Embed Size (px)
Citation preview

JOURNAL CANADIEN DE BOTANIQUE Date de publication, volume 56, 1978
N U M E R O
ler 2 3 4 5 6 7 8 9
10 l1
MOIS
janvier janvier fevrier fevrier mars mars avril avril mai mai juin juin juillet juillet aoiit aoiit septembre septembre octobre octobre novembre novembre decembre decembre
D A T E D E PUBLICATION
27janvier 1978 9 fevrier
13 mars 13 mars 22 mars 30 mars I l avril 25 avril 12 mai 23 mai 22juin 4juillet 7 juillet 2 aoht
28 aoiit I septembre
2 1 septembre 3 octobre
24 octobre 2 novembre
20 novembre 24 novembre 18 decembre 26janvier 1979
CANADIAN JOURNAL O F BOTANY Date of Issue, Volume 56, 1978
NUMBER
I 2 3 4 5 6 7 8 9
10 11 12 13 14 15 16 17 18 19 20 2 1 22 23 24
MONTH
January January February February March March April April May May June June July July August August September September October October November November December December
I S S U E D
January 27, 1978 February 9 March 13 March 13 March 22 March 30 April I I April 25 May l2 May 23 June 22 July 4 July 7 August 2 August 28 September I September 21 October 3 October 24 November 2 November 20 November 24 December 18 January 26, 1979
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Sa
nta
Cru
z (U
CSC
) on
11/
09/1
4Fo
r pe
rson
al u
se o
nly.

Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Sa
nta
Cru
z (U
CSC
) on
11/
09/1
4Fo
r pe
rson
al u
se o
nly.

Canadian Journal of Botany P~rDlisl~rtl hy T H E N A T I O N A L RESEARCH COUNCIL OF C A N A D A
Journal canadien de botanique PrrbliP ptrr LE CONSEIL N A T I O N A L DE RECHERCHES DU C A N A D A
Volume 57 Number 7 April l , 1979 Volume 57 numero 7 1 avril 1979
Bud development in Larix occidentalis. I. Growth and development of vegetative long shoot and vegetative short shoot
JOHN N. OWENS A N D MARJE MOLDER Biology Dc~ptrr t t~ir t~t , U t~ iwrs i ty of Victoritr, Victoritr. B . C . , Ctr t~t~t lo V8W 2 YZ
Received August 9, 1978
OWENS. J . N., and M. MOLDER. 1979. Bud development in L u r k occitler~tcrlis. I . Growth and development of vegetative long shoot and vegetative short shoot buds. Can. J . Bot. 57: 687-700.
Vegetative terminal long shoot buds (TLSB) and short shoot buds (SSB) were studied through- out the annual growthcycle in several trees over several years. TLSB were not totally preformed. . . The dormant TLSB consisted of bud scales enclosing some basal leaves and both were borne on a broad receptacle. Centripetal to the basal leaves, a series of axial leaf primordia was borne on the flanks of the apex. After dormancy a second series of axial leaves was initiated above those initiated before dormancy. Basal and both series of axial leaves elongated during shoot elongation as the terminal apex again initiated axial leaves, bud scales. and then basal leaves. After shoot elongation the first series of axial leaves was initiated before the TLSB became dormant in October. No dimorphism occurred between predormancy and postdol-mancy axial leaves or axial and basal leaves. Axilliary buds were initiated in the TLSB about the time of flushing. All leaves did not bearaxillary buds. All axillal-y buds rapidly initiated a series of bud scales and then entered a slower phase of butl-scale initiation and rapid apical enlargement. Leaf primordia then were initiated at the base of the apex and borne on the broad receptacle. Apices then differentiated into nxillary long shoot buds (ALSB) or SSB. ALSB developed similarly to TLSB. whereas axillary SSB initiated leaf primordia at the base of the apex and all but the last primordia to be initiated were borne on the broad receptacle. Axillary SSB were preformed but ALSB were not com- pletely preformed and both became dormant in mid-October. The apex of a short shoot lived for up to 8 to 10 years. In each successive year it passed through phases of bud-scale initiation and leaf initiation to form a dormant preformed SSB which flushed after overwintering. Annual short shoot elongation was about 1 mm. The LSB and SSB apices varied in shape and size during the year but apical zonation was similar in all apices. Loris vegetative bud development is compared with that found in othergymnosperms.
