Embed Size (px)
Citation preview

Plant Physiol. (1982) 70, 297-3020032-0889/82/70/0297/06/$00.50/0
Cell Division Cycles and Circadian Clocks`3PHASE-RESPONSE CURVES FOR LIGHT PERTURBATIONS IN SYNCHRONOUS CULTURES OFEUGLENA
Received for publication October 6, 1981 and in revised form February 25, 1982
LELAND N. EDMUNDS, JR., DORANNE E. TAY, AND DANIELLE L. LAVAL-MARTIN4Department ofAnatomical Sciences, School of Medicine, Health Sciences Center, State University ofNewYork, Stony Brook, New York 11794
ABSTRACT
The cell division rhythm in Eugkia gracilis Klebs (Z strain) freerunswith a circadian period (30.2 + 1.8 hours for 156 monitored oscillations) inaerated, magnetically stirred, 8-liter, axenic batch cultures grown photoau-totrophically at 25°C in LD: 3,3, (7,500 lux, cool-white fluorescent) 6-hourlight cycles from the moment of inoculation. Cell number was measured at2-hour intervals with an automatic fraction collector and Coulter ElectronicParticle Counter. At different circadian times throughout the 30-hourdivision cycle, 3-hour light perturbations were imposed on free-running cellpopulations by giving light during one of the intervals when dark wouldhave fallen in the LD: 3,3 regimen. Using the onset of division as the phasereference point, the net steady-state phase advance or delay (±4) of therhythm was determined after transients, if any, had subsided (usually inone or two days) relative to an unperturbed control culture. Both +A4 and-A0 were found, with maximum values of approximately ±11 to 12 hoursbeing obtained at circadian time (CT) 20 to 22 (the 'breakpoint'); little, ifany phase shift occurred if the light signal was given between CT 6 and CT12. The phase-resetting curve obtained by plotting new phase ()') versusold phase (4) was of the type 0 ('strong') variety. Light perturbations, nomatter when imposed, engendered new phases which mapped to a relativelyrestricted portion (CT 6 to CT 13) of the circadian cycle.
These data provide the first detailed phase-response curve for a circadianmitotic clock. The findings, therefore, not only further support the hypoth-esis that a circadian osciOlator (perhaps exhibiting limit cycle behavior) canmodulate cell division in eukaryotic cells, but also provide a useful basisfor the dissection of the nature and extent of the coupling between celldivision and circadian cycles.
type (33) to those exhibiting limit cycle behavior (18, 21, 23, 40).Additionally, the role of 'quantal' cells cycles (2, 19) and circadianoscillators (1, 7, 8, 10) and their interaction with the CDC5 havebeen considered.
Inherent in these models for cell cycle oscillators is the phenom-enon of phase perturbation: The response of cells to externalinfluences is often strongly dependent on the time of the CDC atwhich the agent is imposed. Thus, synchronous cultures of eu-karyotic unicells, such as the ciliate Tetrahymena pyriformis (32,42-45) or the fission yeast Schizosaccharomyces pombe (31, 45),and ofanimal cells, such as mouse fibroblast L cells (28) all exhibitincreased division setback (or "excess division delay") to heatshock and to a variety of chemical agents applied at progressivelylater times during their CDC. Perhaps more germane is the findingin Schizosaccharomyces (36) that the same agent can producedelays when imposed at one point of the CDC but advances whengiven at another: indeed, Klevecz and co-workers have discoveredthat synchronous mammalian V79 cells perturbed by serum (22),heat shock (23), or ionizing radiation (21) at 0.5-h intervals acrosstheir 8.5-h CDC display a biphasic, 4-h PRC comprising bothadvances and delays m subsequent cell divisions.
Despite these investigations, however, and chiefly for technicalreasons, a detailed PRC for any circadian mitotic clock has beenlacking. Inasmuch as the algal flagellate Euglena gracilis (Z) hasa well-defined circadian system with many mapped persistingcircadian rhythms (6, 9, 12), and because one of themost inten-sively studied periodicities has been that of cell division (5, 7, 10),we undertook an investigation to explore further the relationbetween the CDC and the circadian cycle in this organism. Thisstudy reports the derivation of a PRC for the phase-shifting effectsof light pertubations on the freerunning rhythm of cell division inEuglena.
Discussions of cell cycle control mechanisms (for reviews, see10, 15, 16, 26, 29) include the notion of timers and clocks inregulating cell cycle sequences culminating in mitosis and celldivision (10, 19, 27). These, in turn, embrace various types ofmacromolecular oscillators, ranging from those of the relaxation
'Dedicated to the memory of William S. Hiliman, critical scientist,colleague, and good friend.
2Supported in part by National Science Foundation Grant PCM76-10273 and United States Public Health Service Biomedical ResearchSupport Grant 2S07RR07067-16 to L. N. E.
3 Some of these results were reported at the XV International Congressof the International Society for Chronobiology, September 13-16, 1981,Minneapolis.
4 Permanent address: Laboratoire des Membranes Biologiques, Uni-versite Paris VII, 2, place Jussieu, 75005 Paris, France.
