Embed Size (px)
Citation preview

1
Cdc28 Regulate Many Cell Cycle Events
How can one enzyme regulate multiple cell cycle events?
Synchronized cdc28ts cells can’t initiate S-phase
Cdc28 drives entry into S-phase as well as Mitosis
2
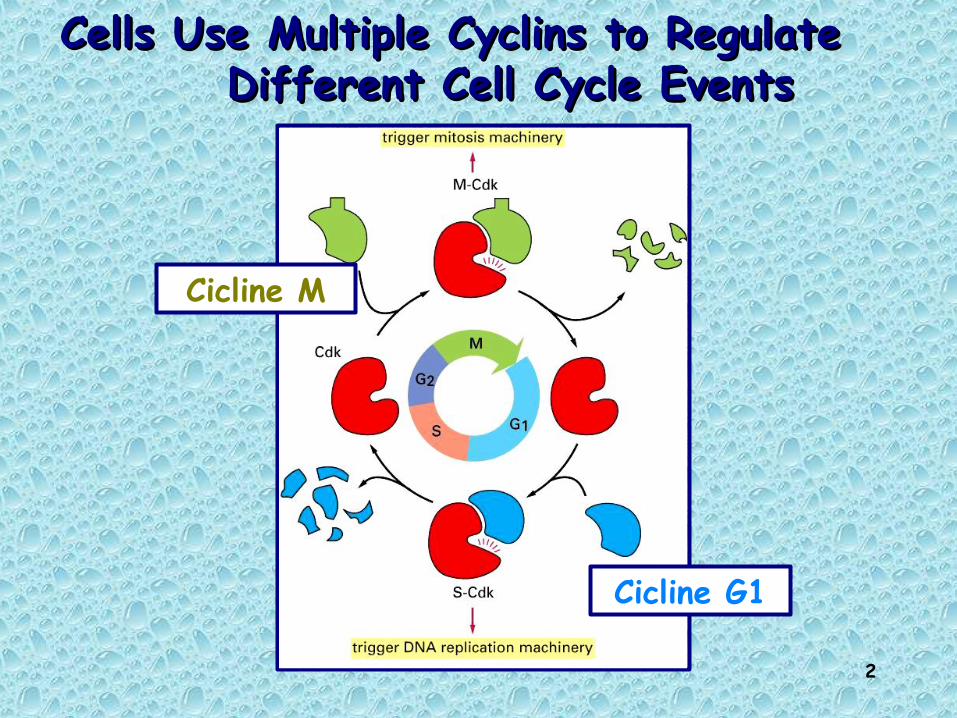
Cells Use Multiple Cyclins to Regulate Cells Use Multiple Cyclins to Regulate Different Cell Cycle EventsDifferent Cell Cycle Events
Cicline G1
Cicline M

3
Il complesso MPFMPF è formato da Cdc28/cicline mitotiche in S. cerevisiae e
Cdc2/Cdc13 in S. pombe
Il complesso SPFSPF è formato da Cdc28/cicline G1 in S. cerevisiae
Il complesso Il complesso MPFMPF regola l’ingresso mitotico regola l’ingresso mitotico (studi su (studi su S. pombeS. pombe))
Il complesso Il complesso SPFSPF regola l’ingresso in S regola l’ingresso in S (studi su (studi su S. cerevisiaeS. cerevisiae))

4
Modello di azione del complesso Cdc28/cicline Modello di azione del complesso Cdc28/cicline in in S. cerevisiaeS. cerevisiae

5
FunctionExpressionIdentificationCyclin
S-phase entryG1AccidentalClb6
S-phase entryG1AccidentalClb5
SpindleSHomologyClb4
SpindleSHomologyClb3
MitosisMHomologyClb2
MitosisMHomologyClb1
B-type Cyclins
STARTG1mutantCln3
STARTG1mutantCln2
STARTG1mutantCln1
G1 Cyclins
Budding Yeast Cyclin Genes

6
Cyclins NomenclatureCyclins Nomenclature
Cyclin ACyclin B’sS. pombe cdc13...S. cerevisiae CLB1,2,3,4,5,6
- G1’s more similar to each other-PEST domains (Pro,Glu,SerThr)
All contain “cyclin box” domain cdc2 binding domain
A and B’s closely related- destruction box- required for S and M- “A” accumulates earlier,- degraded earlier
G1 CyclinsS. cerevisiae CLN1,2,3

7
Cyclins can be grouped by expression patternsCyclins can be grouped by expression patterns
- CLN1 and CLN2 and CLB5 and CLB6 -
CLN1,2,CLB5,6
CLN3
CLB1,2
- CLB 1 and CLB2
- CLB3 and CLB4
CLB3,4
G1 Start Metaphase
[Cyc
lin]
[Cyc
lin]

