Embed Size (px)
Citation preview

Caractérisation des interactions phage-minerai et
développement de bio-réactifs potentiels pour les
procédés de flottation
Mémoire
Karl Tremblay-Bouliane
Maîtrise en génie chimique
Maître ès sciences (M.Sc.)
Québec, Canada
© Karl Tremblay-Bouliane, 2015


iii
RÉSUMÉ
La flottation est un procédé de séparation important dans l’industrie minière. Un in-
térêt particulier est porté aux réactifs employés dans ce procédé en raison de leur impact sur
les performances économiques des mines. Avec pour objectif de développer des réactifs
efficaces d’origine biologique et moins nocifs pour l’environnement, une librairie
d’expression phagique a été criblée afin d’identifier des ligands pour différents minerais
d’intérêt industriel, notamment l'or, la chalcopyrite, la sphalérite, la pyrite et la silice. Après
plusieurs rondes de sélection et d'enrichissement, des séquences peptidiques ont été isolées
pour chacun de ces minerais. Toutefois, la détermination des isothermes d'adsorption pour
chacune de ces interactions a révélé une faible spécificité. L’effet de l’adsorption de bacté-
riophages, arborant certaines séquences, à la surface de ces minerais a été étudié afin
d’évaluer leur potentiel en tant que bio-réactifs de flottation. Ceci a permis de confirmer
une diminution de l’hydrophobicité, c’est-à-dire un effet déprimant. Les bactériophages
seraient donc des candidats potentiels pour certains types d'application en flottation, cepen-
dant leur coût de production devrait être significativement réduit afin d'en faire une alterna-
tive viable.


v
SUMMARY
Flotation is an important separation process in mining industry. Because of their
impact on mines economic performances, special attention is directed to flotation reagents.
In order to develop efficient bio-based reagents with a lower environmental footprint, a
phage display library was screened as a mean to identify peptides able to bind to ores of
economical interest, including gold, chalcopyrite, pyrite, sphalerite and silica. After many
biopanning rounds, peptide sequences were successfully isolated for each of these ores.
However, adsorption isotherm determination for these interactions revealed a low specifici-
ty of the obtained sequences. The possibility to use bacteriophages as flotation bio-reagents
was assessed by studying effect produced by adsorption of phages displaying selected pep-
tide sequences on surface properties of some of the ores. A decrease in hydrophobicity was
confirmed, suggesting a depressing effect on ores. Thus, bacteriophages might be potential
candidates for some types of flotation applications but their production cost will have to be
brought down significantly in order to be considered a viable alternative.


vii
TABLE DES MATIÈRES
Résumé ............................................................................................................................. iii
Summary ........................................................................................................................... v
Liste des tableaux ............................................................................................................. ix
Liste des figures ............................................................................................................... xi
Remerciements ............................................................................................................... xiii
Avant-propos .................................................................................................................. xv
Chapitre 1 : Introduction et objectifs ................................................................................ 1
1.1. Mise en contexte et problématique ....................................................................... 3
1.2. Procédés de flottation ............................................................................................ 5
1.2.1. Concept général........................................................................................... 5
1.2.2. Réactifs de flottation ................................................................................... 6
1.2.3. Flottation des sulfures ................................................................................. 8
1.3. Revue de littérature : Interactions peptides/minerais et bio-réactifs de flottation 9
1.3.1. Bactéries ...................................................................................................... 9
1.3.2. Polymères d’origine naturelle ................................................................... 10
1.3.3. Peptides ..................................................................................................... 12
1.4. Objectifs .............................................................................................................. 19
Chapitre 2 : Matériel et méthodes ................................................................................... 21
2.1. Caractéristiques et traitement des phases minérales ........................................... 23
2.1.1. Caractéristiques des minerais .................................................................... 23
2.1.2. Traitement à l’EDTA ................................................................................ 23
2.2. Criblage de la librairie de phages ........................................................................ 24
2.2.1. Choix de la librairie................................................................................... 24
2.2.2. Procédures générales reliées au bio-criblage ............................................ 26
2.3. Caractérisation de l’adsorption ........................................................................... 32
2.3.1. Modèle d’interaction récepteur/ligand ...................................................... 33
2.3.2. Génération des isothermes et calcul des paramètres ................................. 33
2.4. Mesure de temps d’induction .............................................................................. 34
2.4.1. Concept de temps d’induction................................................................... 34
2.4.2. Conditions expérimentales ........................................................................ 35

viii
Chapitre 3: Résultats présentés sous forme d'article ....................................................... 37
3.1. Résumé en français .............................................................................................. 39
3.2. Abstract ................................................................................................................ 40
3.3. Introduction ......................................................................................................... 41
3.4. Materials and Methods ........................................................................................ 42
3.4.1. Phage library and biopanning .................................................................... 42
3.4.2. Adsorption isotherms................................................................................. 43
3.4.3. Induction time measurements .................................................................... 44
3.5. Results and discussion ......................................................................................... 45
3.5.1. Biopanning ................................................................................................ 45
3.5.2. Adsorption isotherms................................................................................. 47
3.5.3. Induction time measurements .................................................................... 51
3.6. Conclusion ........................................................................................................... 54
Chapitre 4: Résultats et éléments de discussion supplémentaires ................................... 55
4.1. Expériences de bio-criblage supplémentaires ..................................................... 57
4.2. Efficacité de la titration des phages ..................................................................... 61
4.3. Calcul du coût potentiel de l'utilisation de phages comme réactif de flottation .. 63
Chapitre 5 : Conclusion et recommandations ................................................................. 65
Références ....................................................................................................................... 69
ANNEXES ...................................................................................................................... 75
Annexe 1 : Milieux et solutions .................................................................................. 77
Annexe 2 : Protocoles détaillés .................................................................................. 79
Annexe 3 : Résultats détaillés du bio-criblage ........................................................... 83
Annexe 4: Abréviations et catégories d’acides aminés .............................................. 89

ix
LISTE DES TABLEAUX
Tableau 1: Types de minerai traités par flottation et concentrés produits aux États-Unis, en
millions de tonnes métriques ............................................................................... 4
Tableau 2: Adhésion d'acides aminés sur différentes surfaces inorganiques avec de l'eau
comme solvant .................................................................................................. 13
Table 3: Biopanning results ................................................................................................. 46
Table 4: Results found in literature for biopanning on ores ................................................ 47
Tableau 5: Séquences obtenues lors du bio-criblage avec pré-incubation .......................... 58
Tableau 6: Efficacité de sélection de différents groupes de clones ..................................... 59
Tableau 7: Concentrations types récupérées lors de l'élution et de l'amplification ............. 60
Tableau 8: Coût du milieu de culture pour la production de bactériophages ...................... 64
Tableau 9: Détails à propos de la préparation des milieux et des solutions ........................ 77
Tableau 10: Acides aminés et abréviations ......................................................................... 89
Tableau 11: Catégories et structure d'acides aminés ........................................................... 89


xi
LISTE DES FIGURES
Figure 1: Croissance économique de l'industrie minière canadienne de 1983 à 2014,
excluant les combustibles ....................................................................................... 3
Figure 2: Schéma conceptuel d’une cellule mécanique majoritairement employée pour les
procédés de flottation ............................................................................................. 6
Figure 3: Représentation du xanthate d'éthyle de potassium, largement utilisé pour la
flottation des sulfures ............................................................................................. 7
Figure 4: Adhésion de R. opacus sur l'hématite et le quartz en fonction du pH .................. 10
Figure 5: Flottation de mélanges de chalcopyrite et de galène selon le pH, avec ajout de
chitosane ............................................................................................................... 11
Figure 6: Récupération de la sphalérite et de la galène par flottation en fonction de
différentes concentrations d'ADN ........................................................................ 12
Figure 7: Structure du bactériophage M13 .......................................................................... 15
Figure 8: Isothermes d'adsorption démontrant l'affinité des peptides obtenus suite au
criblage d'une librairie de phages sur du carbone de type diamant ...................... 16
Figure 9: Représentation de l'adsorption de bactériophages sur la silice et la sphalérite par
l'entremise de mesure de fluorescence ................................................................. 16
Figure 10: Comparaison de l'adsorption de bactériophages décorés de séquences
octapeptidiques différentes sur la chalcopyrite, la sphalérite, la silice et la pyrite
............................................................................................................................ 17
Figure 11: Temps d'induction pour la chalcopyrite en présence et en absence de phages
adsorbés .............................................................................................................. 18
Figure 12: Agrégation de fines particules de chalcopyrite induite par l'addition de
bactériophages .................................................................................................... 18
Figure 13: Séquences insérées pour la fabrication de librairies peptidiques (Ph.DMD, NEB)
sur le bactériophage M13 ................................................................................... 25
Figure 14: Représentation des différentes étapes du bio-criblage ....................................... 28
Figure 15: Exemple d'essai sur plaque ................................................................................ 29
Figure 16: Appareil de mesure de temps d’induction (induction timer) ............................. 35
Figure 17: Isotherms comparing adsorption of different phage clones on chalcopyrite and
sphalerite. ........................................................................................................... 48
Figure 18: Estimated parameters for receptor-ligand interaction model. ............................ 50
Figure 19: Induction time measurements of chalcopyrite, sphalerite and pyrite with and
without (control) addition of phages at different concentrations. ...................... 53
Figure 20: Calcul de la représentation des différents groupes et de chacun des différents
clones présents dans chaque groupe suite aux différentes rondes du bio-criblage.
............................................................................................................................ 60

xii
Figure 21: Vérification de l'efficacité de mise en culture. Comparaison du titre de
différents stocks de phages par la méthode d’essai sur plaque et la méthode
spectrophotométrique. ....................................................................................... 62
Figure 22: Production de bactériophages en bioréacteur de 20 L. ...................................... 63

xiii
REMERCIEMENTS
Pour son aide dans la réalisation de ce travail et dans cette introduction au monde de
la recherche, je voudrais remercier mon directeur de recherche au département de génie
chimique de l’Université Laval, M. Alain Garnier. J’aimerais également mentionner l’aide
apportée par M. Bruno Gaillet, co-directeur, ainsi que celle apportée par les membres des
équipes de recherche que j’ai côtoyés, qui ont contribué à une ambiance de travail enrichis-
sante et constructive.
Je tiens également à souligner la contribution très importante de Caroline Olsen,
superviseure du projet dans le milieu industriel d’accueil (COREM). Son expérience en
recherche et ses connaissances en traitement du minerai ont été d’une grande aide dans ce
projet et pour la rédaction de ce mémoire. Merci également à tout le personnel de recherche
et le personnel technique de COREM avec qui j’ai eu l’occasion de travailler et sans qui la
réalisation de ce travail aurait été impossible.
Enfin, j’aimerais remercier M. Sylvain Moineau, Mme Denise Tremblay et leur
équipe au département de biochimie, microbiologie et bio-informatique de l’Université
Laval pour leur aide et générosité, particulièrement en ce qui a trait à mon initiation à la
culture et la caractérisation des bactériophages.


xv
AVANT-PROPOS
Dans le cadre des travaux reliés à ce programme de maîtrise, un article a été rédigé
et inséré dans le présent mémoire. Cet article n’a toutefois pas encore été soumis ni publié.
Au moment de la rédaction de ce mémoire, des démarches de révision sont entamées et
certains périodiques scientifiques sont considérés pour la publication. D’autre part, des pré-
sentations ont été faites lors de diverses conférences scientifiques
1. Insertion d’article
Titre de l’article:
Screening a Phage Library Looking for Peptide-Ore Interactions for Bio-Flotation Applica-
tion
Auteurs:
Karl Tremblay-Bouliane1, Caroline Olsen3, Denise Tremblay2, Sylvain Moineau2 et Alain
Garnier1,*
1Université Laval, Department of Chemical Engineering, Québec, QC, Canada
2Université Laval, Department of Microbiology and Bio-informatic, Québec, QC, Canada
3COREM, Research Group in Flotation and Extractive Metallurgy, Québec, QC, Canada
*Auteur de correspondance: [email protected]
Auteur principal:
Karl Tremblay-Bouliane
Co-auteurs:
Caroline Olsen, Denise Tremblay, Sylvain Moineau, Bruno Gaillet et Alain Garnier
Périodique scientifique considéré :
CIM magazine (Canadian Institute of Mining, Metallurgy and Petroleum)
Rôle de l’auteur principal:
- Réalisation des expériences
- Analyse des résultats
- Rédaction de la première version de l’article (révision et contribution par les co-
auteurs)

xvi
2. Présentations lors de conférences
15e Rencontre Québec-Ontario en biotechnologie
Date : 28/05/2013
Type de présentation : Présentation par affiche
Titre : Screening a Phage Library Looking for Peptide-Ore Interactions
Auteurs : Karl Tremblay-Bouliane1, Caroline Olsen3, Denise Tremblay2, Sylvain Moineau2
et Alain Garnier1,*
63e Congrès canadien de génie chimique
Date : 22/10/2013
Type de présentation : Présentation orale
Titre : Peptide-Ore Interaction Screening by use of Phage Display Combinatorial Library
Auteurs : Karl Tremblay-Bouliane1, Caroline Olsen3, Denise Tremblay2, Sylvain Moineau2
et Alain Garnier1,*
14e Symposium annuel de PROTEO
Date : 09/05/2014
Type de présentation : Présentation par affiche
Titre : Recherche et caractérisation d’interactions peptide-minerai par l’entremise du cri-
blage d’une librairie de phages dans une optique de bio-flottation
Auteurs : Karl Tremblay-Bouliane1, Caroline Olsen3, Denise Tremblay2, Sylvain Moineau2
et Alain Garnier1,*
1Université Laval, Département de génie chimique, Québec, QC, Canada
2Université Laval, Département de microbiologie et bio-informatique, Québec, QC, Canada
3COREM, Groupe de recherché en flottation et métallurgie extractive, Québec, QC, Canada
*Auteur de correspondance: [email protected]

1
CHAPITRE 1 : INTRODUCTION ET OBJECTIFS

3
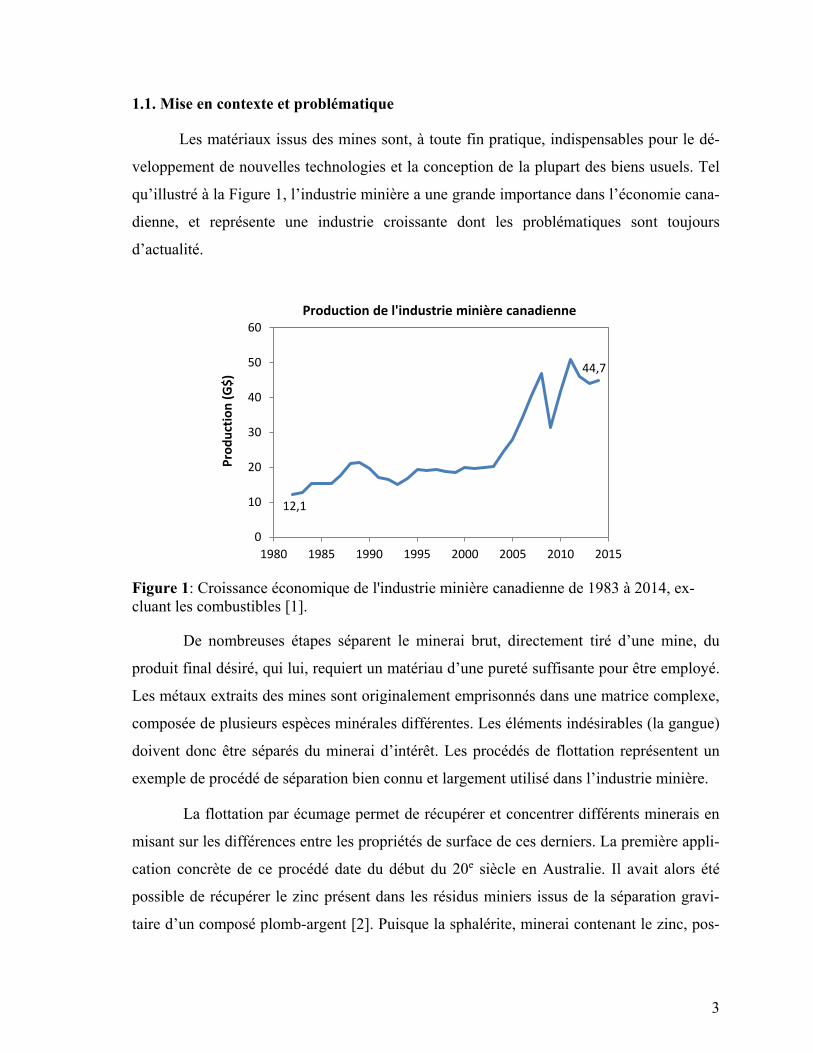
1.1. Mise en contexte et problématique
Les matériaux issus des mines sont, à toute fin pratique, indispensables pour le dé-
veloppement de nouvelles technologies et la conception de la plupart des biens usuels. Tel
qu’illustré à la Figure 1, l’industrie minière a une grande importance dans l’économie cana-
dienne, et représente une industrie croissante dont les problématiques sont toujours
d’actualité.
Figure 1: Croissance économique de l'industrie minière canadienne de 1983 à 2014, ex-
cluant les combustibles [1].
De nombreuses étapes séparent le minerai brut, directement tiré d’une mine, du
produit final désiré, qui lui, requiert un matériau d’une pureté suffisante pour être employé.
Les métaux extraits des mines sont originalement emprisonnés dans une matrice complexe,
composée de plusieurs espèces minérales différentes. Les éléments indésirables (la gangue)
doivent donc être séparés du minerai d’intérêt. Les procédés de flottation représentent un
exemple de procédé de séparation bien connu et largement utilisé dans l’industrie minière.
La flottation par écumage permet de récupérer et concentrer différents minerais en
misant sur les différences entre les propriétés de surface de ces derniers. La première appli-
cation concrète de ce procédé date du début du 20e siècle en Australie. Il avait alors été
possible de récupérer le zinc présent dans les résidus miniers issus de la séparation gravi-
taire d’un composé plomb-argent [2]. Puisque la sphalérite, minerai contenant le zinc, pos-
44,7
12,1
0
10
20
30
40
50
60
1980 1985 1990 1995 2000 2005 2010 2015
Pro
du
ctio
n (
G$
)
Production de l'industrie minière canadienne
4
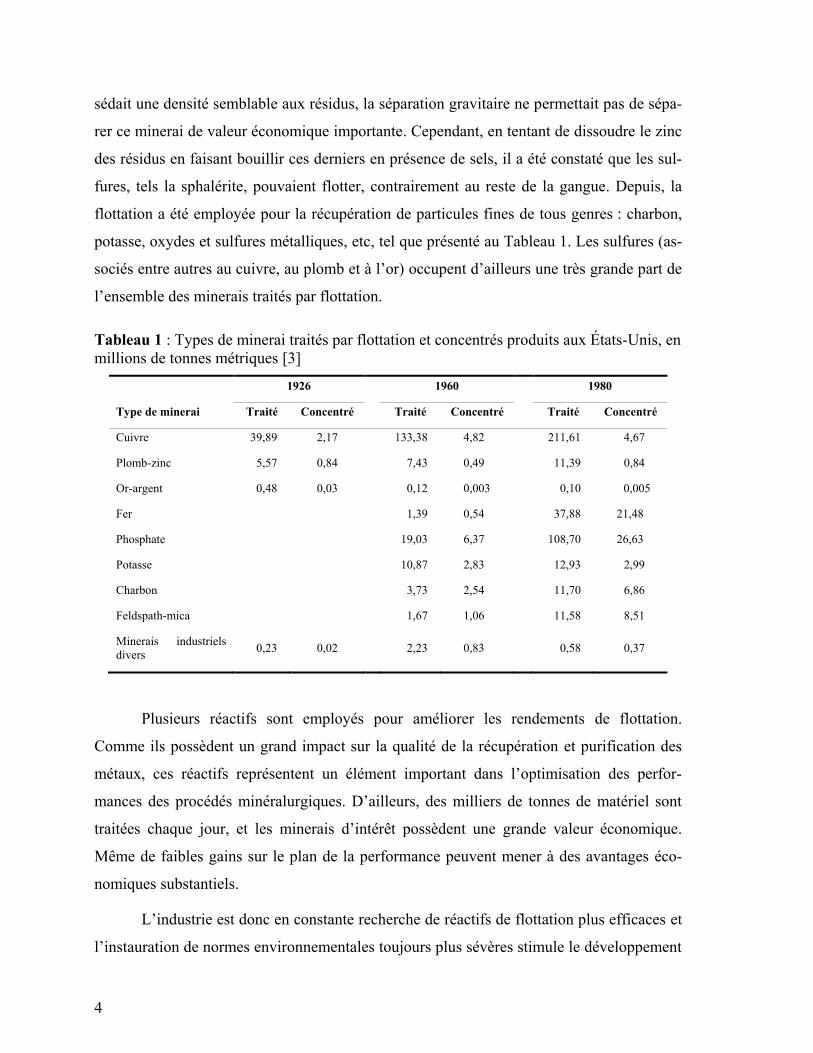
sédait une densité semblable aux résidus, la séparation gravitaire ne permettait pas de sépa-
rer ce minerai de valeur économique importante. Cependant, en tentant de dissoudre le zinc
des résidus en faisant bouillir ces derniers en présence de sels, il a été constaté que les sul-
fures, tels la sphalérite, pouvaient flotter, contrairement au reste de la gangue. Depuis, la
flottation a été employée pour la récupération de particules fines de tous genres : charbon,
potasse, oxydes et sulfures métalliques, etc, tel que présenté au Tableau 1. Les sulfures (as-
sociés entre autres au cuivre, au plomb et à l’or) occupent d’ailleurs une très grande part de
l’ensemble des minerais traités par flottation.
Tableau 1 : Types de minerai traités par flottation et concentrés produits aux États-Unis, en
millions de tonnes métriques [3]
1926 1960 1980
Type de minerai Traité Concentré Traité Concentré Traité Concentré
Cuivre 39,89 2,17 133,38 4,82 211,61 4,67
Plomb-zinc 5,57 0,84 7,43 0,49 11,39 0,84
Or-argent 0,48 0,03 0,12 0,003 0,10 0,005
Fer 1,39 0,54 37,88 21,48
Phosphate 19,03 6,37 108,70 26,63
Potasse 10,87 2,83 12,93 2,99
Charbon 3,73 2,54 11,70 6,86
Feldspath-mica 1,67 1,06 11,58 8,51
Minerais industriels
divers 0,23 0,02 2,23 0,83 0,58 0,37
Plusieurs réactifs sont employés pour améliorer les rendements de flottation.
Comme ils possèdent un grand impact sur la qualité de la récupération et purification des
métaux, ces réactifs représentent un élément important dans l’optimisation des perfor-
mances des procédés minéralurgiques. D’ailleurs, des milliers de tonnes de matériel sont
traitées chaque jour, et les minerais d’intérêt possèdent une grande valeur économique.
Même de faibles gains sur le plan de la performance peuvent mener à des avantages éco-
nomiques substantiels.
L’industrie est donc en constante recherche de réactifs de flottation plus efficaces et
l’instauration de normes environnementales toujours plus sévères stimule le développement