MATERIALS AND METHODS
Organism and Culture Conditions. The cells used in all experi-ments belonged to the Z strain of Euglena gracilis Klebs, which
5Abbreviations: CDC, cell division cycle; LL, continuous illumination;DD, constant darkness; LD: x,y, a repetitive light-dark cycle comprisingx hours of light and y hours of dark, whose period (T) is given by x + y;CR, circadian rhythm, whose free-running period is denoted by T (takenhere to be the time between onsets of cell division in a population of cells);4), phase of the rhythm; AOv, change in phase (phase shift); -OR, phasemarker, or phase reference point (here, the onset of the division burst; ss,average stepsize, or factorial increase in cell concentration (plateau toplateau) after a phased or synchronized division step; g, average generationtime (doubling time) of a population of cells; PRC, phase-responsecurve(s); CT, circadian time(s) (CT 0 indicates the phase point of a free-running rhythm that has been normalized to 24 h which occurs at the onsetof light in a LD: 12,12 reference cycle; ZT, environmental (Zeitgeber) time(where ZT 0 = onset of light).
297 www.plantphysiol.orgon June 23, 2018 - Published by Downloaded from
Copyright © 1982 American Society of Plant Biologists. All rights reserved.
Plant Physiol. Vol. 70, 1982
have been maintained in this laboratory since 1965. Axenic,aerated (575-625 ml min-'), magnetically stirred, 8-L batch cul-tures were grown photoautotrophically at 25°C in environmentalchambers on a modified Cramer and Myers medium, supple-mented with vitamins B1 and B12, as previously described (11).Illumination was provided by clock-programmed WestinghouseEcon-o-watt cool-white fluorescent bulbs (7,500 lux incident in-tensity). Cell number was monitored every 2 h by a miniaturizedfraction collector and automatic pipetting device (4, 11) and a
Coulter model ZB Electronic Particle Counter (Coulter Electron-ics, Hialeah, FL).Methodology for Deriving PRC. Newly inoculated cultures were
placed either into LL for 1 or 2 d and then into LD: 3,3, or directlyinto LD: 3,3; earlier results have indicated that this (13) andsimilar (I 1, 14) short-period regimens elicit free-running6 circadianrhythms of cell division. Growth curves were followed for severaldays in LD: 3,3 until T could be accurately defined. Subsequently,at a cell titer ranging between I x 104 to 4 x 104 cells ml , a 3-hlight signal was imposed on the culture at different CT, corre-
sponding to different phase points of the CDC, by replacing one
of the usual 3-h dark intervals in the LD: 3,3 regimen with lightof the same intensity as that ofthe preceding light interval (therebycreating a 9-h light span, whose midpoint was arbitrarily used insubsequent calculations). After any transients had subsided (in 1-2 d) and the steady-state, free-running period had been reestab-lished, the difference in phase (A4) between the shifted rhythm
LLmC)U-
0
z
Eug/enc grGc///s (Z)25Cc, 7700lux A
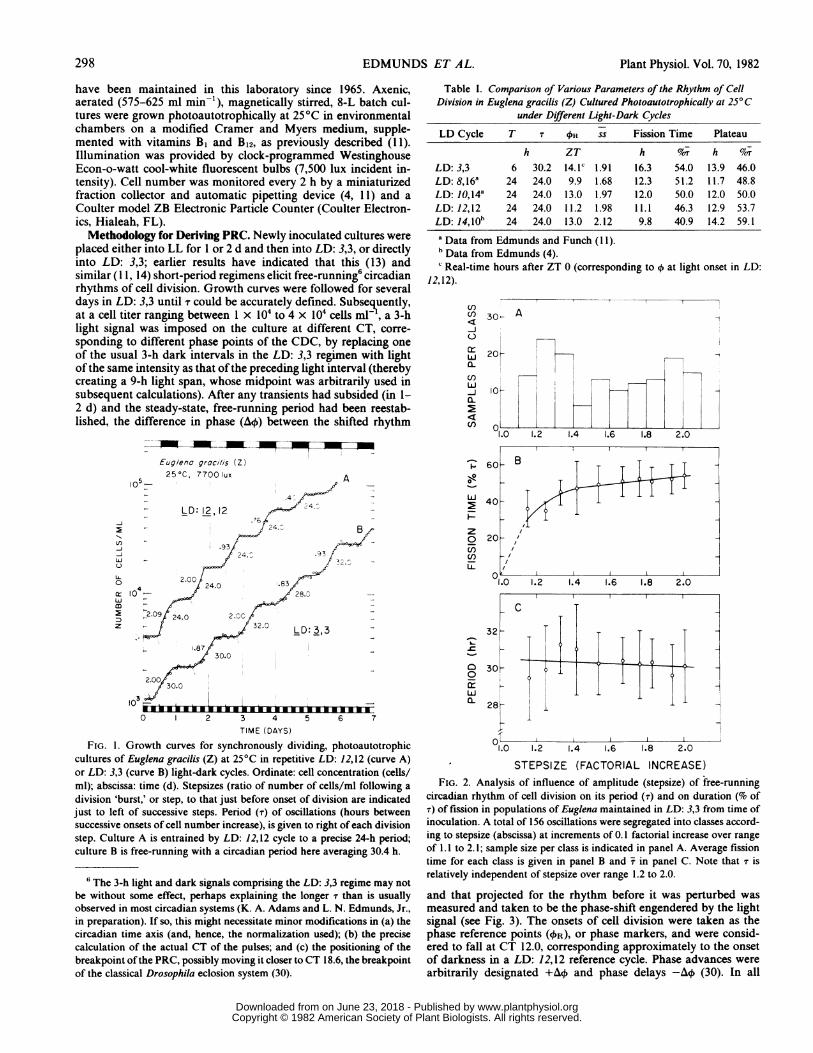
Table I. Comparison of Various Parameters of the Rhythm of CellDivision in Euglena gracilis (Z) Cultured Photoautotrophically at 25°C
under Different Light-Dark Cycles
LD Cycle T T Oi ss Fission Time Plateau
h ZT h N/r h NorLD: 3,3 6 30.2 14.1C 1.91 16.3 54.0 13.9 46.0LD: 8,16a 24 24.0 9.9 1.68 12.3 51.2 11.7 48.8LD: 10,14a 24 24.0 13.0 1.97 12.0 50.0 12.0 50.0LD: 12,12 24 24.0 11.2 1.98 11.1 46.3 12.9 53.7LD: 14,10b 24 24.0 13.0 2.12 9.8 40.9 14.2 59.1a Data from Edmunds and Funch (I 1).h Data from Edmunds (4).c Real-time hours after ZT 0 (corresponding to o at light onset in LD:
12,12).