OWENS, J . N. , et M. MOLDER. 1979. Bud development in Ltrrix- occitlrr~ttrlis. I. Growth and development of vegetative long shoot and vegetative short shoot buds. Can. J . Bot. 57: 687-700.
Les bourgeons vegktatifs des mmeaux longs terminaux (TLSB) et des bourgeons des rameaux courts (SSB) d e L t r k occitler~ttrlis ont ete CtudiCs pendant toute la saison d e croissance annuelle chez plusieurs arbres pendant plusieurs annees. Les TSLB ne sont pas complttement prCformks. L e TLSB dormant consiste en des Ccailles de bour-geon enveloppant quelques feuilles basales, et les deux sont portCes sur un rkceptacle large. En position centripete par rapport aux feuilles basales, on retrouve une s k i e d e primordiums foliaires axiaux les flancs d e I'apex. Apres la dormance, une seconde serie d e feuilles axiales sont amorcees au-dessus de celles qui ont Cte amorcees avant la dormance. Lesfeuilles basales et les deux series de feuilles axialks s'allongent d u m t I'elongation d e la pousse, alors que I'apex terminal amorce encore une fois des feuilles axiales, des kcailles d e bourgeon, puis des feuilles basales. Apres I'elongation d e la pousse, la
'Supported by a research contract from the Research Division of the British Columbia Forest Service and the National Re- search Council of Canadaglant A-1982.
'Appreciation is extended to the British Columbia Forest Service, Skimikin Nursery, for collecting specimens, to Mrs.Ann Smr-eciu for technical assistance, and to Mrs. OldriskaCeska for preparation of illustrations.
0008-4026/79/070687- l4EO I .OO/O @ 1979 National Research Council of Canada/Conseil national d e recherches du Canada
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Sa
nta
Cru
z (U
CSC
) on
11/
09/1
4Fo
r pe
rson
al u
se o
nly.

CAN. J . BOT. VOL. 57, 1979
premiere serie de feuilles axiales sont amorcees avant que le TSLB entre en dormance en octobre. Les feuilles axiales amorcees avant et apres la dormance ne prisentent aucun dimorphisme, pas plus que les feuilles axiales et les feuilles basales. Les bourgeons axillaires sont amorcCs dans le TSLB h peu pres au moment du debourrement. II n'y a pas de bourgeon axillaire hchaque feuille. Tous les bourgeons axillaires amorcent rapidement une serie d'ecailles de bourgeon, puis ils entrent dans une phase ou les ecailles de bourgeon sont amorcees plus lentement et ou I'apex grossit rapidement. Les primordiums foliaires sont alors amorces h la base de I'apex et sont portes sur le large receptacle. Les apex se differencient alors en bourgeons de rameaux longs axillaires (ALSB) ou en SSB. Les ALSB se developpent comme les TLSB, tandis que chez les SSB axillaires les primordiums foliaires sont amorcCs B la base de I'apex et tous les primordiums amorces sauf les derniers sont portes sur le large receptacle. Les SSB axillaires sont preformes, mais les ALSB ne sont pas completement preformes; les deux entrent en dormance B la mi- octobre. L'apex des lameaux courts vit jusqu'i 8 B 10 ans. Chaque annie, i l passe par des phases de formation d'ecailles de bourgeon et de primordiums foliaires et forme un SSB priforme dormant qui debourre apres I'hiver. L'allongement annuel des lameaux courts est d'environ I mm. La dimension et la forme de I'apex des LSB et des SSB varient au cours de I'annee, mais la zonation apicale est identique chez tous les apex. Le developpement des bourgeons vegetatifs chez Lc~ris est compare B celui d'autre gyrnnospermes.
[Traduit par le journal]
Introduction Western larch (Lur i ,~ occide~~talis Nutt.) is the
largest of North American larches and it ranges from southeastern British Columbia, east to the foot hills of southern Alberta, and south through western Montana, northern Idaho, and the north- eastern portion of Washington and Oregon. It also occurs along the eastern slopes of the Cascade mountains in Washington and Oregon (Boe 1958; Fowells 1965; Hosie 1973). This species is being harvested in British Columbia and is being consid- ered as an important species for reforestation.