Yeast Cyclin Genes: CLNs and CLBsYeast Cyclin Genes: CLNs and CLBs
Triple cln1,2,3 mutantTriple cln1,2,3 mutant
Quadruple clb1,2 mutantQuadruple clb1,2 mutant
Dominant CLN MutationsDominant CLN Mutations
CLN1,2,3CLN1,2,3Sense cell size, commit to divisionSense cell size, commit to division
Clb1,2Clb1,2Spindle assemblySpindle assembly
anaphaseanaphase
Clb5,6Clb5,6G1 entryG1 entry
Clb3,4Clb3,4Activate replication Activate replication originsorigins

9
Vertebrate Cells, like budding yeast, express multiple cyclins, but they also express several CDK’s

10
Regulation of G1/S transition (start/restriction point)
Identification of mammalian G1 cyclins
Human cDNA libraries screened for ability to complement CLN3 mutantlead to cloning of three human G1 cyclins:Cyclin CCyclin D1 Cyclin E
Cyclin D2 and Cyclin D3 subsequentlyidentified
D-type cyclins: respond to growth factors G0 to G1 transition
E-type cyclin: expression is periodic peaks at G1/S transition controls ability of mammalian cells to enter S-phase

11
Regulation of G1/S transition (start/restriction point)Regulation of G1/S transition (start/restriction point)
Identification of mammalian cyclin-dependent kinases (CDKs)Identification of mammalian cyclin-dependent kinases (CDKs)
Human cDNA libraries screened for ability to complement budding yeast CDC28 mutant
Three cDNA clones identified which could complement CDC28 mutant
Mammalian cdk1 acts at G2/M transition cdk2 acts at G1/S transition cdk3 unknown function
Cdk4 identified in an anti-cyclin D co-immunoprecipitation experiment
Unable to complement CDC28

12

13
Budding yeastBudding yeast MammiferiMammiferi
Punti di controllo del ciclo cellulare
START PUNTO DI RESTRIZIONE

14
Restriction point (START)
Point at which cell is irreversibly committed to traversing the cell cycleMammals: restriction pointYeast: START
Cell cycle proceeds without influence from environment (only stopped by damage)
Late in G1

15
To divide or not to divide:that is the question
Yeast cells make decision based on cell size, which is dependent on nutrient availability
Mammalian cells make decision based on the presence of protein growth factors called mitogens that stimulate cell growth

16
In assenza di fattori di crescita le cellule di mammifero si arrestano in G0 (cellule quiescenticellule quiescenti);
L’aggiunta di mitogeni causa il passaggio attraverso il punto di restrizione (dopo 14-16 oredopo 14-16 ore);
Le cellule in proliferazione entrano in S (dopo altre 6-8 oredopo altre 6-8 ore)

17
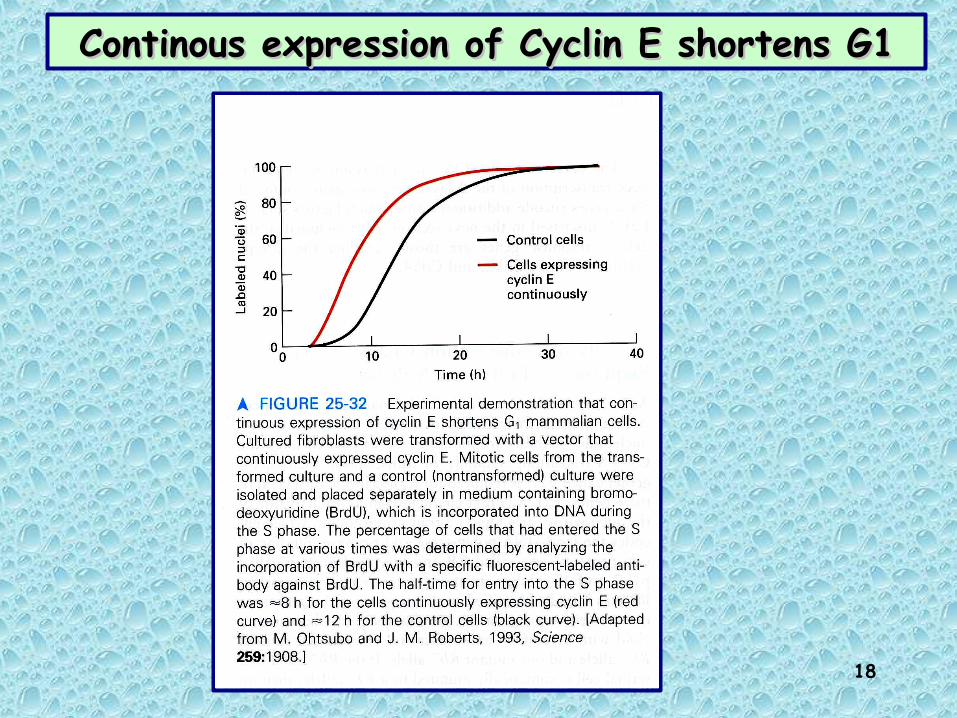
Cyclin D is required to pass restriction point
Colorazione 16 ore dopo la microiniezione e l’aggiunta di BrdU al mezzo.
Colorante per DNA
Anticorpi anti-BrdU
Anticorpi anti-cicl.D
R point
18
Continous expression of Cyclin E shortens G1Continous expression of Cyclin E shortens G1

