Embed Size (px)
Citation preview

Cell, Vol. 111, 77–89, October 4, 2002, Copyright !2002 by Cell Press
Asymmetries in H!/K!-ATPase and Cell MembranePotentials Comprise a Very Early Stepin Left-Right Patterning
anisms that act upstream of asymmetric gene expres-sion, however, remain poorly characterized. Thus, ourwork has focused on pursuing the determining mecha-nisms backward from the point of asymmetric gene ex-pression.
Michael Levin,1,2 Thorleif Thorlin,3,4
Kenneth R. Robinson,4 Taisaku Nogi,2
and Mark Mercola1,5,6
1Department of Cell BiologyHarvard Medical School240 Longwood Avenue Several mechanisms have been proposed to act up-
stream of sided gene expression to orient LR asymme-Boston, Massachusetts 021152 Cytokine Biology Department try. One model is based on the observation that the
vortical movement of individual monocilia that protrudeThe Forsyth Institute140 The Fenway from the ventral surface of cells in the mouse node
propel the leftward movement of an unknown determi-Boston, Massachusetts 021153 Sahlgrens University Hospital nant (Nonaka et al., 1998). This model is attractive be-
cause it provides a means of generating body axis asym-Institute for Clinical NeuroscienceBla Straket 7 metry from a molecular level chirality and is consistent
with the involvement of dynein and kinesin proteins as-SE 413 45 GothenburgSweden sociated with monocilia (Supp et al., 1999, 1997; Takeda
et al., 2000, 1999). However, the difficulty in reconciling4 Department of Biological SciencesPurdue University this model with experiments showing asymmetric ex-
pression of genes and patterning prior to node formationWest Lafayette, Indiana 47907in the chick (Levin and Mercola, 1999; Pagan-Westphaland Tabin, 1998; Psychoyos and Stern, 1996; Yuan andSchoenwolf, 1998) has prompted speculation that pro-Summaryfound differences exist between the asymmetry de-termining mechanisms used across vertebrate speciesA pharmacological screen identified the H! and K!
and/or that multiple, possibly reinforcing, mechanismsATPase transporter as obligatory for normal orienta-operate (Capdevila et al., 2000; Mercola and Levin,tion of the left-right body axis in Xenopus. Maternal2001). Xenopus embryos have been widely used toH!/K!-ATPase mRNA is symmetrically expressed inprobe early patterning steps and several lines of experi-the 1-cell Xenopus embryo but becomes localized dur-mentation suggest that components of the LR determi-ing the first two cell divisions, demonstrating thatnation mechanism act prior to the formation of Spe-asymmetry is generated within two hours postfertiliza-mann’s Organizer, which has signaling functions intion. Although H!/K!-ATPase subunit mRNAs are sym-common with the amniote node (for instance, see Hyattmetrically localized in chick embryos, an endogenousand Yost, 1998; Lohr et al., 1997). In previous studies,H!/K!-ATPase-dependent difference in membranewe have shown that intercellular communication via gapvoltage potential exists between the left and rightjunctions is an essential component of LR patterningsides of the primitive streak. In both species, pharma-upstream of early asymmetric gene expression in bothcologic or genetic perturbation of endogenous H!/K!-chick and Xenopus embryos (Levin and Mercola, 1998,ATPase randomized the sided pattern of asymmetri-1999). Importantly, genetic or pharmacologic disruptioncally expressed genes and induced organ heterotaxia.of gap junctional communication randomizes normallyThus, LR asymmetry determination depends on a veryasymmetric organ anatomy and gene expression. Al-early differential ion flux created by H!/K!-ATPase ac-though clearly required for LR asymmetry, the endoge-tivity.nous pattern of open junctional channels would seeminsufficient to orient the LR axis without a mechanismIntroductionto rectify the direction of propagation of determinantsthrough the gap junctions.The vertebrate body plan is outwardly bilaterally sym-
Here, we report the results of a pharmacologic screenmetric yet has consistent asymmetries in the positionindicating that the H!/K! ATPase transporter is essentialand anatomy of the visceral organs and brain. Muchfor normal LR asymmetry in developing Xenopus andprogress has been made recently in unraveling the mo-chicks. Although endogenous ion gradients and volt-lecular mechanisms involved in the generation of left-ages have been implicated in a number of embryonicright (LR) asymmetries during embryonic developmentprocesses (Altizer et al., 2001; Borgens and Shi, 1995;(Burdine and Schier, 2000; Capdevila et al., 2000; Mer-Hotary and Robinson, 1992, 1994; Jaffe, 1981; Shi andcola and Levin, 2001; Yost, 2001). The best understoodBorgens, 1994, 1995), our results identify a hydrogen/aspect of LR asymmetry determination is the geneticpotassium flux as a very early step in LR asymmetrycascade that transduces an initial asymmetry into cuesdetermination in chick and Xenopus embryos. We sug-interpretable by the developing visceral organs. Mech-gest that asymmetrically localized ion flux controlsasymmetric gene expression, possibly by directing the5 Correspondence: [email protected] low molecular weight determinants through6 Present address: The Burnham Institute, 10901 North Torrey Pines
Road, La Jolla, California 92037. gap junction channels.

Cell78
Results in the absence of other defects (Figure 1A and Supple-mental Table S1 available at http://www.cell.com/cgi/content/full/111/1/77/DC1). As these affect potassiumA Pharmacological Screen Implicates H!
flux, we then examined lansoprazole, a blocker of theand K! Ion Flux in LR AsymmetryH!/ K!-ATPase pump (Sanders et al., 1992) and foundIn order to determine whether endogenous ion flux isthat it induced heterotaxia in 51% of the embryos.important for LR patterning during development, we
took advantage of the ease of exposing large numbersof Xenopus embryos to compounds in medium, as well Inhibitors of H!/K!-ATPase Activity
Cause Heterotaxiaas the plethora of available pharmacological agents thattarget various ion channels and pumps. Drugs of in- The H!/K!-ATPase is a well-studied heterodimer con-
sisting of a larger " subunit responsible for transportcreasing specificity were used in successive reiterationsof the screen in order to narrow down the lead candidate and catalytic functions and a smaller glycosylated #
subunit with structural and membrane-targeting func-proteins. In preliminary studies, we determined the lev-els of each drug that did not cause general develop- tions (Gottardi and Caplan, 1993a, 1993b; Sachs et al.,
1995). In a number of epithelial cells, including gastricmental defects in Xenopus. Surprisingly, embryosreadily tolerated high concentrations of a number of ion parietal cells, H!/K!-ATPase in the apical plasma mem-
brane pumps H! out of the cell in exchange for K! atchannel and pump blockers. In each experiment, eggsfrom individual females were divided into groups of the expense of ATP hydrolysis. Three specific inhibitors
confirm that LR defects are caused by disruption ofabout 100 embryos each. The internal control group wascultured in 0.1$MMR containing the same amount of H!/K!-ATPase function (Figure 1B). Xenopus embryos
exposed between stages 1 and 16 to omeprazole (Mizu-vehicle as the experimental groups. Experimentalgroups were cultured in 0.1$MMR containing levels (see nashi et al., 1993), SCH28080 (Munson et al., 2000), and
lansoprazole (Sanders et al., 1992) exhibited 30%, 42%,Experimental Procedures) of each compound that weretitered to sufficiently low concentrations to permit abso- and 51% incidences of heterotaxia, respectively. In con-
trast, Na!/H! exchanger blockers such as amiloridelutely normal embryonic development and morphogene-sis of the heart and gut other than situs anomalies. The (Harris and Fliegel, 1999), ethylisopropylamiloride (EIPA;
Pizzonia et al., 1996), and cariporide (Mathur and Kar-normal dorsoanterior development (dorsoanterior index,DAI, of 5) and normally spaced eyes of the treated em- mazyn, 1997) had only a very modest or no effect (6%,
4%, and 1% incidences of heterotaxia, respectively).bryos indicate that the effects on LR asymmetry de-scribed below are not the consequence of compromised Likewise, treatment with the Na!/K!-ATPase inhibitor
ouabain (Croyle et al., 1997) or the mitochondrialdorsoanterior patterning. Embryos were allowed to de-velop in solution until stage 16, at which point they were F-ATPase inhibitor aurovertin (Weber and Senior, 1998)
induced only 8% and 1% incidences, respectively. Thus,washed extensively and cultured in 0.1$MMR untilstage 45. At that stage, anesthetized tadpoles were the pharmacological profile strongly implicates H!/K!-