Although many studies have been made on shoot growth in woody plants (Kozlowski 1964), few studies have described vegetative bud and shoot development in Larix. Fujimoto (1978) studied shoot formation in L. leptolcpis and briefly de- scribed long shoot buds (LSB) and short shoot buds (SSB) of L. decidua and L. grnelini. Seasonal growth periodicity of long shoots was described for L. decidm (Frampton 1960) and L. laricitm (Clausen and Kozlowski 1967, 1970; Kozlowski and Clausen 19660) but no studies have compared apical changes in long and short shoots or have related these changes to the phenology of vegeta- tive bud development and shoot elongation.
In Larix, pollen cones and seed cones form by the transition of a previously vegetative apex into a reproductive apex. The purpose of this study is to describe the ontogeny of vegetative buds and apices and to relate this to lateral shoot elongation. Such information will be used in a subsequent study as a basis for determining when changes occur in developing apices to indicate that cone initiation has occurred. Such information is useful also in other studies because it delimits the true annual growth cycle of the different types of vegetative buds and makes it possible tocorrelate microscopic
developmental stages in the apices with visible ex- ternal stages of bud and shoot development.
Materials and Methods Collections were made from several 10- to 20-year-old trees
growing near sea level in the Pacific Forest Research Center aboretum in Victoria, British Columbia, in 1969, 1970, 1971. 1976, and 1978. In 1969, collections from several trees were made weekly from April 22 through July 28 and every fortnight from August 18 through December 18. In 1970, collections from several trees were made once in January and once in March and then weekly from April 9 through August 19. In 1971, collections from several trees were made weekly from May 4 through June 16 and from nine trees from June 23 through August 24. Collec- tions then were made from two trees every fortnight from Au- gust 17 through November24. In 1978, weekly collections were made from several trees from March I through March 23.
On each collection date branches were sampled from the upperone-third and the lowerone-third of the crown ofeach tree and taken to the University of Victoria where they were dis- sected and fixed. Specimens from these trees provided detailed anatomical information on bud development. In 1976, four lat- eral long shoots on each of four trees were measured weekly to relate shoot elongation with bud development.
In 1976, 1977, and 1978, collections were made also from three 50-year-old trees growing at a 390-m-elevation site 20 km north- west of Salmon Arm, British Columbia. This is the northern limit of L. occiclet~lcrlis distribution and provided specimens for comparison ofanatomy and phenology with the trees sampled in Victoria. In 1976, weekly collections were made from April 23 through November 9. In 1977, weekly collections were made from June 21 through August 31. In 1978, collections were made every fortnight from March 1 through April 26. On each col- lection date blanches were sampled from upper and lower re- gions of the crown, twigs were cut into 10-cm lengths, fixed, and shipped to the University of Victoria where buds were dis- sected.
On all collection dates, several terminal long shoot buds (TLSB), newly initiated axillary long shoot buds (ALSB), newly initiated axillary short shoot buds (SSB), and SSB on l-year-old short shoots were collected and dissected by slicing along both sides toallow better penetration of the fixative. Periodically SSB from 2-year-old and older short shoots also were dissected and fixed. In 1969, 1970, and 1971, all specimens were fixed in formalin - acetic acid - alcohol (FAA), and in 1976, 1977, and
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Sa
nta
Cru
z (U
CSC
) on
11/
09/1
4Fo
r pe
rson
al u
se o
nly.

OWENS A N D MOLDER 689
1978, all specimens were fixed in Navashin's Craf solution (Sass 1958). Fixed specimens were dehydrated in a tertiary butyl alcohol series (Johansen 1940) and embedded in Paraplast. Se- rial longitudinal sections of buds were cut at 6pm and stained with safranin and hematoxylin. Median sections were selected, projected using a microprojector, and traced. Apical height and width were measured from tracings and apical zonation was noted on the tracings while viewing the slides through a com- pound microscope.