Cl)
(I)
cIcn
-LJ
a-
(I)
(I)
30- A
60
0-
> 40
zo 20U)n)n
105
.4
12,20
40 J24. B
.93
_ 9 2~~~~4.^ .93J
24.0 .83
10 w 28.0-
X > ~~~32.0 LD:3,3 -
1.87 f/t30.0
2.0030.0
0 2 3 4
TIME (DAYS)
-
0
0~
crLL
5 6 7
FIG. 1. Growth curves for synchronously dividing, photoautotrophiccultures of Euglena gracilis (Z) at 25°C in repetitive LD: 12,12 (curve A)or LD: 3,3 (curve B) light-dark cycles. Ordinate: cell concentration (cells/ml); abscissa: time (d). Stepsizes (ratio of number of cells/ml following a
division 'burst,' or step, to that just before onset of division are indicatedjust to left of successive steps. Period (T) of oscillations (hours betweensuccessive onsets of cell number increase), is given to right of each divisionstep. Culture A is entrained by LD: 12,12 cycle to a precise 24-h period;culture B is free-running with a circadian period here averaging 30.4 h.
6The 3-h light and dark signals comprising the LD: 3,3 regime may notbe without some effect, perhaps explaining the longer T than is usuallyobserved in most circadian systems (K. A. Adams and L. N. Edmunds, Jr.,in preparation). If so, this might necessitate minor modifications in (a) thecircadian time axis (and, hence, the normalization used); (b) the precisecalculation of the actual CT of the pulses; and (c) the positioning of thebreakpoint ofthe PRC, possibly moving it closer to CT 18.6, the breakpointof the classical Drosophila eclosion system (30).
w
1.0 1.2 1.4 1.6 1.8 2.0
28
Th.0 1.2 1.4 1.6 1.8 2.0
STEPSIZE (FACTORIAL INCREASE)FIG. 2. Analysis of influence of amplitude (stepsize) of free-running
circadian rhythm of cell division on its period (T) and on duration (% ofT) of fission in populations of Euglena maintained in LD: 3,3 from time ofinoculation. A total of 156 oscillations were segregated into classes accord-ing to stepsize (abscissa) at increments of 0.1 factorial increase over rangeof 1.1 to 2.1; sample size per class is indicated in panel A. Average fissiontime for each class is given in panel B and X in panel C. Note that T isrelatively independent of stepsize over range 1.2 to 2.0.
and that projected for the rhythm before it was perturbed was
measured and taken to be the phase-shift engendered by the lightsignal (see Fig. 3). The onsets of cell division were taken as thephase reference points (OR), or phase markers, and were consid-ered to fall at CT 12.0, corresponding approximately to the onsetof darkness in a LD: 12,12 reference cycle. Phase advances were
arbitrarily designated +A4 and phase delays -A4 (30). In all
,,B
298 EDMUNDS ET AL.
www.plantphysiol.orgon June 23, 2018 - Published by Downloaded from Copyright © 1982 American Society of Plant Biologists. All rights reserved.

CELL DIVISION CYCLES AND CIRCADIAN CLOCKS
cn-J
U-
0
z: 104
z
TIME (hr)
FIG. 3. Examples and methodology used in calculation of phase-shiftdelays (-A4) and advances (+A4) of free-running rhythm of cell divisionobtained in cultures of Euglena maintained in LD: 3,3 at 25°C andperturbed by 3-h light (7,500 lux) signals given at different circadian times.Two in-phase, synchronously dividing cultures, at approximately the sametiter (ordinate) and exhibiting a stable period of 30 h (vertical dashedlines) over a time span of at least 72 h (abscissa), were subjected to a lightpulse whose midpoint occurred at either hour 79.5 or hour 85.5, corre-
sponding to circadian times CT 19.6 or CT 24.0 (=CT 0), respectively.Onsets of cell division were taken as phase-reference points (4R) andconsidered to fall at CT 12.0, corresponding to onset of darkness in LD:12,12. After transients had subsided and free-running T of 30 h had beenreestablished, the difference in phase (AO) between shifted rhythm (verticaldotted lines) and that projected (solid markers) for rhythm before it wasperturbed was measured and taken to be steady-state phase shift engen-
dered by light signal. Other notation as for Figure 1.
cases, both CT and data for AO were normalized to 24 h tofacilitate comparison among different values of T.