5
de réactifs plus verts. En plus de l’aspect environnemental, les réactifs traditionnels peuvent
représenter un souci important sur le plan de la santé et de la sécurité. Les xanthates, par
exemple, largement employés en flottation des sulfures, sont toxiques pour le milieu récep-
teur. Ce type de réactifs induit également la formation de sous-produits (disulfure de car-
bone) pouvant porter atteinte à la santé humaine en cas de trop grande exposition (limite à 1
ppm pour le CS2 en Ontario [4]).
Bien que le domaine ait été bien peu exploré comparativement aux technologies tra-
ditionnelles, quelques produits d’origine biologique ont fait l’objet d’études cherchant à
développer des réactifs capables de remplacer les réactifs traditionnels [5-9]. Cette nouvelle
avenue est une option intéressante puisque les bio-réactifs peuvent permettre un impact
réduit sur l’environnement et un traitement plus facile dans l’effluant des usines. C’est donc
dans ce contexte de développement de bio-réactifs de flottation qu’est situé le présent tra-
vail.
1.2. Procédés de flottation
1.2.1. Concept général
Une fois extrait de la mine, le minerai est broyé afin de libérer le plus possible les
différentes espèces minérales présentes dans la matrice rocheuse [3, 10]. Ceci implique un
broyage considérablement fin, habituellement de l’ordre de 100 μm et moins. Le matériel
fraîchement broyé est classifié par des tamis ou des hydrocyclones. Avant d’effectuer la
flottation, différents réactifs chimiques sont ajoutés pour contrôler le pH et
l’hydrophobicité des particules d’intérêt. Ces réactifs sont ajoutés au broyage ou encore lors
d’une étape de conditionnement préalable à la flottation. Le matériel est ensuite transféré
dans des équipements de flottation, dans lesquels de l’eau est ajoutée au minerai pour obte-
nir une pulpe de 30 à 35 % solide. Par la suite, de l’air est injecté et dispersé de sorte à for-
mer de fines bulles. Il existe différents types d’appareils de flottation tels que les cellules de
flottation mécaniques (Figure 2), les cellules contact, les colonnes de flottation, les cellules
Jameson, qui impliquent chacun des méthodes différentes permettant de maximiser le con-
tact entre les bulles et les particules minérales. Les particules les plus hydrophobes
s’accrochent donc préférentiellement aux bulles, ce qui leur permet de flotter jusqu’à la

6
surface pour être récupérées dans l’écume qui s’y forme. Il est également commun de pla-
cer plusieurs appareils de flottation en série afin d’augmenter le temps de résidence des
particules et maximiser le contact avec les bulles d’air (et donc augmenter les performances
du procédé). Généralement, les rejets d’une première cellule servent d’apport à une seconde
afin de répartir et de maximiser la récupération en fonction du tonnage et du temps de ré-
tention. En ce qui concerne le concentré, c’est-à-dire la portion flottée riche en minerai, il
peu subir une étape de nettoyage lorsque nécessaire. Il est alors acheminé à une autre cel-
lule ou colonne afin d’en augmenter la teneur (pourcentage de minerai d’intérêt).
Figure 2: Schéma conceptuel d’une cellule mécanique majoritairement employée pour les
procédés de flottation [11].
1.2.2. Réactifs de flottation
Collecteurs et déprimants
Plusieurs minerais possèdent un certain degré de flottabilité naturelle. Toutefois des
produits chimiques sont ajoutés afin d’altérer sélectivement les propriétés de surface des
espèces minérales en question, leur conférant un caractère plus ou moins hydrophobe, selon

7
le cas [3, 12]. Ainsi, les collecteurs sont ajoutés dans le but d’augmenter l’hydrophobicité
des particules sur lesquelles ils s’adsorbent. Les molécules employées contiennent généra-
lement un groupement polaire responsable de la liaison sur le minerai (principalement une
liaison ionique ou covalente) et un résidu non-polaire hydrophobe conférant le caractère
désiré à la surface (Figure 3). Contrairement aux collecteurs, les déprimants ont pour rôle
de diminuer la flottabilité des particules qu’on souhaite éliminer du concentré de minerai à
récupérer [3, 12].
Figure 3: Représentation du xanthate d'éthyle de potassium, largement utilisé pour la flot-
tation des sulfures. À gauche, on retrouve le résidu hydrocarboné (non-polaire); à droite on
retrouve le groupement polaire responsable de l'adsorption sur le minerai [13].
Moussants
D’autres produits peuvent être ajoutés afin d’optimiser les conditions de flottation
[10, 12]. L’ajout de collecteur au procédé permet d’augmenter l’hydrophobicité des espèces
minérales ciblées, mais n’implique pas nécessairement la formation d’une écume stable, qui
facilite la récupération du concentré. Dans ces conditions, des moussants sont employés
pour diminuer la taille des bulles et stabiliser l’écume; les molécules employées sont des
tensio-actifs qui possèdent un groupement polaire (hydrophile) et un groupement alipha-
tique (hydrophobe) qui, en s’arrangeant à l’interface air-liquide, induisent la formation de
mousse. Ils permettent de diminuer la tension de surface de l’eau.
Agents modifiants
Les agents modifiants sont quant à eux employés pour améliorer les performances
de flottation en modulant les interactions entre collecteurs, déprimants et minerais, de sorte
à augmenter la sélectivité et la performance du procédé [12]. Parmi ces agents on compte,
par exemple, les agents activants, qui permettent de faciliter l’adhésion des collecteurs sur

8
le minerai, des agents floculants et dispersants, utiles pour contrôler la récupération de par-
ticules fines, ainsi que des acides et des bases, servant à réguler le pH (paramètre important
qui influence grandement les propriétés de surface du minerai).
1.2.3. Flottation des sulfures
La flottation des sulfures est l’une des applications les plus répandues de ce type de
procédé dans l’industrie minière [3, 12]. La chalcopyrite (CuFeS2), la chalcocite (Cu2S), la
bornite (Cu5FeS4) et la covellite (CuS) sont parmi les sulfures de cuivres les plus répandus,
avec la chalcopyrite qui vient en tête. La chalcopyrite possède d’ailleurs un degré appré-
ciable de flottabilité naturelle. De plus, il est assez fréquent de retrouver de l’or parmi le
minerai de cuivre, ce qui augmente la valeur d’un gisement. Les xanthates sont les collec-
teurs les plus couramment utilisés pour les sulfures de cuivre.
La sphalérite (ZnS), un minerai de zinc, est également un sulfure de valeur écono-
mique considérable fréquemment traité dans l’industrie minière, tout comme la galène (PbS)
fréquemment associée à l’argent [3].
Toutefois, les minerais de valeur décrit ci-haut sont souvent retrouvés en compagnie
de sulfures de fer comme la pyrite (FeS2), la pyrrhotite (Fe(1-x)S, où x = 0 à 0,2) et
l’arsénopyrite (FeAsS), qui possèdent une bien moindre valeur et doivent donc être séparés
du cuivre et de l’or [3, 12, 14]. De plus, comme ils ont aussi la capacité de flotter en pré-
sence de xanthates, des procédures particulières doivent être mises en œuvre pour éviter la
récupération de sulfure de fer dans le concentré. Dans certains cas, il est favorable de dé-
primer ces sulfures de fer. Parmi les déprimants conventionnels de la pyrite, on retrouve
d’abord la chaux et l’air. Une augmentation de pH provoque en surface des sulfures ferreux
une formation d’oxyhydroxides de fer (FeOOH). Ces dépôts hydrophiles limitent
l’attachement des sulfures de fer aux bulles d’air lors de la flottation. Par exemple, en pré-
sence de xanthate à pH alcalin, la pyrite ne flotte pas très bien alors qu’un pH neutre ou
acide lui permet de flotter [12]. Tout comme l’ajout de chaux, une pré-aération de la pulpe
avec de l’air ou de l’oxygène provoque la même formation d’espèces hydrophiles de type
FeOOH en surface des sulfures de fer. D’autres déprimants comme le cyanure de sodium
(plus connu pour la séparation Cu-Pb), l’hydrogénosulfure de sodium, de l’amidon oxydé et
des dextrines dans certains cas, peuvent être employés. Le choix des déprimants varie d’un

9
procédé à l’autre, selon la composition du minerai, la taille des particules ou encore l’étape
de séparation (séparation grossière, concentrateur). La silice est un autre élément de la
gangue qui est largement répandu dans les gisements miniers et qu’on cherche habituelle-
ment à éliminer du concentré par l’emploi de déprimants. Toutefois, il est généralement
possible de séparer cette espèce minérale ainsi que d’autres éléments non sulfurés de la
gangue par l’emploi de xanthates uniquement [12], à condition que les sulfures d’intérêt
soient bien libérés au broyage (absence d’associations minéralogique avec la gangue). Il est
donc possible de moduler les conditions d’opération du procédé (variation du pH, ajout
d’agents modifiants) pour limiter l’activation de la gangue et diminuer sa flottabilité.
1.3. Revue de littérature : Interactions peptides/minerais et bio-réactifs de flottation
Plusieurs travaux ont permis d’évaluer le potentiel de différents produits d’origine
biologique en tant qu’agents de bio-flottation ou encore comme agent de chélation pour
divers matériaux. Des bactéries ont d’ailleurs fait l’objet de plusieurs études, de même que
certains polymères d’origine naturelle. L’interaction entre peptides et matériaux inorga-
niques a également été étudiée par l’entremise de différents acides aminés ainsi que par
l’emploi de bactériophages comme véhicule d’expression peptidique.
1.3.1. Bactéries
Les bactéries ont d’abord été considérées dans l’industrie minière pour des applica-
tions en biolixiviation. Ce concept est utilisé afin de récupérer des métaux à partir de rési-
dus miniers qu’il était difficile de récupérer par des méthodes conventionnelles[15].
La capacité des micro-organismes à utiliser de tels substrats implique en premier
lieu la capacité à se lier à la surface du minerai, par l’entremise de la paroi cellulaire char-
gée et la présence de polysaccharides et de protéines imbriquées dans cette même paroi [7].
Ceci est intéressant pour les procédés de flottation également puisqu’il y a démonstration
du potentiel des micro-organismes en tant que ligands efficaces. Plusieurs travaux de re-
cherche ont été publiés par rapport à l’utilisation de bactéries en tant que bio-réactifs de
flottation [7, 16-19].
10
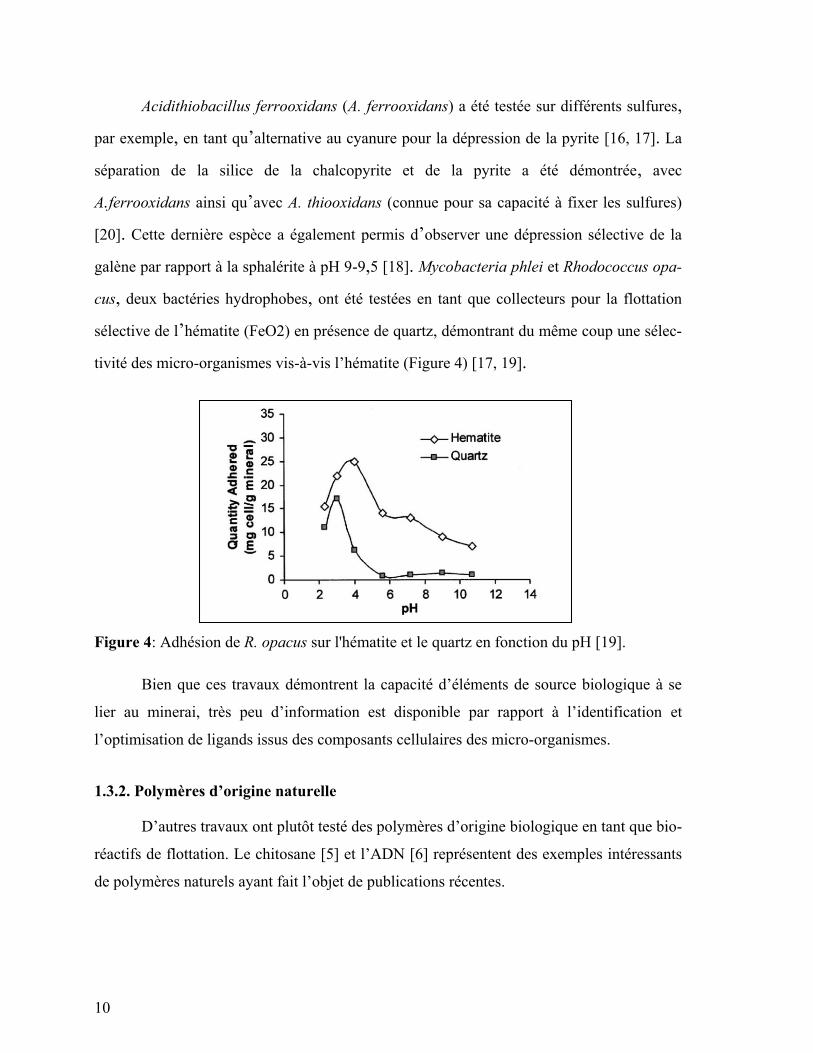
Acidithiobacillus ferrooxidans (A. ferrooxidans) a été testée sur différents sulfures,
par exemple, en tant qu’alternative au cyanure pour la dépression de la pyrite [16, 17]. La
séparation de la silice de la chalcopyrite et de la pyrite a été démontrée, avec
A.ferrooxidans ainsi qu’avec A. thiooxidans (connue pour sa capacité à fixer les sulfures)
[20]. Cette dernière espèce a également permis d’observer une dépression sélective de la
galène par rapport à la sphalérite à pH 9-9,5 [18]. Mycobacteria phlei et Rhodococcus opa-
cus, deux bactéries hydrophobes, ont été testées en tant que collecteurs pour la flottation
sélective de l’hématite (FeO2) en présence de quartz, démontrant du même coup une sélec-
tivité des micro-organismes vis-à-vis l’hématite (Figure 4) [17, 19].
Figure 4: Adhésion de R. opacus sur l'hématite et le quartz en fonction du pH [19].
Bien que ces travaux démontrent la capacité d’éléments de source biologique à se
lier au minerai, très peu d’information est disponible par rapport à l’identification et
l’optimisation de ligands issus des composants cellulaires des micro-organismes.
1.3.2. Polymères d’origine naturelle
D’autres travaux ont plutôt testé des polymères d’origine biologique en tant que bio-
réactifs de flottation. Le chitosane [5] et l’ADN [6] représentent des exemples intéressants
de polymères naturels ayant fait l’objet de publications récentes.

11
Chitosane
La Chalcopyrite et la galène sont généralement séparées par flottation. Des dépri-
mants inorganiques sont souvent utilisés, toutefois ces derniers sont toxiques, dangereux et
ne parviennent pas à diminuer l’entraînement des minéraux déprimés, surtout lorsque ceux-
ci sont présents en particules ultra fines. Une étude a exploré le potentiel du chitosane, un
polymère naturel non toxique issu de la chitine (élément retrouvé entre autres comme cons-
tituant de la carapace des invertébrés [21]), en tant que déprimant pouvant floculer sélecti-
vement les minéraux à être déprimés, aidant ainsi à contrer le phénomène d’entraînement
[5].
Suite aux tests de flottation individuels, il est apparu que la présence de chitosane
diminuait considérablement la récupération par flottation, passant de plus de 90 % à envi-
ron 60 % en présence de chitosane, tant pour la chalcopyrite que pour la galène. Cependant,
en présence d’un mélange galène et chalcopyrite, une flottation plus importante de la galène
a été observée, ce qui expose la possibilité d’utiliser le chitosane comme déprimant sélectif,
surtout à bas pH (Figure 5). Il y aurait donc présence d’une adsorption préférentielle du
chitosane sur la chalcopyrite, permettant d’expliquer que cette dernière soit plus faiblement
récupérée que la galène lors de tests de flottation.
Figure 5: Flottation de mélanges de chalcopyrite et de galène selon le pH, avec ajout de
chitosane. (a) Teneur des métaux dans l’écume; (b) Récupération des métaux dans l’écume
[5].
12
ADN
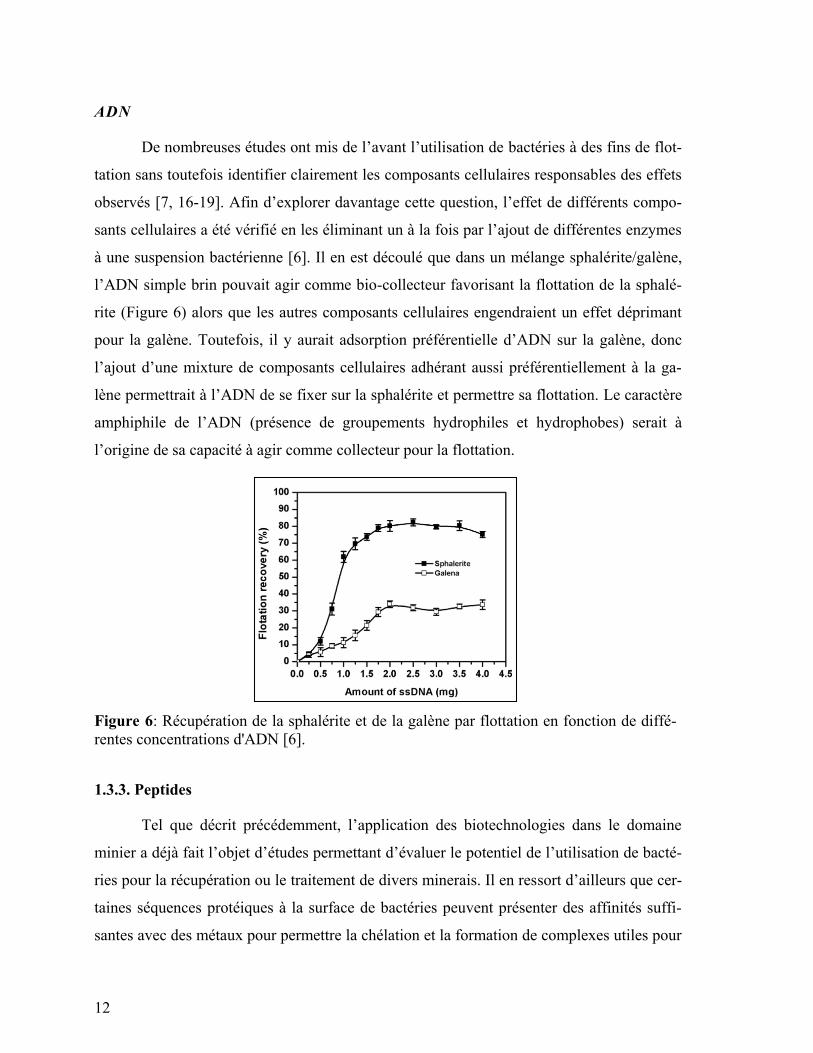
De nombreuses études ont mis de l’avant l’utilisation de bactéries à des fins de flot-
tation sans toutefois identifier clairement les composants cellulaires responsables des effets
observés [7, 16-19]. Afin d’explorer davantage cette question, l’effet de différents compo-
sants cellulaires a été vérifié en les éliminant un à la fois par l’ajout de différentes enzymes
à une suspension bactérienne [6]. Il en est découlé que dans un mélange sphalérite/galène,
l’ADN simple brin pouvait agir comme bio-collecteur favorisant la flottation de la sphalé-
rite (Figure 6) alors que les autres composants cellulaires engendraient un effet déprimant
pour la galène. Toutefois, il y aurait adsorption préférentielle d’ADN sur la galène, donc
l’ajout d’une mixture de composants cellulaires adhérant aussi préférentiellement à la ga-
lène permettrait à l’ADN de se fixer sur la sphalérite et permettre sa flottation. Le caractère
amphiphile de l’ADN (présence de groupements hydrophiles et hydrophobes) serait à
l’origine de sa capacité à agir comme collecteur pour la flottation.
Figure 6: Récupération de la sphalérite et de la galène par flottation en fonction de diffé-
rentes concentrations d'ADN [6].
1.3.3. Peptides
Tel que décrit précédemment, l’application des biotechnologies dans le domaine
minier a déjà fait l’objet d’études permettant d’évaluer le potentiel de l’utilisation de bacté-
ries pour la récupération ou le traitement de divers minerais. Il en ressort d’ailleurs que cer-
taines séquences protéiques à la surface de bactéries peuvent présenter des affinités suffi-
santes avec des métaux pour permettre la chélation et la formation de complexes utiles pour

13
la séquestration ou la récupération de métaux. Il apparait donc pertinent d’étudier les inte-
ractions entre peptides et minerais.
Différents acides aminés testés séparément
Quelques publications [22, 23] traitent de l’étude des interactions entre des compo-
sants de base des structures biologiques (peptides) et des surfaces inorganiques. Dans l’une
de ces publications [23], différentes surfaces inorganiques ont été exposées à des solutions
de peptides, chacune impliquant des polypeptides composés d’un acide aminé spécifique,
avec marqueurs fluorescents. La fluorescence a été mesurée pour quantifier la densité de
peptides adsorbés sur ces surfaces. Les matériaux inorganiques testés incluent des métaux
(Au, Pd, Pt, Ti et Al), des semi-conducteurs (GaAs, AlGaAs) et des isolants (Si3N4, SiO2).
Les tests ont permis d’observer une adhésion significative d’acides aminés chargés (chaînes
latérales acides et basiques) principalement sur les isolants (la silice et le nitrure de sili-
cium), tel que présenté au Tableau 2.
Tableau 2: Adhésion d'acides aminés sur différentes surfaces inorganiques avec de l'eau
comme solvant [23]