19
Attività dei complessi Attività dei complessi Cdk/ciclina dei mammiferiCdk/ciclina dei mammiferi
All’inizio della fase S le cicline D ed E vengono degradate. I livelli di Cdk4 e 6 crollano repentinamente
Queste proteine non sono Queste proteine non sono necessarie per la progressione necessarie per la progressione
nella fase Snella fase SAll’inizio della fase S viene sintetizzata la ciclina A che si associa a Cdk2. La distruzione della ciclina A o una sua modificazione inibiscono la sintesi di DNA. Il complesso Cdk2/ciclina A è Il complesso Cdk2/ciclina A è
indispensabile per la indispensabile per la progressione nella fase Sprogressione nella fase S

20
Cell-cycle phase-specific Cell-cycle phase-specific Cdk complexesCdk complexes
Three classes:G1 Cdk complexes
S-phase Cdk complexesMitotic Cdk complexes (also known as MPF)
Cell-cycle phase specificity determined by cyclin type and, in some cells, Cdk type

21
Controllo del ciclo cellulare dei mammiferiControllo del ciclo cellulare dei mammiferi
Ingresso in fase S (complesso SPF)Ingresso in fase S (complesso SPF)
Progressione in fase SProgressione in fase S
Ingresso in fase M (complesso MPF)Ingresso in fase M (complesso MPF)Cdk1/ciclina ACdk1/ciclina B
Cdk4,6/ciclina D punto di restrizioneingresso in S
Cdk2/ciclina A
Cdk2/ciclina E

22
In addition to forming complexes between In addition to forming complexes between specific Cdks and their cyclin binding specific Cdks and their cyclin binding partners, all complexes require partners, all complexes require phosphorylation and dephosphorylation to phosphorylation and dephosphorylation to become fully activebecome fully active
phosphorylation of thr160/161 is phosphorylation of thr160/161 is important for the activation of all major important for the activation of all major Cdk/cyclin complexesCdk/cyclin complexes
catalysed by Cdk activating kinase CAKcatalysed by Cdk activating kinase CAK

23
Regolazione della Cdk1 mediante Regolazione della Cdk1 mediante fosforilazione-defosforilazionefosforilazione-defosforilazione
Equilibrio tra fosfatasi (Cdc25) e kinasi (Wee1)
Siti di inattivazione
Sito di attivazione