ATPase activity in LR axis orientation.scored visually for the situs of the heart, gut coiling, andVisceral organs of treated Xenopus embryos devel-gallbladder. A defect in the laterality of any of those
oped asymmetrically, but with either normal or LR re-three organs was scored as an instance of heterotaxia.versed anatomy (Figures 1C–1G). Note that the headsFigure 1A summarizes a sample of the data resultingof treated embryos developed normally, with widelyfrom the pharmacological screen. The maximum levelspaced eyes. Head and eye anatomy are sensitive indi-of heterotaxia theoretically possible from the random-cators of early dorsoanterior patterning; thus, the condi-ization of three organs is 87.5% if each occurs indepen-tions of the assay did not alter dorsoanterior pattern.dently. Lower incidences are generally observed be-Although LR reversal of the viscera (situs inversus) wascause situs of an individual organ can be influenced byobserved frequently, individual reversals of the heart,that of adjacent organs [e.g., disturbances in gut rotationas well as reversal of gut and gallbladder together werecorrelate with an inversion of gallbladder situs (Casey,also common (Supplemental Table S1 available at above1998)] and the drug doses, which were chosen to pre-website) and similar for the three drugs. Thus, disruptionclude defects that might secondarily affect situs (suchof H!/K!-ATPase activity resulted in heterotaxia (discor-as effects on dorsoanterior patterning), are likely to bedant asymmetry of visceral organs), rather than situssuboptimal for perturbing laterality. Untreated controlinversus or isomerism (loss of asymmetry). Similarly,embryos exhibit approximately 1% incidence of hetero-chick embryos treated by injection of lansoprazole,taxia. Numerous compounds were found to have noomeprazole, or SCH28080 into the albumin prior to incu-specific effect on LR asymmetry, including spermine,bation and maintained in ovo until HH stage 15 hadp-chloromercuribenzenesulphonate (PCMB), amanta-frequent heart reversals but were otherwise anatomi-dine, indanyloxyacetic acid (IAA), 5-nitro-2-(3-phenyl-cally correct [Figures 1H and 1I; 0% of untreated em-propylamino)benzoic acid (NPPB, PAA), tetrodotoxin,bryos (n % 62) and 14% of injected embryos (n % 28)veratridine, quinidine, lanthanum, and amiodarone. Theexhibited reversals indicating a significant effect, p &pharmacological profile of the compounds that do not0.01 from chi-square analysis].affect organ laterality argues against the involvement
of a broad range of targets (assuming no permeabilitybarriers), including aquaporins, calcium, sodium, chlo- Inhibitors of H!/K!-ATPase Affect an Early
Developmental Process and Disrupt Normalride, and ligand-gated NMDA channels.In contrast, BaCl2 and the KvLQT-1 channel blocker Asymmetric Gene Expression
The current understanding of LR determination is thatchromanol 293B (Jiang and MacKinnon, 2000) caused32% and 45% incidences of heterotaxia, respectively, an early biasing mechanism orients the LR axis and
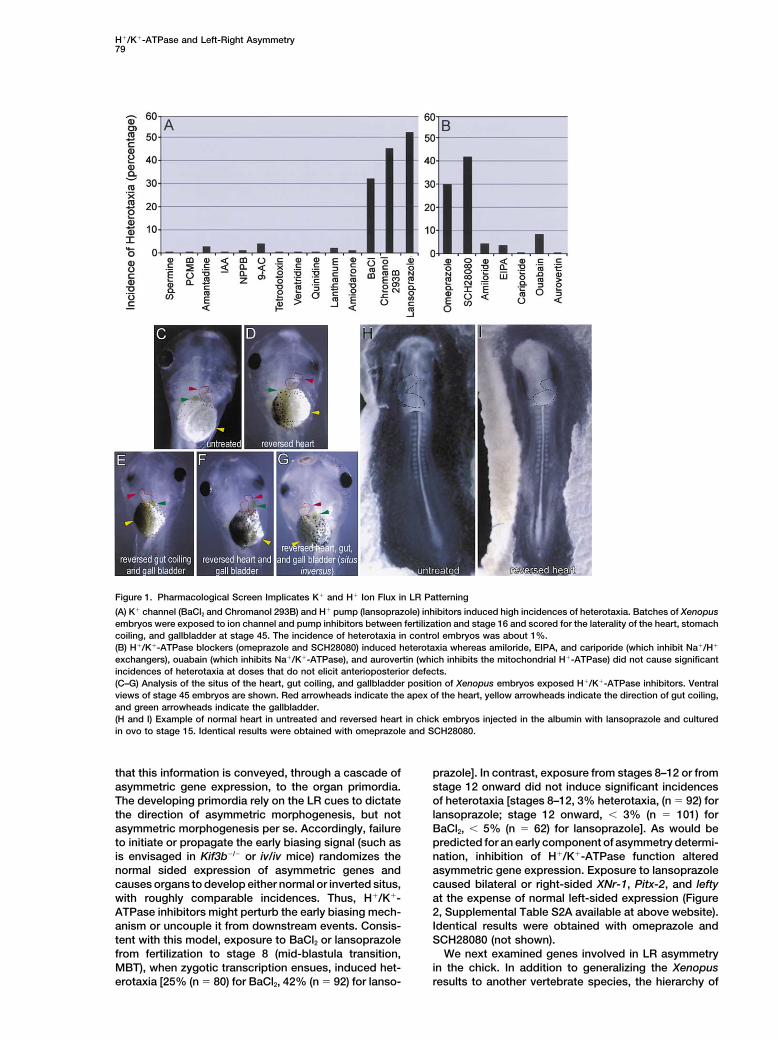
H!/K!-ATPase and Left-Right Asymmetry79
Figure 1. Pharmacological Screen Implicates K! and H! Ion Flux in LR Patterning
(A) K! channel (BaCl2 and Chromanol 293B) and H! pump (lansoprazole) inhibitors induced high incidences of heterotaxia. Batches of Xenopusembryos were exposed to ion channel and pump inhibitors between fertilization and stage 16 and scored for the laterality of the heart, stomachcoiling, and gallbladder at stage 45. The incidence of heterotaxia in control embryos was about 1%.(B) H!/K!-ATPase blockers (omeprazole and SCH28080) induced heterotaxia whereas amiloride, EIPA, and cariporide (which inhibit Na!/H!
exchangers), ouabain (which inhibits Na!/K!-ATPase), and aurovertin (which inhibits the mitochondrial H!-ATPase) did not cause significantincidences of heterotaxia at doses that do not elicit anterioposterior defects.(C–G) Analysis of the situs of the heart, gut coiling, and gallbladder position of Xenopus embryos exposed H!/K!-ATPase inhibitors. Ventralviews of stage 45 embryos are shown. Red arrowheads indicate the apex of the heart, yellow arrowheads indicate the direction of gut coiling,and green arrowheads indicate the gallbladder.(H and I) Example of normal heart in untreated and reversed heart in chick embryos injected in the albumin with lansoprazole and culturedin ovo to stage 15. Identical results were obtained with omeprazole and SCH28080.
that this information is conveyed, through a cascade of prazole]. In contrast, exposure from stages 8–12 or fromstage 12 onward did not induce significant incidencesasymmetric gene expression, to the organ primordia.
The developing primordia rely on the LR cues to dictate of heterotaxia [stages 8–12, 3% heterotaxia, (n % 92) forlansoprazole; stage 12 onward, & 3% (n % 101) forthe direction of asymmetric morphogenesis, but not
asymmetric morphogenesis per se. Accordingly, failure BaCl2, & 5% (n % 62) for lansoprazole]. As would bepredicted for an early component of asymmetry determi-to initiate or propagate the early biasing signal (such as
is envisaged in Kif3b'/' or iv/iv mice) randomizes the nation, inhibition of H!/K!-ATPase function alteredasymmetric gene expression. Exposure to lansoprazolenormal sided expression of asymmetric genes and
causes organs to develop either normal or inverted situs, caused bilateral or right-sided XNr-1, Pitx-2, and leftyat the expense of normal left-sided expression (Figurewith roughly comparable incidences. Thus, H!/K!-
ATPase inhibitors might perturb the early biasing mech- 2, Supplemental Table S2A available at above website).Identical results were obtained with omeprazole andanism or uncouple it from downstream events. Consis-
tent with this model, exposure to BaCl2 or lansoprazole SCH28080 (not shown).We next examined genes involved in LR asymmetryfrom fertilization to stage 8 (mid-blastula transition,
MBT), when zygotic transcription ensues, induced het- in the chick. In addition to generalizing the Xenopusresults to another vertebrate species, the hierarchy oferotaxia [25% (n % 80) for BaCl2, 42% (n % 92) for lanso-
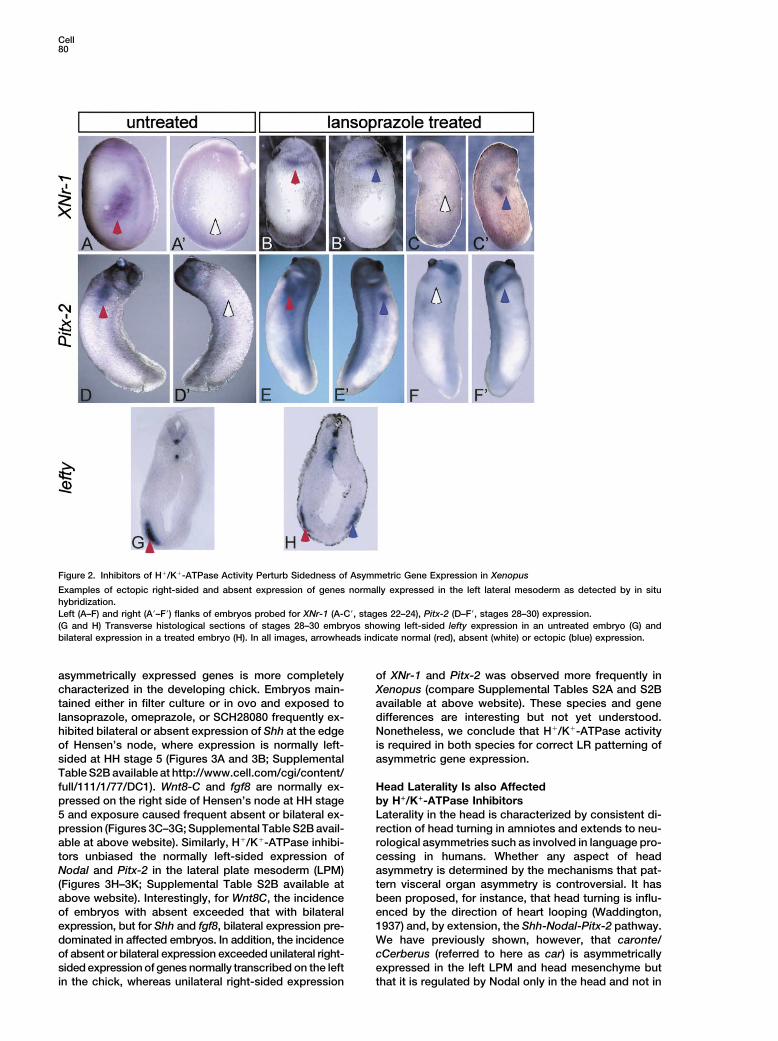
Cell80
Figure 2. Inhibitors of H!/K!-ATPase Activity Perturb Sidedness of Asymmetric Gene Expression in Xenopus
Examples of ectopic right-sided and absent expression of genes normally expressed in the left lateral mesoderm as detected by in situhybridization.Left (A–F) and right (A(–F() flanks of embryos probed for XNr-1 (A-C(, stages 22–24), Pitx-2 (D–F(, stages 28–30) expression.(G and H) Transverse histological sections of stages 28–30 embryos showing left-sided lefty expression in an untreated embryo (G) andbilateral expression in a treated embryo (H). In all images, arrowheads indicate normal (red), absent (white) or ectopic (blue) expression.
asymmetrically expressed genes is more completely of XNr-1 and Pitx-2 was observed more frequently inXenopus (compare Supplemental Tables S2A and S2Bcharacterized in the developing chick. Embryos main-
tained either in filter culture or in ovo and exposed to available at above website). These species and genedifferences are interesting but not yet understood.lansoprazole, omeprazole, or SCH28080 frequently ex-
hibited bilateral or absent expression of Shh at the edge Nonetheless, we conclude that H!/K!-ATPase activityis required in both species for correct LR patterning ofof Hensen’s node, where expression is normally left-
sided at HH stage 5 (Figures 3A and 3B; Supplemental asymmetric gene expression.Table S2B available at http://www.cell.com/cgi/content/full/111/1/77/DC1). Wnt8-C and fgf8 are normally ex- Head Laterality Is also Affected
by H!/K!-ATPase Inhibitorspressed on the right side of Hensen’s node at HH stage5 and exposure caused frequent absent or bilateral ex- Laterality in the head is characterized by consistent di-
rection of head turning in amniotes and extends to neu-pression (Figures 3C–3G; Supplemental Table S2B avail-able at above website). Similarly, H!/K!-ATPase inhibi- rological asymmetries such as involved in language pro-
cessing in humans. Whether any aspect of headtors unbiased the normally left-sided expression ofNodal and Pitx-2 in the lateral plate mesoderm (LPM) asymmetry is determined by the mechanisms that pat-
tern visceral organ asymmetry is controversial. It has(Figures 3H–3K; Supplemental Table S2B available atabove website). Interestingly, for Wnt8C, the incidence been proposed, for instance, that head turning is influ-
enced by the direction of heart looping (Waddington,of embryos with absent exceeded that with bilateralexpression, but for Shh and fgf8, bilateral expression pre- 1937) and, by extension, the Shh-Nodal-Pitx-2 pathway.
