fig. Id). Recoveries of those amino acids present in large amounts (larger peaks) gave values within 10- 15% on duplicate runs, but the reproducibility of yields from very small peaks was poor. As an example, the nanomolar recovery ratio of tyrosine sulfate to tyrosine was 0.1/2.2 in one run as shown, and 0.03/2.5 in a second run. 35S and 3H radioactivity peaked at 1 min and 7.5 min, respectively (Fig. le). The sample collected at 6.5 min had counts for both isotopes, close to the position at which cold tyrosine sulfate was demonstrated. The results indicate that a small portion of the tyrosine had become sulfated.
The data presented here demonstrate that cell-derived MAF (defined by molecular size and by co-precipitation with specific anti-MAF block 2 antibody)-and perhaps other sponge pro- teins-possess tyrosine sulfate residues, but in small amounts
relative to non-sulfated tyrosine. This finding is consistent with others in established literature (12). The presence of tyrosine sulfate in MAF conforms to the sequence data derived from the translated MAF core protein cDNA, from which strongly acidic consensus sequences could be identified. Some of them also contained other favorable residues such as a low count of basic amino acids, a minimum of hydrophobic residues, and one or more turn-inducing amino acids such as glycine. The translated sequence that appears to be most favorable as a sulfation substrate is given by the 15 amino acid sequence VGFDPT- DYEVNEADG (9), and it compares well with peptide segments in other systems in which sulfate activation converts a non- binding ligand to one with cell-binding properties (4). The information from these experiments may be of value in future studies on the sulfation of tyrosine residues as it applies to such issues as environmental stress or cell-cell communication.
The authors express their gratitude to Dr. Glen Horton who kindly supplied the tyrosine-0-sulfate used as a reference standard in these assays.
Literature Cited
1. Huttner, W. 1988. Annu. Rev. Physiol. 50: 363-376. 2. Pouyani, T., and B. Seed. 1995. Cell 83: 333-343. 3. Dong, J., C. Li, and J. Lopez. 1994. Biochemistry 33: 13946-
13953. 4. Sako, D., K. Comess, K. Barone, R. Camphausen, D. Cumming,
and G. Shaw. 1995. Cell 83: 323-331. 5. Mutt, V. 1980. Pp. 169-221 in Gastrointestinal Hormones, G. B. J.
Glass, ed. Raven Press, New York. 6. Baeuerle, P. 1987. J. Cell Biol. 105: 2655-2664. 7. Lin, W., K. Larsen, G. Hortin, and J. Roth. 1992. J. Biol. Chem.
267: 2876-2878. 8. Hille, A., A. Braulke, K. von Figura, and W. Huttner. 1990. Eur.
J. Biochem. 188: 577-586. 9. Fernandez-Busquets, X., D. Gerosa, D. Hess, and M. Burger. 1998.
J. Biol. Chem. 273: 29545-29553. 10. Kuhns, W., O. Popescu, M. Burger, and G. Misevic. 1995. J. Cell.
Biochem. 57: 71-89. 11. Bidlingmeyer, B., S. Cohen, and T. Tarvin. 1984. J. Chromatogr.
336: 93-104. 12. Huttner, W., and P. Baeuerle. 1988. Mod. Cell Biol. 6: 97-140.
relative to non-sulfated tyrosine. This finding is consistent with others in established literature (12). The presence of tyrosine sulfate in MAF conforms to the sequence data derived from the translated MAF core protein cDNA, from which strongly acidic consensus sequences could be identified. Some of them also contained other favorable residues such as a low count of basic amino acids, a minimum of hydrophobic residues, and one or more turn-inducing amino acids such as glycine. The translated sequence that appears to be most favorable as a sulfation substrate is given by the 15 amino acid sequence VGFDPT- DYEVNEADG (9), and it compares well with peptide segments in other systems in which sulfate activation converts a non- binding ligand to one with cell-binding properties (4). The information from these experiments may be of value in future studies on the sulfation of tyrosine residues as it applies to such issues as environmental stress or cell-cell communication.
The authors express their gratitude to Dr. Glen Horton who kindly supplied the tyrosine-0-sulfate used as a reference standard in these assays.