14
Protéines et peptides naturels se liant aux métaux
Les interactions avec les métaux sont courantes dans différents processus biolo-
giques. Par exemple, le fer est un élément essentiel de l’hémoglobine qui sert au transport
d’oxygène dans le sang. La fixation du fer est un phénomène important et résulte
d’interactions avec des composants de l’organisme. À cet effet, des travaux ont permis
d’isoler un polypeptide (NLGEQYFKG) pouvant se lier au fer. Ce nonapeptide a été obte-
nu par hydrolyse d’une protéine de plasma porcin [24]. Des peptides pouvant se lier au fer
ont également été isolés à partir de l’hydrolysat de sous-produits provenant du traitement de
crevettes [25]. Ces études évoquaient toutefois l’utilité de tels peptides en tant qu’additif
alimentaire.
Dans d’autres cas, il est question du rôle des peptides et protéines dans la fixation
des métaux dans une optique de bioremédiation [26, 27]. La capacité de protéines et pep-
tides à se lier aux métaux lourds tels que le cadmium et le mercure a été rapportée, souli-
gnant le potentiel de l’utilisation de plantes et micro-organismes exprimant ces protéines en
tant qu’agents de phytoremédiation et bioremédiation.
Polypeptides exprimés sur des bactériophages
L’identification de polypeptides affinitaires à divers matériaux inorganiques a fait
l’objet de quelques publications. En fusionnant une multitude de séquences peptidiques
aléatoires différentes sur l’une des protéines présentes à la surface de bactériophages fila-
menteux (de type M13 ou fd), une librairie d’expression phagique (ou encore « phage di-
play libraries ») peut être générée [28].
Ces phages filamenteux, de la famille des Inoviridae, dont la structure est illustrée à
la Figure 7) sont des phages à ADN simple brin spécifiques à E. coli [29]. L’infection des
bactéries par ces virus ne provoque pas la lyse des cellules mais provoque un ralentisse-
ment de rythme de reproduction. Le cycle de réplication de ce type de phages est bien do-
cumenté [29-32] et les étapes principales sont résumées ci-après. Le cycle débute par
l’adsorption de la protéine PIII sur le pilus F d’E. coli. L’infection se poursuit avec le dé-
sassemblage des protéines structurales principales PVIII (qui s’insèrent dans la membrane
cellulaire), et l’intrusion de l’ADN viral dans la cellule hôte. Les enzymes de l’hôte conver-

15
tissent ensuite cet ADN en ADN double brin qui permet d’abord la production des pro-
téines du phage, et ensuite à la duplication du matériel génétique effectuée par un méca-
nisme de réplication circulaire. Il y a alors formation d’un complexe nucléoprotéique avec
la protéine d’assemblage-réplication PV, qui protège l’ADN jusqu’à l’étape d’extrusion de
la particule virale, au cours de laquelle la protéine PV est remplacée par les protéines struc-
turales entreposées dans la membrane cellulaire. Le particule virale mature est finalement
relâchée dans le milieu et peut infecter d’autres cellules.
Figure 7: Structure du bactériophage M13. L’ADN du virus est enveloppé de 5 protéines
structurales différentes (PIII, PVI, PVIII, PVII et PIX).
Une librairie d’expression phagique mise en contact avec une cible définie permet
de tester des millions de candidats potentiels (tout dépendant du degré de complexité de la
librairie) au cours d’une même expérience et de sélectionner les meilleurs ligands de cette
très grande réserve de peptides. Des séquences peptidiques affinitaires, découlant d’un pro-
cessus de criblage d’une librairie de phages, ont été obtenues, entre autres, pour l’or [33],
des oxydes de silice et de titane [34], des oxydes de zinc [35], ainsi que pour du carbone de
type diamant [36]. Dans le dernier cas, il a d’ailleurs été démontré par une méthode
d’isothermes d’adsorption que l’affinité des peptides obtenus était de beaucoup supérieure à
celle des contrôles (Figure 8). Cependant, les matériaux mentionnés précédemment ont
surtout un intérêt lié à des applications nano technologiques.
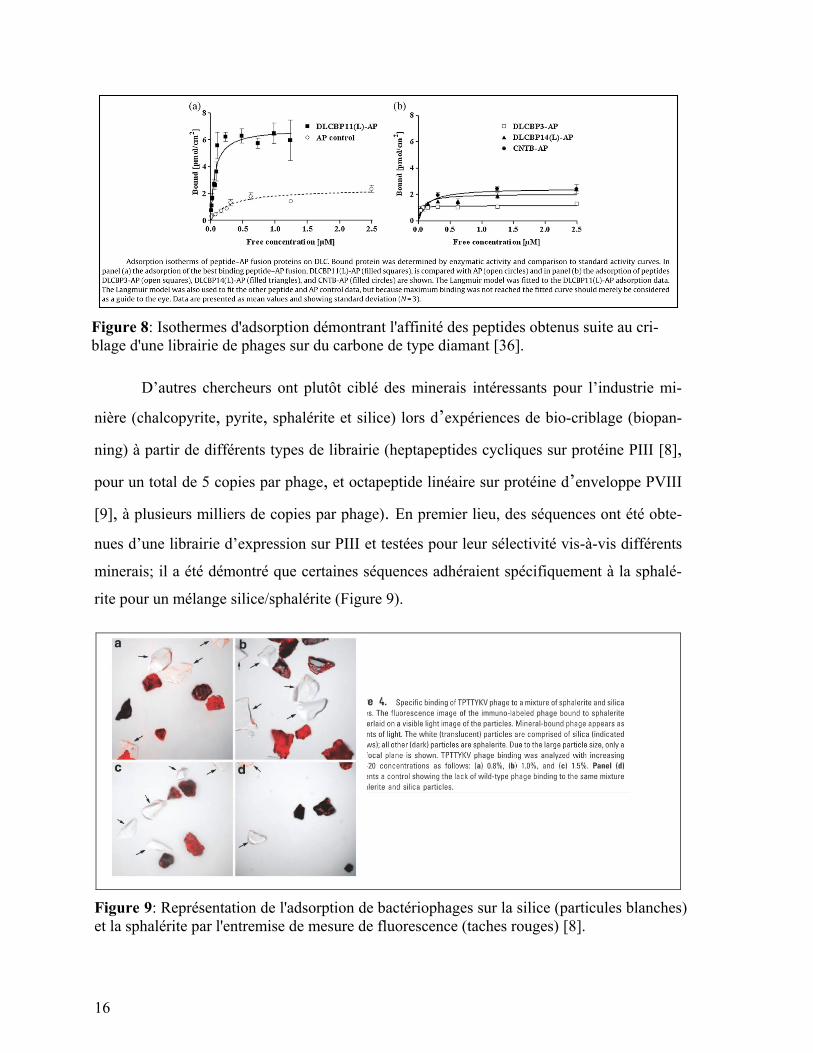
16
Figure 8: Isothermes d'adsorption démontrant l'affinité des peptides obtenus suite au cri-
blage d'une librairie de phages sur du carbone de type diamant [36].
D’autres chercheurs ont plutôt ciblé des minerais intéressants pour l’industrie mi-
nière (chalcopyrite, pyrite, sphalérite et silice) lors d’expériences de bio-criblage (biopan-
ning) à partir de différents types de librairie (heptapeptides cycliques sur protéine PIII [8],
pour un total de 5 copies par phage, et octapeptide linéaire sur protéine d’enveloppe PVIII
[9], à plusieurs milliers de copies par phage). En premier lieu, des séquences ont été obte-
nues d’une librairie d’expression sur PIII et testées pour leur sélectivité vis-à-vis différents
minerais; il a été démontré que certaines séquences adhéraient spécifiquement à la sphalé-
rite pour un mélange silice/sphalérite (Figure 9).
Figure 9: Représentation de l'adsorption de bactériophages sur la silice (particules blanches)
et la sphalérite par l'entremise de mesure de fluorescence (taches rouges) [8].

17
L’expression d’un peptide en plus grande quantité sur PVIII a résulté en une adsorp-
tion beaucoup plus spécifique de bactériophages (Figure 10) [9]. Ainsi, des clones sélectifs
à la chalcopyrite et la pyrite ont pu être isolés. La sélectivité respective des différents clones
vis-à-vis les différents minerais a été vérifiée en comparant l’adsorption par méthode im-
muno-enzymatique (ELISA).
Figure 10: Comparaison de l'adsorption de bactériophages décorés de séquences octapep-
tidiques différentes sur la chalcopyrite, la sphalérite, la silice et la pyrite [9].
De plus, l’effet de l’adsorption de bactériophages sur le caractère hydrophobe ou
hydrophile de la surface du minerai a été testé [9]. Des mesures de temps d’induction ont
permis de constater une diminution de l’hydrophobicité des minerais en présence de bacté-
riophages (augmentation du temps d’induction, Figure 11). Le temps d’induction représente
le temps requis pour observer l’attachement de particules de minerai suite au contact avec
une bulle d’air, simulant le comportement d’attachement dynamique tel que retrouvé en
condition de flottation.
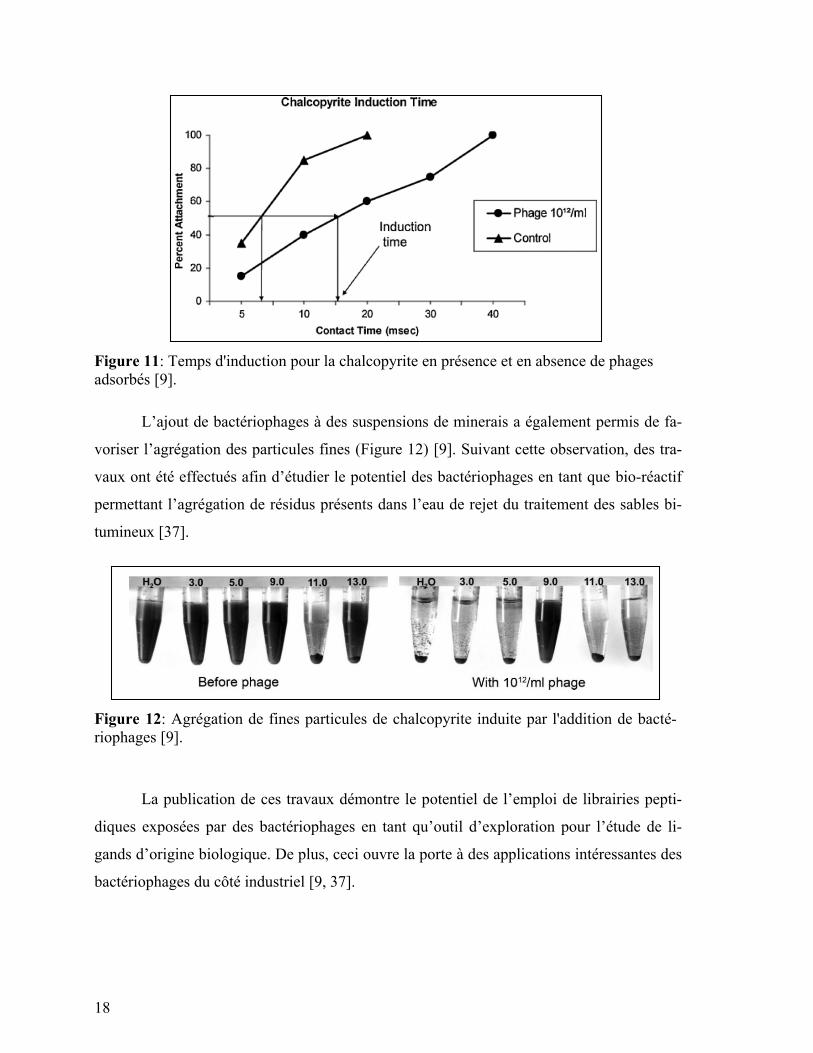
18
Figure 11: Temps d'induction pour la chalcopyrite en présence et en absence de phages
adsorbés [9].
L’ajout de bactériophages à des suspensions de minerais a également permis de fa-
voriser l’agrégation des particules fines (Figure 12) [9]. Suivant cette observation, des tra-
vaux ont été effectués afin d’étudier le potentiel des bactériophages en tant que bio-réactif
permettant l’agrégation de résidus présents dans l’eau de rejet du traitement des sables bi-
tumineux [37].
Figure 12: Agrégation de fines particules de chalcopyrite induite par l'addition de bacté-
riophages [9].
La publication de ces travaux démontre le potentiel de l’emploi de librairies pepti-
diques exposées par des bactériophages en tant qu’outil d’exploration pour l’étude de li-
gands d’origine biologique. De plus, ceci ouvre la porte à des applications intéressantes des
bactériophages du côté industriel [9, 37].

19
1.4. Objectifs
En tenant compte des préoccupations environnementales reliées aux réactifs de flot-
tation traditionnels, l’exploration de nouvelles options axées sur les biotechnologies s’avère
pertinente. D’ailleurs, comme il s’agit d’un procédé de séparation, la sélectivité des réactifs
employés est cruciale. Dans une optique de développement de bio-réactifs, l’un des aspects
les plus importants est donc d’identifier des ligands sélectifs par rapport aux différentes
cibles. À cet effet, l’emploi d’outils permettant d’étudier une grande variété de ligands (tel
qu’une librairie d’expression phagique) est approprié, tout comme l’application d’une dé-
marche permettant de vérifier la sélectivité des ligands à l’étude. En dernier lieu, afin
d’examiner le potentiel des ligands sélectionnés pour la flottation, une méthode permettant
de vérifier l’effet procuré à la surface du minerai est nécessaire.
La flottation est employée pour le traitement d’une multitude de minerais, mais la
séparation des sulfures représente l’une des applications les plus répandues. D’autre part,
l’importance économique des métaux précieux justifie l’intérêt qui leur est porté au niveau
industriel. Combinant ces considérations avec les besoins du partenaire industriel (CO-
REM), les minerais ciblés dans ce travail sont la chalcopyrite, la sphalérite et l’or, pour leur
valeur économique. En contrepartie, la silice et la pyrite sont des composantes importantes
de la gangue associée aux minerais mentionnés précédemment et doivent généralement être
éliminés des concentrés, ce pourquoi ils sont également considérés dans cet ouvrage.
Les objectifs de ce mémoire peuvent donc être résumés en trois étapes principales :
1. Identifier, par l’entremise du criblage d’une librairie de bactériophages, des sé-
quences peptidiques ayant la capacité de se lier à différents minerais d’intérêt indus-
triel, tels que la chalcopyrite, la sphalérite, l'or, la pyrite et la silice.
2. Vérifier la sélectivité des séquences obtenues en comparant l’adsorption des clones
arborant ces séquences sur les différents minerais.
3. Examiner l’effet de l’adsorption de bactériophages sur la surface des minerais dans
une optique d’application à un procédé de flottation.


21
CHAPITRE 2 : MATÉRIEL ET MÉTHODES


23
2.1. Caractéristiques et traitement des phases minérales
2.1.1. Caractéristiques des minerais
Les échantillons de chalcopyrite, de pyrite et de sphalérite ont été obtenus de
Ward’s Science et proviennent respectivement de Zacatecas au Mexique (chalcopyrite et
pyrite), et des mines Piedras Verdes (état de Chihuahua, Mexique). La silice a quant à elle
été obtenue de Sigma Aldrich (produit # 274739, lot # MKBG7577V) alors que la poudre
d’or a été obtenue d’Alfa Aesar (lot # B27Y009). La taille de particules était inférieure à 38
μm pour les tests de bio-criblage et les phases minérales ont été conservées à -20°C pour
limiter l’oxydation des sulfures exposés à la surface du minerai. La surface spécifique de
chacun des minerais a été obtenue par analyse BET, effectuées au laboratoire du professeur
Serge Kaliaguine du département de génie chimique de l’Université Laval. Un prétraite-
ment à 150°C d’une durée de deux heures a été effectué (pour retirer toute trace d’eau) et
l’expérience BET a été accomplie par adsorption d’azote à -196°C (refroidissement à
l’azote liquide). Pour les tests de temps d’induction, la tranche granulométrique de 45 à 90
μm a été employée afin de permettre la visualisation des particules (détails à la section 2.4.).
2.1.2. Traitement à l’EDTA
Afin d’obtenir une qualité de surface se rapprochant le plus possible de celle retrou-
vée immédiatement après le broyage, les sulfures ont été soumis à un traitement à l’EDTA
avant chacune des expériences impliquant l’adsorption de bactériophages. Cette étape per-
met d’extraire sélectivement les oxydes métalliques, les hydroxydes, les carbonates et les
sulfates formés à la surface des sulfures, sans altérer ces derniers.[38]
Le traitement a été effectué avec une solution 3 % (m/m) d’EDTA ajustée à pH 7,5
(NaOH). La solution a été désoxygénée au métabisulfite de sodium afin d’éviter la forma-
tion de nouveaux oxydes suite à la dissolution de la couche initiale. Cette solution a ensuite
été ajoutée aux échantillons à traiter de manière à former une pulpe de 25 % massique de
minerai et les contenants (tubes, erlenmeyer, etc.) ont alors été scellés et incubés avec agita-
tion pour environ une heure. Le surnageant contenant les oxydes dissouts a été jeté et le
minerai rincé à l’eau distillée désoxygénée ou à l’aide de la solution tampon utilisée pour

24
les expériences prévues. En cas d’utilisation ultérieure, le matériel préparé a été conservé
dans l’éthanol à -20°C, puis rincé au moment de l’emploi.
2.2. Criblage de la librairie de phages
Afin de développer des réactifs de flottation efficaces, il est essentiel d’identifier des
ligands sélectifs par rapport aux différents minerais. Bien que quelques travaux aient été
publiés sur le sujet [5, 6, 8, 9], davantage de connaissances par rapport aux interactions
peptides/minerais sont requises. D’autre part l’identification de la nature de telles interac-
tions ou la simulation numérique basée sur des modèles fondamentaux d’interactions molé-
culaires représente un travail extensif et difficile à réaliser. L’absence de données expéri-
mentales exhaustives fait en sorte qu’il serait ardu d’obtenir par simulation des séquences
polypeptidiques typiques ayant un bon potentiel en tant que ligand pour les minerais. Dans
cette optique, le criblage d’une librairie de peptides présentée par des phages permet de
mettre en contact de nombreuses séquences d’acides aminés avec les minerais à tester, et ce
au cours d'une même expérience. Il est alors possible d’identifier des ligands sans avoir à
étudier fondamentalement la nature des interactions entre la cible et les polypeptides, ce qui
simplifie grandement le processus. D’ailleurs, comme les séquences d’acides aminées fu-
sionnées font partie du génome des phages, elles sont identifiables par séquençage de
l’ADN des virus. De plus, les bactériophages peuvent être produits rapidement et en grande
quantité par l’entremise d’un hôte bactérien, ce qui facilite la transition vers des étapes liées
à la caractérisation de l’adsorption des phages.
2.2.1. Choix de la librairie
Les travaux ont été effectués avec la librairie d’expression Ph.D.MD C7C (New En-
gland Biolabs). Cette librairie est construite de sorte à inclure toutes les combinaisons pos-
sibles d’heptapeptides, exprimés à la surface de différents bactériophages M13. Le proces-
sus de génération de la librairie implique l’insertion de séquences d’ADN randomisées
(correspondant à différents heptapeptides) au gène codant pour la production de la protéine
PIII du bactériophage (responsable de l’adhésion au pilus F d’E. coli lors de l’infection),
située à l’extrémité de la particule virale (Figure 7). Ces phages génétiquement altérés ex-

25
priment donc une protéine PIII naturelle, mais à laquelle est fusionnée une séquence de 13
acides aminés supplémentaires incluant l’heptapeptide variable (Figure 13).
Figure 13: Séquences insérées pour la fabrication de librairies peptidiques (Ph.DMD, NEB)
sur le bactériophage M13 [39].
En considérant un total de 20 acides aminés, une librairie d’heptapeptides comporte
virtuellement 207 combinaisons différentes, soit 1,28 milliards de types de clones distincts
(un clone représente une population de phages possédant la même altération génétique).
Ces séquences peptidiques sont fusionnées à chacune des cinq copies de la protéine PIII.
De plus, contrairement aux autres librairies disponibles, la séquence variable d’acides ami-
nés est intercalée entre une paire de cystéines (librairie Ph.D.-C7C, Figure 13) qui, par la

26
formation d’un pont disulfure entre ces deux résidus, impose une contrainte conformation-
nelle au peptide. Cette contrainte engendre la présentation du peptide sous forme d’une
boucle. Cette particularité, combinée au fait que les peptides soient présents en seulement
cinq exemplaires sur les phages, devrait favoriser l’obtention de séquences plus sélectives
[39, 40]. Un autre type de librairie intéressante aurait été celle développée par Petrenko et
al [28] et testée avec succès[9], où l'heptapeptide était exprimé sur toutes les protéines d'en-
veloppe (PVIII) du phage filamenteux de type fd. Cependant, cette librairie n’est pas dispo-
nible commercialement et la construction complète d’une libraire d’expression phagique à
partir du phage original n'était pas réalisable dans le cadre de ce projet.
2.2.2. Procédures générales reliées au bio-criblage
Le bio-criblage ("biopanning"), qui représente le processus requis pour la sélection
et l’enrichissement de séquences peptidiques affinitaires, a été réalisé en suivant le proto-
cole fourni avec la librairie de phages de New England Biolabs [39]. Quelques modifica-
tions ont été apportées afin d’adapter certaines sections de ce protocole au criblage de la
librairie sur une cible composée de fines particules de minerai. Le protocole complet, de
même que les détails concernant la préparation des tampons et des solutions, sont dispo-
nibles en annexe.
Préparation du minerai (cible)
Le minerai a été nettoyé à l’EDTA (tel que décrit à la section précédente) pour éli-
miner les oxy-hydroxydes sans altérer les sulfures. Un échantillon de minerai de 75 mg [34]
a ensuite été suspendu dans du tampon TBS (tampon salin Tris 50 mM, NaCl 150 mM, pH
7,5) additionné de polysorbate 20 (TweenMD 20) (croissant d’une ronde à l’autre: 0,1 % v/v
pour la première ronde, 0,5 % pour la deuxième ronde et 0,75 % pour les suivantes). La
masse de minerai a été abaissée à 25 mg à partir de la deuxième ronde. L’augmentation de
la concentration de polysorbate et la diminution de la quantité de minerai avait pour but
d’augmenter la rigueur des conditions de sélection lors des rondes pour lesquelles le ren-
dement était moins important. Lors des rondes initiales, moins de copies des clones étaient
présentes; il était donc souhaitable de permettre une sélection moins sévère afin d’éviter de
perdre des candidats potentiels [39, 41].