We have previously shown, however, that caronte/dominated in affected embryos. In addition, the incidenceof absent or bilateral expression exceeded unilateral right- cCerberus (referred to here as car) is asymmetrically
expressed in the left LPM and head mesenchyme butsided expression of genes normally transcribed on the leftin the chick, whereas unilateral right-sided expression that it is regulated by Nodal only in the head and not in
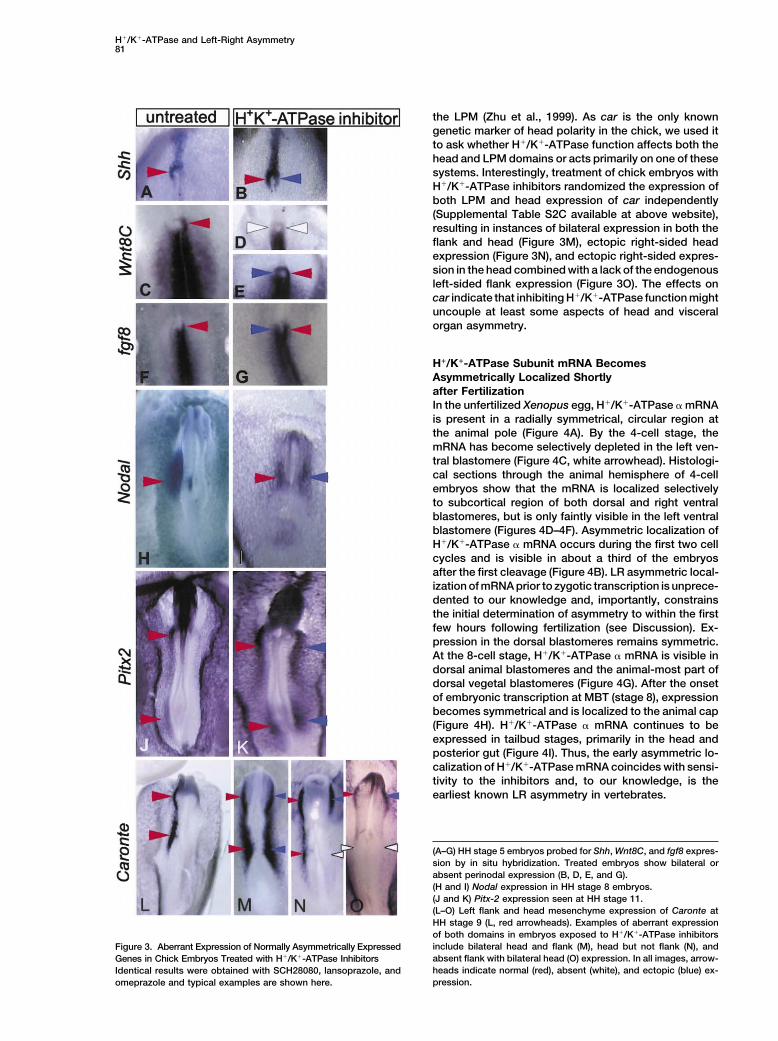
H!/K!-ATPase and Left-Right Asymmetry81
the LPM (Zhu et al., 1999). As car is the only knowngenetic marker of head polarity in the chick, we used itto ask whether H!/K!-ATPase function affects both thehead and LPM domains or acts primarily on one of thesesystems. Interestingly, treatment of chick embryos withH!/K!-ATPase inhibitors randomized the expression ofboth LPM and head expression of car independently(Supplemental Table S2C available at above website),resulting in instances of bilateral expression in both theflank and head (Figure 3M), ectopic right-sided headexpression (Figure 3N), and ectopic right-sided expres-sion in the head combined with a lack of the endogenousleft-sided flank expression (Figure 3O). The effects oncar indicate that inhibiting H!/K!-ATPase function mightuncouple at least some aspects of head and visceralorgan asymmetry.
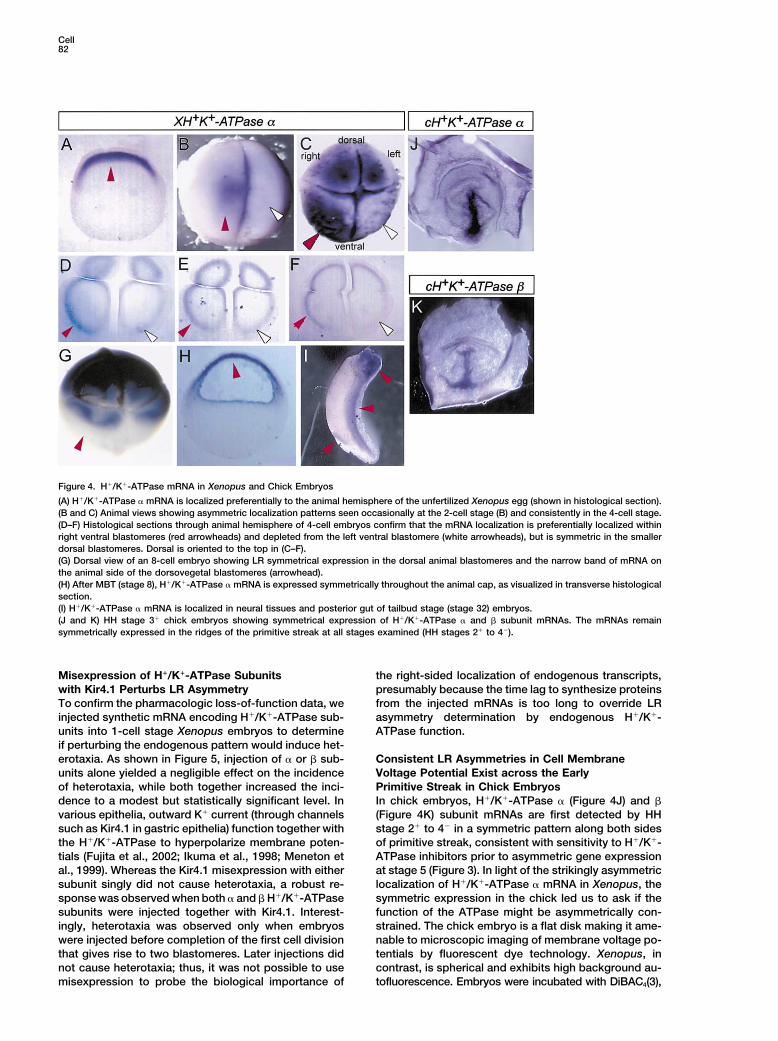
H!/K!-ATPase Subunit mRNA BecomesAsymmetrically Localized Shortlyafter FertilizationIn the unfertilized Xenopus egg, H!/K!-ATPase " mRNAis present in a radially symmetrical, circular region atthe animal pole (Figure 4A). By the 4-cell stage, themRNA has become selectively depleted in the left ven-tral blastomere (Figure 4C, white arrowhead). Histologi-cal sections through the animal hemisphere of 4-cellembryos show that the mRNA is localized selectivelyto subcortical region of both dorsal and right ventralblastomeres, but is only faintly visible in the left ventralblastomere (Figures 4D–4F). Asymmetric localization ofH!/K!-ATPase " mRNA occurs during the first two cellcycles and is visible in about a third of the embryosafter the first cleavage (Figure 4B). LR asymmetric local-ization of mRNA prior to zygotic transcription is unprece-dented to our knowledge and, importantly, constrainsthe initial determination of asymmetry to within the firstfew hours following fertilization (see Discussion). Ex-pression in the dorsal blastomeres remains symmetric.At the 8-cell stage, H!/K!-ATPase " mRNA is visible indorsal animal blastomeres and the animal-most part ofdorsal vegetal blastomeres (Figure 4G). After the onsetof embryonic transcription at MBT (stage 8), expressionbecomes symmetrical and is localized to the animal cap(Figure 4H). H!/K!-ATPase " mRNA continues to beexpressed in tailbud stages, primarily in the head andposterior gut (Figure 4I). Thus, the early asymmetric lo-calization of H!/K!-ATPase mRNA coincides with sensi-tivity to the inhibitors and, to our knowledge, is theearliest known LR asymmetry in vertebrates.
(A–G) HH stage 5 embryos probed for Shh, Wnt8C, and fgf8 expres-sion by in situ hybridization. Treated embryos show bilateral orabsent perinodal expression (B, D, E, and G).(H and I) Nodal expression in HH stage 8 embryos.(J and K) Pitx-2 expression seen at HH stage 11.(L–O) Left flank and head mesenchyme expression of Caronte atHH stage 9 (L, red arrowheads). Examples of aberrant expressionof both domains in embryos exposed to H!/K!-ATPase inhibitorsinclude bilateral head and flank (M), head but not flank (N), andFigure 3. Aberrant Expression of Normally Asymmetrically Expressedabsent flank with bilateral head (O) expression. In all images, arrow-Genes in Chick Embryos Treated with H!/K!-ATPase Inhibitorsheads indicate normal (red), absent (white), and ectopic (blue) ex-Identical results were obtained with SCH28080, lansoprazole, andpression.omeprazole and typical examples are shown here.