RESULTS
Characterization of the Free-Running Circadian Rhythm of CellDivision. Growth curves for typical cultures of Euglena dividingsynchronously in a reference LD: 12,12 cycle and in the experi-mental LD: 3,3 short-period (T = 6 h) regimen are shown inFigure 1. In the former cycle, the rhythm is directly synchronized,or entrained, to a precise 24.0-h period, matching that of theimposed Zeitgeber. The population approximately doubles (ss =2.00) during each dark interval until limiting light intensities are
reached (due to mutual shading by the cells); but even when ss
< 2.0, the period as displayed by the population averages 24 h (4,1 1). In the LD: 3,3 regimen, however, a different situation obtains:the cell division rhythm freeruns, exhibiting a period of approxi-mately 30 h. These longer periods are similar to those found alsoin other short-period (such as LD: 1/3,1/3 or LD: 1,1) and even
'random' light cycles (1 1, 13, 24) and are in striking contrast to theprecise 24-h periods observed under directly entraining LD cycles(for example, LD: 8,16 or LD: 10,14) having driving periods of 24h (Table I).A statistical analysis of a total of 156 oscillations in the cell
division rhythm displayed by cultures maintained in LD: 3,3yielded a value for T of 30.21 + 1.85 h. (In other strains, under the
D 8 10 12 14 16
CIRCADIAN TIME (hr)
FIG. 4. Phase-response curve for action of 3-h light signals (7,500 lux)on free-running rhythm of cell division in photoautotrophic cultures ofEuglena maintained in LD: 3,3. Steady-state phase shift (±A4), determinedas described in legend to Figure 3, is plotted as a function of CT(normalized to 24 h) at which midpoint of signal was given. "Breakpoint,"indicated by dashed line between CT 20 and 22, would disappear if curvewere drawn in monotonic form with all phase shifts being treated asdelays.
identical light regimen, values of T may be different, sometimesaveraging as little as 27 or 28 h, indicating that entrainment to 30-h period by a 5-fold frequency demultiplication of the shorter, 6-h driving period was not occurring.) This value for the free-running period for all intents and purposes was independent ofthe amplitude of the rhythm (Fig. 2, C): T was conserved over ssranging between 1.12 and 2.20. Similarly, the duration of thefission burst during any given oscillation was relatively unaffectedby ss over the range 1.4 to 2.0 (Fig. 2, B). Values of T over theentire range of cell concentration utilized in our batch cultures,therefore, could be taken with confidence.
Derivation of a Phase-Response Curve for Light Signals. Oncethe free-running rhythm of cell division in LD: 3,3 had beencharacterized, it was systematically scanned by single 3-h lightperturbations (replacing a given dark interval in the LD cycle)imposed at different subjective CT (corresponding to differentphases of the 30-h CDC). Two such phase-shifting experiments,with signals being given at CT 0 (corresponding to the beginningof the CDC at the onset of light in a reference LD: 12,12 cycle)and at CT 19.6 (middle of the division burst), are illustrated inFigure 3. In both cases, a large phase shift occurred but ofoppositesign (+ 12.0 h and -8.0 h, respectively, on a real-time scale). Theexperiments were facilitated by the high stability of the free-running period, both before and after the light perturbation.
This type of experiment yielded a family of 20 curves fromwhich a classic phase-response curve (30, 41) was generated (Fig.4). It comprised a portion of (CT 22 to CT 6) yielding phaseadvances as great as 11 to 12 h, a plateau (CT 6 to CT 12)relatively "insensitive" to light perturbations, and a descendinglimb (CT 12 to CT 20) giving phase delays. The discontinuity, or"breakpoint" ("phase-jump"), inherent in this type of plot laybetween CT 20 and CT 22, where delays became advances. If allphase shifts were treated alternatively as delays (with the advancesbeing displaced 3600), the breakpoint would disappear and onewould obtain a smooth, monotonic form of the PRC. The precisepositioning of the PRC was dependent on two factors. By conven-tion, we used the midpoint of the light signal as the CT of thepulse, but if we had instead chosen the onset (or end) of the signal,
299
www.plantphysiol.orgon June 23, 2018 - Published by Downloaded from Copyright © 1982 American Society of Plant Biologists. All rights reserved.
Plant Physiol. Vol. 70, 1982
+
w
z
-j
C
I-
11J
z
0
z
CIRCADIAN TIME OF SIGNAL (4)
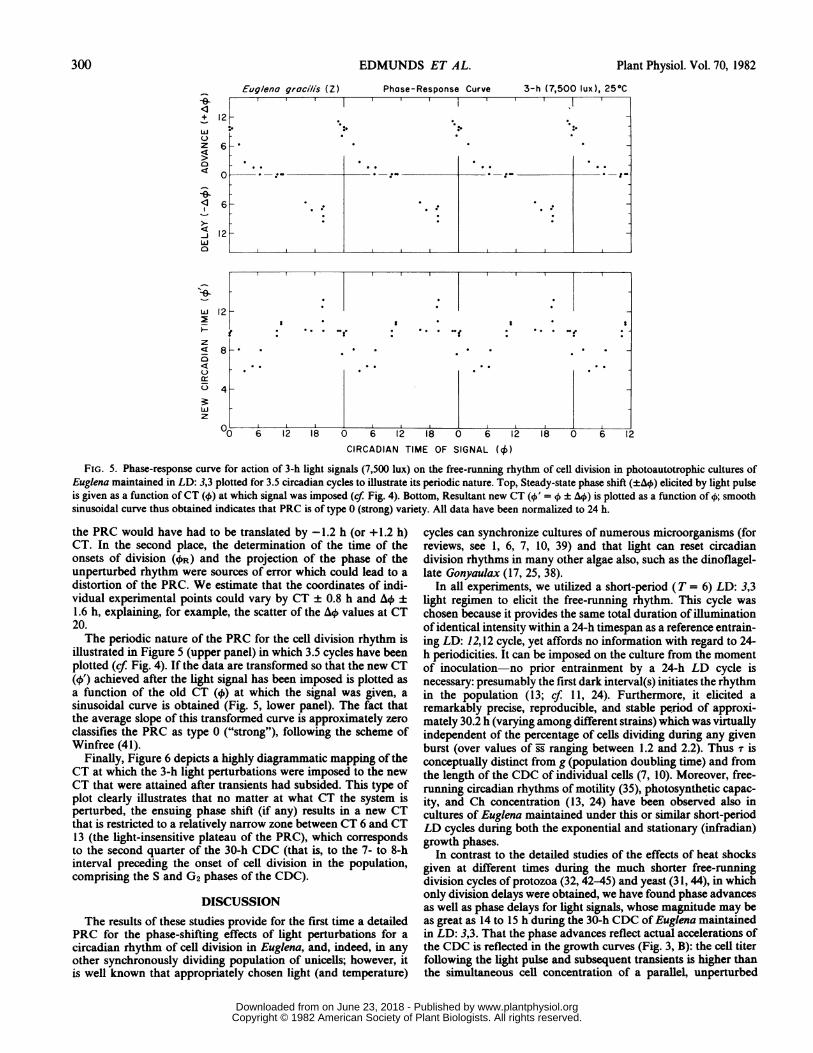
FIG. 5. Phase-response curve for action of 3-h light signals (7,500 lux) on the free-running rhythm of cell division in photoautotrophic cultures ofEuglena maintained in LD: 3,3 plotted for 3.5 circadian cycles to illustrate its periodic nature. Top, Steady-state phase shift (±AO) elicited by light pulseis given as a function ofCT (4)) at which signal was imposed (cf. Fig. 4). Bottom, Resultant new CT (4)' = ± AO\) is plotted as a function of 4); smoothsinusoidal curve thus obtained indicates that PRC is of type 0 (strong) variety. All data have been normalized to 24 h.