Although bud development occurred over a shorter time in trees near Salmon Arm than in those from Victoria, the different bud types passed through the same stages of development. Consequently, all graphs are based on apices of a particular type which were at a specific stage and the time sequence is an average of that which occurred in Victoriaover the several years of this study. Measurements were taken from 137 apices of which 33 were TLSB, 34 were newly initiated axillary SSB, 6 were newly initiated ALSB, 31 were SSB on l-year-old short shoots, 29 were SSB on 2-year-old or older short shoots. and 4 were TLSB which had become SSB. Numerous other buds were observed but were not included because of poor fixation, sec- tions being off angle, or because the buds had differentiated in reproductive buds. The latter aspect will be described in a subsequent paper.
Buds of all types were dissected periodically, and unfixed or fixed buds and apices were observed with a JEOL-35U scanning electron microscope operating at 10 kV. Some specimens for the scanning electron microscope were fixed in Zirkle-Erliki fluid (Conn et NI. 1960), dehydrated in a graded series of ethanol solutions to pentyl acetate, dried using a carbon dioxide critical-point dryer, and coated with gold. The number of leaf primordia present at various stages of development was deter- mined by counts of fresh buds or buds fixed for the scanning electron microscope.
Observations In L. occidentalis, as in other species of Lcrrix-,
the terminal shoot and usually the distal one to three lateral shoots on each vigorous branch are long shoots. Less vigorous branches may have no lateral long shoots and occasionally no terminal long shoot. In both instances these are replaced by short shoots or reproductive buds. Long shoots sampled bore about 100 helically arranged leaves of which the proximal one third (basal leaves) were separated by very short internodes and appeared as a cluster and the distal two thirds (axial leaves) were separated by measurable internodes (Fig. 3). During shoot elongation a bud developed in the axil of many of the distal leaves. Axillary buds occurred in groups of two or three separated by 1 or 2cm where there were no axillary buds. Usually one to three of the most distal axillary buds developed into LSB, whereas the remaining basal axillary buds developed into SSB. Externally these were indis- tinguishable during the 1st year (Fig. 4), but they became distinct after they flushed in the spring of the 2nd year (Fig. 3). Vegetcrtiue Terminal Long Shoots
Externally, dormant TLSB were small, rounded,
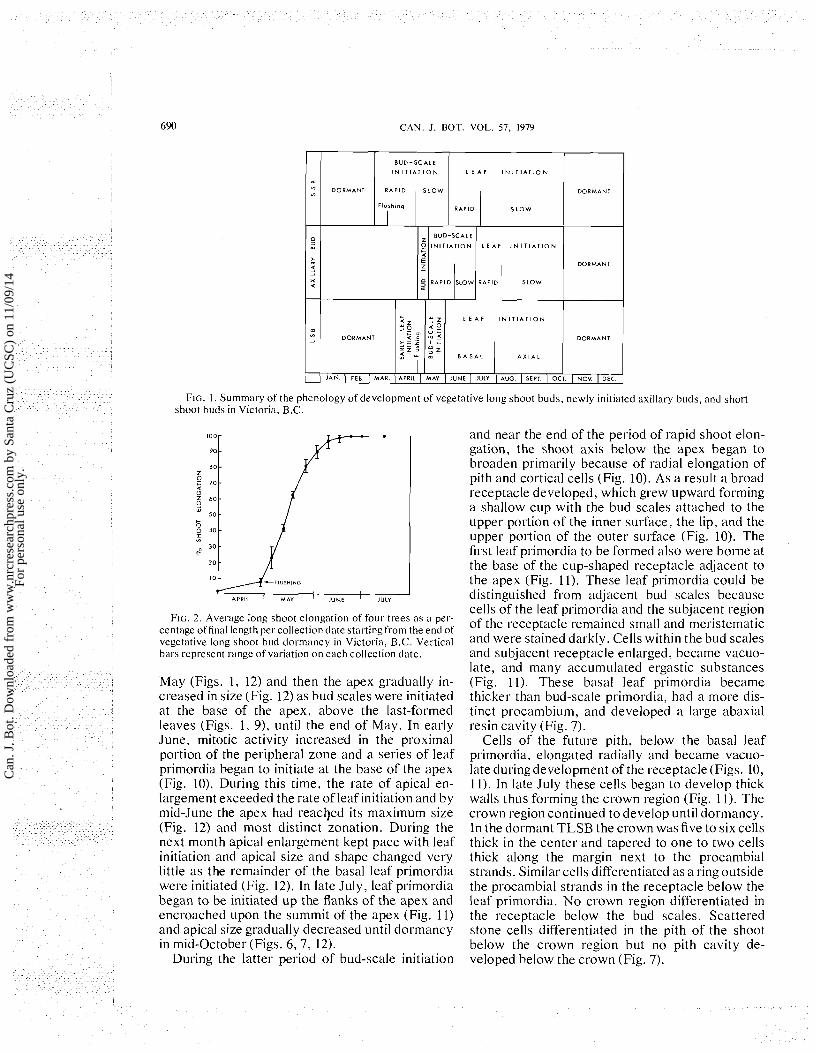