Cell82
Figure 4. H!/K!-ATPase mRNA in Xenopus and Chick Embryos
(A) H!/K!-ATPase " mRNA is localized preferentially to the animal hemisphere of the unfertilized Xenopus egg (shown in histological section).(B and C) Animal views showing asymmetric localization patterns seen occasionally at the 2-cell stage (B) and consistently in the 4-cell stage.(D–F) Histological sections through animal hemisphere of 4-cell embryos confirm that the mRNA localization is preferentially localized withinright ventral blastomeres (red arrowheads) and depleted from the left ventral blastomere (white arrowheads), but is symmetric in the smallerdorsal blastomeres. Dorsal is oriented to the top in (C–F).(G) Dorsal view of an 8-cell embryo showing LR symmetrical expression in the dorsal animal blastomeres and the narrow band of mRNA onthe animal side of the dorsovegetal blastomeres (arrowhead).(H) After MBT (stage 8), H!/K!-ATPase " mRNA is expressed symmetrically throughout the animal cap, as visualized in transverse histologicalsection.(I) H!/K!-ATPase " mRNA is localized in neural tissues and posterior gut of tailbud stage (stage 32) embryos.(J and K) HH stage 3! chick embryos showing symmetrical expression of H!/K!-ATPase " and # subunit mRNAs. The mRNAs remainsymmetrically expressed in the ridges of the primitive streak at all stages examined (HH stages 2! to 4').
Misexpression of H!/K!-ATPase Subunits the right-sided localization of endogenous transcripts,presumably because the time lag to synthesize proteinswith Kir4.1 Perturbs LR Asymmetry
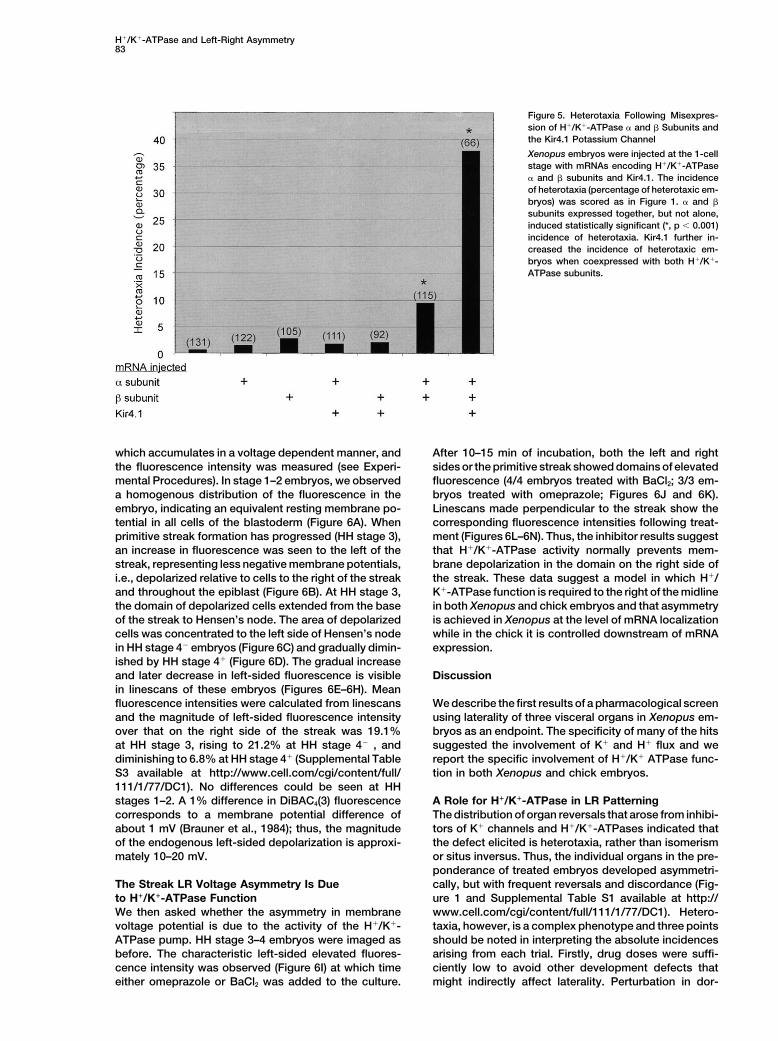
To confirm the pharmacologic loss-of-function data, we from the injected mRNAs is too long to override LRasymmetry determination by endogenous H!/K!-injected synthetic mRNA encoding H!/K!-ATPase sub-
units into 1-cell stage Xenopus embryos to determine ATPase function.if perturbing the endogenous pattern would induce het-erotaxia. As shown in Figure 5, injection of " or # sub- Consistent LR Asymmetries in Cell Membrane
Voltage Potential Exist across the Earlyunits alone yielded a negligible effect on the incidenceof heterotaxia, while both together increased the inci- Primitive Streak in Chick Embryos
In chick embryos, H!/K!-ATPase " (Figure 4J) and #dence to a modest but statistically significant level. Invarious epithelia, outward K! current (through channels (Figure 4K) subunit mRNAs are first detected by HH
stage 2! to 4' in a symmetric pattern along both sidessuch as Kir4.1 in gastric epithelia) function together withthe H!/K!-ATPase to hyperpolarize membrane poten- of primitive streak, consistent with sensitivity to H!/K!-
ATPase inhibitors prior to asymmetric gene expressiontials (Fujita et al., 2002; Ikuma et al., 1998; Meneton etal., 1999). Whereas the Kir4.1 misexpression with either at stage 5 (Figure 3). In light of the strikingly asymmetric
localization of H!/K!-ATPase " mRNA in Xenopus, thesubunit singly did not cause heterotaxia, a robust re-sponse was observed when both " and # H!/K!-ATPase symmetric expression in the chick led us to ask if the
function of the ATPase might be asymmetrically con-subunits were injected together with Kir4.1. Interest-ingly, heterotaxia was observed only when embryos strained. The chick embryo is a flat disk making it ame-
nable to microscopic imaging of membrane voltage po-were injected before completion of the first cell divisionthat gives rise to two blastomeres. Later injections did tentials by fluorescent dye technology. Xenopus, in
contrast, is spherical and exhibits high background au-not cause heterotaxia; thus, it was not possible to usemisexpression to probe the biological importance of tofluorescence. Embryos were incubated with DiBAC4(3),
H!/K!-ATPase and Left-Right Asymmetry83
Figure 5. Heterotaxia Following Misexpres-sion of H!/K!-ATPase " and # Subunits andthe Kir4.1 Potassium Channel
Xenopus embryos were injected at the 1-cellstage with mRNAs encoding H!/K!-ATPase" and # subunits and Kir4.1. The incidenceof heterotaxia (percentage of heterotaxic em-bryos) was scored as in Figure 1. " and #subunits expressed together, but not alone,induced statistically significant (*, p & 0.001)incidence of heterotaxia. Kir4.1 further in-creased the incidence of heterotaxic em-bryos when coexpressed with both H!/K!-ATPase subunits.
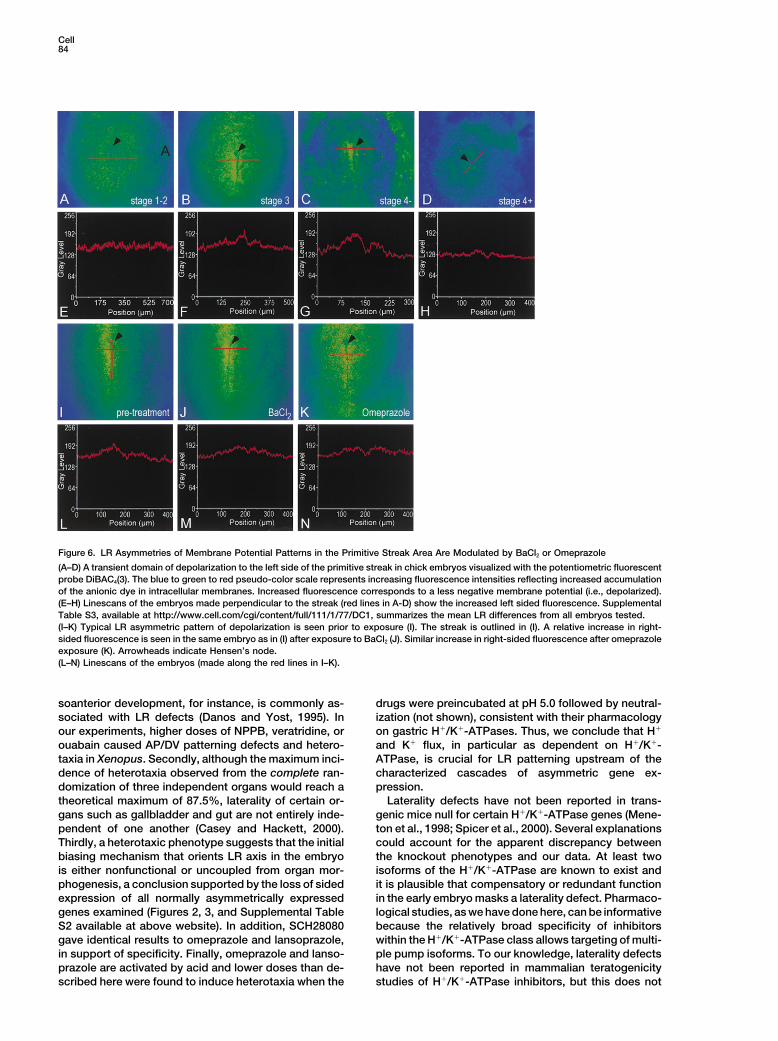
which accumulates in a voltage dependent manner, and After 10–15 min of incubation, both the left and rightsides or the primitive streak showed domains of elevatedthe fluorescence intensity was measured (see Experi-
mental Procedures). In stage 1–2 embryos, we observed fluorescence (4/4 embryos treated with BaCl2; 3/3 em-bryos treated with omeprazole; Figures 6J and 6K).a homogenous distribution of the fluorescence in the
embryo, indicating an equivalent resting membrane po- Linescans made perpendicular to the streak show thecorresponding fluorescence intensities following treat-tential in all cells of the blastoderm (Figure 6A). When
primitive streak formation has progressed (HH stage 3), ment (Figures 6L–6N). Thus, the inhibitor results suggestthat H!/K!-ATPase activity normally prevents mem-an increase in fluorescence was seen to the left of the
streak, representing less negative membrane potentials, brane depolarization in the domain on the right side ofthe streak. These data suggest a model in which H!/i.e., depolarized relative to cells to the right of the streak
and throughout the epiblast (Figure 6B). At HH stage 3, K!-ATPase function is required to the right of the midlinein both Xenopus and chick embryos and that asymmetrythe domain of depolarized cells extended from the base
of the streak to Hensen’s node. The area of depolarized is achieved in Xenopus at the level of mRNA localizationwhile in the chick it is controlled downstream of mRNAcells was concentrated to the left side of Hensen’s node
in HH stage 4' embryos (Figure 6C) and gradually dimin- expression.ished by HH stage 4! (Figure 6D). The gradual increaseand later decrease in left-sided fluorescence is visible Discussionin linescans of these embryos (Figures 6E–6H). Meanfluorescence intensities were calculated from linescans We describe the first results of a pharmacological screen
using laterality of three visceral organs in Xenopus em-and the magnitude of left-sided fluorescence intensityover that on the right side of the streak was 19.1% bryos as an endpoint. The specificity of many of the hits
suggested the involvement of K! and H! flux and weat HH stage 3, rising to 21.2% at HH stage 4' , anddiminishing to 6.8% at HH stage 4! (Supplemental Table report the specific involvement of H!/K! ATPase func-
tion in both Xenopus and chick embryos.S3 available at http://www.cell.com/cgi/content/full/111/1/77/DC1). No differences could be seen at HHstages 1–2. A 1% difference in DiBAC4(3) fluorescence A Role for H!/K!-ATPase in LR Patterning