the PRC would have had to be translated by -1.2 h (or +1.2 h)CT. In the second place, the determination of the time of theonsets of division (4)R) and the projection of the phase of theunperturbed rhythm were sources of error which could lead to a
distortion of the PRC. We estimate that the coordinates of indi-vidual experimental points could vary by CT ± 0.8 h and AO ±
1.6 h, explaining, for example, the scatter of the AO) values at CT20.The periodic nature of the PRC for the cell division rhythm is
illustrated in Figure 5 (upper panel) in which 3.5 cycles have beenplotted (cf. Fig. 4). If the data are transformed so that the new CT(41) achieved after the light signal has been imposed is plotted asa function of the old CT (4) at which the signal was given, asinusoidal curve is obtained (Fig. 5, lower panel). The fact thatthe average slope of this transformed curve is approximately zeroclassifies the PRC as type 0 ("strong"), following the scheme ofWinfree (41).
Finally, Figure 6 depicts a highly diagrammatic mapping of theCT at which the 3-h light perturbations were imposed to the newCT that were attained after transients had subsided. This type ofplot clearly illustrates that no matter at what CT the system isperturbed, the ensuing phase shift (if any) results in a new CTthat is restricted to a relatively narrow zone between CT 6 and CT13 (the light-insensitive plateau of the PRC), which correspondsto the second quarter of the 30-h CDC (that is, to the 7- to 8-hinterval preceding the onset of cell division in the population,comprising the S and G2 phases of the CDC).
DISCUSSION
The results of these studies provide for the first time a detailedPRC for the phase-shifting effects of light perturbations for a
circadian rhythm of cell division in Euglena, and, indeed, in anyother synchronously dividing population of unicells; however, itis well known that appropriately chosen light (and temperature)
cycles can synchronize cultures of numerous microorganisms (forreviews, see 1, 6, 7, 10, 39) and that light can reset circadiandivision rhythms in many other algae also, such as the dinoflagel-late Gonyaulax (17, 25, 38).
In all experiments, we utilized a short-period (T = 6) LD: 3,3light regimen to elicit the free-running rhythm. This cycle was
chosen because it provides the same total duration of illuminationof identical intensity within a 24-h timespan as a reference entrain-ing LD: 12,12 cycle, yet affords no information with regard to 24-h periodicities. It can be imposed on the culture from the momentof inoculation-no prior entrainment by a 24-h LD cycle isnecessary: presumably the first dark interval(s) initiates the rhythmin the population (13; cf. 11, 24). Furthermore, it elicited aremarkably precise, reproducible, and stable period of approxi-mately 30.2 h (varying among different strains) which was virtuallyindependent of the percentage of cells dividing during any givenburst (over values of ss ranging between 1.2 and 2.2). Thus T isconceptually distinct from g (population doubling time) and fromthe length of the CDC of individual cells (7, 10). Moreover, free-running circadian rhythms of motility (35), photosynthetic capac-ity, and Ch concentration (13, 24) have been observed also incultures of Euglena maintained under this or similar short-periodLD cycles during both the exponential and stationary (infradian)growth phases.
In contrast to the detailed studies of the effects of heat shocksgiven at different times during the much shorter free-runningdivision cycles of protozoa (32, 42-45) and yeast (31, 44), in whichonly division delays were obtained, we have found phase advancesas well as phase delays for light signals, whose magnitude may beas great as 14 to 15 h during the 30-h CDC of Euglena maintainedin LD: 3,3. That the phase advances reflect actual accelerations ofthe CDC is reflected in the growth curves (Fig. 3, B): the cell titerfollowing the light pulse and subsequent transients is higher thanthe simultaneous cell concentration of a parallel, unperturbed
12 2
Kl 6 1I 2 18 o 6 1 18 0 6 12 18 0 6 12
300 EDMUNDS ET AL.
www.plantphysiol.orgon June 23, 2018 - Published by Downloaded from Copyright © 1982 American Society of Plant Biologists. All rights reserved.