and had brown, glabrous, nonresinous bud scales (Fig. 4). In 1976, mitotic activity in leaf primordia indicated that dormancy of TLSB ended in late March at the Victoria site and about 3 weeks later at the Salmon Arm site. Divisions began in the apex about 1 week later. Measurable shoot elongation began within the bud about 2 weeks after TLSB dormancy ended (Fig. 1). During April shoot growth was slow but leaf elongation was rapid. Consequently, leaf elongation was primarily re- sponsible for causing the buds to swell and flush in late April. Leaf elongation was completed before shoot elongation. This was followed by about 1 month of rapid shoot elongation, during which shoots reached about 90% of their final length. Shoot elongation again slowed for l to 3 weeks until elongation was completed by middle to late June (Fig. 2).
Internally, the TLSB apex varied in size and shape during the growing season but retained es- sentially the same cytohistological zonation at all stages. The zonation and terminology used here (Figs. 6, 8) are similar to that first described for Ginkgo (Foster 1938). At the summit of the apex there was a layer of apical initials that were usually slightly enlarged and underwent infrequent divi- sion. Directly below was a cup-shaped zone of en- larged central mother cells. In this study the two zones were combined and discussed as an apical zone. Central mother cells divided infrequently in all planes, giving rise to the pith rib meristem di- rectly below and lateral to the peripheral zone. The pith rib meristem and the peripheral zone consisted of small actively dividing cells during the growing season which gave rise to the pith and lateral ap- pendages respectively. The surface layer of the peripheral zone, the protoderm, divided both an- ticlinally and periclinally .
Dormant TLSB apices were broadly dome shaped and the width was two to three times the height (Figs. 5, 6, 7). The outer third of the total number of leaf primordia were long, overarched the apex, and were basal leaf primordia borne on a broad, cup-shaped receptacle just above the bud scales. The next third of the leaf primordia were predormancy axial leaf primordia and were borne on the narrower conical axis of the bud (Figs. 5,7).
When apical growth resumed in late March, api- cal size did not change for about 3 weeks (Fig. 12). Then leaf primordia were initiated up the flanks of the apex decreasing its size (Figs. 8, 12). This last set of leaf primordia elongated and developed into mature leaves as the shoot elongated. This post- dormancy period of leaf initiation ended in early
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Sa
nta
Cru
z (U
CSC
) on
11/
09/1
4Fo
r pe
rson
al u
se o
nly.
CAN. J . BOT. VOL. 57, 1979
BUD-SCALE
l N l l l A l l O N L E A F l N l T l A T l O N
Flu,hing RAPID S L O W
0 BUD-SCALE 3 m p l N l r l A r l O N L E A F I N l T l A r l O N -
4 > ,. 4 z DORMANT
> - X 4 4 RAPID SLOW RAPID SLOW
m
l I I I I I I 1 J A N 1 FE0 1 MAR. 1 APRIL ) MAY 1 JUNE 1 JULY ) AUG I SEPT I OCT. I NOV. I DEC
FIG. I . Summary of the phenology of development of vegetative long shoot buds, newly initiated axillasy buds, and short shoot buds in Victoria, B.C.
FIG. 2. Aver-age long shoot elongation of four trees as a per- centage offinal length per collection date startingfrom the end of vegetative long shoot bud dormancy in Victoria, B.C. Vertical bals represent range of variation on each collection date.