The distribution of organ reversals that arose from inhibi-corresponds to a membrane potential difference ofabout 1 mV (Brauner et al., 1984); thus, the magnitude tors of K! channels and H!/K!-ATPases indicated that
the defect elicited is heterotaxia, rather than isomerismof the endogenous left-sided depolarization is approxi-mately 10–20 mV. or situs inversus. Thus, the individual organs in the pre-
ponderance of treated embryos developed asymmetri-cally, but with frequent reversals and discordance (Fig-The Streak LR Voltage Asymmetry Is Due
to H!/K!-ATPase Function ure 1 and Supplemental Table S1 available at http://www.cell.com/cgi/content/full/111/1/77/DC1). Hetero-We then asked whether the asymmetry in membrane
voltage potential is due to the activity of the H!/K!- taxia, however, is a complex phenotype and three pointsshould be noted in interpreting the absolute incidencesATPase pump. HH stage 3–4 embryos were imaged as
before. The characteristic left-sided elevated fluores- arising from each trial. Firstly, drug doses were suffi-ciently low to avoid other development defects thatcence intensity was observed (Figure 6I) at which time
either omeprazole or BaCl2 was added to the culture. might indirectly affect laterality. Perturbation in dor-
Cell84
Figure 6. LR Asymmetries of Membrane Potential Patterns in the Primitive Streak Area Are Modulated by BaCl2 or Omeprazole
(A–D) A transient domain of depolarization to the left side of the primitive streak in chick embryos visualized with the potentiometric fluorescentprobe DiBAC4(3). The blue to green to red pseudo-color scale represents increasing fluorescence intensities reflecting increased accumulationof the anionic dye in intracellular membranes. Increased fluorescence corresponds to a less negative membrane potential (i.e., depolarized).(E–H) Linescans of the embryos made perpendicular to the streak (red lines in A-D) show the increased left sided fluorescence. SupplementalTable S3, available at http://www.cell.com/cgi/content/full/111/1/77/DC1, summarizes the mean LR differences from all embryos tested.(I–K) Typical LR asymmetric pattern of depolarization is seen prior to exposure (I). The streak is outlined in (I). A relative increase in right-sided fluorescence is seen in the same embryo as in (I) after exposure to BaCl2 (J). Similar increase in right-sided fluorescence after omeprazoleexposure (K). Arrowheads indicate Hensen’s node.(L–N) Linescans of the embryos (made along the red lines in I–K).
soanterior development, for instance, is commonly as- drugs were preincubated at pH 5.0 followed by neutral-ization (not shown), consistent with their pharmacologysociated with LR defects (Danos and Yost, 1995). In
our experiments, higher doses of NPPB, veratridine, or on gastric H!/K!-ATPases. Thus, we conclude that H!
and K! flux, in particular as dependent on H!/K!-ouabain caused AP/DV patterning defects and hetero-taxia in Xenopus. Secondly, although the maximum inci- ATPase, is crucial for LR patterning upstream of the
characterized cascades of asymmetric gene ex-dence of heterotaxia observed from the complete ran-domization of three independent organs would reach a pression.
Laterality defects have not been reported in trans-theoretical maximum of 87.5%, laterality of certain or-gans such as gallbladder and gut are not entirely inde- genic mice null for certain H!/K!-ATPase genes (Mene-
ton et al., 1998; Spicer et al., 2000). Several explanationspendent of one another (Casey and Hackett, 2000).Thirdly, a heterotaxic phenotype suggests that the initial could account for the apparent discrepancy between
the knockout phenotypes and our data. At least twobiasing mechanism that orients LR axis in the embryois either nonfunctional or uncoupled from organ mor- isoforms of the H!/K!-ATPase are known to exist and
it is plausible that compensatory or redundant functionphogenesis, a conclusion supported by the loss of sidedexpression of all normally asymmetrically expressed in the early embryo masks a laterality defect. Pharmaco-
logical studies, as we have done here, can be informativegenes examined (Figures 2, 3, and Supplemental TableS2 available at above website). In addition, SCH28080 because the relatively broad specificity of inhibitors
within the H!/K!-ATPase class allows targeting of multi-gave identical results to omeprazole and lansoprazole,in support of specificity. Finally, omeprazole and lanso- ple pump isoforms. To our knowledge, laterality defects
have not been reported in mammalian teratogenicityprazole are activated by acid and lower doses than de-scribed here were found to induce heterotaxia when the studies of H!/K!-ATPase inhibitors, but this does not

H!/K!-ATPase and Left-Right Asymmetry85
seem to have been specifically examined. Thus, it will al., 2000) and Ash1 in yeast (Beach et al., 2000; Munchowet al., 1999; Takizawa and Vale, 2000). Axonemal dyneinbe of interest to analyze phenotypes arising from combi-
natorial deletions of genes encoding H!/K!-ATPase iso- (Supp et al., 1997) and kinesins (Takeda et al., 1999)have been demonstrated genetically to be involved informs or from drug treatment in the mouse. In the ab-
sence of such data, it is not possible to predict whether LR asymmetry in the mouse where they are thoughtto drive monociliary movement at the node. Our data,or not LR asymmetry determination in the mouse in-
volves H!/K!-ATPases or another system of pumps and however, hint at novel functions for cytoplasmic motorsin the determination of LR asymmetry.channels. Pennekamp et al. (2002) provided evidence
for the latter by observing heterotaxia and perturbationsin asymmetric gene expression in a mouse with a tar- Models of Ion Flux in LR Patterninggeted mutation in the polycystic kidney disease (PCKD) The H!/K! exchanger works together with K! channelschannel. Interestingly, at least one ion pump gene (en- (Ikuma et al., 1998). The intracellular accumulation ofcoding the sodium-calcium exchanger NCX-1) is ex- K! ions is an active process mediated by a class ofpressed asymmetrically in Hensen’s node of later stage ATPases (P-type) that convert energy from the ATP/ADPchick embryos (Linask et al., 2001). ratio into an electrochemical K! gradient. Such ATPases
exchange K! ions reciprocally with either Na! or H!
ions, as for the H!/K!-ATPase implicated here in LRLocalization of H!/K!-ATPase mRNA in XenopusUnexpectedly, H!/K!-ATPase " mRNA in Xenopus is asymmetry determination. The K! accumulated in living
cells exits passively through a variety of K! channels.asymmetrically localized by the 4-cell stage (Figures4B–4F). This finding places a very early limit on when Thus, the net potassium current, IK, associated with H!/
K!-ATPase function leads to a more negative (i.e., morethe initial biasing mechanism operates in the embryo.The amphibian egg is radially symmetrical and the point polarized) membrane potential (Meneton et al., 1999).