CELL DIVISION CYCLES AND CIRCADIAN CLOCKS
.~~~~~2:\'1i /:2i0~~~~~~~~~NI5
0-~ ~ -
FIG. 6. Diagrammatic mapping ofCT at which 3-h phase-shifting lightperturbations were given (0) to new CT that were attained (0) for free-running rhythm of cell division in photoautotrophic cultures of Euglenamaintained in LD: 3,3 at 25°C. Inner ring represents circadian cycle (T =
30 h) normalized to 24 h for convenience, hour zero (CT 0 = CT 24)corresponds to onset of light in LD: 12,12, and stippled area beginning atCT 12 represents subjective night during which cell division occurs. Notethat no matter when system is perturbed, ensuing phase shift (if any)results in attainment of a new CT that is restricted to a relatively narrow
zone between CT 6 and CT 13 (cf. Fig. 5, bottom). Arcs (dashed lines)connecting old and new phase points are provided merely to facilitatevisualization of phase shifts and probably do not represent their actualtrajectories.
control. Although this might be attributed to the additional energyfor photosynthetic growth afforded by the 3-h light signal itself,the same signal given at other CT results in a reduction in growth(titer achieved), lengthening of the CDC, and a phase delay in thecell division rhythm (Fig. 3, A). An additional implication of thesedata (and suggested also by the cell cohorts that do not divideduring a given division burst in an unperturbed population) is thatthe circadian clock continues to function and measure time eventhough mitosis may be skipped during a particular circadian cycle.The PRC for cell division in Euglena (Fig. 4) comprises a large
advance portion during the late subjective night (CT 22 to CT 24)and early subjective day (CT 0 to CT 6), a "plateau" segment (CT6 to CT 12) relatively insensitive to light signals, a strong delayportion (CT 12 to CT 22) during the subjective night, and abreakpoint, or phase-jump (at approximately CT 22, or perhapsearlier) where the sign of the engendered phase shift reverses. Itsgeneral form is remarkably similar to that found for the PRC fora variety of other circadian rhythms in Drosophila and other lowerorganisms (30, 41). It also resembles those derived for the rhythmsof phototaxis (14) and dark motility (35) in Euglena, if allowancesare made for the different methods used in plotting. (Note thatthe precise positioning of our PRC along either axis could vary by2 or 3 h, particularly if the LD: 3,3 cycle is assumed to have someeffect on the rhythm.) If all phase shifts elicited by the lightperturbations are treated as delays, a smooth monotonic curve isobtained, with a maximum delay of the rhythm (approximately
equal to t) being found at CT 6. Whether or not the discontinuityin the PRC is "real" (in the sense of reflecting some discretemetabolic change in the cellular oscillator) or merely an "artifact"in plotting the data is still an open question (41), although it istempting to speculate about short "trigger-phase" intervals leadingto mitosis. Finally, the PRC falls into the class denoted by Winfree(41) as type 0 (or strong), as is seen most readily by plotting thenew phase achieved (4/) following a light-induced phase shift asa function of the CT (4) at which the perturbation was adminis-tered (Fig. 5, lower panel): A sinusoidal oscillation is obtainedwith values of 4' falling into a zone ranging between CT 6 andCT 13.
Several other interesting features of the PRC emerge. In thefirst place, the CT at which increasing delay or maximum advancephase shifts are achieved correspond, respectively, to the positionof the CDC during which division occurs (commencing at ap-proximately CT 12) and to the first few hours of the G1 phase(This correspondence cannot be resolved precisely, however, in-asmuch as the values of ss for the division cycles perturbed bylight signals occasionally were a little less than 2.00 jss = 1.91; seeTable I], indicating some heterogeneity in CDC states within thepopulation.) These times, furthermore, are exactly those when thefree-running rhythm of photosynthetic capacity (13, 24) in LD:3,3 displays the lowest values; maximum values (CO2 fixed/cell/h) occur at the very time (CT 6 to CT 12) that 3-h light signals arevirtually ineffective in phase shifting the cell division rhythm.These observations demonstrate that although light is needed asa substrate for photosynthesis and the progression of the CDC inphotoautotrophically cultured Euglena, it serves a quite distinctand separable function in phase shifting and entraining the cir-cadian oscillator(s) hypothesized to underlie the circadian rhythmof cell division (7, 10), whose rhythmic sensitivity to light signalsis reflected in the PRC. Finally, our data show (Fig. 6; cf. Fig. 5,lower panel) that no matter at what time a light signal is imposedduring the circadian cycle, the resulting new CT achieved aftertransients have subsided are limited to the range between approx-imately CT 6 and CT 13 (the second half of the subjective day,prior to the onset of division and corresponding to the S and G2CDC phases [4, 10]).We have no evidence, despite our relatively detailed PRC, that
a higher frequency quantal component (Gq) exists for Euglena, ashas been suggested by Klevecz (19, 20) for mammalian cells onthe basis of biphasic PRC obtained for serum pulses, heat shocks,and ionizing radiation administered at different intervals duringthe CDC (21-23). (Of course, the "gating" of cell division tocircadian "windows" constitutes a quantizement of g in itself,where Gq = t.) Indeed, Sweeney (37) has suggested that thispossibility of Gq cycling has been excluded in the large, slowlydividing (g = 5 to 6 d) dinoflagellate, Pyrocystisfusiformis Murray,in which five morphological stages of the CDC in addition todivision all appear to be gated, or phased, by a circadian clock inisolated cells. As Klevecz and co-workers (19, 23) have cautioned,however, heterogeneity of cell generation times in populations ofcells makes the detection of a short Gq difficult in lines havinglonger values of g.The derivation of a PRC for the cell division rhythm in Euglena
now allows us not only to proceed with a formal analysis (30, 41)of the putative light-sensitive circadian oscillator and its entrain-ment by light cycles (and other Zeitgeber), but also to undertakeother experimental lines of investigation on its mechanism. Itwould be particularly interesting to know whether or not a similarPRC could be obtained for pulses of inhibitors of metabolism andmacromolecular synthesis, as has been done with cycloheximidefor the CDC of Schizosaccharomyces (31) and Physarum (34), andmost recently for the circadian rhythm of bioluminescence inGonyaulax (3, 39), in which the PRC thus obtained was similar tothat for light (39).