24
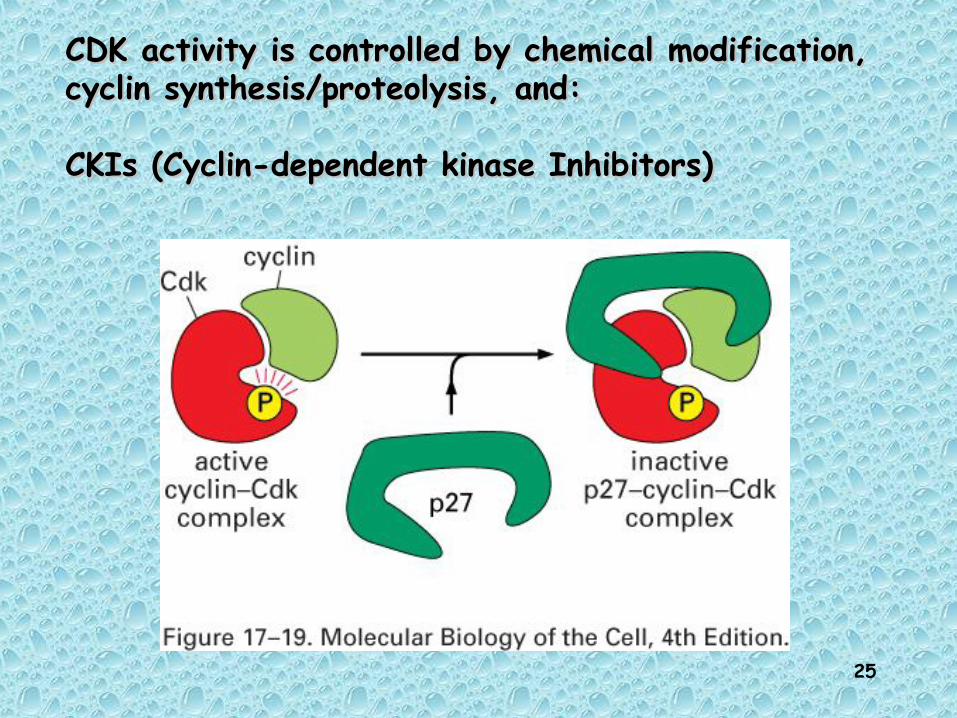
Another mechanism by which Another mechanism by which CDKs canCDKs can
regulate Multiple Transitions regulate Multiple Transitions is by usingis by using
different CDK inhibitors different CDK inhibitors (CKI’s)(CKI’s)
25
CDK activity is controlled by chemical modification, CDK activity is controlled by chemical modification, cyclin synthesis/proteolysis, and:cyclin synthesis/proteolysis, and:
CKIs (Cyclin-dependent kinase Inhibitors)CKIs (Cyclin-dependent kinase Inhibitors)
26
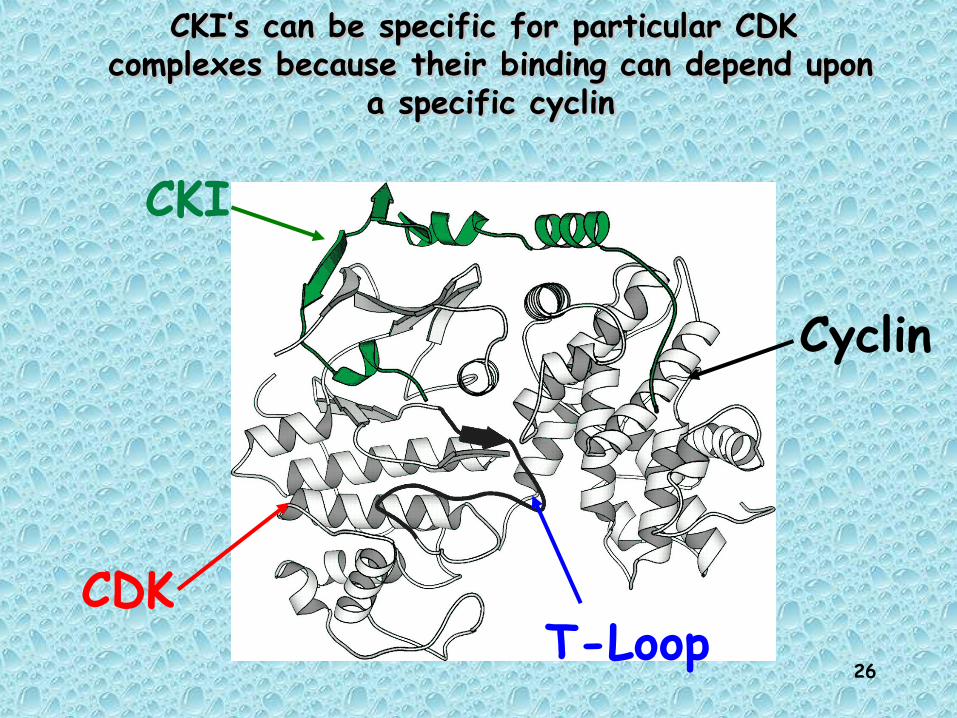
CDK
Cyclin
CKI
T-Loop
CKI’s can be specific for particular CDK CKI’s can be specific for particular CDK complexes because their binding can depend upon complexes because their binding can depend upon
a specific cyclina specific cyclin

27
Cdc28/Cln1,2Sic1
Down-regulation of a CKI can, in turn, be Down-regulation of a CKI can, in turn, be regulated by a CDK-mediated phosphorylationregulated by a CDK-mediated phosphorylationbecause its ubiquitin-dependent proteolysis because its ubiquitin-dependent proteolysis can be regulated by its phosphorylation statecan be regulated by its phosphorylation state

28
Wee1 kinases
Cdc25 phosphatases
Cdk-Activating Kinases (CAK)
Kap1 phosphatases
T160
Y15
PhosphorylationControl
Ubiquitin-dependent Proteolytic Control
Transcriptional Control
Cyclin-dependent KinaseInhibitors (CKI’s)
We can add another limb to our regulation treeWe can add another limb to our regulation tree
Transcriptional Control

29
Inibitori delle CdkInibitori delle CdkMembri diversi di questa famiglia inibiscono Cdk diverse
Cdc2/Clb P40
Cdc2/ClnFar1
Inibitore lievito
Cdk4/ciclinaDCdk6/ciclinaDCdk2/ciclinaECdk2/ciclinaA
p21, p27
Cdk4/ciclinaDCdk6/ciclinaD
p15, p16
Cyclin-Cdk complexInibitore cellule animali

30
Meccanismi di controllo dell’attività dei complessi ciclina/CDK
Concentrazione delle cicline -livello di sintesi tramite controllo trascrizionale-proteolisi-localizzazione cellulare
Fosforilazione delle CDK
Inibitori delle CDK

31
Proliferazione neoplastica: innesco della proliferazione in assenza di
programma, per mutazione di un gene cellulare o infezione di virus
Embriogenesi: proliferazione attivacicli cellulari brevirapida successione di fasi di replicazione (S) e mitosi (M) crescita "logaritmica".
Differenziamento: graduale attivazione di funzioni specializzate espansione clonale di cellule che si specializzano. contemporaneamente il ciclo di divisione viene modulato.
Differenziamento terminale: arresto programmato del ciclo (GO).
Reversibilità: cellule differenziate possono ripristinare il programma di divisione in risposta a stimoli (danni meccanici, agenti fisici etc.).
Situazioni biologiche in cui la proliferazione è controllata