27
Culture bactérienne en vue du titrage ou de l’amplification
La souche E. coli ER2738, recommandée pour le bio-criblage, a été obtenue de New
England Biolabs et les méthodes de culture sont tirées des recommandations du fournisseur
[39]. La souche a d’abord été cultivée sur boîte de Petri composée de milieu LB avec anti-
biotique (tétracycline) à 37°C pour une nuit. Cette souche possède un gène de résistance à
la tétracycline lié à la production du pilus F, qui sert de site d’adsorption des bactério-
phages. L’utilisation de l’antibiotique lors des pré-cultures assure donc le maintien de cette
caractéristique des cellules, essentielle à la réplication des phages. L’inoculum utilisé lors
du titrage ou de l’amplification provient de pré-cultures préparées dans du milieu
LB+Tet20 (20 mg/L de tétracycline), inoculées à partir d’une colonie de la boîte de Petri
puis incubées à 37°C avec forte agitation (200-250 rpm). La culture sur boîte de Petri a été
conservée pour un maximum d’un mois à 4°C alors que les pré-cultures liquides ont été
conservées pour 4-5 jours seulement, également à 4°C.
Bio-criblage
Une portion de la librairie a été ajoutée aux échantillons de minerais de sorte à obte-
nir une représentation équivalente à 100 copies de chaque clone, chacun représentant une
séquence distincte de 7 acides aminés (heptapeptide). Après une incubation d’une heure
sous légère agitation à la température de la pièce, les échantillons ont été brièvement centri-
fugés afin d’en récupérer le surnageant contenant les phages non liés. Le culot a été rincé
une dizaine de fois avec du TBST (concentration croissante de polysorbate, comme pour la
préparation des échantillons). L’élution des phages liés aux minerais a été effectuée avec
une solution de glycine-HCl (pH 2,2). L’éluat récupéré a été neutralisé avec une solution
Tris-HCl pH 9,1 afin d’éviter la dégradation des phages, puis une titration virale a été effec-
tuée afin de vérifier le rendement de l’expérience. Le rendement devrait augmenter d’une
ronde à l’autre puisque les meilleurs candidats devraient être enrichis au cours du processus.
Le reste de l’éluat a été amplifié en milieu LB inoculé à 1 % avec E. coli ER2738. Les
phages contenus dans le surnageant de l’amplification ont été purifiés par une double préci-
pitation au PEG, puis resuspendus dans du TBS. L’éluat amplifié et purifié a été titré afin
d’en connaître la concentration et d’évaluer le volume à ajouter lors de la ronde suivante,
pour obtenir un apport de phages comparable à la première ronde. Le tout a été répété pour
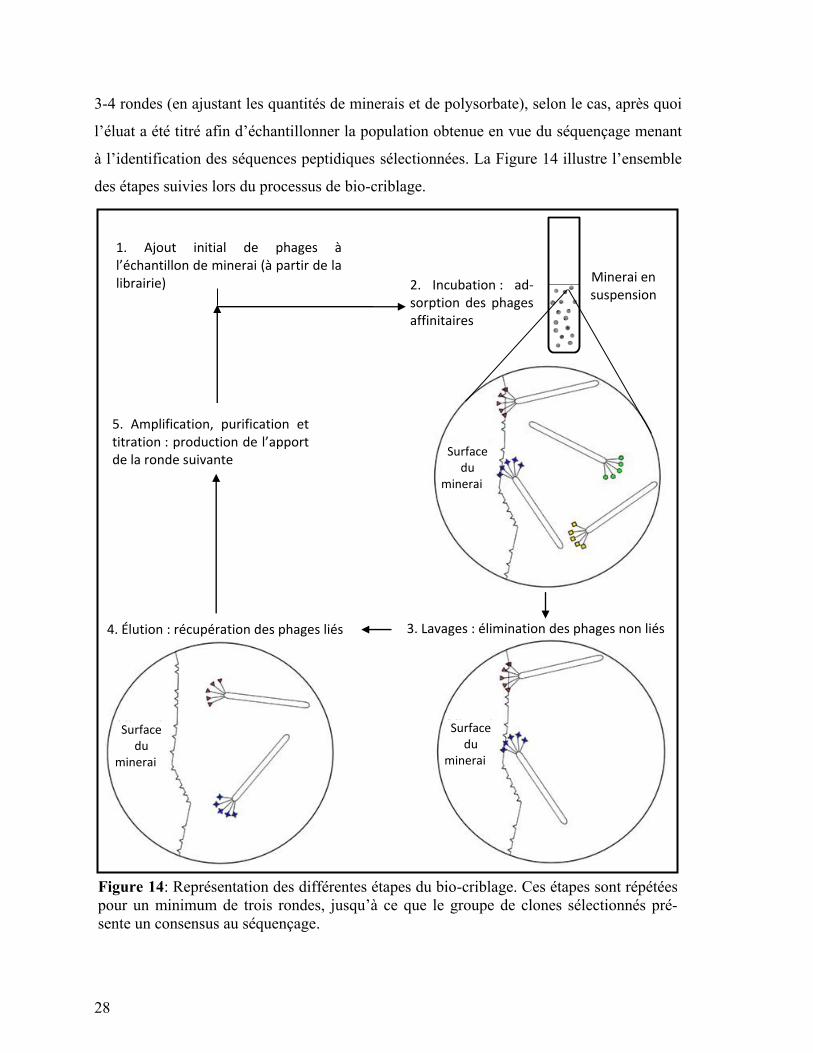
28
3-4 rondes (en ajustant les quantités de minerais et de polysorbate), selon le cas, après quoi
l’éluat a été titré afin d’échantillonner la population obtenue en vue du séquençage menant
à l’identification des séquences peptidiques sélectionnées. La Figure 14 illustre l’ensemble
des étapes suivies lors du processus de bio-criblage.
1. Ajout initial de phages à l’échantillon de minerai (à partir de la librairie) Minerai en
suspension 2. Incubation : ad-sorption des phages affinitaires
3. Lavages : élimination des phages non liés 4. Élution : récupération des phages liés
5. Amplification, purification et titration : production de l’apport de la ronde suivante
Figure 14: Représentation des différentes étapes du bio-criblage. Ces étapes sont répétées
pour un minimum de trois rondes, jusqu’à ce que le groupe de clones sélectionnés pré-
sente un consensus au séquençage.
Surface du
minerai
Surface du
minerai
Surface du
minerai

29
Titration des bactériophages
Essais sur plaque
La titration a été effectuée en employant la méthode de plages d'infection [39, 42].
L’échantillon à quantifier est dilué plusieurs fois (facteur 10 pour chaque dilution) dans du
milieu LB, puis mélangé à un petit volume de « top agar » inoculé avec E. coli.
L’échantillon est ensuite versé sur une boîte de Petri composée de milieu de culture auquel
a été ajouté de l’IPTG et du X-gal. Comme le bactériophage M13 ne forme pas de plages de
lyse mais plutôt des zones de croissance bactérienne plus faible, les clones de la librairie
sont conçus de manière à inclure dans leur génome le gène lacZα (lié à la production de β-
galactosidase). L’expression de ce gène par les bactéries suite à l’infection par les phages
leur confère la capacité de dégrader le X-Gal (un galactoside, hétéroside du galactose); son
hydrolysation produit un substrat bleu, facilement identifiable lors d’essais sur plaque (Fi-
gure 15). Il faut donc ajouter de l’IPTG et du X-Gal à la composition du milieu de culture
des boîtes de Petri utilisés pour ces essais. Après incubation, la quantité de phages de
l’échantillon peut être déterminée en dénombrant les plages bleues formées sur le tapis bac-
térien (unités formatrices de plages ou « plaque forming unit », pfu/mL), en appliquant le
facteur de dilution utilisé.
Figure 15: Exemple d'essai sur plaque. Chaque bactériophage actif présent dans l'échantil-
lon initial produit une zone de croissance bleutée.

30
Méthode par absorbance UV
Pour des mesures rapides d’échantillons purifiés, resuspendus dans du TBS, (pour
évaluer le résultat d’une amplification par exemple) une méthode spectrophotométrique a
été employée [43]. En raison de la contribution importante des protéines (pic d'absorbance
à 280 nm) par rapport à l’ADN (pic d'absorbance à 260 nm) dans le spectre d’absorbance
des bactériophages, une absorbance dans cette plage, avec un maximum à 269 nm peut être
mesurée. La valeur de cette absorbance maximale est proportionnelle à la concentration de
phages de l’échantillon (moyennant la soustraction de l’absorbance mesurée à 320 nm per-
mettant de compenser pour l’absorbance des chromophores de phage). Un blanc est égale-
ment requis pour soustraire l’absorbance du tampon. Finalement, la formule employée
(équation 1) tient compte de la taille du génome du phage, proportionnelle à la taille du
phage et donc de la quantité de protéines.
𝑇𝑖𝑡𝑟𝑒 (𝑝ℎ𝑎𝑔𝑒𝑠
𝑚𝐿⁄ ) =(𝐴269 − 𝐴320) ∙ 6 ∙ 1016
𝑡𝑎𝑖𝑙𝑙𝑒 𝑑𝑢 𝑔é𝑛𝑜𝑚𝑒 𝑑𝑢 𝑝ℎ𝑎𝑔𝑒 (𝑛𝑏 𝑑𝑒 𝑏𝑎𝑠𝑒)∙ 𝑓𝑎𝑐𝑡𝑒𝑢𝑟 𝑑𝑒 𝑑𝑖𝑙𝑢𝑡𝑖𝑜𝑛
Équation 1: Évaluation du titre phagique par spectrophotométrie
Cette méthode a l’avantage de pouvoir être réalisée en une quinzaine de minutes
comparativement aux essais sur plaque qui requièrent au moins 10 heures d’incubation. En
contrepartie, l’échantillon doit être suffisamment pur (suspendu dans du TBS) afin d’éviter
d’affecter le spectre d’absorption. La valeur obtenue ne tient également pas compte de
l’activité des phages, mais plutôt d’un nombre total de particules virales, impliquant un
ratio phage infectieux/phage totale pouvant être variable (sujet traité plus en détail à la sec-
tion 4.2). D’autre part, le seuil de détection de cette méthode est assez élevé: une concentra-
tion minimale d’environ 1011 phages/mL est requise. La spectrophotométrie n’a donc pas
été employée pour des mesures précises à basses concentrations.

31
Amplification de clones individuels
Petite échelle
Suite à la titration de la dernière ronde de bio-criblage, des plages infectieuse sont
sélectionnées et amplifiées afin d’obtenir suffisamment de matériel pour procéder au sé-
quençage d’une partie dy génome des phages [39]. Chaque plage a été piquée à l’aide d’un
embout de micropipette et employée pour infecter 1 mL de milieu LB préalablement inocu-
lé avec E. coli. Après une incubation à 37°C pendant 4-5 heures, les échantillons ont été
centrifugés pour en séparer les bactéries et les surnageants transférés dans des tubes stériles.
Une méthode d’extraction et purification d’ADN a ensuite été effectuée pour procéder au
séquençage. Pour conservation à long terme, un volume de culture d’un clone amplifié peut
être entreposé à -20°C, dilué dans un volume égal de glycérol.
Afin de produire davantage de matériel biologique pour des tests subséquents, les
clones individuels préalablement préparés sont employés pour infecter 20 mL de milieu LB
inoculé à 1 % (v/v) d’une culture E. coli (cultivée pendant une nuit). Cette pré-culture, in-
cubée pour 2-3 heures à 37°C, sert ensuite d’inoculum pour une culture d’un litre, qui est
incubée pour 4 heures supplémentaires à 37°C [44].
Fermentation en bioréacteur de 20 litres
À un certain point lors des travaux, une quantité importante de bactériophages était
requise pour la réalisation de diverses expériences autant à l’université que chez le parte-
naire industriel. D’autre part, il est pertinent d’avoir une idée du potentiel de production de
bactériophages afin de vérifier comment la technologie pourrait être transposée à l’échelle
industrielle advenant l’emploi de bactériophages comme bio-réactifs de flottation.
À cet effet, des travaux d’optimisation des conditions de fermentation pour la pro-
duction de phages filamenteux fd sont déjà rapportés dans la littérature [45, 46]. Par la réa-
lisation d’un plan d’expérience et d’une analyse par surface des réponses, il a été démontré
qu’une température de 28°C et un pH de 7 représentaient des conditions optimales; ces
conditions ont d’ailleurs permis d’obtenir une production finale de l’ordre de 3,5E11
pfu/mL [46]. Ces conditions, en plus d’une quantité d’oxygène dissous fixée à 100 % ont
été employées pour effectuer une production de 20 L en bioréacteur. Une pré-culture équi-

32
valant à 2 % du volume du réacteur, préparée tel que décrit pour l’amplification à petite
échelle, a servi d’inoculum pour cette expérience. La croissance bactérienne en cours de
production a été mesurée par densité optique à 600 nm (OD600) alors que la concentration
de bactériophages a été mesurée par essai sur plaque.
Purification d’échantillons d’ADN et séquençage
Une fois amplifiés, les échantillons de clones sélectionnés (obtenus à partir d’une
plage de lyse) ont été purifiés par une courte précipitation au PEG/NaCl (PEG 20 % / NaCl
2,5 M), puis centrifugés. L’ADN a été extrait par suspension du culot dans du tampon io-
dure et précipité par l’ajout d’éthanol. Le culot d’ADN obtenu a été resuspendu dans du
tampon EB et quantifié par spectrophotométrie à 260 nm avec un NanoDrop 2000c (Ther-
mo Scientific). Les échantillons d’ADN ont été séquencés par la Plate-forme d’Analyses
Génomiques de l’Université Laval. Un total de 16 clones issus de la dernière ronde ont été
sélectionnés pour le séquençage afin d’assurer une représentation suffisante pour
l’identification de séquences consensus.
2.3. Caractérisation de l’adsorption
Le processus de bio-criblage permet d’isoler des séquences peptidiques qui de-
vraient a priori avoir une affinité pour la cible à laquelle elles auront été exposées. Toute-
fois, il incombe de vérifier cette hypothèse en caractérisant plus précisément l’adsorption
des différents clones identifiés sur les minerais à l’étude. Dans cette optique, la détermina-
tion d’isothermes d’adsorption est un choix adéquat puisque ces derniers fournissent de
l’information quantitative sur le degré d’affinité et la spécificité des phages pour leurs
cibles et la capacité d’adsorption maximale des matériaux. De plus, cette méthode est assez
simple à réaliser comparativement à d’autres méthodes (telles que l’ELISA [9]). D’autre
part, cette méthode a l’avantage de représenter le phénomène dans des conditions
d’équilibre, ce qui se rapproche davantage de la sorption de réactifs de flottation en milieu
industriel que des méthodes impliquant le rinçage des échantillons, éliminant les ligands
non fortement liés.

33
2.3.1. Modèle d’interaction récepteur/ligand
L’application d’un modèle d’interactions récepteur/ligand (équation 2) sur les don-
nées expérimentales mènera à l'obtention des paramètres qui permettront la comparaison
quantitative des différentes expériences [47].
𝜑𝑙𝑖é =𝐵𝑚𝑎𝑥 ∙ 𝜑𝑙𝑖𝑏𝑟𝑒
𝐾𝑑 + 𝜑𝑙𝑖𝑏𝑟𝑒+ 𝑁𝑆 ∙ 𝜑𝑙𝑖𝑏𝑟𝑒
Équation 2: Modèle d'interaction récepteur/ligand [47]
Dans cette équation, 𝜑𝑙𝑖é représente la quantité de phages liés à la cible et 𝜑𝑙𝑖𝑏𝑟𝑒
correspond à la quantité de phages libres. D'autre part, la constante de dissociation (Kd) est
indicatrice du degré d’affinité du ligand par rapport au récepteur. Plus Kd est faible, plus
l'affinité du ligand (le phage) pour le récepteur (le minerai) est grande. Le paramètre Bmax
représente quant à lui la capacité d’adsorption maximale de phage par le minerai, alors que
l’importance de l’adsorption non spécifique sera proportionnelle au paramètre NS.
2.3.2. Génération des isothermes et calcul des paramètres
Les clones à tester ont été amplifiés et purifiés afin d’obtenir des quantités suffi-
santes pour effectuer les expériences d’adsorption. Pour chaque point expérimental, un
échantillon de 125 mg de minerai a été préparé et nettoyé à l’EDTA (dans le cas des sul-
fures). Des concentrations variables de phages, préparées dans du tampon TBST (0,25 %
Tween 20) ont été ajoutées aux minerais et incubés pendent une heure à la température de
la pièce, permettant ainsi d’atteindre l’équilibre. Après centrifugation, le surnageant (conte-
nant les phages non liés) a été récupéré et titré; par bilan de masse (équation 3), il a alors
été possible de déterminer la quantité de phages adsorbés et d’obtenir les points de
l’isotherme.
𝜑𝑙𝑖é =𝑉𝜑 ∙ (𝜑𝑙𝑖𝑏𝑟𝑒,0 − 𝜑𝑙𝑖𝑏𝑟𝑒,é𝑞)
𝑚𝑚 ∙ 𝑎𝑚
Équation 3 : Calcul de la quantité de phages liés

34
Où la quantité de phages liés (𝜑𝑙𝑖é) est en pfu/cm2, le volume de la solution de
phages exposée au minerai (𝑉𝜑 ) est en mL, la quantité de phages libres initiale et à
l’équilibre (𝜑𝑙𝑖𝑏𝑟𝑒,0 𝑒𝑡 𝜑𝑙𝑖𝑏𝑟𝑒,𝑒𝑞) sont en pfu/mL, la masse de minerai (𝑚𝑚) en grammes (g)
et la surface spécifique du minerai (𝑎𝑚) en cm2/g.
Ainsi, pour satisfaire l’équation 2, la constante de dissociation (Kd), la capacité
maximale d’adsorption (Bmax) et la constante d’adsorption non-spécifique (Ns) doivent res-
pectivement être rapportées en pfu/mL, pfu/cm2 et mL/cm2. La valeur de ces paramètres a
été calculée par régression non-linéaire. Des valeurs de paramètres sont d’abord fixées,
permettant de calculer des concentrations de phages liés (𝜑𝑙𝑖é) en fonction de la quantité de
phages libres ( 𝜑𝑙𝑖𝑏𝑟𝑒 ) expérimentale. À l’aide d’un logiciel (tel que l’outil Solveur
d’ExcelTM), la valeur des paramètres a été optimisée de sorte à minimiser l’erreur entre le
calcul issu de l’équation 2 et celui provenant de l’équation 3.
2.4. Mesure de temps d’induction
Une fois des séquences polypeptidiques identifiées et l’affinité vérifiée, il importe
de vérifier l’effet de l’adsorption des bactériophages sur les propriétés de surface du mine-
rai, dans une optique d’application aux procédés de flottation. Pour ce faire, un appareil
permettant de mesurer ce qui est appelé le temps d'induction a été utilisé chez le partenaire
industriel (COREM).
2.4.1. Concept de temps d’induction
Le temps d’induction est une mesure permettant de comparer le niveau
d’hydrophobicité de différents échantillons [3]. Cette méthode a été préférée à d’autres mé-
thodes, telles que la mesure d’angle de contact, puisqu’elle permet de prendre des mesures
qui se rapprochent davantage du modèle dynamique qui représente l’attachement d'une par-
ticule de minerai à une bulle dans les procédés de flottation (c'est-à-dire l'effet net d'une
compétition entre des phénomènes d’attachement et de détachement).
Une bulle est mise en contact avec un lit de particules pour un temps donné et l'atta-
chement ou non de particules à sa surface est détecté. L’expérience est répétée plusieurs
fois, de sorte que la proportion des essais qui résultent en l’attachement de particule(s)

35
permet d’obtenir un taux d’attachement pour un temps de contact donné. Des mesures sont
faites pour différents de contact. Le temps d’induction est défini comme le temps requis
pour engendrer un pourcentage d’attachement de 50 % [48]. Les essais ont été effectués
avec un minuteur d'induction (« induction timer »). L’appareil est composé d’une caméra
permettant d’observer des particules de faible diamètre, d’une plateforme supportant le lit
de particules et d’un système précis de déplacement actionné par haut parleur permettant de
mettre la bulle d’air en contact avec le lit (Figure 16).
Figure 16: Schéma de l’appareil de mesure de temps d’induction (induction timer).
2.4.2. Conditions expérimentales
La taille des particules employées lors de ces essais était de 45 à 90 μm, ce qui re-
présente le minimum requis pour bien visualiser le phénomène à l’aide de la caméra haute
définition utilisée sur le montage. De plus, comme pour les méthodes décrites dans les sec-
tions précédentes, le minerai a été nettoyé à l’EDTA. Les échantillons ont été rincés plu-
sieurs fois afin de dégager les fines particules du lit, assurant ainsi une bonne visibilité et
une plus grande constance de la taille des particules. Ces étapes ont été effectuées en limi-
tant le plus possible l’exposition à l’oxygène (utilisation d’eau désoxygénée pour les rin-

36
çages) afin d’éviter l’altération des surfaces par oxydation. L’espacement initial entre le lit
et la bulle, le déplacement, et le diamètre de bulle sont des facteurs qui doivent être bien
contrôlés afin d’offrir des conditions expérimentales semblables pour tous les essais. Ces
paramètres ont été définis respectivement à 200 μm, 400 μm et 2 mm. Il est également es-
sentiel de faire les répétitions sur des emplacements variables du lit de particules afin de
tenir compte de l’hétérogénéité de l’échantillon.

37
CHAPITRE 3: RÉSULTATS PRÉSENTÉS SOUS FORME D'ARTICLE


39
Criblage d’interactions peptide-minerai à l’aide d’une librairie d’expression phagique
pour une application en bio-flottation
Screening Peptide-Ore Interactions with a Phage Display Library for Bio-flotation appli-
cation
Karl Tremblay-Bouliane1, Caroline Olsen3, Denise Tremblay2, Sylvain Moineau2 Bruno
Gaillet1 and Alain Garnier1,*
1Université Laval, Department of Chemical Engineering 2Université Laval, Department of Biochemistry, Microbiology and Bio-informatics 3COREM, Research Group in Flotation and Extractive Metallurgy
*Corresponding author: [email protected]
3.1. Résumé en français
Une librairie d'heptapeptides présentés sur des bactériophages a été criblée dans le
but d’identifier des séquences peptidiques affinitaires à différents minerais (or, chalcopyrite,
sphalérite, pyrite et silice) et d’évaluer le potentiel de ces phages en bio-flottation. Ce bio-
criblage a permis d’isoler plusieurs séquences peptidiques différentes de celles disponibles
dans la littérature. Des isothermes d’adsorption ont été déterminées et l’application d’un
modèle d’interaction récepteur-ligand a permis de comparer l’affinité de chacune des sé-
quences peptidiques isolées par rapport aux minerais. Ainsi, certains phages possèdent une
affinité légèrement inférieure pour la sphalérite comparativement aux autres sulfures, ce-
pendant, cette affinité ne serait pas significativement différente du contrôle négatif, l'expo-
sition des minerais au phage d'origine. L’adsorption sur les sulfures est significativement
plus importante que sur la silice. En dernier lieu, des mesures de temps d’induction suggè-
rent que les bactériophages diminuent l’hydrophobicité de la surface des minerais (effet
déprimant).
*Cette section des résultats a été présentée sous forme d’article en anglais qui sera soumis
sous peu au journal CIM (Canadian Institute of Mining).

40
3.2. Abstract
A heptapeptide library displayed on bacteriophages was screened to identify peptide
sequences able to specifically bind to different ores (gold, chalcopyrite, sphalerite, pyrite
and silica) and evaluate phages potential as bio-flotation reagents. Many peptide sequences
were isolated through this biopanning and differed from other sequences found in the litera-
ture. Adsorption isotherms were determined and the application of a receptor/ligand inter-
action model allowed to compare the affinity of obtained peptide sequences towards the
ores. Some of the phages have a slightly lower affinity to sphalerite compared to other sul-
fides, but that affinity is not significantly different from the negative control, wildtype virus
exposition. Adsorption was found to be significantly higher on sulfides compared to silica.
Lastly, induction time measurements suggest that bacteriophages contribute to lower the
hydrophobicity of the ores surface (depressing effect).