Our measurements using DiBAC4(3) indicate that a do-of sperm entry determines the dorsoventral and antero-posterior body axes of the embryo (Elinson, 1975; Ger- main of depolarization exists on the left side of the primi-
tive streak in untreated HH stage 3 to 4 chick embryoshart et al., 1981; Scharf and Gerhart, 1980). LR asymme-try, therefore, is thought to be determined after (Figure 6, Supplemental Table S3 available at http://
www.cell .com/ cgi/content/ful l/111/1/77/D C1).fertilization. Our finding of consistently asymmetricmRNA in the 4-cell stage embryo suggests that the LR Treatment with the H!/K!-ATPase inhibitor omeprazole
or with BaCl2 leads to bilateral domains (Figure 6), indi-axis is oriented by about 2 hr of development in Xeno-pus. It is usually argued that LR asymmetry is deter- cating that the H!/K!-ATPase and IK normally functions
to maintain the more negative membrane potential onmined later during cleavage or gastrula stages in verte-brates. In mice, for instance, the earliest known the right side of the streak (Figure 7A). These data, to-
gether with the early right-sided localization of H!/K!-manifestation of LR asymmetry is the vortical rotationof monocilia on the ventral surface of node cells and ATPase mRNA in Xenopus and the observation that
drugs affecting K! flux cause heterotaxia and perturbthe resultant fluid flow has been proposed to initiateasymmetry determination (Nonaka et al., 2002, 1998; sided gene expression, implicate H!/K!-ATPase func-
tion as a very early step in LR asymmetry determination.Takeda et al., 1999). In the chick, the AP axis is firstmanifest by the location of Kohler’s sickle and primitive How might H!/K!-ATPase function regulate asym-
metric gene expression? A simple model is that H!/K!-streak in the blastoderm, which contains thousands ofcells at this time. Whether LR asymmetry follows from ATPase function directly regulates synthesis or secre-
tion of a diffusible determinant that then initiates thethis point, or if both LR and AP are patterned earlier inthe chick remain uncertain. There are reports, however, known cascades of asymmetrically expressed genes
(Figure 7B). Another possibility (Figure 7C) is that theof consistent LR differences in as early as the 2-cellstage in amphibians. These are manifest in the organ H!/K!-ATPase-dependent asymmetry in membrane
voltage in the left and right sides of the embryo mightsitus of embryos derived from separated left and rightblastomeres (Spemann, 1938) and in the number of mito- influence intracellular communication through gap junc-
tions. Although the endogenous pattern of gap junctionchondria that are found in each of the first 2 blastomeresin frog (Marinos, 1986). Indeed, Brown and Wolpert communication (GJC) is essential for LR asymmetry de-
termination in both chicks and Xenopus (Levin and Mer-(1990) presented a prescient model of asymmetry deter-mination in amphibians in which LR asymmetry determi- cola, 1998, 1999), GJC cannot be the initiating process,
since an upstream mechanism must orient the directionnation was linked directly to the process of cortical rota-tion that establishes the AP and DV axes. that putative LR determinants pass through the chan-
nels. Electrophoresis of determinants has been pro-Localization of H!/K!-ATPase " mRNA in Xenopusprecedes the known activation of the embryonic ge- posed for endogenous electric fields (Cooper et al.,
1989; Lange and Steele, 1978; Robinson and Messerli,nome at the MBT (stage 8) when the embryo has roughly1000 cells. Localization might involve selective mRNA 1996; Rose, 1970) and is known to operate in the trans-
port of material into Drosophila oocytes from sur-degradation or cytoplasmic transport prior to completeseparation of the cleaving blastomeres. Selective degra- rounding nurse cells (Diehl-Jones, 1993; Woodruff et al.,
1988; Woodruff and Telfer, 1980). By analogy, a voltagedation is thought to localize Drosophila mRNAs such asnanos in the posterior polar plasm (Bashirullah et al., difference, such as across the streak in the chick or
ventral midline in Xenopus, might direct the intercellular1999) and linkage to motor proteins mediates the trans-port of mRNAs such as bicoid and oskar in Drosophila propagation of low molecular weight, charged determi-
nants through gap junctions. Net asymmetric accumula-(Micklem et al., 2000; Schnorrer et al., 2000; Wilhelm et

Cell86
Figure 7. Models for Involvement of H!/K!-ATPase in Early LR Asymmetry
(A) Data presented here indicate that H!/K!-ATPase activity is localized to the right sideof early Xenopus and chick embryos. An ini-tiating mechanism to orient LR asymmetryhas not been described experimentally in ei-ther chick or Xenopus but is envisaged to beresponsible for the asymmetric H!/K!-ATPase activities, by localization of mRNA(Xenopus) or posttranslationally (chick), asdescribed in the text.(B) H!/K!-ATPase function might directlyregulate the secretion of an early determinantof asymmetric gene expression.(C) Alternatively, the H!/K!-ATPase might in-fluence the propagation of unknown, low mo-lecular weight LR determinants (green dots)between cells. Unidirectional propagationmight rely simply on electrophoresis ofcharged determinants through open gapjunction channels (depicted) or connexongating (see text). The segregation of determi-nants triggers asymmetric expression ofgenes in multicellular fields.
tion or localization might then bias asymmetric gene middle phase of LR patterning features large-scale,multi-cellular fields of gene expression. Our models haveexpression. An alternate model is that H!/K!-ATPase
activity might influence GJC via voltage and/or pH gating the advantage of using asymmetries in K! and H! fluxand GJC to leverage cell-based chirality into large-scale(Brink, 2000; Morley et al., 1997; White et al., 1994),
which has been demonstrated in the frog embryo (Turin multi-cellular asymmetry.and Warner, 1980).
Regardless of how H!/K!-ATPase function and K!Experimental Procedures
flux are coupled to gene expression, it is important toXenopus Embryologynote that asymmetric H!/K!-ATPase function cannot beXenopus embryos were dejellied in 2% cysteine 30( after artificialthe initiating step of LR asymmetry determination, sincefertilization and washed in 0.1$MMR. Batches of eggs from a singlesome upstream factor must localize H!/K!-ATPasefemale were divided into several experimental and control groups.
mRNA (as in Xenopus) or function (as in chick). One of Each group was put into either 10 ml of 0.1$MMR (Marc’s modifiedthe principal findings in this paper is that H!/K!-ATPase Ringer’s solution) (controls) or 0.1$MMR containing drugs at the
following doses: spermine, 395 )M; PCMB (p-chloromercuribenzoicmRNA becomes localized in the 4-cell Xenopus embryo,acid), 277 )M; amantadine, 266 )M; IAA-94, 55 )M; NPPB (PAA),thus restricting the timing of the initiating event to the3 )M; 9-AC (ACA), 68 )M; tetrodotoxin, 31 )M; veratridine, 0.2 )M;first few hours of development. Brown and Wolpertquinidine, 1.5 mM; lanthanum, 100 )M; amiodarone, 88 )M; barium(1990) proposed a conceptual model in which LR asym-chloride, 4 mM; Chromanol 293B, 100 )M; lansoprazole, 27 )M;
metry is polarized when a chiral molecule becomes ori- omeprazole, 28 )M; SCH28080, 100 )M; amiloride, 2.2 mM; EIPA,ented with respect to the AP and DV axes. It is easiest 83 )M; ouabain, 10 )M; cariporide, 2.6 mM; and aurovertin, 400
)M. In all cases, levels of vehicle (DMSO for compounds which areto envision the chiral molecule as subcellular, while the

H!/K!-ATPase and Left-Right Asymmetry87
not water soluble) was & 0.5% (which permits normal LR develop- bis-(1,3-dibutyrlbarbituric acid)thrimethine oxonol) [DiBAC4(3)] (Mo-lecular Probes Inc, Eugene, OR, USA; Epps et al., 1994). DiBAC4(3)ment). Embryos were allowed to develop in the drug-containing
medium until stage 16 (Nieuwkoop and Faber, 1967), at which point is an anionic fluorescent molecule that is distributed into intracellularmembranes depending on the electrical potential across the plasmathey were washed three times in 0.1$MMR and allowed to develop
in 0.1$MMR ! 0.1% gentamycin until stage 45. membrane. The more depolarized, i.e., the less negative the mem-brane potential is, the more anionic probe is accumulated, givingan increased fluorescent signal from the cell. The experiments wereXenopus Injectionsperformed in ovo. A round opening was cut in the egg after 8–18Capped, synthetic mRNA was transcribed in vitro (Ambion Messagehr of incubation at 38*C, and some of the thick albumen was removedMachine) from linearized plasmid templates. mRNA was mixed withby the use of a fire polished Pasteur pipette. A Plexiglas ring (diame-50 ng of rhodamine-lysinated dextran (10,000 MW, Molecularter 1.5 cm) was placed on the yolk surrounding the embryo to mini-Probes) and 320 pg of mRNA encoding #-galactosidase (as lineagemize the amount of chemicals used. DiBAC4(3) was dissolved inlabels) and injected into embryos within 30–45 min of fertilization.70% ethanol at 1 mg/ml and further diluted in deionized water to aThe approximate doses (per cell) were H!/K!-ATPase (Xenopus ",final concentration of 100 )g/ml. Portions of 100 )l from these100 pg) (Bufo #, 200 pg), mammalian Kir4.1/Bir10 (200 pg). H!/solutions were added to 900 )l of Panett-Compton buffer [pH 7.4],K!-ATPase cDNAs were obtained from Kaithi Geering (Institute ofto give final dye concentrations of 10 )g/ml, and the embryos werePharmacology and Toxicology, Switzerland); and the Kir4.1/Bir10incubated with the probe for 20–30 min at room temperature. ThecDNA was from John Adelman (Oregon Health Sciences University).fluorescence measurements were made in an upright epifluorescentmicroscope (Zeiss Axiophot) with a 2.5$ Neofluor lens (NA 0.075),Chick Embryologyand the images were sampled with a cooled CCD camera (PrincetonAll experimental manipulations were performed on standard patho-Instruments Inc., NJ, USA). High potassium concentrations (50 mM)gen-free white leghorn chick embryos obtained from Charles Riverwere used to test for dye efficacy and cell viability. This treatmentLaboratories (SPAFAS). Embryos were staged according to Ham-depolarized the cells, as seen by an increase in the fluorescence.burger and Hamilton (1992). Embryos were treated with inhibitorsIn the blocking experiments, 1 mg/ml BaCl2 or 100 )g/ml omeprazoleeither by addition to filter culture or by injection directly into the(Sigma Chemical Co. St. Louis, MO, USA) were added after thealbumin in ovo. Filter culture of control embryos does not alter20–30 min probe incubation or together with the probe at the startasymmetric gene expression (Levin and Mercola, 1999). Generally,of the incubation. The data were analyzed with a MetaMorph imagehowever, drug treatment in ovo yielded a lower incidence of asym-analysis program (Universal Imaging Corp., PA, USA). The fluores-metric gene expression, presumably because of a lower local con-cence (F) was calculated from a linescans of the left sided (FL %centration after injection into the egg. In ovo treatment was usedFleft/Fmin) and right-sided (FR % Fright/Fmin) fluorescences, respectively.for examination of heart looping (Figure 1) and for some of theFmin represents the lowest fluorescence intensity value in transverseembryos examined for Pitx2 and car expression (Supplemental Ta-linescans over the whole embryo; FL and FR were peak intensityble S2B and S2C available at http://www.cell.com/cgi/content/full/values. The difference between left- and right-sided fluorescence111/1/77/DC1). For in ovo exposure, 0.5 ml of drug at the statedis dF % FL-FR, and the change in fluorescence presented asdose (in albumin) was injected directly into the albumin of eachdF/FL$100. The values obtained were reproducible for different em-unincubated egg. The needle hole was covered with electrical tapebryos (Supplemental Table S3 available at http://www.cell.com/cgi/and the egg incubated in a nonrotating 38*C incubator. For culture,content/full/111/1/77/DC1).embryos were explanted in Pannett-Compton medium under a dis-
secting microscope. The embryo was placed ventral side upwardon a Costar 1 )m filter (catalog #110410) floating on top of medium Acknowledgmentsconsisting of 0.5 ml of Alpha-MEM medium (with or without drug)in 40 ml of thin albumin. Embryos were then cultured at 38*C until This paper is dedicated to Lionel Jaffe, whose research inspiredthe required stage of development. these studies. We thank Adam Crook, Debra Sorocco, and Shing-
Ming Cheng for their expert technical assistance. We acknowledgeScoring Xenopus Embryonic Situs AstraZeneca, Takeda Chemical Industries, and Schering-Plough Co.The phenotype of embryos was determined by scoring the situs of for their generous gifts of omeprazole, lansoprazole, and SCH28080,the heart, stomach, and gallbladder under a dissecting microscope respectively. Once commercially available, these drugs were pur-using tricaine to immobilize the stage 45 embryos. Only embryos chased from Sigma. We also thank Gary Yellen, Frederic Jaisser,with normal dorsoanterior development (DAI % 5) and clear left- and George Sachs for many helpful discussions, and Ralph Kentsided or right-sided organs were scored. Embryos with ambiguous for advice on statistical analysis. We are grateful for chick H!/K!-(unscoreable) situs comprised less than 5% of each experiment. A ATPase " cDNA from Frazer Murray; the chick H!/K!-ATPase #heterotaxic embryo was considered to be one in which any of those cDNA from K. Takeyasu; the Xenopus H!/K!-ATPase # and Bufo "three organs was reversed in its position. The incidences of organ cDNAs from Kaithi Geering; and the Kir4.1/Bir10 cDNA from Johnsitus were analyzed using the chi-square test with Pearson correc- Adelman. M.L. gratefully acknowledges a Helen Hay Whitney Foun-tion (for sample size). dation postdoctoral fellowship and grants from the American Cancer
Society (RSG-02-046-01), Chestnut Hill Charitable Foundation, andthe American Heart Association (Beginning Grant in Aid, 0160263T).In Situ HybridizationK.R.R. gratefully acknowledges support from the NSF (008517-IBN)Chick embryos were fixed in 4% paraformaldehyde at 4*C overnightand M.M. gratefully acknowledges grants from the NIH (RO1(for cultured embryos, filters with embryos were transferred to 4%HL63271) and American Heart Association (Established Investigatorparaformaldehyde and the embryos were carefully detached). FrogAward, 9740005N).embryos were fixed in MEMFA (Harland, 1991) at 4*C overnight.