32
2 principali livelli di controllo della proliferazione di cellule eucariotiche:
controllo della scelta tra destino proliferativo / entrata in quiescenza (G0) ed eventuale differenziamento della cellula.
REGOLATO DALL’ ESTERN0
controllo delle fasi del ciclo e coordinamento tra i diversi eventi necessari
alla divisione cellulare.MECCANISMI DI CONTROLLO INTRINSECI
Controllo del ciclo cellulare e controllo della proliferazione

33
A cell continues through the cell cycle after passing the restriction point (STARTSTART) unless it encounters genetic damage
If the cell receives a go-ahead signal, it completes the cell cycle and divides otherwise it switches to a nondividing state, the G0 phase. Most human cells are in this phase. Liver cells can be “called back” to the cell cycle by external cues (growth factors), but highly specialized nerve and muscle cells never divide.
Progress though the cell cycle is monitored at four checkpoints
Regulation of cell cycleRegulation of cell cycle

34
Cell cycle checkpointsCell cycle checkpoints ensure integrity of the genome
cell does not enter mitosis until DNA replication is complete and DNA damage is repaired
ensure chromosome segregation does not occur if chromosomes are incorrectly aligned on the mitotic spindle and spindle formation is inhibited
during the cell cycle a number of processes take place and they need to be co-ordinated
each process involves synthesis, assembly and correct function; all the changes that take place during the cell cycle have to integrated and correctly regulated

35
Look out for defect Look out for defect and emit a signal and emit a signal SENSORSSENSORS
Transmission of signals throughout Transmission of signals throughout the nucleus or cell and amplification the nucleus or cell and amplification TRANSDUCERSTRANSDUCERS
Delay cell-cycle progression Delay cell-cycle progression EFFECTORSEFFECTORS
Checkpoint machineryCheckpoint machinery

36
Four checkpointsFour checkpoints

37
THE THE G1/S CHECKPOINTG1/S CHECKPOINT

38
GROWTH FACTORS ARE INVOLVED GROWTH FACTORS ARE INVOLVED IN PASSING THE IN PASSING THE GG11 CHECKPOINT CHECKPOINT
Cyclin Cyclin
CyclinCyclin
Cyclin
Cyclin
CyclinCdK
CdK
CdK
ATP
ADP
Targetprotein
Pi
1. Arrival ofgrowth factorsfrom other cells.
2. Growth factors cause increase in cyclin concentration.
3. Cyclin activatescyclin-dependent kinase.
4. Kinases activate S phase proteins, leading to cell division.

39
Growth factor signalling and Growth factor signalling and transcriptiontranscription
Growth factors increase the expression of specific genes:
1. early response genes: rapid increase in mRNA and protein levels include transcription factors such as E2F, c-myc, c-
fos, c-jun
2. delayed response genes: include cell cycle proteins such as Cdks, cyclins expression regulated by early response genes

40
Regulated expression of two classes of Regulated expression of two classes of genes returns Ggenes returns G00 mammalian cells to mammalian cells to
the cell cyclethe cell cycle
Early-response genes:
Transcription factors (E2F)
Delayed-response genes:
CyclinD, ECdk2, 4, 6

41
Il passagggio attraverso il punto di Il passagggio attraverso il punto di restrizione dipende dalla attivazione di restrizione dipende dalla attivazione di
fattori di trascrizione E2Ffattori di trascrizione E2F
I fattori E2F attivi stimolano la propria I fattori E2F attivi stimolano la propria sintesi e quella della Cdk2 e della Ciclina E;sintesi e quella della Cdk2 e della Ciclina E;
Il complesso Cdk2/CyclE porta all’aumento di Il complesso Cdk2/CyclE porta all’aumento di fattori E2F attivi fattori E2F attivi

42
E2F is a transcription factor that by itself E2F is a transcription factor that by itself activates transcriptionactivates transcription
E2F
However, Rb binds to E2F and represses its activation function
Rb
gene expressiongene expression

43
When Rb repression is inhibited by phosphorylation of Rb by early G1 Cyclin-CDK protein kinases (Cyclin D-CDK4/6), E2F stimulates the expression of genes required for S-phase, including genes encoding DNA polymerases and other proteins required for DNA synthesis, enzymes that synthesize dNTPs, and genes encoding late G1 Cyclin-CDKs (Cyclin E and CDK2) and the major S-phase Cyclin (Cyclin A).
Rb repression is regulated by Rb repression is regulated by early Gearly G11 Cyclins-CDKs Cyclins-CDKs
PP P P
E2FE2F
RbRbS-phase gene transcription

44
The G1/S transition in human cells: Rb (retinoblastoma protein) regulates E2F proteins, which are transcription
factors for S phase genes

45
Tumor-suppressor genes Tumor-suppressor genes Genes capable of suppressing the tumor-
forming potential of transformed cellsTumor suppressor genes normally function
to suppress cell growth and division.Tumor-suppressor genes encode :
Regulatory proteins eg rbrb, p53p53Intra-cellular signaling proteins (nf1-
Neurofibrosarcomas)Cell adhesion proteins (dcc–colon carcinomas)