41
3.3. Introduction
Froth flotation is a well known process widely used in the mineral processing indus-
try to purify and concentrate mineral ores [3, 12]. After being harvested from the mine, ores
are ground to a range going from a few micrometers up to about 100 μm (depending on the
process). Chemicals (collectors or depressants) are then added to specifically alter ores sur-
face properties while frothers are added to reduce bubble size and stabilize froth. Collectors,
such as xanthates in copper flotation, selectively increase hydrophobicity of desired parti-
cles and allow them to float with the help of an uprising air bubble flow. Depressants help
to decrease floatability of gangue to achieve better separation. Ores are harvested in the
froth thus generated at the top of the flotation cell/column.
Although this technology has been developed and used for over 100 years [2], envi-
ronmental regulations and constant need for improved flotation performances drives the
development of greener and better flotation reagents. For example, xanthate based collec-
tors are toxic (at variable concentrations) in water effluent [49, 50]. They can also generate
toxic and flammable decomposition products such as carbon disulphide, and cause sponta-
neous combustion as well as acute irritation when inhaled or absorbed [51]. Thus, they are
increasingly regulated by government authorities (1 ppm threshold for carbon disulfide in
Ontario, Canada [4]).
Bio-related products have the advantage of being relatively easy to treat in the efflu-
ent as they are generally easier to biodegrade and often pose little threat to human health.
They also have the potential of offering good selectivity, considering the almost endless
variety of bio-compounds found in nature. Many of those products, such as bacterial strains
[7] and bio-polymers like chitosan [5], or single stranded DNA [6] have been tested for
their capacity to specifically bind to different mineral ores and for their effect on flotation
performances. Recent works brought out promising results using bacteriophages peptide
libraries. Following the biopanning of such libraries on sulfide ores, heptapeptide and oc-
tapeptide sequences were isolated as specific binders to sphalerite and chalcopyrite. Phages
presenting the selected peptides were found to decrease hydrophobicity of ores and induce
aggregation [8, 9].

42
To follow up on the above results, this work focuses on studying the potential inter-
actions of polypeptide presenting phages with ores of economical interest as well as their
bio-flotation potential. The study was specifically aimed at the recovery of chalcopyrite
(CuFeS2), sphalerite (ZnS) and gold (often present in sulfide deposits), as opposed to re-
moval of silica and pyrite (FeS2), which are often found as gangue in sulfide ores. In order
to identify peptide sequences with potential affinity to those ores, a biopanning procedure
was carried out for each mineral phase and receptor-ligand interactions were studied by
adsorption isotherms, in order to assess peptide selectivity towards ores. Effect on surface
properties was also tested by induction time measurements and allowed to study the poten-
tial in bio-flotation application.
3.4. Materials and Methods
3.4.1. Phage library and biopanning
Biopanning with phage display libraries allows the screening of a large number of
potential ligands in a timely and effective manner. Phage display libraries are created by
modifying phage genome, with the insertion of DNA sequences that lead to the expression
of a short polypeptide on one of the phage structural proteins. The use of these bacterio-
phages allows rapid amplification and enrichment of selected ligands during biopanning
experiments. In this work, a M13 phage display library was obtained from New England
Biolabs (Ph.D C7CTM) and displays all possible heptapeptides combinations (complexity up
to 1.28∙109 different peptide sequences). A disulfide bond between two cysteins found at
both ends of the inserted peptides imposes a conformational constraint causing the peptide
to be presented as a loop. This library was chosen because a fixed conformation would pre-
sumably yield higher affinity ligands [40].
Chalcopyrite, sphalerite, and pyrite samples were obtained from Ward’s Science.
Chalcopyrite and sphalerite were mined in Zacatecas (Mexico) while pyrite was obtained
from Piedras Verdes mines (Chihuahua State, Mexico). Gold powder was obtained from
Alfa Aesar while silica was obtained from Sigma Aldrich (# 274739). Particle size was less
than 38 μm except for gold, which was 100 μm. Sulfide ores were leached with a deoxy-
genated 0.1 M EDTA solution pH 7.5 (50 % w/w ore and leaching solution) for an hour in

43
order to remove oxide deposits. Materials were stored in ethanol 95 % at -20°C until need-
ed.
Biopanning was conducted following NEB protocols. For the first round, 15 mg of
ore was incubated for an hour with a 100 fold representation of the library (100 copies of
each sequence, about 1011 phages) diluted in 1 mL TBS (50 mM tris-HCl pH 7.5, 150 mM
NaCl) + 0.1 % v/v polysorbate 20 (Tween 20; TBST 0.1 %). Samples were washed 10
times with TBST 0.1 % and elution was carried with 1 mL of 0.2 M glycine HCl (pH 2.2).
The eluate was neutralized with 150 μL of 1 M Tris-HCl (pH 9.1). Eluates were amplified
with E. coli ER2738 after each round and served as input stock for subsequent rounds (up
to four), in which polysorbate concentration was increased to 0.25, 0.5 and 0.75 % v/v for
rounds 2 to 4, respectively. Between each round, phages were quantified using plaque assay
in which IPTG/Xgal LB plates were used to allow the formation of easily identifiable blue
plaques. Following the biopanning procedure (after three to four rounds), 16 phage plaques
were picked from a titer plate, amplified for DNA extraction and DNA samples were sent
for sequencing, allowing the identification of deduced peptide sequences.
3.4.2. Adsorption isotherms
Ore samples (125 mg) were washed with EDTA and rinsed with deoxygenated
dH2O, to remove oxide deposits. Various titers of phages diluted in TBST 0.25 % (to limit
non specific interactions), for a total volume of 500 μL, were added to ores and incubated
for an hour under shaking at room temperature. Samples were briefly centrifuged and ti-
tered by plaque assay.
By measuring the partition coefficient between the two phases of the phage, either
free or bound, a quantitative evaluation of the sequence selectivity could be obtained. A
receptor-ligand interaction model [47] (equation 4) was applied to data and parameters
were estimated by non-linear regression (Figure 18). The non-specific interaction term (𝑁𝑆 ∙
𝜑𝑓𝑟𝑒𝑒 ) was omitted for computation purposes since polysorbate was added during adsorp-
tion experiments, thus limiting this type of interactions. The formation of a plateau on the
adsorption isotherms (Figure 17) justified that assumption. Bound phage was calculated by

44
subtracting the amount of phages remaining in the supernatant after the incubation to the
total quantity of initially added phages, as shown in equation 5.
𝜑𝑏𝑜𝑢𝑛𝑑 =𝐵𝑚𝑎𝑥 ∙ 𝜑𝑓𝑟𝑒𝑒,𝑒𝑞
𝐾𝑑 + 𝜑𝑓𝑟𝑒𝑒,𝑒𝑞+ 𝑁𝑆 ∙ 𝜑𝑓𝑟𝑒𝑒,𝑒𝑞
Equation 4: Receptor-ligand interaction model used for parameter determinationn
𝜑𝑏𝑜𝑢𝑛𝑑 =𝑉𝜑 ∙ (𝜑𝑓𝑟𝑒𝑒,0 − 𝜑𝑓𝑟𝑒𝑒,𝑒𝑞)
𝑚 ∙ 𝑎
Equation 5: Calculation of bound phages
In equations 4 and 5, 𝜑𝑏𝑜𝑢𝑛𝑑 represents the amount of bound phages (pfu/cm2),
while 𝜑𝑓𝑟𝑒𝑒,0 and 𝜑𝑓𝑟𝑒𝑒,𝑒𝑞 are the initial and equilibrium free phage concentrations in
pfu/mL. The total volume of phage solution (𝑉𝜑) is in mL, the mass of ore used (𝑚) in
grams and the specific surface area (a) in cm2/g. To satisfy equation 4, maximum binding
(Bmax), the dissociation constant (Kd) and non-specific binding parameter (NS) should be
respectively reported in pfu/cm2, pfu/mL and mL/cm2.
Specific surface area values were determined by BET experiments, consisting of a
pretreatment at 150°C for two hours followed by nitrogen adsorption at -196°C. Specific
surface area allowed the conversion of raw adsorption data to surface based values.
3.4.3. Induction time measurements
Induction time was measured to evaluate the effect of phage adsorption on ore sur-
face properties. This method requires a bubble to be lowered on a particle bed multiple
times for increasing contact times. Induction time is defined as the contact time required for
a 50 % particle attachment occurrence ratio over a series of assays [48]. A total of 20 assays
were carried for each contact time value. Particle size was 45-90 μm and ores were washed
with EDTA and stored in ethanol until used. Tests were performed using 1 g samples,
washed multiple times with deoxygenated dH2O in order to remove ethanol and smaller
particles. Different phage concentrations were prepared and incubated with ores (1 mL
phage solution per gram of pure ore) for one hour at room temperature. Bubble distance

45
from the bed was set at 200 μm for a displacement of 400 μm. Time to approach/retract
were kept at 5 ms, unless contact time was under 5 ms, in which case time to ap-
proach/retract was set equal to the contact time (time to approach/retract must be shorter or
equal to contact time for adequate measurements).
3.5. Results and discussion
3.5.1. Biopanning
Biopanning was successfully carried on each of the five target ores. Phage clones
were picked after 3-4 rounds and consensus heptapeptide sequences were identified (con-
sensus being assessed by observing higher than average representation of peptide sequences
in the sample pool; results presented in Table 3, detailed biopanning results in Appendix 3
of the thesis). Sulfide ores yielded similar sequences; phage clone C2 was obtained from
the screening on chalcopyrite, sphalerite, pyrite and gold, while phage Sp1 was isolated
from both chalcopyrite and sphalerite. Silica yielded considerably different sequences con-
sisting mostly of basic amino acids, as previously found in literature [23, 34, 52]. In fact,
most sequences (all ores considered) were mainly composed of polar amino acids, a fact
also mentioned in other studies implying phage display and inorganic materials [8, 34].
Sequence G1 was obtained from the biopanning of gold and from the control exper-
iment (in which biopanning was conducted in a tube without any ore particles), and can
therefore be considered a false positive. The G1 clone was highly represented (54 %) in the
phage pool obtained from the biopanning on gold, implying the absence of high affinity
binders, causing accumulation of non-specific sequences during the biopanning process.
Experiments were made in polypropylene microfuge tubes, for which false positives are
less likely to occur [53]. Since it can be assumed that binding to the tubes was negligible,
false positives obtained in this experiment could be the result of non-uniform amplification
of the eluted phages between biopanning rounds, leading to the enrichment of phages relat-
ed to infectivity rather than binding affinity to the target [54].
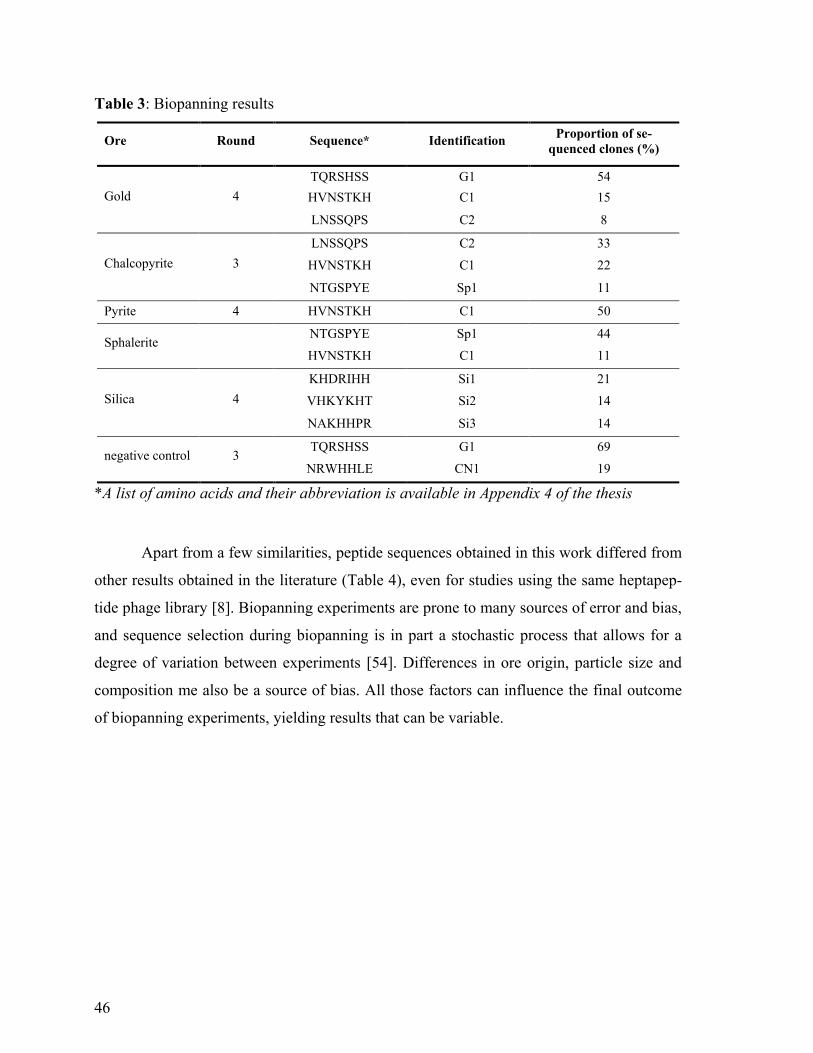
46
Table 3: Biopanning results
Ore Round Sequence* Identification Proportion of se-
quenced clones (%)
Gold 4
TQRSHSS G1 54
HVNSTKH C1 15
LNSSQPS C2 8
Chalcopyrite 3
LNSSQPS C2 33
HVNSTKH C1 22
NTGSPYE Sp1 11
Pyrite 4 HVNSTKH C1 50
Sphalerite NTGSPYE Sp1 44
HVNSTKH C1 11
Silica 4
KHDRIHH Si1 21
VHKYKHT Si2 14
NAKHHPR Si3 14
negative control 3 TQRSHSS G1 69
NRWHHLE CN1 19
*A list of amino acids and their abbreviation is available in Appendix 4 of the thesis
Apart from a few similarities, peptide sequences obtained in this work differed from
other results obtained in the literature (Table 4), even for studies using the same heptapep-
tide phage library [8]. Biopanning experiments are prone to many sources of error and bias,
and sequence selection during biopanning is in part a stochastic process that allows for a
degree of variation between experiments [54]. Differences in ore origin, particle size and
composition me also be a source of bias. All those factors can influence the final outcome
of biopanning experiments, yielding results that can be variable.
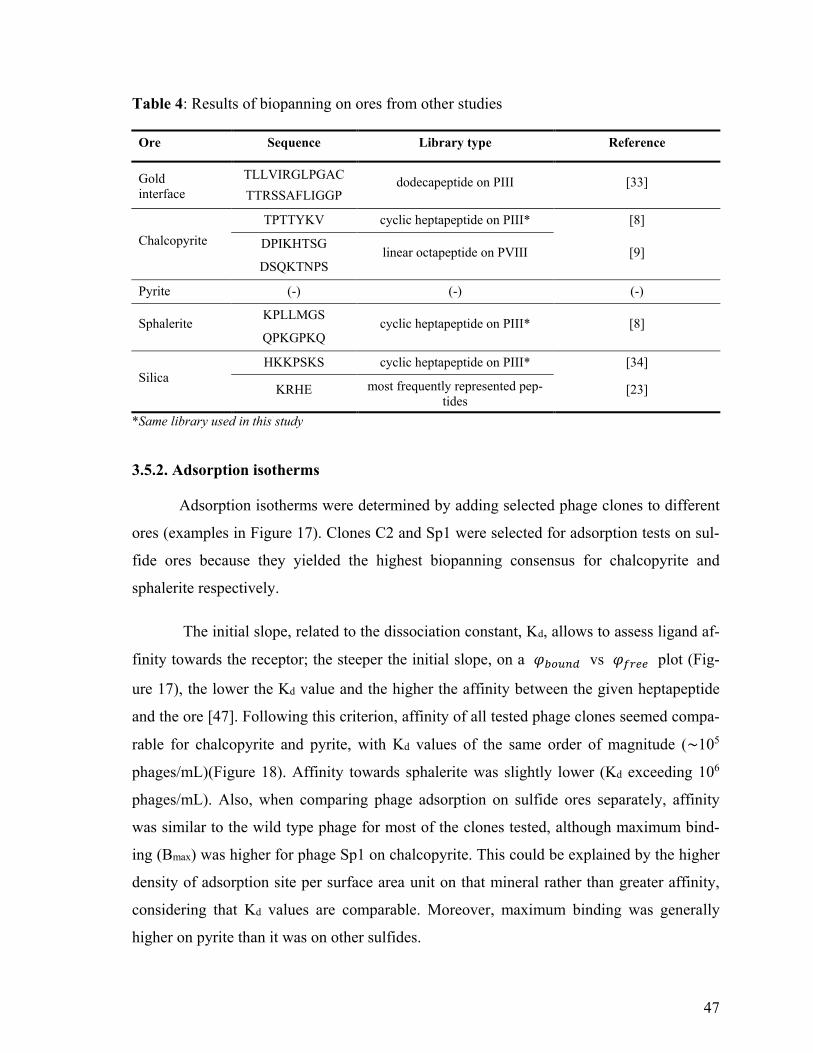
47
Table 4: Results of biopanning on ores from other studies
Ore Sequence Library type Reference
Gold
interface
TLLVIRGLPGAC dodecapeptide on PIII [33]
TTRSSAFLIGGP
Chalcopyrite
TPTTYKV cyclic heptapeptide on PIII* [8]
DPIKHTSG linear octapeptide on PVIII [9]
DSQKTNPS
Pyrite (-) (-) (-)
Sphalerite KPLLMGS
cyclic heptapeptide on PIII* [8] QPKGPKQ
Silica
HKKPSKS cyclic heptapeptide on PIII* [34]
KRHE most frequently represented pep-
tides [23]
*Same library used in this study
3.5.2. Adsorption isotherms
Adsorption isotherms were determined by adding selected phage clones to different
ores (examples in Figure 17). Clones C2 and Sp1 were selected for adsorption tests on sul-
fide ores because they yielded the highest biopanning consensus for chalcopyrite and
sphalerite respectively.
The initial slope, related to the dissociation constant, Kd, allows to assess ligand af-
finity towards the receptor; the steeper the initial slope, on a 𝜑𝑏𝑜𝑢𝑛𝑑 vs 𝜑𝑓𝑟𝑒𝑒 plot (Fig-
ure 17), the lower the Kd value and the higher the affinity between the given heptapeptide
and the ore [47]. Following this criterion, affinity of all tested phage clones seemed compa-
rable for chalcopyrite and pyrite, with Kd values of the same order of magnitude (~105
phages/mL)(Figure 18). Affinity towards sphalerite was slightly lower (Kd exceeding 106
phages/mL). Also, when comparing phage adsorption on sulfide ores separately, affinity
was similar to the wild type phage for most of the clones tested, although maximum bind-
ing (Bmax) was higher for phage Sp1 on chalcopyrite. This could be explained by the higher
density of adsorption site per surface area unit on that mineral rather than greater affinity,
considering that Kd values are comparable. Moreover, maximum binding was generally
higher on pyrite than it was on other sulfides.

48
A
B
Figure 17: Isotherms comparing adsorption of different phage clones on chalcopyrite
(A) and sphalerite (B), wild type phage served as a control experiment.
0,0
0,5
1,0
1,5
2,0
2,5
0,0 0,1 0,2 0,3 0,4 0,5
Ad
sorb
ed
ph
age
s (1
09
pfu
/cm
2)
Supernatant concentration (109 pfu/mL)
Sp1
C2
Wildtype
0,0
0,5
1,0
1,5
2,0
2,5
0,0 0,1 0,2 0,3 0,4 0,5
Ad
sorb
ed
ph
age
s (1
09p
fu/c
m2)
Supernatant concentration (109 pfu/mL)
Sp1
C2
Wildtype

49
The fact that comparable affinity between the control (M13KE clone, obtained from
NEB) and other clones obtained through the biopanning process was observed may suggest
that the phage particle itself tends to bind to sulfide ores, overcoming the possible affinity
of the fused heptapeptide. Only five copies of the inserted peptide sequence are available
on the entire phage (on phage protein PIII) while the phage coat contains roughly 3,000
proteins PVIII [55]. Such a library was chosen for its potential in yielding better ligand af-
finity, but this could also imply significant overall binding from the rest of the phage struc-
tural proteins during adsorption measurements. Adsorption isotherms are determined with
equilibrium values, without washing. This could allow weaker, but more numerous interac-
tions to play a role in phage adsorption, enough to mask the intrinsic interaction of the pep-
tide of interest [8, 54]. Low peptide affinity could also explain the similar binding observed
between control and some of the isolated phages. For example, a relatively simple simula-
tion can show that a 1 % selection efficiency advantage during biopanning would be
enough to yield consensus sequences over 4 rounds (details available as supplementary
material). Therefore, phages displaying sequences with an affinity level only slightly higher
than the wild type phage could still be enriched an isolated.
Despite displaying a very similar binding affinity on sulfide ores, phages showed
much less binding to silica (Figure 18A). Maximum binding (Bmax) was about ten times
lower compared to pyrite and about 5 times lower compared to chalcopyrite and sphalerite.
Differences in binding affinity were also noted for different phages tested on silica (Figure
18C). The wild type phage presented less affinity than phage Si1, a clone obtained from the
biopanning carried on silica.