May (Figs. 1, 12) and then the apex gradually in- creased in size (Fig. 12) as bud scales were initiated at the base of the apex, above the last-formed leaves (Figs. 1, 9), until the end of May. In early June, mitotic activity increased in the proximal portion of the peripheral zone and a series of leaf primordia began to initiate at the base of the apex (Fig. 10). During this time, the rate of apical en- largement exceeded the rate of leaf initiation and,by midJune the apex had reached its maximum size (Fig. 12) and most distinct zonation. During the next month apical enlargement kept pace with leaf initiation and apical size and shape changed very little as the remainder of the basal leaf primordia were initiated (Fig. 12). In late July, leaf primordia began to be initiated up the flanks of the apex and encroached upon the summit of the apex (Fig. 11) and apical size gradually decreased until dormancy in mid-October (Figs. 6 ,7 , 12).
During the latter period of bud-scale initiation
and near the end of the period of rapid shoot elon- gation, the shoot axis below the apex began to broaden primarily because of radial elongation of pith and cortical cells (Fig. 10). As a result a broad receptacle developed, which grew upward forming a shallow cup with the bud scales attached to the upper portion of the inner surface, the lip, and the upper portion of the outer surface (Fig. 10). The first leaf ~rimordia to be formed also were borne at the base'of the cup-shaped receptacle adjacent to the apex (Fig. 11). These leaf primordia could be distinguished from adjacent bud scales because cells of the leaf primordia and the subjacent region of the receptacle remained small and meristematic and were stained darkly. Cells within the bud scales and subjacent receptacle enlarged, became vacuo- late, and many accumulated ergastic substances (Fig. 11). These basal leaf primordia became thicker than bud-scale ~rimordia. had a more dis- tinct procambium, and developed a large abaxial resin cavity (Fig. 7).
Cells of the future pith, below the basal leaf primordia, elongated radially and became vacuo- late during development of the receptacle (Figs. 10, 1 1). In late July these cells began to develop thick walls thus forming the crown region (Fig. 11). The crown region continued to develop until dormancy. In the dormant TLSB the crown was five to six cells thick in the center and tapered to one to two cells thick along the margin next to the procambial strands. Similar cells differentiated as a ring outside the procambial strands in the receptacle below the leaf primordia. No crown region differentiated in the receptacle below the bud scales. Scattered stone cells differentiated in the pith of the shoot below the crown region but no pith cavity de- veloped below the crown (Fig. 7).
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Sa
nta
Cru
z (U
CSC
) on
11/
09/1
4Fo
r pe
rson
al u
se o
nly.

OWENS AND MOLDER 69 1
FIG. 3. Branch showing long shoots with basal leaves (BL) and axial leaves (AL), some of which bear axillary buds (AB). One-year-old long shoots bearing short shoots (SS). X 0.66. FIG. 4. Enlarged distal end of long shoot showing terminal long shoot bud (TLSB), axillary long shoot buds (LSB), and axillary short shoot buds (SSB). X 4. FIG. 5. Scanning electron micrograph of a dormant TLSB with bud scales removed showing larger basal and smaller predormancy axial leaf primordia and the apex (A). x 90.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Sa
nta
Cru
z (U
CSC
) on
11/
09/1
4Fo
r pe
rson
al u
se o
nly.

692 CAN. J BOT. VOL 57, 1979
Asi1l~i.y Biid Iiliticrtioi7 cirzct Deuelopinent Axillary bud initiation began in the TLSB in late
Apsil, about the time of flushing, and continued for about 2 weeks. No axillary buds were initiated in the axils of the basal leaves. The first axillary buds were initiated in the axils of the proximal predor- mancy axial leaves. Initiation proceeded acropet- ally ~ ~ n t i l the last axillary buds were initiated in early May in the axils of the most distal post- dormancy leaves just below the bud scales (Figs. 9, 15).