46
Cells become malignant by losing tumor suppressor gene activity.
These act as Mendelian recessive traits. A cell has to be homozygous for a nonfunctional (or missing) tumor suppressor gene for it to have an effect.
AnalogyTumor suppressor brake pedalLack of suppressor no brake pedal
Tumor-suppressor genes Tumor-suppressor genes

47
Gene oncosoppressore RbGene oncosoppressore Rb• Raro tumore dell’occhio che
colpisce circa un bambino su 14000 nati
• Provocato da due mutazioni consecutive che interessano entrambe le copie del gene Rb

48
What is the defect in hereditary retinoblastoma?
Rb Rb Rb
Normal person:2 good copies ofRb gene
Hereditary retinoblastomapatient: 1 good copy ofRb gene, 1 defective copy
Compromise this good one, then problems!
or
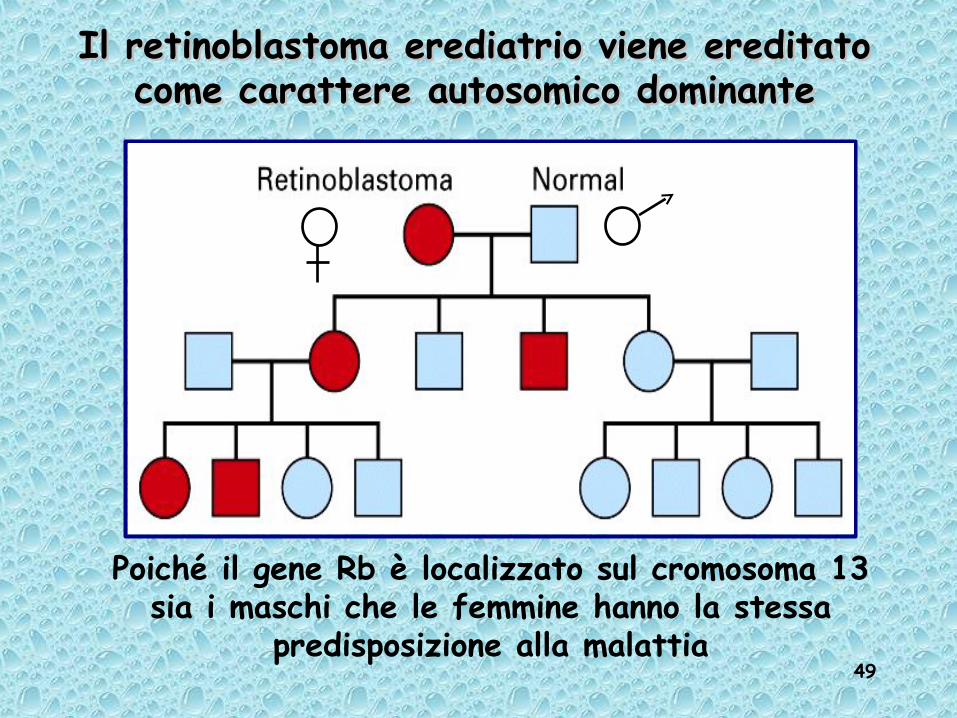
49
Il retinoblastoma erediatrio viene ereditato Il retinoblastoma erediatrio viene ereditato come carattere autosomico dominantecome carattere autosomico dominante
Poiché il gene Rb è localizzato sul cromosoma 13 sia i maschi che le femmine hanno la stessa
predisposizione alla malattia

50
Perdita dell’eterozigosi degli antioncogeniPerdita dell’eterozigosi degli antioncogeni

51
Meccanismi genetici alla base del retinoblastoma

52
Retinoblastoma and the “Two-hit” Retinoblastoma and the “Two-hit” model of carcinogenesismodel of carcinogenesis
Knudsons “two-hit” hypothesis:familial cases (high frequency, early onset): retinoblastoma caused by a germline mutation of one Rb allele + an acquired somatic mutation of the remaining allele of the Rb gene = both inactivatedsporadic cases (low frequency, late onset): retinoblastom caused by two acquired somatic mutations in both alleles = both inactivated

53
Knudson’s 2-hit mutation model for retinoblastoma

54
RB = tumor suppressor geneRB was the first tumor
suppressor to be identified.
RB is absent or mutated in at least one-third of all human tumors.
Cloning of the retinoblastoma gene mapped to 13q14 (loss of heterozygosity) rb-1 gene cloned 1986-87 Mutated or lost in all cases of retinoblastomas Also found mutated in osteosarcoma and small-cell lung cancer