301
www.plantphysiol.orgon June 23, 2018 - Published by Downloaded from Copyright © 1982 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 70, 1982
Acknowledgment-The authors thank Edmund S. Monchak for his expert technicalassistance and help in carrying out some of these experiments.
LITERATURE CITED
1. CHISHOLM SW 1981 Temporal patterns of cell division in unicellular algae. In TPlatt, ed, Physiological Phytoplankton Ecology. Published for the NATOAdvanced Study Institute. Lipari, Italy In press
2. CHISHOLM SW, FMM MOREL, WS SLOCUM 1980 The phasing and distributionof cell division cycles in marine diatoms. In PG Falkowski, ed, PrimaryProductivity in the Sea. Brookhaven Symposium Biology No. 31. PlenumPress, New York, pp 281-300
3. DUNLAPJC, WT TAYLOR, JW HASTINGS 1980 The effects of protein synthesisinhibitors on the Gonyaulax clock. I. Phase-shifting effects of cycloheximide.J Comp Physiol B 138: 1-8
4. EDMUNDS LN JR 1964 Replication of DNA and cell division in synchronouslydividing cultures of Euglena gracilis. Science 145: 266-268
5. EDMUNDS LN JR 1966 Studies on synchronously dividing cultures of Euglenagracilis Klebs (strain Z). III. Circadian components of cell division. J CellPhysiol 67: 35-44
6. EDMUNDS LN JR 1975 Temporal differentiation in Euglena: circadian phenomenain non-dividing populations and in synchronously dividing cells. In: M Lefort-Tran, R Valencia, eds., Les Cycles Cellulaires et leur Blocage chez PlusieursProtistes. ColloquesInt. C.N.R.S. No. 240. Centre National dela RechercheScientifique, Paris, pp 53-67
7. EDMUNDS LN JR 1978 Clocked cell cycle clocks: implications toward chrono-pharmacology and aging. In HV Samis Jr, S Capobianco, eds, Aging andBiological Rhythms. Plenum, New York, pp 125-184
8. EDMUNDS LN JR 1981 Clocked cell cycle clocks: ultradian, circadian, andinfradian interfaces. In HG Schweiger, ed, International Cell Biology 1980-1981. Springer-Verlag, Berlin and Heidelberg, pp 831-845
9. EDMUNDS LN JR 1981 Circadian and infradian rhythms. In DE Buetow, ed, TheBiology of Euglena, Vol 3. Academic Press, New York In press
10. EDMUNDS LN JR, KJ ADAMS 1981 Clocked cell cycle clocks. Science 211: 1002-1013
11. EDMUNDS LN JR, R FUNCH 1969 Effects of "skeleton" photoperiods and highfrequency light-dark cycles on the rhythm of cell division in synchronizedcultures of Euglena. Planta 87: 134-163
12. EDMUNDS LN JR, F HALBERG 1981 Circadian time structure of Euglena: a modelsystem amenable to quantification. In HE Kaiser, ed, Neoplasms-Compara-tive Pathology of Growth in Animals, Plants and Man. Williams & Wilkins,Baltimore, pp 105-134
13. EDMUNDS LN JR, DL LAVAL-MARTIN 1981 "Free-running" circadian rhythms ofphotosynthesis elicited by short-period cycles of light and darkness in synchro-nously dividing and nondividing Euglena. In G Akoyunoglou, ed, Photosyn-thesis. Proceedings of 5th International Congress on Photosynthesis, September7-13, 1980, Halkidiki, Greece, Vol VI. Balaban International Science Services,Jerusalem, pp 313-322
14. FELDMAN JF 1967 Lengthening the period of a biological clock in Euglena bycycloheximide, an inhibitor of protein synthesis. Proc Natl Acad Sci USA 73:4012-4016
15. GILBERT DA 1980 Mathematics and cancer. In W Getz, ed, MathematicalModelling in Biology and Ecology. Lecture Notes in Biomathematics, Vol 33.Springer-Verlag, New York, pp 97-116
16. HARTWELL L, J CULATTI, JR PRINGLE, BT REID 1974 Genetic control of the celldivision cycle in yeast. Science 183: 46-51
17. HASTINGS JW, BM SWEENEY 1964 Phased cell division in marine dinoflagellates.In E Zeuthen, ed, Synchrony in Cell Division and Growth. Interscience, Wiley& Sons, New York, pp 307-321
18. KAUFFMAN S, JJ WILLE 1975 The mitotic oscillator in Physarum polycephalum.J Theor Biol 55: 47-93
19. KLEVECZ RR 1976 Quantized generation time in mammalian cells as an expres-sion of the cellular clock. Proc Natl Acad Sci USA 73: 4012-4016
20. KLEVECZ RR 1978 The clock in animal cells is a limit cycle oscillator. In ER
Dirksen, DM Prescott, CF Fox, eds, Cell Reproduction: in Honor of DanielMazia. Academic Press, New York, pp 139-146
21. KLEVECz RR, GA KING, RM SHYMKO 1980 Mapping the mitotic clock by phaseperturbation. J Supramol Struct 14: 329-342
22. KLEVECZ RR, J KROS, SD GROSS 1978 Phase response versus positive andnegative division delay in animal cells. Exp Cell Res 116: 285-290