Axillary buds were not initiated in the axils of all elongating leaves (Figs. 3, 4). Axillary buds were distributed equally around, but not along, the stem and showed no dorsoventral relationship. Just dis- tal to the basal leaves, there was little internode elongation and several buds occurred in a cluster. More distally, internode elongation was greater and buds were initiated in groups of two or three in a cluster and each group was separated by l to 2 cm where there were no axillary buds (Figs. 3,4). Most axillary buds which were initiated developed, and abortion of young axillary buds was rare.
Axillary buds arose as small protuberances of meristematic cells frorn the developing cortex in the leaf axils (Figs. 9, 15). Within 2 to 3 weeks all apices had enlarged enough for zonation, similar to that of TLSB apices, to become evident (Fig. 16). The first two prophylls were initiated oppositely but subsequent bud-scale primordia were initiated spirally. Early bud-scale initiation was rapid but from mid-June until early July bud-scale initiation was slower and apices increased in height and width (Figs. 1, 13). Frequent cell divisions, parallel to the bud axis, and subsequent radial cell elonga- tion resulted in very broad flanks at the base of the apex and a broad receptacle at the base of the bud (Fig. 17). All axillary buds initiated about 20 to 25 bud scales, which was fewer than on the TLSB. Bud-scale initiation on axillary buds ended in mid- June (Figs. l , 13).
Leaf initiation in the new axillary buds began in early July, about 4 to 5 weeks later than in the TLSB (Figs. 1, 13). Until the middle of this period of rapid leaf initiation the two types of axillary buds were still indistinguishable (Figs. 13, 18) and possi- bly ~~ndetermined as to their pathway of develop- ment. By late July, at the end of rapid leaf initiation, one to three of the more distal apices increased more in height and width than did other apices, and leaf primordia began to be initiated up the flanks instead of only at the base of the apex. These apices differentiated into ALSB and during later stages of development were similar in size and shape to the TLSB (Figs. 11, 13). Other distal and all proximal
apices became broadly conical (Fig. 15) and differ- entiated into axillary SSB (Figs. 19,20).
Axillary SSB development was different in that the apex remained as a broad low cone, and leaf primordia were initiated at the base of the apex (Figs. 18, 19). Only the last few primordia were initiated on the flanks of the apex (Fig. 11). Most leaf primordia in SSB were borne on a continuousl y broadening cup-shaped receptacle so that the bases of early leaf primordia often were above the summit of the apex (Fig. 19). Leaf primordia were initiated rapidly for the 1st month and then more slowly until early October when cells in the apex stopped di- viding and 20 to 25 leaf primordia were present. Cells continued to divide in the leaf primordia until mid-October when SSB became dormant. By the time the SSB were dormant, most of the leaf primor-dia had elongated to about 1 mm and were erect with the tips incurved slightly, enclosing the apex (Fig. 20). The outer first-formed leaf primor- dia had a distinct procambial strand which ex- tended from the bud axis nearly to the tip of the primordium (Fig. 19), and two resin ducts, one in each margin, which extended frorn just below the tip of the leaf primordium to the base but did not continue into the shoot axis. Future mesophyll and most protoderm cells contained abundant ergastic substances. The inner younger leaf primordia were shorter (Fig. 20), contained few ergastic com- pounds, and lacked a distinct procambium and resin ducts (Fig. 19). No abscission region was evident in dormant leaf primordia but this began to develop when leaves elongated in the spring. The abscission region became well defined as a band of small, less differentiated protoderm and mesophyll cells when short shoots flushed in early April. As a result of intercalary elongation of leaves, the abscission region developed about 1 mm above the leaf axil and when leaves abscised, distinct leaf bases remained on the short shoot.
Development of the crown region in axillary SSB apices was similar to that in TLSB and ALSB apices but began 1 month later. Cells of the future pith, below the basal leaf primordia, elongated and became vacuolate during development of the re- ceptacle (Fig. 17). In late August, four to five layers of these cells began to develop thick cell walls forming the crown region. The crown region con- tinued to develop until the buds became dormant. In the dormant axillary SSB the crown was five to six cells thick in the center and tapered to one to two cells thick at the periphery of the disc-shaped crown region. Similar cells differentiated as a ring outside the procambial strands, below the outer leaf primordia, but no crown differentiated in the
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Sa
nta
Cru
z (U
CSC
) on
11/
09/1
4Fo
r pe
rson
al u
se o
nly.