50
A
B
C
Figure 18: Estimated parameters for receptor-ligand interaction model. Maximum bind-
ing, Bmax (A), shows that phages bind significantly less to silica compared to sulfide
ores. Affinity of phages, related to Kd, is slighly lower for sphalerite compared to other
sulfide ores (B). Affinty of phage clone Si1 towards silica is significantly higher than con-
trol, considering the lower value of Kd parameter.
* Bmax was evaluated with 95 % confidence while Kd was evaluated with 85 % confi-
dence intervals.
0
1
2
3
4
5
6
Sp1 C2 Control
Bm
ax(1
09p
fu/c
m2)
Tested phage clones
Maximum phage binding*
ChalcopyriteSphaleritePyriteSilica
1E+04
1E+05
1E+06
1E+07
Sp1 C2 Control
Kd
(p
fu/m
L)
Tested phage clones
Kd parameter for sulfide ores*
Chalcopyrite
Sphalerite
Pyrite
1E+04
1E+05
1E+06
1E+07
1E+08
Si1 Control
Kd
(p
fu/m
L)
Tested phage clones
Kd parameter for silica*

51
3.5.3. Induction time measurements
Induction times were measured to verify the effect of phage Sp1 adsorption at dif-
ferent concentrations on ore surface properties (Figure 19). Phage clone Sp1 was used be-
cause it showed comparable adsorption on each of the sulfide ores (chalcopyrite, pyrite and
sphalerite). A higher induction time indicates a lower hydrophobicity; strong hydrophobi-
city facilitates the adhesion of particles to air bubbles, thus leading to a shorter induction
time.
Some variability was notable throughout the particle bed during measurements
(mainly for short contact times). Results should therefore be interpreted more as trends (or
in a comparative way) than as absolute, precise values. As shown on Figure 19, phage addi-
tion to sulfide ores increased induction times (curves shifted to the right), especially at the
highest phage concentration, thus decreasing the particle hydrophobicity. Similar results
were reported in the literature for experiments involving phages adsorption on chalcopyrite
[9]. In our experiment on chalcopyrite (panel A), induction time was roughly equal to 5 ms
for control experiments (experiments on bare ores). Addition of phage up to 1012 pfu/mL
caused negligible effect; a concentration of 1013 pfu/mL was needed to allow observation of
a significant induction time shift of 13 ms (260 % increase) bringing the induction time to
18 ms.
Sphalerite’s induction time appeared to be less affected by phage binding. At 1013
pfu/mL phage addition, while induction time increased to 18 and 17 ms respectively for
chalcopyrite and pyrite, it only went up to 9 ms for sphalerite. However, this observation
may not be very useful for flotation since sphalerite does not normally possess native flota-
tion properties and would require considerable amount of collector with proper activation
to be floated [3]. In fact, the low induction time observed for bare sphalerite, comparable to
chalcopyrite (known for being more hydrophobic), is surprising. This could be explained
by the presence of a certain amount of copper in the sphalerite used for these experiments.
Micro-flotation tests would be required to confirm this hypothesis, but nevertheless it
doesn’t question the general depressing effect of phage addition on sulfide ores flotation
that was observed.

52
Even though direct process application might not yet be possible, these observations
show the potential of designing peptidic bio-reagents with specific effects on different ores,
such as the one seen with sphalerite or the sequence selectivity observed for silica.
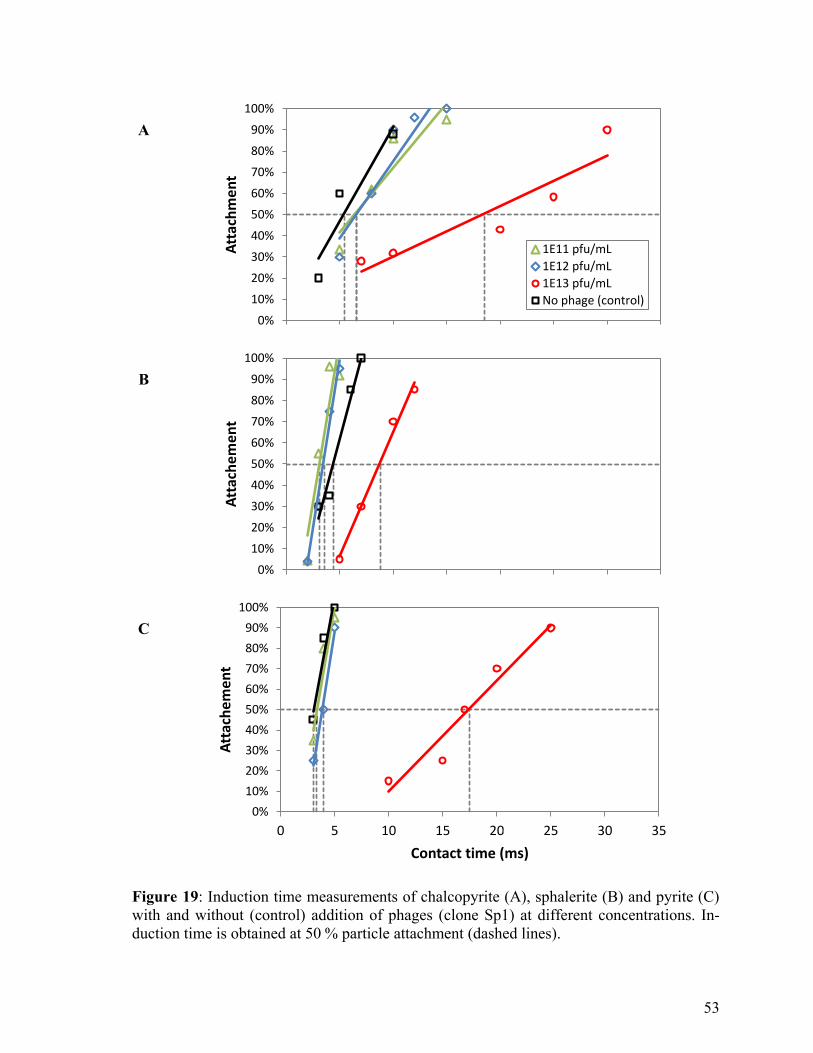
53
A
B
C
Figure 19: Induction time measurements of chalcopyrite (A), sphalerite (B) and pyrite (C)
with and without (control) addition of phages (clone Sp1) at different concentrations. In-
duction time is obtained at 50 % particle attachment (dashed lines).
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Att
ach
me
nt
1E11 pfu/mL
1E12 pfu/mL
1E13 pfu/mL
No phage (control)
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Att
ach
em
en
t
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
0 5 10 15 20 25 30 35
Att
ach
em
en
t
Contact time (ms)

54
3.6. Conclusion
As an attempt to develop novel bio-flotation agents, a phage display library was
screened, searching for specific peptide-ore interactions. Biopanning yielded consensus
sequences associated with different ores. Even though slight similarities could be noted, our
results generally differed from previous work [8, 9], implying alternative selection factors
that are yet to be thoroughly explored.
Adsorption isotherm determination showed that phages bound significantly more to
sulfide ores compared to silica, but a strong and specific affinity of the heptapeptides to
their target ore could not be confirmed through that method. Binding of the phage coat pro-
teins (present in larger number) with the ores tested may be involved in this phenomenon.
This could be the case if the specific interaction that allowed screening was weak and only
slightly superior to the binding of wild-type phage. The polysorbate concentration used for
adsorption experiments (0.25 % v/v compared to 0.75 % for final biopanning rounds) could
also be linked to the observed low specificity. More work would be required for a thorough
assessment of the affinity of isolated peptides. This could be achieved by the fusion of the
obtained sequences to the major coat protein pVIII, thus greatly increasing peptide expres-
sion. Others have had some success using a similar solution (directly using a pVIII phage
display library)[9].
The potential of using phages in bio-flotation was assessed by induction time meas-
urements. Phage adsorption on ores conferred a less hydrophobic surface, opening the pos-
sibility for the development of bio-depressants for froth flotation. Further testing must be
done to confirm this tendency in flotation conditions and to evaluate the optimal amount of
phage needed. It could also be interesting to study phage coat protein modification as a
mean to confer more or less hydrophobic properties to phage particles.
In this study, phages were studied as an option for flotation bio-reagents using
phage as an exploratory method for ligand identification. Considering that phage produc-
tion may not be viable economically at this time compared to traditional flotation reagents,
screening cheap, natural material for peptidic sequences similar to the ones identified in this
work could contribute to the development of a protein-based flotation bio-reagents.

55
CHAPITRE 4: RÉSULTATS ET ÉLÉMENTS DE DISCUSSION SUPPLÉMEN-
TAIRES


57
4.1. Expériences de bio-criblage supplémentaires
La première série d’expériences de bio-criblage a rendu des résultats intéressants,
toutefois des avenues supplémentaires ont été étudiées. Comme plusieurs séquences pepti-
diques identifiées lors de ces expériences étaient communes à plusieurs minerais (pour les
sulfures plus particulièrement) une procédure a été testée dans le but de favoriser
l’obtention de séquences avec un meilleur potentiel sur le plan de la sélectivité.
Dans l’espoir d’éliminer les séquences communes aux différents minerais le proces-
sus de criblage a été effectué en pré-incubant d'abord un échantillon de la librairie sur diffé-
rents minerais et en récupérant ceux qui n'étaient pas adsorbés. Par exemple, l’échantillon
de phages employé pour la première ronde de criblage sur la chalcopyrite a préalablement
été incubé en présence de sphalérite et de pyrite afin d’éliminer le plus possible les ligands
possédant une affinité pour ces minerais. Cette pré-incubation a été effectuée en présence
de détergent (0,5 % polysorbate Tween20) et les minerais lavés une fois afin de récupérer
autant de phages non fortement liés que possible, permettant un apport suffisant pour
l’identification de ligands sur la chalcopyrite. Les résultats de ces expériences sont présen-
tés au Tableau 5.
Après trois rondes de criblage sur la chalcopyrite (suite à la pré-incubation), tous les
clones séquencés comportaient la séquence Sp1, la même qui avait été identifiée dans les
expériences précédentes impliquant le criblage de la sphalérite et la chalcopyrite (voir Ta-
bleau 3). L’objectif de la pré-incubation était d’éliminer le plus possible les ligands de forte
affinité pour les autres sulfures, toutefois, il semble évident que l’efficacité de sélection de
cette étape n’est pas de 100 %. Dans le cas de l’expérience décrite ci-haut, il aurait suffit
qu’un seul clone Sp1 échappe à la pré-incubation pour que celui-ci soit amplifié et donc
retenu lors du processus de criblage. Cette observation renforce l’hypothèse selon laquelle
l’efficacité de sélection des séquences affinitaires aux minerais est relativement faible, tout
comme le degré d’affinité.
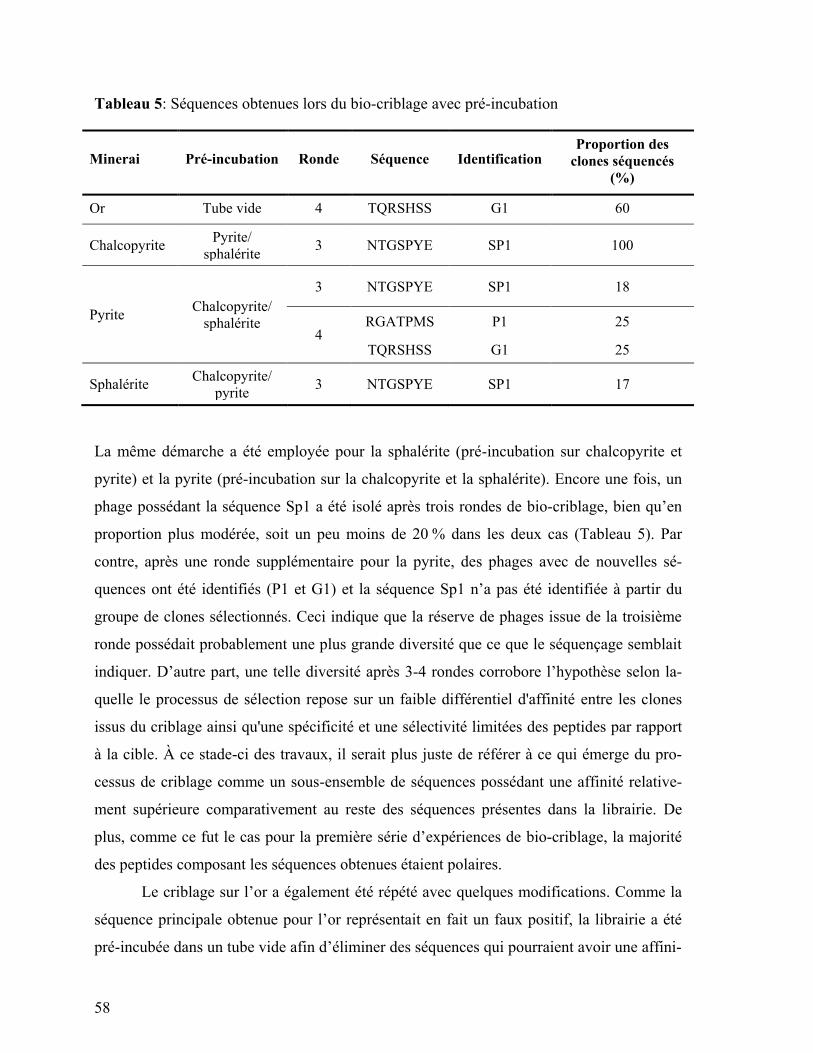
58
Tableau 5: Séquences obtenues lors du bio-criblage avec pré-incubation
Minerai Pré-incubation Ronde Séquence Identification Proportion des
clones séquencés
(%)
Or Tube vide 4 TQRSHSS G1 60
Chalcopyrite Pyrite/
sphalérite 3 NTGSPYE SP1 100
Pyrite Chalcopyrite/
sphalérite
3 NTGSPYE SP1 18
4 RGATPMS P1 25
TQRSHSS G1 25
Sphalérite Chalcopyrite/
pyrite 3 NTGSPYE SP1 17
La même démarche a été employée pour la sphalérite (pré-incubation sur chalcopyrite et
pyrite) et la pyrite (pré-incubation sur la chalcopyrite et la sphalérite). Encore une fois, un
phage possédant la séquence Sp1 a été isolé après trois rondes de bio-criblage, bien qu’en
proportion plus modérée, soit un peu moins de 20 % dans les deux cas (Tableau 5). Par
contre, après une ronde supplémentaire pour la pyrite, des phages avec de nouvelles sé-
quences ont été identifiés (P1 et G1) et la séquence Sp1 n’a pas été identifiée à partir du
groupe de clones sélectionnés. Ceci indique que la réserve de phages issue de la troisième
ronde possédait probablement une plus grande diversité que ce que le séquençage semblait
indiquer. D’autre part, une telle diversité après 3-4 rondes corrobore l’hypothèse selon la-
quelle le processus de sélection repose sur un faible différentiel d'affinité entre les clones
issus du criblage ainsi qu'une spécificité et une sélectivité limitées des peptides par rapport
à la cible. À ce stade-ci des travaux, il serait plus juste de référer à ce qui émerge du pro-
cessus de criblage comme un sous-ensemble de séquences possédant une affinité relative-
ment supérieure comparativement au reste des séquences présentes dans la librairie. De
plus, comme ce fut le cas pour la première série d’expériences de bio-criblage, la majorité
des peptides composant les séquences obtenues étaient polaires.
Le criblage sur l’or a également été répété avec quelques modifications. Comme la
séquence principale obtenue pour l’or représentait en fait un faux positif, la librairie a été
pré-incubée dans un tube vide afin d’éliminer des séquences qui pourraient avoir une affini-

59
té avec le contenant (tube micro-centrifuge). La séquence G1 (obtenue lors du contrôle né-
gatif) a tout de même été obtenue en majorité. Toutefois, des études soutiennent que le po-
lypropylène, dont sont composés les tubes ne possède pas d’affinité particulière pour les
peptides exprimés dans les librairies de phages [39, 53]. De plus, la concentration de
phages dans l’éluat récupéré pour cette expérience contrôle était à peu près constante d’une
ronde à l’autre, malgré l’accumulation de la séquence G1. Cette séquence n’aurait donc
probablement pas d’affinité pour le contenant mais pourrait être le résultat d’une amplifica-
tion non-uniforme, un phénomène qui peut survenir en l’absence de ligand d’affinité suffi-
sante [39, 54].
D’ailleurs, le phénomène d’enrichissement de clones affinitaires pourrait être simu-
lé moyennant quelques hypothèses relativement simples. En fractionnant la librairie en di-
vers groupes hypothétiques de clones ayant une affinité variable pour la cible (mais cons-
tante d’une ronde à l’autre) et en extrapolant leur amplification à partir de données expéri-
mentales (voir Tableau 6 et Tableau 7), il est possible d’estimer leur progression à travers
les rondes de bio-criblage. En supposant, par exemple, que le clone ayant la plus grande
affinité possède une efficacité de sélection aussi faible que 1 % (pourcentage des copies du
clone étant retenu lors d’une ronde de criblage), il est tout de même possible d’obtenir un
consensus de presque 40 % au bout de plusieurs rondes (Figure 20).
Tableau 6: Efficacité de sélection de différents groupes de clones
Groupe Nombre de clones Efficacité de sélection
(%)
1 1 1,00
2 2 0,75
3 10 0,50
4 100 0,25
5 1000 0,10
6 ~1,28E9 0,00005
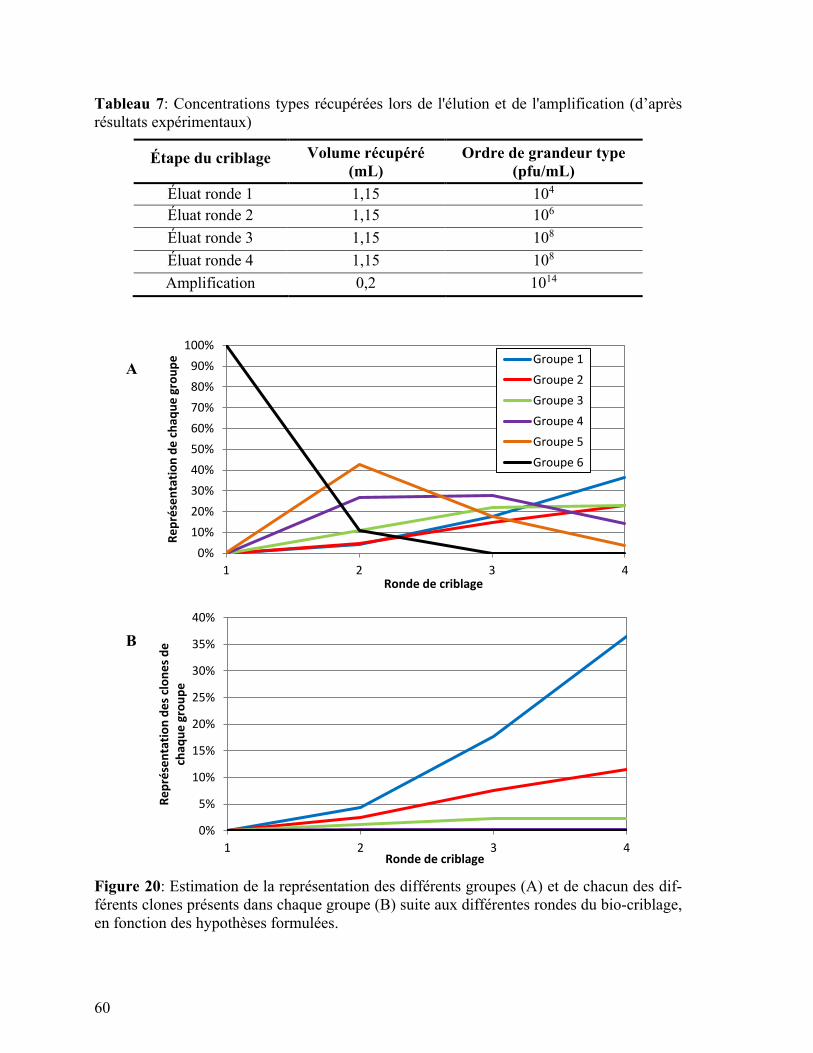
60
Tableau 7: Concentrations types récupérées lors de l'élution et de l'amplification (d’après
résultats expérimentaux)
Étape du criblage Volume récupéré
(mL)
Ordre de grandeur type
(pfu/mL)
Éluat ronde 1 1,15 104
Éluat ronde 2 1,15 106
Éluat ronde 3 1,15 108
Éluat ronde 4 1,15 108
Amplification 0,2 1014
A
B
Figure 20: Estimation de la représentation des différents groupes (A) et de chacun des dif-
férents clones présents dans chaque groupe (B) suite aux différentes rondes du bio-criblage,
en fonction des hypothèses formulées.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
1 2 3 4
Re
pré
sen
tati
on
de
ch
aqu
e g
rou
pe
Ronde de criblage
Groupe 1
Groupe 2
Groupe 3
Groupe 4
Groupe 5
Groupe 6
0%
5%
10%
15%
20%
25%
30%
35%
40%
1 2 3 4
Re
pré
sen
tati
on
de
s cl
on
es
de
ch
aqu
e g
rou
pe
Ronde de criblage