23. KLEVECZ RR,J KROs, GA KING 1980 Phase response to heat shock as evidencefor a timekeeping oscillator in synchronous animal cells. Cytogenet Cell Genet26: 236-243
24. LAVAL-MARTIN DL, DJ SHUCH, LN EDMUNDS JR 1979 Cell cycle-related andendogenously controlled circadian photosynthetic rhythms in Euglena. PlantPhysiol 63: 495-502
25. MCMURRY L, JW HASTINGS 1972 No desynchronization among four differentcircadian rhythms in the unicellular alga, Gonyaulax polyedra. Science 175:1137-1139
26. MITCHISON JM 1971 The Biology of the Cell Cycle. Cambridge University Press,London
27. MITCHISON JM 1974 Sequences, pathways and timers in the cell cycle. In GMPadilla, IL Cameron, A Zimmerman, eds, Cell Cycle Controls. AcademicPress, New York, pp 125-142
28. MIYAMOTO H, L RASMUSSEN, E ZEUTHEN 1973 Studies of the effect of temperatureshocks on preparation for cell division in mouse fibroblast cells (L cells).J CellSci13: 889-900
29. PADILLA GM, IL CAMERON, A ZIMMERMAN, eds. 1974 Cell Cycle Controls.Academic Press, New York
30. PITTENDRIGH CS 1965 On the mechanism of the entrainment of a circadianrhythm by light cycles. InJ Aschoff, ed, Circadian Clocks. North-HollandPublishing Co, Amsterdam, pp 277-297
31. POLANSHEK MM 1977 Effects of heat shock and cycloheximide on growth anddivision of the fission yeast, Schizosaccharomyces pombe.J Cell Sci 23: 1-23
32. RASMUSSEN L, E ZEUTHEN 1962 Cell division and protein synthesis in Tetrahy-mena, as studied with p-fluorophenylalanine. C R Trav Lab Carlsberg 32: 333-358
33. SACHSENMAIER W 1976 Control of synchronous nuclear mitosis in Physarumpolycephalum. In JW Hastings, HG Schweiger, eds, The Molecular Basis ofCircadian Rhythms. Dahlem Konferenzen, Berlin, pp 409-420
34. SCHEFFEY C, JJ WILLE 1978 Cycloheximide-induced mitotic delay in Physarumpolycephalum. Exp Cell Res 113: 259-262
35. SCHNABEL G 1968 Der Einfluss von Licht auf die circadiane Rhythmik vonEuglena gracilis bei Autotrophie und Mixotrophie. Planta 81: 49-63
36. SMITH HTB,IM MITCHISON 1976 Anaesthetics delay and accelerate division inthe fission yeast Schizosaccharomyces pombe. Exp Cell Res 99: 432-435
37. SWEENEY BM 1982 Interaction of the cireadian cycle with the cell cycle inPyrocystisfusiformis. Plant Physiol. 70: 272-276
38. SWEENEY BM, JW HASTINGS 1958 Rhythmic cell division in populations ofGonyaulaxpolyedra. J Protozol 5: 217-224
39. WALz B, BM SWEENEY 1979 Kinetics of the cycloheximide-induced phasechanges in the biological clock in Gonyaulax. Proc Natl Acad Sci USA 76:6443-6447
40. WILLE JJ JR 1979 Biological rhythms in protozoa. In M Levandowsky, SHHutner, eds, Biochemistry and Physiology of Protozoa, Ed 2, Vol 2. AcademicPress, New York, pp 67-149
41. WINFREE AT 1980 The Geometry of Biological Time. Springer-Verlag, NewYork
42. ZEUTHEN E 1970 Independent synchronization of DNA synthesis and of celldivision in same culture of Tetrahymena cells. Exp Cell Res 61: 311-325
43. ZEUTHEN E 1971 Synchronization of the Tetrahymena cell cycle. In DM Prescott,L Goldstein, E McConkey, eds, Advances in Cell Biology, Vol 2. Appleton-Century-Crofts, New York, pp 111-152
44. ZEUTHEN E 1974 A cellular model for repetitive and free-running synchrony inTetrahymena and Schizosaccharomyces. In GM Padilla, IL Cameron, AMZimmerman, eds, Cell Cycle Controls. Academic Press, New York, pp 1-30
45. ZEUTHEN E, L RASMUSSEN 1972 Synchronized cell division in protozoa. In TTChen, ed, Research in Protozoology, Vol 4. Pergamon Press, New York, pp 9-145
302 EDMUNDS ET AL.
www.plantphysiol.orgon June 23, 2018 - Published by Downloaded from Copyright © 1982 American Society of Plant Biologists. All rights reserved.



















![A globally convergent incremental Newton method...Incremental Newton 3 extended Kalman lter (EKF) method with variable stepsize or equivalently the EKF-S algorithm [20], and shows](https://img.dokumen.tips/doc/110x75/607645be86b0bf621f2f18ba/a-globally-convergent-incremental-newton-method-incremental-newton-3-extended.jpg)





![course.ece.cmu.eduece796/documents/MPEG-1... · Web viewThe following table indicates the scale factor multiplier used in the requantization equation for each stepsize. scalefac_scale[gr]](https://img.dokumen.tips/doc/110x75/60a73c6f53206e0f46230324/ece796documentsmpeg-1-web-view-the-following-table-indicates-the-scale-factor.jpg)