Literature Cited
1. Huttner, W. 1988. Annu. Rev. Physiol. 50: 363-376. 2. Pouyani, T., and B. Seed. 1995. Cell 83: 333-343. 3. Dong, J., C. Li, and J. Lopez. 1994. Biochemistry 33: 13946-
13953. 4. Sako, D., K. Comess, K. Barone, R. Camphausen, D. Cumming,
and G. Shaw. 1995. Cell 83: 323-331. 5. Mutt, V. 1980. Pp. 169-221 in Gastrointestinal Hormones, G. B. J.
Glass, ed. Raven Press, New York. 6. Baeuerle, P. 1987. J. Cell Biol. 105: 2655-2664. 7. Lin, W., K. Larsen, G. Hortin, and J. Roth. 1992. J. Biol. Chem.
267: 2876-2878. 8. Hille, A., A. Braulke, K. von Figura, and W. Huttner. 1990. Eur.
J. Biochem. 188: 577-586. 9. Fernandez-Busquets, X., D. Gerosa, D. Hess, and M. Burger. 1998.
J. Biol. Chem. 273: 29545-29553. 10. Kuhns, W., O. Popescu, M. Burger, and G. Misevic. 1995. J. Cell.
Biochem. 57: 71-89. 11. Bidlingmeyer, B., S. Cohen, and T. Tarvin. 1984. J. Chromatogr.
336: 93-104. 12. Huttner, W., and P. Baeuerle. 1988. Mod. Cell Biol. 6: 97-140.
Reference: Biol. Bull. 197: 281-282. (October 1999)
Prophenoloxidase Is Not Activated by Microbial Signals in Limulus polyphemus Aimee Vasse, Alice Child, and Norman Wainwright (Marine Biological Laboratory,
Woods Hole, Massachusetts 02543)
Reference: Biol. Bull. 197: 281-282. (October 1999)
Prophenoloxidase Is Not Activated by Microbial Signals in Limulus polyphemus Aimee Vasse, Alice Child, and Norman Wainwright (Marine Biological Laboratory,
Woods Hole, Massachusetts 02543)
Prophenoloxidase (proPO) has been found in many inverte- brates as a zymogen component of the non-inducible defense cascade involved in local melanization. The microbial cell wall constituents, lipopolysaccharide (LPS), 13-1,3-glucan, and peptido- glycan are known to activate proPO, and thus, the melanin cas- cade, by activating prophenoloxidase-activating-enzyme (ppA). PpA proteolytically cleaves proPO into two active peptides, which
Prophenoloxidase (proPO) has been found in many inverte- brates as a zymogen component of the non-inducible defense cascade involved in local melanization. The microbial cell wall constituents, lipopolysaccharide (LPS), 13-1,3-glucan, and peptido- glycan are known to activate proPO, and thus, the melanin cas- cade, by activating prophenoloxidase-activating-enzyme (ppA). PpA proteolytically cleaves proPO into two active peptides, which
can then function as opsonins (1, 2), catalyze the hydroxylation of tyrosine to dopa, or both. Dopa is further oxidized to dopaquinone, which autocatalytically produces melanin as a modified clot at a wound site, or as an encapsulation of microbial invaders in the circulatory system.
Soderhall and coworkers (2, 3, 4, 5) deemed proPO absent from the horseshoe crab Limulus polyphemus, claiming that the primary
can then function as opsonins (1, 2), catalyze the hydroxylation of tyrosine to dopa, or both. Dopa is further oxidized to dopaquinone, which autocatalytically produces melanin as a modified clot at a wound site, or as an encapsulation of microbial invaders in the circulatory system.
Soderhall and coworkers (2, 3, 4, 5) deemed proPO absent from the horseshoe crab Limulus polyphemus, claiming that the primary
281 281
This content downloaded from 188.72.126.47 on Wed, 25 Jun 2014 01:04:19 AMAll use subject to JSTOR Terms and Conditions











![Azacytidine Enhances Efficacy of Tyrosine Kinase …...methylation in leukemia, lymphoma and multiple myeloma [73,74]. Moreover, over expression of bone marrow proteoglycan 2 (PRG2)](https://img.dokumen.tips/doc/110x75/5f3350b79b285c36e55d3a8a/azacytidine-enhances-efficacy-of-tyrosine-kinase-methylation-in-leukemia-lymphoma.jpg)
