61
Suivant ce raisonnement, un échantillonnage plus étendu au moment du séquençage
pourrait permettre une meilleure analyse de la diversité des séquences issues du bio-
criblage. Ceci permettrait également de mieux suivre l’enrichissement des diverses sé-
quences à travers les rondes et d’établir un modèle plus complet se rapprochant davantage
des conditions expérimentales.
Bref, les résultats de bio-criblage avec pré-incubation et les données de simulation
suggèrent que les séquences obtenues lors du criblage ne sont pas nécessairement des li-
gands possédant une forte affinité mais font plutôt partie d’une population assez diversifiée
de phages avec une affinité qui ne pourrait être que légèrement supérieure à la moyenne.
4.2. Efficacité de la titration des phages
La méthode sir boîte de Petri a été employée pour déterminer la concentration de
bactériophages des stocks de clones individuels amplifiés, de même que pour évaluer les
concentrations dans le surnageant pour les expériences de détermination des isothermes
d’adsorption. Ainsi, les valeurs obtenues par cette méthode représentent en réalité la quanti-
té de particules virales infectieuses au sein de l’échantillon. La valeur du titre obtenu dé-
pend également de l’efficacité de la mise en culture. Quelques facteurs peuvent affecter la
capacité des phages à infecter une cellule hôte et à se mulitplier [56]. La capacité d’un
phage à produire une plage d’infection visible n’est donc pas le fait de toutes les particules
virales et la valeur obtenue lors d’un essai sur plaque sera vraisemblablement inférieure à la
quantité totale réelle de phages présents dans l’échantillon.
Cette efficacité d'infection a été vérifiée en comparant la concentration de phages
obtenue par la méthode spectrophotométrique et la méthode sur plaques (Figure 21). À par-
tir de la pente obtenue par l’application d’une régression linéaire sur les données expéri-
mentales (Tableau 8), il est possible d’estimer l’efficacité de formation de plages à 72 %
(pente=0,724). La pente calculée possède d’ailleurs une erreur type de 0,207 (près 30 % de
la valeur du paramètre) et un faible coefficient de détermination (R2) de 0,605. Il y a donc
une variabilité importante du ratio obtenu entre les deux méthodes lors de la titration
d’échantillons différents, ce qui expose une certaine incertitude liée à la comparaison de
concentrations issues de différents stocks de phages.
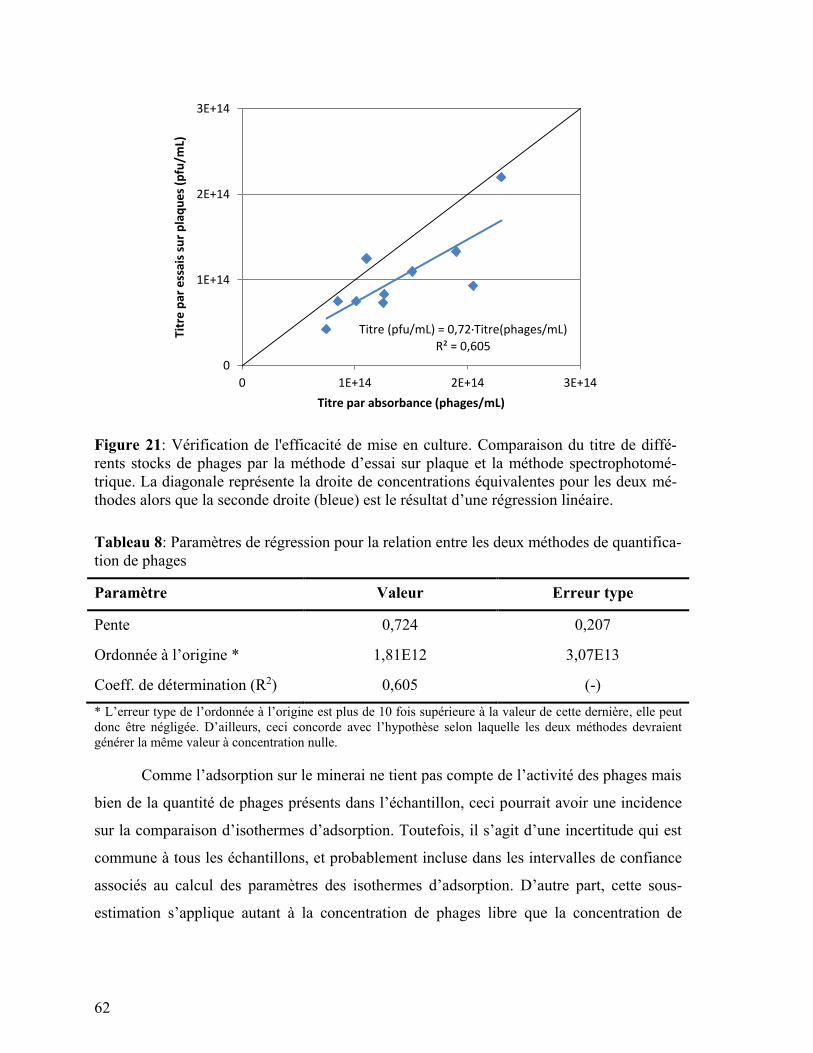
62
Figure 21: Vérification de l'efficacité de mise en culture. Comparaison du titre de diffé-
rents stocks de phages par la méthode d’essai sur plaque et la méthode spectrophotomé-
trique. La diagonale représente la droite de concentrations équivalentes pour les deux mé-
thodes alors que la seconde droite (bleue) est le résultat d’une régression linéaire.
Tableau 8: Paramètres de régression pour la relation entre les deux méthodes de quantifica-
tion de phages
Paramètre Valeur Erreur type
Pente 0,724 0,207
Ordonnée à l’origine * 1,81E12 3,07E13
Coeff. de détermination (R2) 0,605 (-)
* L’erreur type de l’ordonnée à l’origine est plus de 10 fois supérieure à la valeur de cette dernière, elle peut
donc être négligée. D’ailleurs, ceci concorde avec l’hypothèse selon laquelle les deux méthodes devraient
générer la même valeur à concentration nulle.
Comme l’adsorption sur le minerai ne tient pas compte de l’activité des phages mais
bien de la quantité de phages présents dans l’échantillon, ceci pourrait avoir une incidence
sur la comparaison d’isothermes d’adsorption. Toutefois, il s’agit d’une incertitude qui est
commune à tous les échantillons, et probablement incluse dans les intervalles de confiance
associés au calcul des paramètres des isothermes d’adsorption. D’autre part, cette sous-
estimation s’applique autant à la concentration de phages libre que la concentration de
Titre (pfu/mL) = 0,72∙Titre(phages/mL)
R² = 0,605
0
1E+14
2E+14
3E+14
0 1E+14 2E+14 3E+14
Titr
e p
ar e
ssai
s su
r p
laq
ue
s (p
fu/m
L)
Titre par absorbance (phages/mL)

63
phage liés, mesurées lors des essais d’adsorption, ce qui annule probablement l’effet global
de cette erreur lors de la comparaison d’expériences utilisant la même réserve de phages.
Il serait tout de même intéressant d’explorer d’autres méthodes de quantification qui
permettraient de diminuer cette incertitude liée à la quantification des phages. L’utilisation
de la PCR (réaction en chaîne par polymérase) quantitative a déjà été considérée [57, 58] et
pourrait représenter une solution à ce problème, tout comme l’emploi d’un cytomètre de
flux, qui a également été rapportée [59].
4.3. Calcul du coût potentiel de l'utilisation de phages comme réactif de flottation
Afin d’obtenir de plus grandes quantités de phages et de faire une évaluation préli-
minaire du potentiel de transposition à l’échelle industrielle (advenant l’emploi de phages
en tant que bio-réactifs de flottation), l’un des clones obtenu suite au criblage de la librairie
de phages a été amplifié en bioréacteur de 20 L.
Après une période de fermentation d’environ 8h, la concentration en phages montre
une stabilisation autour d’une valeur de 2,2E12 pfu/mL (Figure 22). Sachant de plus qu'un
ratio de 1013 phages par gramme de minerai pur est requis pour fournir un effet observable,
ou encore 1019 phages par tonne de minerai, un calcul sommaire du coût de cette technolo-
gie peut être fait.
Figure 22: Production du phage C1 en bioréacteur de 20 L. Axe vertical gauche: densité
cellulaire mesurée par densité optique à 600 nm, axe vertical droit: concentration pha-
gique, déterminée par titrage infectieux.
0,0
0,5
1,0
1,5
2,0
2,5
3,0
0
0,5
1
1,5
2
2,5
0 2 4 6 8 10 12
[ph
age
](1
01
2p
fu/m
L)
OD
60
0
Temps (h)
OD600[phage]

64
En faisant une évaluation sommaire (Tableau 9), en tenant compte uniquement du
coût du milieu de culture, les coûts de production peuvent être estimés à environ
3 300 $/(tonne de minerai d’intérêt). Pour des applications de bioprocédés à grande échelle,
si on estimait que le coût du milieu de culture représentait à lui seul 50 % du coût total de
production des phages [60], les coûts de production s'élèveraient alors à environ
6 600 $/(tonne de minerai d'intérêt).
Tableau 9: Coût du milieu de culture pour la production de bactériophages
Milieu LB Format
(kg) Coût ($)*
Besoin (kg/L)
Coût (k$/tonne minerai d’intérêt)
Tryptone 1 39 0,10 1,77
Extrait de levure 1 32 0,5 1,45
NaCl 1 2 0,5 0,05
Total: 3,27
* Prix obtenus de BioShop Canada pour une quantité minimale de 1000 kg
(communication électronique, 2014-09-02)
En contrepartie, le coût des réactifs de flottation traditionnels se situent générale-
ment entre 3 et 15 $/kg, et leur utilisation moyenne peut être fixée en moyenne à 20 g/tonne
de minerai d'intérêt (données fournies par COREM, Québec, Canada). Ceci correspond
donc à un coût moyen de 0,06 à 0,30 $/tonne de minerai d'intérêt. Il y aurait donc 4 ordres
de grandeur de différence entre le coût des phages et celui des réactifs traditionnels. Même
en considérant la possibilité d’économies d’échelle supplémentaires, une augmentation des
performances suite à des étapes de recherche et développement, ou avec des études de fai-
sabilité plus précises, il est peu vraisemblable que cet écart puisse être comblé.
Bref, même dans l’éventualité où des réactifs intéressants pouvaient être développés
à partir de bactériophages, il semble que les coûts de production seraient trop élevés pour
qu’un usage direct représente une solution économiquement viable. Des alternatives moins
couteuses devraient donc être envisagées afin d'amener cette technologie à un niveau com-
pétitif, comme l’identification de séquences affinitaires présentes sur des protéines déjà
abondantes dans la nature.

65
CHAPITRE 5 : CONCLUSION ET RECOMMANDATIONS


67
Dans une optique de développement de bio-réactifs de flottation, une librairie de
phages a été criblée afin d’identifier des séquences d’heptapeptides affinitaires à l’or, la
chalcopyrite, la pyrite, la sphalérite et la silice. Après plusieurs rondes de bio-criblage, di-
vers clones comportant des séquences différentes ont été identifiés comme étant potentiel-
lement affinitaires aux minerais ciblés. Quelques-unes de ces séquences ont également
montré une affinité générale pour les sulfures laissant présager une sélectivité limitée à
l'intérieur de cette classe de minerais. Par ailleurs, aucune séquence différente d’un contrôle
négatif n’a pu être obtenue pour l’or.
Des isothermes d’adsorption ont permis de quantifier plus précisément la spécificité
et le degré d'interaction des phages portant des séquences bio-criblées aux minerais sous
étude. Un écart significatif été noté entre l’adsorption sur la silice par rapport à celle ayant
été observée sur les sulfures. La capacité maximale d’adsorption (Bmax) de la silice était
environ 10 fois plus faible comparativement à la pyrite et environ 5 fois plus faible compa-
rativement à la chalcopyrite et la sphalérite. Toutefois, l’adsorption était comparable sur les
différents sulfures. Il semble y avoir une affinité légèrement inférieure des phages par rap-
port à la sphalérite, mais il a été impossible d’attribuer cet effet à l’heptapeptide isolé
puisque l’affinité est similaire au contrôle, constitué du phage d'origine naturelle, pour cha-
cun des sulfures. Ceci serait probablement dû à une affinité très faible des peptides identi-
fiés vis-à-vis la cible à l’étude. Il est également probable que la faible représentation du
peptide d’intérêt par rapport au nombre important de protéines structurales du phage M13
masque l’affinité de la séquence lors de tests d’adsorption. À cet effet, une méthode per-
mettant d’étudier l’adsorption intrinsèque du peptide plutôt que celle de l’entièreté de la
particule phagique permettrait de mieux définir l’affinité des séquences peptidiques.
L’expression du peptide sur la protéine majeure de structure du phage (PVIII) [9] ou encore
la production de protéines de fusion incluant les séquences à tester [36] sont des options qui
pourraient être considérées.
D’autre part, les séquences peptidiques identifiées diffèrent de celles déjà publiées,
probablement en raison d’une faible affinité des séquences, de facteurs de probabilité liés à
leur sélection et de différences par rapport aux échantillons de minerais. De plus,
l’obtention de séquences consensus malgré une faible différence d’affinité par rapport au
phage de type naturel (contrôle) suggère que le bio-criblage sur le minerai génère un groupe

68
très varié de séquences ayant une affinité légèrement supérieure à la moyenne. Un échantil-
lonnage plus étendu lors du bio-criblage permettrait de mieux analyser cette diversité.
Il est également possible de conclure que l’adsorption de phages rend la surface des
minerais moins hydrophobe, puisqu’elle engendre une augmentation du temps d’induction.
Les propriétés de surface de la sphalérite ont toutefois été moins altérées par l’ajout de
phages, suggérant la possibilité d’employer les phages en tant que déprimant pour la chal-
copyrite et la sphalérite, même si le phage employé n’y est pas particulièrement spécifique.
Cette possibilité devra toutefois être confirmée par des tests en conditions réelle de flotta-
tion.
Dans cette étude, le potentiel des phages en tant qu’agents de flottation a été analysé.
Considérant l’importance de la sélectivité dans les procédés de flottation, la faible diffé-
rence d’affinité observée rendrait difficile une application industrielle directe. D’autre part,
les contraintes économiques liées à la production de tels bio-réactifs en font une option peu
attrayante comparativement aux réactifs traditionnels.
Ce projet a tout de même permis de mieux connaître l’adsorption de peptides sur
des surfaces minérales. Dans le but ultime de développer des bio-réactifs de flottation re-
présentant une option intéressante pour l’industrie, il serait toutefois préférable d’étendre le
spectre des ligands étudiés. Par exemple, la découverte d’une séquence peptidique ou d’une
protéine affinitaire, existante et abondante dans la nature (un résidu industriel, par exemple)
représenterait une avancée importante dans le développement de bio-réactifs pouvant être
employés dans l’industrie minière puisque les contraintes économiques de production pour-
raient être atténuées. Aussi, le développement d'un système auto-suffisant et auto-régulable
de production de bio-agents directement sur les sites des usines minéralurgiques pourrait
être investigué à cette fin.
Il n’en demeure pas moins que les librairies d’expression phagique sont des outils
exploratoires avec de nombreux avantages, tels que la grande variabilité de ligands impli-
quée et la facilité de propagation des séquences. Les résultats présentés dans ce travail de-
meurent pertinents et la démarche décrite dans le présent ouvrage peut être considérée
comme une étape préliminaire importante dans l’exploration de ligands d’origine pepti-
diques, ouvrant la porte à de nombreuses avenues de recherche.

69
RÉFÉRENCES


71
1. Government of Canada., Mineral production - Annual statistics. 2014;
[http://sead.nrcan.gc.ca/prod-prod/ann-ann-eng.aspx] consultation: 03/2015
2. Clark, M.E., et al., Creating value through application of flotation science and
technology. Minerals Engineering, 2006. 19(6–8): p. 758-765.
3. Fuerstenau, M.C., G.J. Jameson, and R.H. Yoon, Froth flotation: a century of
innovation. 2007: Society for Mining, Metallurgy, and Exploration.
4. Government of Ontario, Current occupational exposure limits for Ontario
workplaces required under regulation 833, Ministry of Labour, 2013.
5. Huang, P., M. Cao, and Q. Liu, Using chitosan as a selective depressant in the
differential flotation of Cu–Pb sulfides. International Journal of Mineral Processing,
2012. 106–109(0): p. 8-15.
6. Vasanthakumar, B., H. Ravishankar, and S. Subramanian, A novel property of DNA
- as a bioflotation reagent in mineral processing. PLoS One, 2012. 7(7): p. e39316.
7. Dwyer, R., et al., Bioflotation and bioflocculation review: microorganisms relevant
for mineral beneficiation. Mineral Processing and Extractive Metallurgy, 2012.
121(2): p. 65-71.
8. Curtis, S.B., et al., Biomining with bacteriophage: selectivity of displayed peptides
for naturally occurring sphalerite and chalcopyrite. Biotechnol Bioeng, 2009.
102(2): p. 644-50.
9. Curtis, S.B., R.T. MacGillivray, and W.S. Dunbar, Effects of bacteriophage on the
surface properties of chalcopyrite (CuFeS₂), and phage-induced flocculation of
chalcopyrite, glacial till, and oil sands tailings. Biotechnol Bioeng, 2011. 108(7): p.
1579-90.
10. Larachi, F., GCH-2000: Technologie des procédés de séparation. 2008,
Département de génie chimique, Université Laval, p. 219-273.
11. Perry, R.H.G., D.W, Perry's chemical engineers' handbook, ed. McGraw-Hill. 1997,
NY.
12. Bulatovic, S.M., Handbook of flotation reagents: chemistry, theory and practice:
Volume 1: flotation of sulfide ores. 2007: Elsevier Science.
13. Charles Tennant, Propesct Chemicals: Xanthates. 2014;
[http://www.ctc.ca/traflesanthates.asp] consultation: 09/2014
14. Chandra, A.P. and A.R. Gerson, A review of the fundamental studies of the copper
activation mechanisms for selective flotation of the sulfide minerals, sphalerite and
pyrite. Adv Colloid Interface Sci, 2009. 145(1-2): p. 97-110.
15. Rohwerder, T., et al., Bioleaching review part A. Applied Microbiology and
Biotechnology, 2003. 63(3): p. 239-248.
16. Misra, M., K. Bukka, and S. Chen, The effect of growth medium of Thiobacillus
ferrooxidans on pyrite flotation. Minerals Engineering, 1996. 9(2): p. 157-168.
17. Misra, M., et al., Mycobacterium phlei as a flotation collector for hematite.
Transaction Society of Mining Engineers of AIME, 1994: p. 170-170.

72
18. Santhiya, D., S. Subramanian, and K.A. Natarajan, Surface chemical studies on
galena and sphalerite in the presence of Thiobacillus thiooxidans with reference to
mineral beneficiation. Minerals Engineering, 2000. 13(7): p. 747-763.
19. de Mesquita, L.M.S., F.F. Lins, and M.L. Torem, Interaction of a hydrophobic
bacterium strain in a hematite–quartz flotation system. International Journal of
Mineral Processing, 2003. 71(1–4): p. 31-44.
20. Natarajan, K.A. and A. Das, Surface chemical studies on ‘Acidithiobacillus’ group
of bacteria with reference to mineral flocculation. International Journal of Mineral
Processing, 2003. 72(1–4): p. 189-198.
21. Goffinet, G. and C. Jeuniaux, Distribution et importance quantitative de la chitine
dans les coquilles de mollusques. Cahiers Biol. Mar, 1979. 20: p. 341-349.
22. Feng, J., et al., Adsorption mechanism of single amino acid and surfactant
molecules to Au {111} surfaces in aqueous solution: design rules for metal-binding
molecules. Soft Matter, 2011. 7(5): p. 2113-2120.
23. Willett, R.L., et al., Differential adhesion of amino acids to inorganic surfaces. Proc
Natl Acad Sci U S A, 2005. 102(22): p. 7817-22.
24. Lee, S.-H. and K.B. Song, Purification of an iron-binding nona-peptide from
hydrolysates of porcine blood plasma protein. Process Biochemistry, 2009. 44(3): p.
378-381.
25. Huang, G., Z. Ren, and J. Jiang, Separation of iron-binding peptides from shrimp
processing by-products hydrolysates. Food and Bioprocess Technology, 2011. 4(8):
p. 1527-1532.
26. Mejáre, M. and L. Bülow, Metal-binding proteins and peptides in bioremediation
and phytoremediation of heavy metals. Trends in Biotechnology, 2001. 19(2): p. 67-
73.
27. Gadd, G.M., Metals, minerals and microbes: geomicrobiology and bioremediation.
Microbiology, 2010. 156(Pt 3): p. 609-43.
28. Petrenko, V.A., et al., A library of organic landscapes on filamentous phage.
Protein Eng, 1996. 9(9): p. 797-801.
29. Barbas, C.F., Phage display : a laboratory manual. 2001, Cold Spring Harbor, NY:
Cold Spring Harbor Laboratory Press.
30. Sidhu, S.S., Engineering M13 for phage display. Biomol Eng, 2001. 18(2): p. 57-63.
31. Hemminga, M.A., et al., Viruses: incredible nanomachines. New advances with
filamentous phages. Eur Biophys J, 2010. 39(4): p. 541-50.
32. Russel, M., Moving through the membrane with filamentous phages. Trends in
Microbiology, 1995. 3(6): p. 223-228.
33. Causa, F., et al., Evolutionary screening and adsorption behavior of engineered
M13 bacteriophage and derived dodecapeptide for selective decoration of gold
interfaces. J Colloid Interface Sci, 2013. 389(1): p. 220-9.

73
34. Chen, H., et al., QCM-D analysis of binding mechanism of phage particles
displaying a constrained heptapeptide with specific affinity to SiO2 and TiO2. Anal
Chem, 2006. 78(14): p. 4872-9.
35. Golec, P., et al., Novel ZnO-binding peptides obtained by the screening of a phage
display peptide library. J Nanopart Res, 2012. 14(11): p. 1218.
36. Gabryelczyk, B., et al., Selection and characterization of peptides binding to
diamond-like carbon. Colloids Surf B Biointerfaces, 2013. 110: p. 66-73.
37. Curtis, S.B., W.S. Dunbar, and R.T. MacGillivray, Bacteriophage-induced
aggregation of oil sands tailings. Biotechnol Bioeng, 2013. 110(3): p. 803-11.
38. Rumball, J.A. and G.D. Richmond, Measurement of oxidation in a base metal
flotation circuit by selective leaching with EDTA. International Journal of Mineral
Processing, 1996. 48(1–2): p. 1-20.
39. NEB, Ph.D. Phage display libraries: Instruction Manual. 2011, New England
Biolabs. p. 41 pages.
40. McLafferty, M.A., et al., M13 bacteriophage displaying disulfide-constrained
microproteins. Gene, 1993. 128(1): p. 29-36.
41. Smith, G.P. and V.A. Petrenko, Phage Display. Chem Rev, 1997. 97(2): p. 391-410.
42. Messing, J., et al., Filamentous coliphage M13 as a cloning vehicle: insertion of a
HindII fragment of the lac regulatory region in M13 replicative form in vitro. Proc
Natl Acad Sci U S A, 1977. 74(9): p. 3642-6.
43. Smith, G.P. Absorption spectroscopy and quantitation of filamentous phage. 2006;
[http://www.biosci.missouri.edu/smithgp/PhageDisplayWebsite/PhageDisplayWebs
iteIndex.html] consultation: 11/2012.
44. NEB. M13 Amplification. 2014;
[https://www.neb.com/protocols/1/01/01/m13-amplification] consultation: 2013-
2014.
45. Grieco, S.H., et al., Maximizing filamentous phage yield during computer-
controlled fermentation. Bioprocess Biosyst Eng, 2009. 32(6): p. 773-9.
46. Grieco, S.H., et al., Optimization of fermentation parameters in phage production
using response surface methodology. J Ind Microbiol Biotechnol, 2012. 39(10): p.
1515-22.
47. Lauffenburger, D.A.L.J., Receptors Models for Binding, Trafficking, and Signaling.
1996.
48. Ye, Y., S.M. Khandrika, and J.D. Miller, Induction-time measurements at a particle
bed. International Journal of Mineral Processing, 1989. 25(3–4): p. 221-240.
49. Webb, M., H. Ruber, and G. Leduc, The toxicity of various mining flotation
reagents to rainbow trout (Salmo gairdneri). Water Research, 1976. 10(4): p. 303-
306.
50. Vigneault, B., Desforges, M. and McGeer, J. Mining Reagents and By-Products (e.g.
Thiosalts) As Potential Toxicants in Mine Effluents. in Environmental Effects