55
Il gene Rb-1Il gene Rb-1GeneticaFrequenza del tumore: 1/14.000 nasciteLe cellule tumorali sono prive di entrambi gli alleli Rb-1 funzionaliAssetto più frequente: una piccola delezione trasmessa dalla linea germinale, poi una mutazione di senso sull’allele omologo dà origine a un clone somatico Associazione di molti casi di tumore con la delezione della banda 13q1.4
Trasmissione mendeliana semplice: predisposizione allo sviluppo di tumori dominante: un cromosoma "difettivo" predispone tutti i portatori a sviluppare tumori;sviluppo di tumori recessivo: il tumore si sviluppa in completa assenza del prodotto, quando entrambi gli alleli sono deleti e/o mutati
Studi in sistemi modello trasfezione del gene Rb1 in cellule proliferanti, microiniezione di proteina pRb, inibiscono il superamento della transizioneG1/S
Topo Rb-/- : letale (difetti nell’eritropoiesi e nei tessuti neuronali)

56
RB - structure of gene and proteinRB - structure of gene and protein Gene
– Highly complex: 200 kb with 27 exons and introns from 80bp to 60kb
Protein– multiple bands Mw= 110-116 kDa– nuclear phosphoprotein– binds DNA non-specifically
Rb contains several functional domains– Domains A and B are highly conserved from humans to
plants, and they interact with each other along an extended interdomain interface to form the central “pocket”, which is critical to the tumour suppressor function of Rb
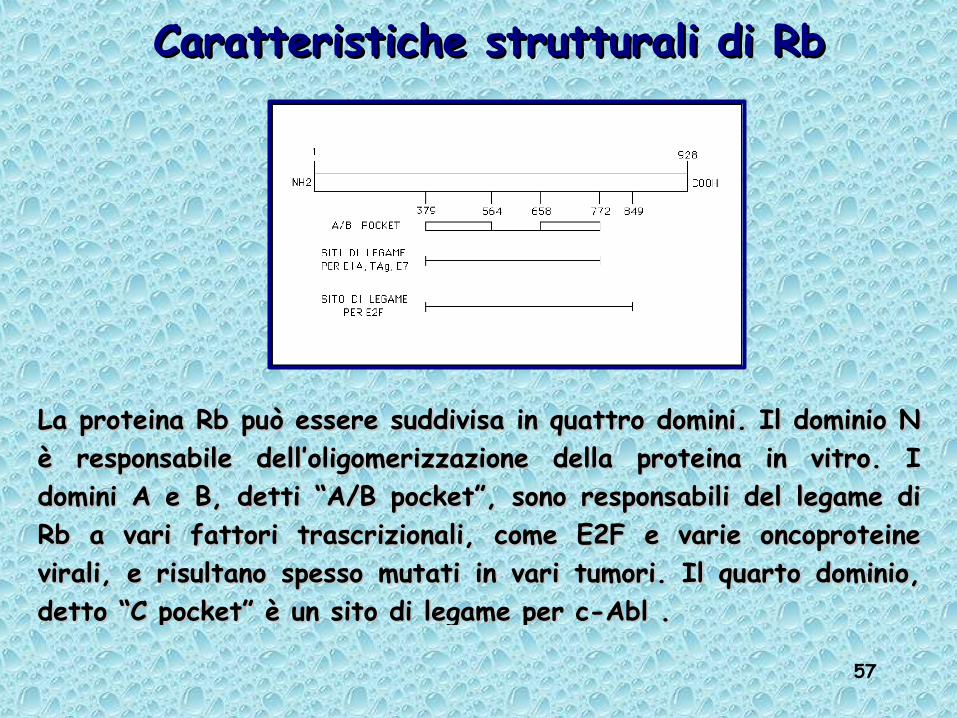
57
La proteina Rb può essere suddivisa in quattro domini. Il dominio N La proteina Rb può essere suddivisa in quattro domini. Il dominio N è responsabile dell’oligomerizzazione della proteina in vitro. I è responsabile dell’oligomerizzazione della proteina in vitro. I domini A e B, detti “A/B pocket”, sono responsabili del legame di domini A e B, detti “A/B pocket”, sono responsabili del legame di Rb a vari fattori trascrizionali, come E2F e varie oncoproteine Rb a vari fattori trascrizionali, come E2F e varie oncoproteine virali, e risultano spesso mutati in vari tumori. Il quarto dominio, virali, e risultano spesso mutati in vari tumori. Il quarto dominio, detto “C pocket” è un sito di legame per c-Abl .detto “C pocket” è un sito di legame per c-Abl .
Caratteristiche strutturali di RbCaratteristiche strutturali di Rb

58
RB FAMILYRB FAMILY p105 RB; p107; p130 pRB binds to the transcriptional activation
domain of E2F and blocks activation E2Fs transactivate expression of genes
that are important for S phase:
dihydrofolate reductasedihydrofolate reductasethymidine kinasethymidine kinasepolymerase polymerase histoneshistones

59
RB´s function: “a signal transducer connecting the cell cycle clock with the
transcriptional machinery”
RB constitutively expressed and relatively stable half-life ≥ 12 hours Still induced increase in levels
resting G0 cells + mitogenic stimuli = RB level increased 4-6x
RB modified by phosphorylation during cell cycle
RbMM
G1G1G2G2
SS
Cell cycle clock
Transcriptional apparatus