cDNA used to transcribe cRNA in situ hybridization probes were:cShh and cNodal (Levin et al., 1995), cCerberus/caronte (Zhu et al., Received: December 12, 20011999), cPitx-2 (Logan et al., 1998), cfgf8 (Boettger et al., 1999; Ohuchi Revised: August 15, 2002et al., 2000), cWnt8c (Levin, 1998; Rodriguez-Esteban et al., 2001),cH!/K!-ATPase " (GenBank accession number AL588035, obtained
Referencesfrom Frazer Murray), cH!/K!-ATPase # (Yu et al., 1994), and XH!/K!-ATPase " (Mathews et al., 1995). Histological sections were
Altizer, A., Moriarty, L., Bell, S., Schreiner, C., Scott, W., and Borgens,obtained by embedding embryos after in situ hybridization in JB4R. (2001). Endogenous electric current is associated with normalaccording to manufacturer’s directions (Polysciences).development of the vertebrate limb. Dev. Dyn. 221, 391–401.
Bashirullah, A., Halsell, S.R., Cooperstock, R.L., Kloc, M., Kara-Chick Membrane Potential MeasurementsFor the study of membrane potential patterns, the embryos were iskakis, A., Fisher, W.W., Fu, W., Hamilton, J.K., Etkin, L.D., and
Lipshitz, H.D. (1999). Joint action of two RNA degradation pathwaysincubated in the fluorescent membrane potential sensitive probe

Cell88
controls the timing of maternal transcript elimination at the midblas- Cell and Molecular Biology, B.K. Kay, and H.B. Peng, eds. (SanDiego: Academic Press), pp. 685–695.tula transition in Drosophila melanogaster. EMBO J. 18, 2610–2620.
Harris, C., and Fliegel, L. (1999). Amiloride and the Na(!)/H(!) ex-Beach, D.L., Thibodeaux, J., Maddox, P., Yeh, E., and Bloom, K.changer protein: mechanism and significance of inhibition of the(2000). The role of the proteins Kar9 and Myo2 in orienting the mitoticNa(!)/H(!) exchanger (review). Int. J. Mol. Med. 3, 315–321.spindle of budding yeast. Curr. Biol. 10, 1497–1506.
Hotary, K., and Robinson, K. (1992). Evidence of a role for endoge-Boettger, T., Wittler, L., and Kessel, M. (1999). FGF8 functions innous electric fields in chick embryo development. Development 114,the specification of the right body side of the chick. Curr. Biol. 9,985–996.277–280.
Hotary, K.B., and Robinson, K.R. (1994). Endogenous electrical cur-Borgens, R., and Shi, R. (1995). Uncoupling histogenesis from mor-rents and voltage gradients in Xenopus embryos and the conse-phogenesis in the vertebrate embryo by collapse of the transneuralquences of their disruption. Dev. Biol. 166, 789–800.tube potential. Dev. Dyn. 203, 456–467.
Hyatt, B., and Yost, H. (1998). The left-right coordinator: the role ofBrauner, T., Hulser, D., and Strasser, R. (1984). Comparative mea-Vg1 in organizing left-right axis. Cell 93, 37–46.surements of membrane potentials with microelectrodes and volt-
age-sensitive dyes. Biochim. Biophys. Acta 771, 208–216. Ikuma, M., Binder, H., and Geibel, J. (1998). Role of apical H-Kexchange and basolateral K channel in the regulation of intracellularBrink, P. (2000). Gap junction voltage dependence: a clear picturepH in rat distal colon crypt cells. J. Membr. Biol. 166, 205–212.emerges. J. Gen. Physiol. 116, 11–12.Jaffe, L. (1981). The role of ionic currents in establishing develop-Brown, N., and Wolpert, L. (1990). The development of handednessmental pattern. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 295,in left/right asymmetry. Development 109, 1–9.553–566.Burdine, R., and Schier, A. (2000). Conserved and divergent mecha-Jiang, Y., and MacKinnon, R. (2000). The barium site in a potassiumnisms in left-right axis formation. Genes Dev. 14, 763–776.channel by x-ray crystallography. J. Gen. Physiol. 115, 269–272.Capdevila, J., Vogan, K., Tabin, C., and Belmonte, J.I. (2000). Mecha-Lange, C.S., and Steele, V.E. (1978). The mechanism of anterior-nisms of left-right determination in vertebrates. Cell 101, 9–21.posterior polarity control in planarians. Differentiation 11, 1–12.Casey, B. (1998). Two rights make a wrong: human left-right malfor-Levin, M. (1998). Left-right asymmetry and the chick embryo. Semin.mations. Hum. Mol. Genet. 7, 1565–1571.Cell Dev. Biol. 9, 67–76.Casey, B., and Hackett, B.P. (2000). Left-right axis malformationsLevin, M., and Mercola, M. (1998). Gap junctions are involved in thein man and mouse. Curr. Opin. Genet. Dev. 10, 257–261.early generation of left right asymmetry. Dev. Biol. 203, 90–105.Cooper, M.S., Miller, J.P., and Fraser, S.E. (1989). ElectrophoreticLevin, M., and Mercola, M. (1999). Gap junction-mediated transferrepatterning of charged cytoplasmic molecules within tissues cou-of left-right patterning signals in the early chick blastoderm is up-pled by gap junctions by externally applied electric fields. Dev. Biol.stream of Shh asymmetry in the node. Development 126, 4703–4714.132, 179–188.Levin, M., Johnson, R., Stern, C., Kuehn, M., and Tabin, C. (1995).Croyle, M.L., Woo, A.L., and Lingrel, J.B. (1997). Extensive randomA molecular pathway determining left-right asymmetry in chick em-mutagenesis analysis of the Na!/K!-ATPase alpha subunit identi-bryogenesis. Cell 82, 803–814.fies known and previously unidentified amino acid residues that
alter ouabain sensitivity—implications for ouabain binding. Eur. J. Linask, K., Han, M., Artman, M., and Ludwig, C. (2001). Sodium-Biochem. 248, 488–495. calcium exchanger (NCX-1) and calcium modulation: NCX protein
expression patterns and regulation of early heart development. Dev.Danos, M., and Yost, H. (1995). Linkage of cardiac left-right asymme-Dyn. 221, 249–264.try and dorsal-anterior development in Xenopus. Development 121,
1467–1474. Logan, M., Pagan-Westphal, S., Smith, D., Paganessi, L., and Tabin,C. (1998). The transcription factor Pitx2 mediates situs-specific mor-Diehl-Jones, W. (1993). Ionic basis of bioelectric currents duringphogenesis in response to left-right asymmetric signals. Cell 94,oogenesis in an insect. Dev. Biol. 158, 301–316.307–317.Elinson, R.P. (1975). Site of sperm entry and a cortical contractionLohr, J., Danos, M., and Yost, H. (1997). Left-right asymmetry of aassociated with egg activation in the frog Rana pipiens. Dev. Biol.nodal-related gene is regulated by dorsoanterior midline structures47, 257–268.during Xenopus development. Development 124, 1465–1472.Epps, D., Wolfe, M., and Groppi, V. (1994). Characterization of theMarinos, E. (1986). Observations on the mitochondrial distributionsteady-state and dynamic fluorescence properties of the potential-in normal, rotated and cold-treated 2-cell stage embryos of Xenopussensitive dye bis-(1,3-dibutylbarbituric acid)trimethine oxonollaevis. Cell Differ. 18, 163–171.(Dibac4(3)) in model systems and cells. Chem. Phys. Lipids 69,
137–150. Mathews, P.M., Claeys, D., Jaisser, F., Geering, K., Horisberger,J.D., Kraehenbuhl, J.P., and Rossier, B.C. (1995). Primary structureFujita, A., Horio, Y., Higashi, K., Mouri, T., Hata, F., Takeguchi, N.,and functional expression of the mouse and frog alpha-subunit ofand Kurachi, Y. (2002). Specific localization of an inwardly rectifyingthe gastric H(!)-K(!)-ATPase. Am. J. Physiol. 268, C1207–1214.K! channel, Kir4.1, at the apical membrane of rat gastric parietal
cells; its possible involvement in K! recycling for the H!-K! pump. Mathur, S., and Karmazyn, M. (1997). Interaction between anesthe-J. Physiol. 540, 85–92. tics and the sodium-hydrogen exchange inhibitor HOE 642 (caripo-
ride) in ischemic and reperfused rat hearts. Anesthesiology 87, 1460–Gerhart, J., Ubbels, G., Black, S., Hara, K., and Kirschner, M. (1981).1469.A reinvestigation of the role of the grey crescent in axis formation
in Xenopus laevis. Nature 292, 511–516. Meneton, P., Schultheis, P.J., Greeb, J., Nieman, M.L., Liu, L.H.,Clarke, L.L., Duffy, J.J., Doetschman, T., Lorenz, J.N., and Shull,Gottardi, C.J., and Caplan, M.J. (1993a). An ion-transporting ATPaseG.E. (1998). Increased sensitivity to K! deprivation in colonic H,K-encodes multiple apical localization signals. J. Cell Biol. 121,ATPase-deficient mice. J. Clin. Invest. 101, 536–542.283–293.Meneton, P., Lesage, F., and Barhanin, J. (1999). Potassium AT-Gottardi, C.J., and Caplan, M.J. (1993b). Molecular requirements forPases, channels, and transporters: an overview. Semin. Nephrol.the cell-surface expression of multisubunit ion-transporting AT-19, 438–457.Pases. Identification of protein domains that participate in Na,K-
ATPase and H,K-ATPase subunit assembly. J. Biol. Chem. 268, Mercola, M., and Levin, M. (2001). Left-right asymmetry determina-14342–14347. tion in vertebrates. Annu. Rev. Cell Dev. Biol. 17, 779–805.