74
Monitoring Investigation of Cause Workshop for Metal Mining. 2012. Gatineau
(QC), Canada.
51. Goode, P., Xanthates in mining, D.o.n.r.a. mines, Editor. 2013: State of Queensland,
Australia. p. 1-3.
52. Naik, R.R., et al., Silica-precipitating peptides isolated from a combinatorial phage
display peptide library. J Nanosci Nanotechnol, 2002. 2(1): p. 95-100.
53. Vodnik, M., et al., Phage display: selecting straws instead of a needle from a
haystack. Molecules, 2011. 16(1): p. 790-817.
54. Levitan, B., Stochastic modeling and optimization of phage display. Journal of
Molecular Biology, 1998. 277(4): p. 893-916.
55. Zeri, A.C., et al., Structure of the coat protein in fd filamentous bacteriophage
particles determined by solid-state NMR spectroscopy. Proc Natl Acad Sci U S A,
2003. 100(11): p. 6458-63.
56. Clokie, M.J., et al., Bacteriophage Plaques: Theory and Analysis, in
Bacteriophages. 2009, Humana Press. p. 161-174.
57. Ballard, V.L., J.M. Holm, and J.M. Edelberg, Quantitative PCR-based approach for
rapid phage display analysis: a foundation for high throughput vascular proteomic
profiling. Physiol Genomics, 2006. 26(3): p. 202-8.
58. Refardt, D., Real-time quantitative PCR to discriminate and quantify lambdoid
bacteriophages of Escherichia coli K-12. Bacteriophage, 2012. 2(2): p. 98-104.
59. Clokie, M.J., A. Kropinski, and C.D. Brussaard, Enumeration of Bacteriophages
Using Flow Cytometry, in Bacteriophages. 2009, Humana Press. p. 97-111.
60. Vickers, C.E., D. Klein-Marcuschamer, and J.O. Krömer, Examining the feasibility
of bulk commodity production in Escherichia coli. Biotechnol Lett, 2012. 34(4): p.
585-96.
61. Jitesh, A. Atomic Structure of Amino Acids. 2011;
[http://www.brighthub.com/science/medical/articles/110734.aspx]
consulation: 08/2014

75
ANNEXES

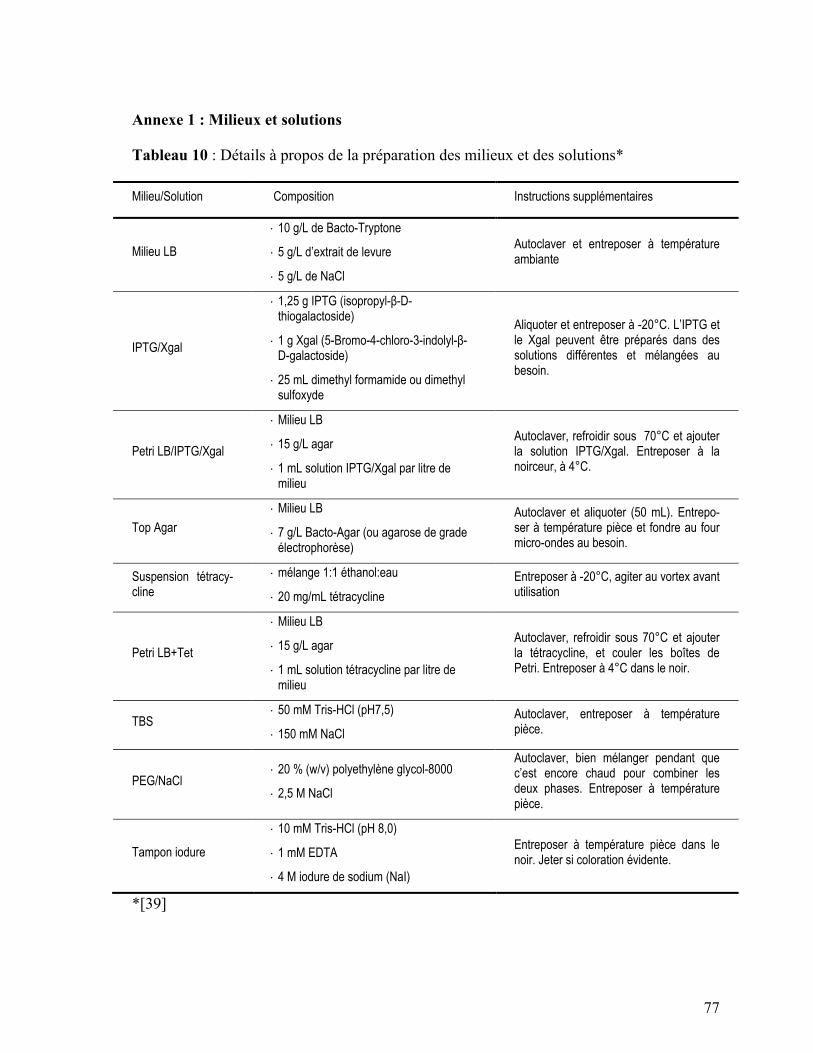
77
Annexe 1 : Milieux et solutions
Tableau 10 : Détails à propos de la préparation des milieux et des solutions*
Milieu/Solution Composition Instructions supplémentaires
Milieu LB
10 g/L de Bacto-Tryptone
5 g/L d’extrait de levure
5 g/L de NaCl
Autoclaver et entreposer à température ambiante
IPTG/Xgal
1,25 g IPTG (isopropyl-β-D-thiogalactoside)
1 g Xgal (5-Bromo-4-chloro-3-indolyl-β-D-galactoside)
25 mL dimethyl formamide ou dimethyl sulfoxyde
Aliquoter et entreposer à -20°C. L’IPTG et le Xgal peuvent être préparés dans des solutions différentes et mélangées au besoin.
Petri LB/IPTG/Xgal
Milieu LB
15 g/L agar
1 mL solution IPTG/Xgal par litre de milieu
Autoclaver, refroidir sous 70°C et ajouter la solution IPTG/Xgal. Entreposer à la noirceur, à 4°C.
Top Agar
Milieu LB
7 g/L Bacto-Agar (ou agarose de grade électrophorèse)
Autoclaver et aliquoter (50 mL). Entrepo-ser à température pièce et fondre au four micro-ondes au besoin.
Suspension tétracy-cline
mélange 1:1 éthanol:eau
20 mg/mL tétracycline
Entreposer à -20°C, agiter au vortex avant utilisation
Petri LB+Tet
Milieu LB
15 g/L agar
1 mL solution tétracycline par litre de milieu
Autoclaver, refroidir sous 70°C et ajouter la tétracycline, et couler les boîtes de Petri. Entreposer à 4°C dans le noir.
TBS 50 mM Tris-HCl (pH7,5)
150 mM NaCl
Autoclaver, entreposer à température pièce.
PEG/NaCl 20 % (w/v) polyethylène glycol-8000
2,5 M NaCl
Autoclaver, bien mélanger pendant que c’est encore chaud pour combiner les deux phases. Entreposer à température pièce.
Tampon iodure
10 mM Tris-HCl (pH 8,0)
1 mM EDTA
4 M iodure de sodium (NaI)
Entreposer à température pièce dans le noir. Jeter si coloration évidente.
*[39]


79
Annexe 2 : Protocoles détaillés
Bio-criblage
1. Pour les sulfures, nettoyer le minerai avec 50 % m/m d’une solution EDTA 0,105 M
désoxygénée. Incuber pour une heure avec agitation (150-200 rpm), rincer à
l’éthanol et entreposer les portions non-utilisées dans l’éthanol à -20°C.
2. Préparer des échantillons de 75 mg pour la première ronde et 25 mg pour les rondes
subséquentes, rincer au TBS pour éliminer l’éthanol. Suspendre l’échantillon dans 1
mL de TBST (0,1 % Tween-20 pour la première ronde, 0,5 % pour la seconde et
0,75 % pour les suivantes). Utiliser aussitôt afin d’éviter l’oxydation du minerai.
3. Ajouter 10 μL de la librairie de phages à chaque échantillon préparé (représentation
de 100x, de l’ordre d’environ 1011 phages) et incuber à la température de la pièce
pendant une heure avec légère agitation.
4. Centrifuger brièvement les échantillons et retirer le surnageant contenant les phages
non liés.
5. Rincer le culot de particules avec 1 mL de TBST (0,1 % Tween-20 pour la première
ronde, 0,5 % pour la seconde et 0,75 % pour les suivantes) à 10 reprises en centri-
fugeant à chaque fois pour retirer la solution de rinçage.
6. Procéder à l’élution des phages adsorbés avec 1 mL de 0,2 M glycine-HCl (pH 2,2)
en agitant doucement pendant 10-20 min. Centrifuger, puis récupérer l’éluat et neu-
traliser avec 150 μL de 1 M Tris-Hcl pH 9,1.
7. Titrer une petite portion de l’éluat (~1 μL) afin vérifier la quantité de phages sélec-
tionnés.
8. Amplifier le reste de l’éluat en l’ajoutant à 20 mL de milieu LB, inoculé à 1 % avec
une culture d’E. coli préalablement préparée. Incuber à 37°C pendant 4,5 heures à
forte agitation (200-250 rpm).
9. Transférer la culture dans un tube et centrifuger 10 minutes à 12 000 g à 4°C. Trans-
férer le surnageant dans un nouveau tube et centrifuger à nouveau (jeter le culot).
Titrer le ou les surnageant(s) (~10 μL) pour valider l’amplification.
10. Transférer les 80% supérieurs du surnageant dans un nouveau tube et ajouter 1/6 du
volume de PEG/NaCl. Laisser les phages précipiter à 4°C pour une nuit.
11. Centrifuger la précipitation à 12 000 g pour 15 minutes à 4°C. Décanter et jeter le
surnageant, centrifuger à nouveau brièvement et retirer le surnageant résiduel à
l’aide d’une pipette. Le culot de phages devrait ressembler à une tache blanche
(taille d’une empreinte) sur le côté du tube.

80
12. Suspendre dans 1 mL de TBS et transférer cette suspension dans un tube micro-
centrifuge et centrifuger à 14 000 rpm pour 5 minutes à 4°C pour culotter les cel-
lules restantes.
13. Transférer le surnageant dans un nouveau tube micro-centrifuge et reprécipiter en
ajoutant 1/6 du volume de PEG/NaCl. Incuber sur glace pour 15-60 minutes. Micro-
centrifuger à 14 000 rpm (~16 000 g) pour 10 minutes à 4°C, jeter le surnageant, re-
centrifuger brièvement et retirer le surnageant avec une micropipette.
14. Suspendre le culot dans 200 μL de TBS. Micro-centrifuger pendant 1 minute pour
culotter ce qui pourrait rester de matières insolubles. Transférer le surnageant dans
un nouveau tube; ceci représente l’éluat amplifié.
15. Titrer l’éluat amplifié (10 μL) sur boîtes de Petri LB/IPTG/Xgal. L’éluat peut être
conservé pendant 3 semaines à 4°C; pour une conservation prolongée, ajouter un
volume égal de glycérol stérile et entreposer à -20°C.
16. Compter les plages bleues des plaques titration et déterminer le titre phagique (de-
vrait être de l’ordre de 1013-14 pfu/mL). Calculer le volume requis pour un apport
équivalent à l’étape 1.
17. Procéder aux prochaines rondes (jusqu’à 3) en augmentant la concentration de
Tween-20 dans le TBST à 0,5 % pour la deuxième ronde, et 0,75 % pour les sui-
vantes lors des étapes de rinçage.
18. Titrer l’éluat non amplifié de la dernière ronde (pas nécessaire de l’amplifier, dilu-
tions de l’ordre de 10-1 à 10-7). Les plages de cette ronde peuvent être utilisés pour
le séquençage; assurer un temps d’incubation à 37°C d’un maximum de 18h (éviter
les délétions). À 4°C, la prise des clones individuels des plages devrait être faite
dans les 1-3 jours suivant la fin de la croissance de 18 h.
19. Pour la préparation de clones individuels pour le séquençage ou l’ELISA, préparer
une culture d’une nuit de 3 mL d’ER2738 à partir d’une colonie, ne pas diluer la
culture de titrage. Procéder à l’amplification des plages sélectionnées. Ne pas pro-
céder à des rondes supplémentaires à moins que le séquençage ne présente aucun
consensus ou que les résultats d’ELISA soient négatifs.
Titrage des bactériophages
1. Inoculer 3 mL de LB+Tet20 avec la souche d’E. coli ER2738 et incuber pour une
nuit.
2. Faire fondre le top agar au four micro-ondes et ajouter 3 mL dans des tubes de cul-
ture stériles, un tube par dilution. Maintenir les tubes à 55°C.
3. Préchauffer une boîte de Petri LB/IPTG/Xgal par dilution à 37°C pour une heure
(facultatif).

81
4. Préparer les dilutions de phages dans du milieu LB; 1 mL de volume final. Pour un
échantillon de phage amplifié, prévoir des dilutions de l’ordre de 108-1011, pour un
échantillon non amplifié, 101-104.
5. Ajouter 100 μL de la culture d’E. coli ER2738 et 100 μL de la dilution de phage au
tubes de top agar et verser sur une boîte de Petri LB/IPTG/Xgal. Agiter doucement
et tourner la boîte de Petri pour étendre l’agar uniformément.
6. Laisser refroidir 5 minutes, et incuber à l’envers à 37°C pour une nuit.
7. Compter les plages bleues sur les boîtes de Petri contenant environ 100 plages.
Amplification de clones individuels (séquençage, ELISA)
1. Diluer une culture d’une nuit d’E. coli ER2738 dans du milieu LB (1:100). Ajouter
1 mL de cette dilution à des tubes de culture, un pour chaque clone à caractériser.
Environ 15-20 clones de la troisième ronde devraient être suffisants.
2. Piquer des plages à partir d’une plaque de titrage (à l’aide d’un embout de micro-
pipette) et transférer chaque clone dans un tube de culture préparé à l’étape 1. Les
plaques de titrages utilisées ne devraient pas dater de plus de 3 jours et contenir au
maximum 100 plages (choisir des plages bien espacées).
3. Incuber à 37°C avec agitation pour 4,5-5 heures (pas plus).
4. Transférer les cultures amplifiées à des tubes micro-centrifuge et centrifuger à
14 000 rpm pour 30 secondes (voir prochaine section pour la purification en vue du
séquençage).
5. Transférer le surnageant dans un nouveau tube et recentrifuger. En utilisant une pi-
pette, transférer les 80 % supérieurs du surnageant à un nouveau tube. Ceci repré-
sente la réserve de phage amplifié et peut être entreposé à 4°C pour plusieurs se-
maines sans perte importante de titre. Pour entreposage prolongé (plusieurs
mois/années) diluer 1:1 dans du glycérol stérile et entreposer à -20°C.
Purification d’échantillons de séquençage
1. Procéder à l’amplification des clones tel que décrit dans la section précédente.
Après l’étape 4, transférer 500 μL du surnageant (contenant le phage) à un tube mi-
cro- centrifuge.
2. Ajouter 200 μL de PEG 20 % / NaCl 2,5 M. Mélanger en inversant plusieurs fois et
laisser reposer 10-20 minutes à température pièce.
3. Micro-centrifuger à 4°C et 14 000 rpm pour 10 min et jeter le surnageant (le culot
pourrait ne pas être visible).

82
4. Re-centrifuger brièvement et retirer soigneusement le surnageant restant à l’aide
d’une pipette.
5. Suspendre complètement le culot dans 100 μL de tampon iodure (iodide buffer) en
tapotant vigoureusement le tube. Ajouter 250 μL d’éthanol et incuber 10-20 min à
température pièce. (Une courte incubation à température pièce fera préférentielle-
ment précipiter l’ADN simple brin, laissant la majorité des protéines du phage en
solution.)
6. Micro-centrifuger à 4°C et 14 000 rpm pour 10 min et jeter le surnageant. Laver le
culot avec 0,5 mL d’éthanol 70 % (entreposé à -20°C), re-centrifuger, jeter le sur-
nageant et sécher brièvement le culot sous vide.
7. Suspendre le culot dans 30 μL de tampon EB ou TE (le TE peut inhiber certaines
réactions, le EB est préférable). L’eau peut être utilisée, mais ceci n’est pas recom-
mandé pour une conservation à long terme. Dans le TE, l’ADN devrait être stable à
-20°C. L’ADN peut être quantifié si désiré par électrophorèse sur gel d’agarose.
L’utilisation de 5 μL d’échantillon remis en suspension devrait donner une bande
d’intensité comparable à 0,5 μg d’ADN simple brin M13mp18 purifié (NEB
#N4040).
Détermination d’isothermes d’adsorption
1. Procéder à l’amplification et purification des clones à tester. Adapter la méthode
décrite à la section 2.1.3.; un volume de culture de 0,5-1 L peut donner suffisam-
ment de matériel pour plusieurs expériences.
2. Préparer des échantillons de 125 mg de minerai (1 pour chaque point expérimental
désiré) dans des tubes de type microcentrifuge.
3. Dans le cas des sulfures, traiter les échantillons à l’EDTA tel que décris dans la sec-
tion 2.1.2. Après le traitement, centrifuger 1 minute, retirer le surnageant, rincer le
minerai à l’eau distillé désoxygénée. Re-centrifuger et retirer le surnageant.
4. Diluer des échantillons de phages dans 500 μL de TBST (0,25 % tween, pour limi-
ter l’importance d’interactions non spécifiques) pour obtenir une plage de concen-
tration croissante. Ajouter une concentration différente à chaque tube contentant les
échantillons de minerais lavés à l’EDTA.
5. Bien agiter pour resuspendre le culot de minerai, incuber pour une heure à tempéra-
ture pièce avec agitation suffisante pour maintenir le minerai en suspension.
6. Centrifuger brièvement les échantillons, récupérer et titrer le surnageant. Connais-
sant la concentration du surnageant et la concentration initiale, la quantité de phages
adsorbés peut être calculée.

83
Annexe 3 : Résultats détaillés du bio-criblage
Les résultats détaillés du séquençage d’échantillons issus du bio-criblage sont pré-
sentés dans cette section. Chaque rangée des tableaux ci-dessous correspond à la séquence
de l’heptapeptide du clone séquencé, précédée du nom donné au clone, le cas échéant.
Code de couleur des acides aminés :
Sulfures Acides
Hydroxylique Amides
Hydrophobes Basiques
Aromatiques
Bio-criblage sur l’or
Or, ronde 3
R S A N I Y T R G A T P M S R G A T P M S S R S D S L V K N S M S P V F N M F S R V
G1 T Q R S H S S
N H T S T N R G Y S S F N R D S F F H N K
Or, ronde 4
G1
T Q R S H S S
T Q R S H S S
T Q R S H S S
T Q R S H S S
T Q R S H S S
T Q R S H S S
T Q R S H S S
C1 H V N S T K H
H V N S T K H
C2 L N S S Q P S
H Y A N S H S E M R D H A A D V S G N W
Or pré-incubation tube vide, ronde 3
H P V S G Q K S I S S L T H T G Q T A M T N G M P A K T L Q N Q G E N H N E G N R A H N T P H H L
G1 T Q R S H S S
T M N P K Y S L N G S M F T S F L N Q G V
Or pré-incubation tube vide, ronde 4
G1
T Q R S H S S
T Q R S H S S
T Q R S H S S
T Q R S H S S
T Q R S H S S
T Q R S H S S
Q Q T K N Y Y
Sp1 N T G S P Y E
S I S S L T H Y S M K Y G S
84
Bio-criblage sur la chalcopyrite
Chalcopyrite, ronde 3
C2
L N S S Q P S
L N S S Q P S
L N S S Q P S
C1 H V N S T K H
H V N S T K H
D L V R G H H
F L N P P S A
W S P F T S E
Sp1 N T G S P Y E
Chalcopyrite avec pré-incubation py-rite/sphalérite, ronde 3
Sp1
N T G S P Y E
N T G S P Y E
N T G S P Y E
N T G S P Y E
N T G S P Y E
N T G S P Y E
N T G S P Y E
N T G S P Y E
N T G S P Y E
N T G S P Y E
N T G S P Y E
N T G S P Y E
N T G S P Y E
N T G S P Y E
N T G S P Y E
N T G S P Y E

85
Bio-criblage sur la sphalérite
Sphalérite, ronde 3
T K H S G N Q
R S W P T Q Q
H S M N F A M
T M N T R N M
N A Q T H A T
Y V S K N N S
T S V L N N T
N R A P Y S Q
N N G T D K L
H P L S N A A
Sphalérite, ronde 4
Sp1
N T G S P Y E
N T G S P Y E
N T G S P Y E
N T G S P Y E
Q G V S P K L P P R T P N I
C1 H V N S T K H
L K L G E K W E N V K H S W
Sphalérite pré-incubation chalcopyrite/pyrite, ronde 3
T S K E Q S Q R G A T P M S S E G T A N M L D G R S T R N E V R S S T
Sp1 N T G S P Y E
N T G S P Y E
L H P N L Q A L V S N S I G I L T N D T F G P S Q I E A G Y S S F N R

86
Bio-criblage sur la pyrite
Pyrite, ronde 3
T K S L A H T
S G L Y A T Y
S G D V Y P M
T W L N S S V
I G N S N T L
L S N D N R A
N T N S L N S
N S S A N T Y
N P K S T L N
A D P L V G R
Pyrite ronde 4
C1
H V N S T K H
H V N S T K H
H V N S T K H
P Q Y K S D L L K A D Y K L G L K A L K E
Pyrite pré-incubation chalcopyrite/sphalérite, ronde 3
E G Q R W M Q N P V H F P R N A Q H I S H N S T G G T R N Q E I A S R S Q L P W Y S
Sp1 N T G S P Y E
N T G S P Y E
T A S H Q Y L G T A Y T K A T S P M H S S
Pyrite pré-incubation chalcopyrite/sphalérite, ronde 4
P1 R G A T P M S
R G A T P M S
S P L T A E S Q I L N T T E M S Q L G T P K T D A H R A
G1 T Q R S H S S
T Q R S H S S

87
Bio-criblage sur la silice
Silice, ronde 3
A I R T W V A
T V R T S A H
T V R T S A D
T V R P S S D
T I R I E V P
S T E H T R S
S S G S D H L
T H A S S H H
D F R H S K H
Si3 N A K H H P R
N H H K H L G
Silice, ronde 4
Si1
K H D R I H H
K H D R I H H
K H D R I H H
K H S A E R H
R H A S S H H
Si2 V H K Y K H T
V H K Y K H T
K A H P T H T
Si3 N A K H H P R
N A K H H P R
H H H F H L
H E H T H R Q
H Y A N S H S
S I S S L T H
Bio-criblage témoin négatif (tube vide)
Témoin négatif, ronde 3
G1
T Q R S H S S
T Q R S H S S
T Q R S H S S
T Q R S H S S
T Q R S H S S
T Q R S H S S
T Q R S H S S
T Q R S H S S
T Q R S H S S
T Q R S H S S
T Q R S H S S
CN1
N R W H H L E
N R W H H L E
N R W H H L E
S R D T N Y V T G L G S H V


89
Annexe 4: Abréviations et catégories d’acides aminés
Tableau 11: Acides aminés et abréviations
Acide aminé Abréviation Acide aminé Abréviation
Alanine A Leucine L
Arginine R Lysine K
Asparagine N Méthionine M
Acide aspar-
tique
D Phénylalanine F
Cystéine C Proline P
Acide Gluta-
mique
E Sérine S
Glutamine Q Thréonine T
Glycine G Tryptophane W
Histidine H Tyrosine Y
Isoleucine I Valine V
Tableau 12: Catégories et structure d'acides aminés [61]





