60
Rb a substrate of CycD-CDKCycE-CDKCycA-CDK
CycC-Cdk3pRb
G0

61
RB is active only within a limited time RB is active only within a limited time window during the cell cyclewindow during the cell cycle
Before the R-point in G1: Rb hypophosphorylated = active repressor of growth (inhibits cell cycle progression) SDS-PAGE: 110 kDa
After the R-point in G1: Rb hyperphosphorylated = inactive repressor of growth (facilitates cell cycle progression) SDS-PAGE: 112 - 116 kDa
Rb is dephosphorylated at the end of mitosis
R
Rb
RbP P P PP
P
activerepressor
Inactiverepressor
MM
G1G1G2G2
SS
P

62
Gate-keeper model for RBGate-keeper model for RB The R-point functions as a door that is kept closed by Rb
G1 arrest upon overexpression of Rb Under conditions favourable for proliferation ⇒ Rb phosphorylated
⇒ R-door is opened In cells with lost Rb-function the door is left open all the time
Such cells will also have lost the ability to respond to growth-promoting/-inhibitory signalsMitogenes (+), TGFβ (-), contact-inhibition (-)
Two key elements in this model: upstream signals ⇒ Rb´s phosphorylation status Rb´s phosphorylations status ⇒ downstream effects
Rb as “signal transducer” Cell cycle-clock ⇒ RB´s phosphorylation status RB´s phosphorylation status ⇒ transcription apparatus involved
in proliferation

63
M
G1G2
S RRb
Cdk4/6Cyclin D
E2F releasedS-phase genes expressed
Gate keeper modelGate keeper model
PPPP
PPPP
PP
PP
PP

64
E2F liberated by Rb inactivationE2F liberated by Rb inactivation
• Rb excert its effects through E2F TFs
Rb = active repressor
Rb = inactivated
R-point
E2F = activated!

65
RB´s phosphorylation status RB´s phosphorylation status ⇒⇒ a signal to the transcription apparatus a signal to the transcription apparatus
• Hypophosphorylated RB binds and inactivates the transcription factor E2F/DP
• Hyperphosphorylation of RB ➨ E2F/DP liberated and free to activate genes necessary for proliferation

66
The functional state of the retinoblastoma protein The functional state of the retinoblastoma protein (pRb) controls cell proliferation(pRb) controls cell proliferation
GFs, mitogens...
cyclin D1 synthesis
Cell cycle progression
G1
RNA pol II
5’..TTTCCGCG…3’
cyclin D1
cdk4/6
Active cyclin/kinasecomplex
Cell cycle genes off
G0
pRb
E2F/DPdimers
5’..TTTCCGCG…3’
wt pRb represses transcription of cell cycle genes
Phosho-pRb becomes inactive
P
P

67
Target genes controlled by Target genes controlled by activating E2Fsactivating E2Fs
E2F sites common consensus binding site: TTTCGCGC No difference in sequence preference between different
E2Fs target genes: E2F controls the transcription of cellular genes
that are essential for cell division: cell cycle regulators
such as cyclin E, cyclin A, Cdc2, Cdc25A, RB and E2F1, enzymes that are involved in nucleotide biosynthesis
such as dihydrofolate reductase, thymidylate synthetase and thymidine kinase

68
E2F/DP only active in a window E2F/DP only active in a window of the cell cycle (late G1 of the cell cycle (late G1 ➜➜ early S) early S)
• Early G1: active RB ➜ E2F/DP turned OFF• The R-point: inactivated RB ➜ E2F/DP turned ON
– E2F/DP liberated ➜ activation of E2F-dependent promoters
• Late S: E2F/DP turned OFF again– cyclin A/cdk2 ➜ phosphorylation of E2F/DP ➜ reduced
DNA-binding ➜ target genes turned off

69
HOW ARE S PHASE PROTEINS ACTIVATED?HOW ARE S PHASE PROTEINS ACTIVATED?
Rb
CyclinCdK
Rb
E2F
Rb
ATP
ADPPi
Pi
E2FE2FE2F
S Phaseproteins
mRNA
DNA
1. In normal cells,Rb protein binds to E2F and shutsdown the cell cycle.
2. If growth factors arrive and activate the cyclin-CdK complex, Rb becomes phosphorylated.
3. E2F is released
4. E2F stimulates the
production of S phase
proteins.
When Rb is mutated, no “tie-up” of E2F, so constant S phase turn on

70
Effetti delle mutazioni del Gene RbEffetti delle mutazioni del Gene Rb
OsteosarcomiOsteosarcomiCarcinomi: Carcinomi: 1) Polmonari 1) Polmonari 2) Mammari2) Mammari3) Prostatici3) Prostatici












![Soils and water [Chapter 3]...34 Soils and the Water Cycle Forest and rangeland soils regulate many important pro-cesses within the water cycle (Fig. 3.1). Not only does the soil strongly](https://img.dokumen.tips/doc/110x75/60c28a8cca00ac21f918578d/soils-and-water-chapter-3-34-soils-and-the-water-cycle-forest-and-rangeland.jpg)
