Hamburger, V., and Hamilton, H. (1992). A series of normal stages Micklem, D.R., Adams, J., Grunert, S., and St Johnston, D. (2000).in the development of the chick embryo. Dev. Dyn. 195, 231–272. Distinct roles of two conserved Staufen domains in oskar mRNA
localization and translation. EMBO J. 19, 1366–1377.Harland, R.M. (1991). In situ hybridization: an improved whole mountmethod for Xenopus embryos. In Xenopus laevis: Practical Uses in Mizunashi, K., Furukawa, Y., Katano, K., and Abe, K. (1993). Effect

H!/K!-ATPase and Left-Right Asymmetry89
of omeprazole, an inhibitor of H!,K(!)-ATPase, on bone resorption dinates for the establishment of embryonic pattern. Dev. Dyn. 202,101–114.in humans. Calcif. Tissue Int. 53, 21–25.
Spemann, H. (1938). Embryonic Development and Induction (NewMorley, G.E., Ek-Vitorin, J.F., Taffet, S.M., and Delmar, M. (1997).Haven, CT: Yale University Press).Structure of connexin43 and its regulation by pHi. J. Cardiovasc.
Electrophysiol. 8, 939–951. Spicer, Z., Miller, M.L., Andringa, A., Riddle, T.M., Duffy, J.J.,Doetschman, T., and Shull, G.E. (2000). Stomachs of mice lackingMunchow, S., Sauter, C., and Jansen, R.P. (1999). Association ofthe gastric H,K-ATPase alpha-subunit have achlorhydria, abnormalthe class V myosin Myo4p with a localised messenger RNA in bud-parietal cells, and ciliated metaplasia. J. Biol. Chem. 275, 21555–ding yeast depends on She proteins. J. Cell Sci. 112, 1511–1518.21565.Munson, K.B., Lambrecht, N., and Sachs, G. (2000). Effects of muta-Supp, D., Witte, D., Potter, S., and Brueckner, M. (1997). Mutationtions in M4 of the gastric H!,K!-ATPase on inhibition kinetics ofof an axonemal dynein affects left-right asymmetry in inversus vis-SCH28080. Biochemistry 39, 2997–3004.cerum mice. Nature 389, 963–966.Nieuwkoop, P.D., and Faber, J. (1967). Normal Table of XenopusSupp, D., Brueckner, M., Kuehn, M., Witte, D., Lowe, L., McGrath,laevis (Daudin), Second Edition (Amsterdam: North-Holland Publish-J., Cirrakes, J., and Potter, S. (1999). Targeted deletion of the ATPing Company).binding domain of left-right dynein confirms its role in specifying
Nonaka, S., Tanaka, Y., Okada, Y., Takeda, S., Harada, A., Kanai,development of left-right asymmetries. Development 126, 5495–
Y., Kido, M., and Hirokawa, N. (1998). Randomization of left-right5504.
asymmetry due to loss of nodal cilia generating leftward flow ofTakeda, S., Yonekawa, Y., Tanaka, Y., Nonaka, Y.O.S., and Hiro-extraembryonic fluid in mice lacking KIF3B motor protein. Cell 95,kawa, N. (1999). Left-right asymmetry and kinesin superfamily pro-829–837.tein KIF3A. J. Cell Biol. 145, 825–836.
Nonaka, S., Shiratori, H., Saijoh, Y., and Hamada, H. (2002). Determi-Takeda, S., Yamazaki, H., Seog, D., Kanai, Y., Terada, S., and Hiro-nation of left-right patterning of the mouse embryo by artificial nodalkawa, N. (2000). Kinesin superfamily protein 3 (KIF3) motor trans-flow. Nature 418, 96–99.ports fodrin-associating vesicles important for neurite building. J.
Ohuchi, H., Kimura, S., Watamoto, M., and Itoh, N. (2000). Involve- Cell Biol. 148, 1255–1265.ment of fibroblast growth factor (FGF)18–FGF8 signaling in specifi-
Takizawa, P.A., and Vale, R.D. (2000). The myosin motor, Myo4p,cation of left-right asymmetry and brain and limb development ofbinds Ash1 mRNA via the adapter protein, She3p. Proc. Natl. Acad.the chick embryo. Mech. Dev. 95, 55–66.Sci. USA 97, 5273–5278.
Pagan-Westphal, S., and Tabin, C. (1998). The transfer of left-rightTurin, L., and Warner, A.E. (1980). Intracellular pH in early Xenopuspositional information during chick embryogenesis. Cell 93, 25–35.embryos: its effect on current flow between blastomeres. J. Physiol.
Pennekamp, P., Karcher, C., Fischer, A., Schweickert, A., Skryabin, 300, 489–504.B., Horst, J., Blum, M., and Dworniczak, B. (2002). The ion channel
Waddington, C. (1937). The dependence of head curvature on thepolycystin-2 is required for left-right axis determination in mice.
development of the heart in the chick embryo. J. Exp. Biol. 14,Curr. Biol. 12, 938–943.
229–231.Pizzonia, J.H., Ransom, B.R., and Pappas, C.A. (1996). Characteriza- Weber, J., and Senior, A.E. (1998). Effects of the inhibitors azide,tion of Na!/H! exchange activity in cultured rat hippocampal dicyclohexylcarbodiimide, and aurovertin on nucleotide binding toastrocytes. J. Neurosci. Res. 44, 191–198. the three F1-ATPase catalytic sites measured using specific trypto-Psychoyos, D., and Stern, C. (1996). Restoration of the organizer phan probes. J. Biol. Chem. 273, 33210–33215.after radical ablation of Hensen’s node and the anterior primitive White, T.W., Bruzzone, R., Goodenough, D.A., and Paul, D.L. (1994).streak in the chick embryo. Development 122, 3263–3273. Voltage gating of connexins. Nature 371, 208–209.Robinson, K., and Messerli, M. (1996). Electric embryos: the embry- Wilhelm, J.E., Mansfield, J., Hom-Booher, N., Wang, S., Turck, C.W.,onic epithelium as a generator of developmental information. In Hazelrigg, T., and Vale, R.D. (2000). Isolation of a ribonucleoproteinNerve Growth and Guidance, C. McCaig, ed. (Portland: Portland complex involved in mRNA localization in Drosophila oocytes. J.Press). Cell Biol. 148, 427–440.Rodriguez-Esteban, C., Capdevila, J., Kawakami, Y., and Izpisua Woodruff, R., and Telfer, W. (1980). Electrophoresis of proteins inBelmonte, J.C. (2001). Wnt signaling and PKA control Nodal expres- intercellular bridges. Nature 286, 84–86.sion and left-right determination in the chick embryo. Dev. Suppl. Woodruff, R., Kulp, J., and LaGaccia, E. (1988). Electrically mediated128, 3189–3195. protein movement in Drosophila follicles. Roux’s Archives of Dev.Rose, S.M. (1970). Differentiation during regeneration caused by Biol. 197, 231–238.migration of repressors in bioelectric fields. Am. Zool. 10, 91–99. Yost, H.J. (2001). Establishment of left-right asymmetry. Int. Rev.Sachs, G., Shin, J.M., Briving, C., Wallmark, B., and Hersey, S. (1995). Cytol. 203, 357–381.The pharmacology of the gastric acid pump: the H!,K! ATPase. Yu, H., Ishii, T., Pearson, W.R., and Takeyasu, K. (1994). PrimaryAnnu. Rev. Pharmacol. Toxicol. 35, 277–305. structure of avian H!/K(!)-ATPase #-subunit. Biochim. Biophys.Sanders, S.W., Tolman, K.G., Greski, P.A., Jennings, D.E., Hoyos, Acta 1190, 189–192.P.A., and Page, J.G. (1992). The effects of lansoprazole, a new Yuan, S., and Schoenwolf, G. (1998). De novo induction of the orga-H!,K(!)-ATPase inhibitor, on gastric pH and serum gastrin. Aliment. nizer and formation of the primitive streak in an experimental modelPharmacol. Ther. 6, 359–372. of notochord reconstitution in avian embryos. Development 125,
201–213.Scharf, S.R., and Gerhart, J.C. (1980). Determination of the dorsal-ventral axis in eggs of Xenopus laevis: complete rescue of uv- Zhu, L., Marvin, M., Gardiner, A., Lassar, A., Mercola, M., Stern, C.,impaired eggs by oblique orientation before first cleavage. Dev. Biol. and Levin, M. (1999). Cerberus regulates left-right asymmetry of the79, 181–198. embryonic head and heart. Curr. Biol. 9, 931–938.Schnorrer, F., Bohmann, K., and Nusslein-Volhard, C. (2000). Themolecular motor dynein is involved in targeting swallow and bicoidRNA to the anterior pole of Drosophila oocytes. Nat. Cell Biol. 2,185–190.
Shi, R., and Borgens, R. (1994). Embryonic neuroepithelial sodiumtransport, the resulting physiological potential, and cranial develop-ment. Dev. Biol. 165, 105–116.
Shi, R., and Borgens, R.B. (1995). Three-dimensional gradients ofvoltage during development of the nervous system as invisible coor-





