OWENS AND MOLDER 693
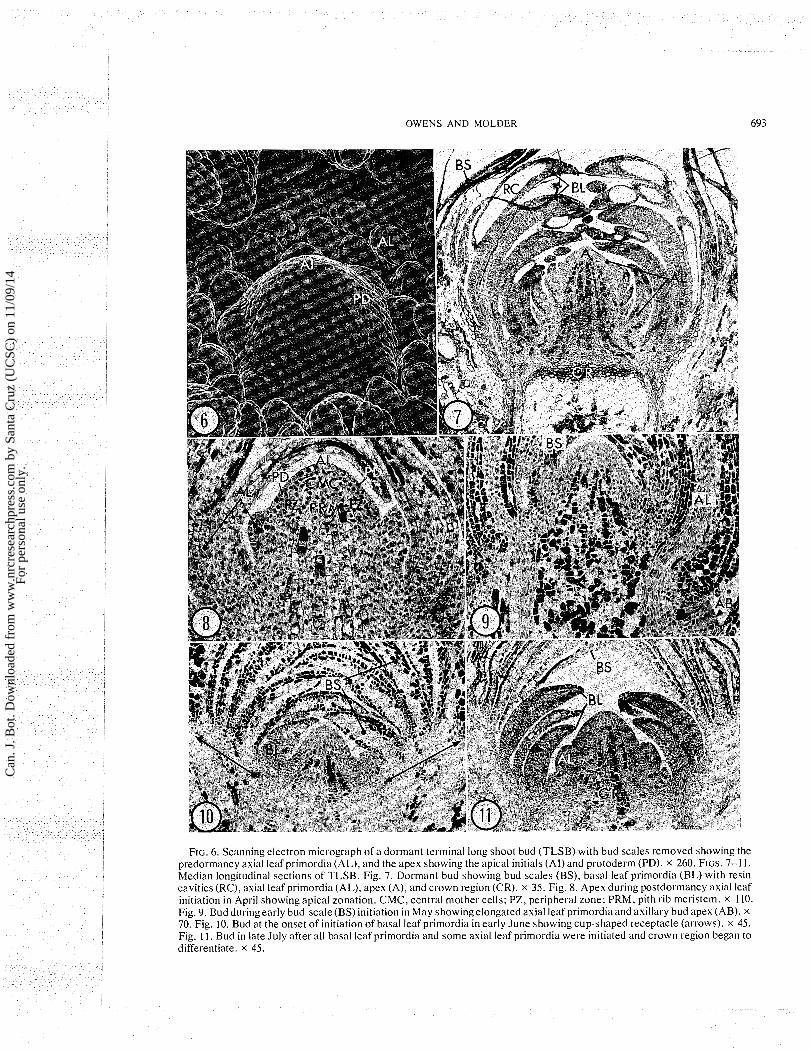
FIG. 6. Scanning electron micrograph of a dormant terminal long shoot bud (TLSB) with bud scales removed showing the predormancy axial leaf primordia (AL), and the apex showing the apical initials (AI) and protoderm (PD). X 260. FIGS. 7-1 1. Median longitudinal sections of TLSB. Fig. 7. Dormant bud showing bud scales (BS), basal leaf primordia (BL) with resin cavities (RC), axial leaf primordia (AL), apex (A), and crown region (CR). X 35. Fig. 8. Apex during postdormancy axial leaf initiation in April showing apical zonation. CMC, central mother cells; PZ, peripheral zone; PRM, pith rib meristem. X 110. Fig. 9. Bud duringearly bud-scale (BS) initiation in May showing elongated axial leaf primordia and axillary bud apex (AB). X
70. Fig. 10. Bud at the onset of initiation of basal leaf primordia in early June showing cup-shaped receptacle (arrows). x 45. Fig. 11. Bud in late July after all basal leaf primordia and some axial leaf primordia were initiated and crown region began to differentiate. X 45.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Sa
nta
Cru
z (U
CSC
) on
11/
09/1
4Fo
r pe
rson
al u
se o
nly.
694 CAN. J . BOT. VOL. 57, 1979
l I MAY I J
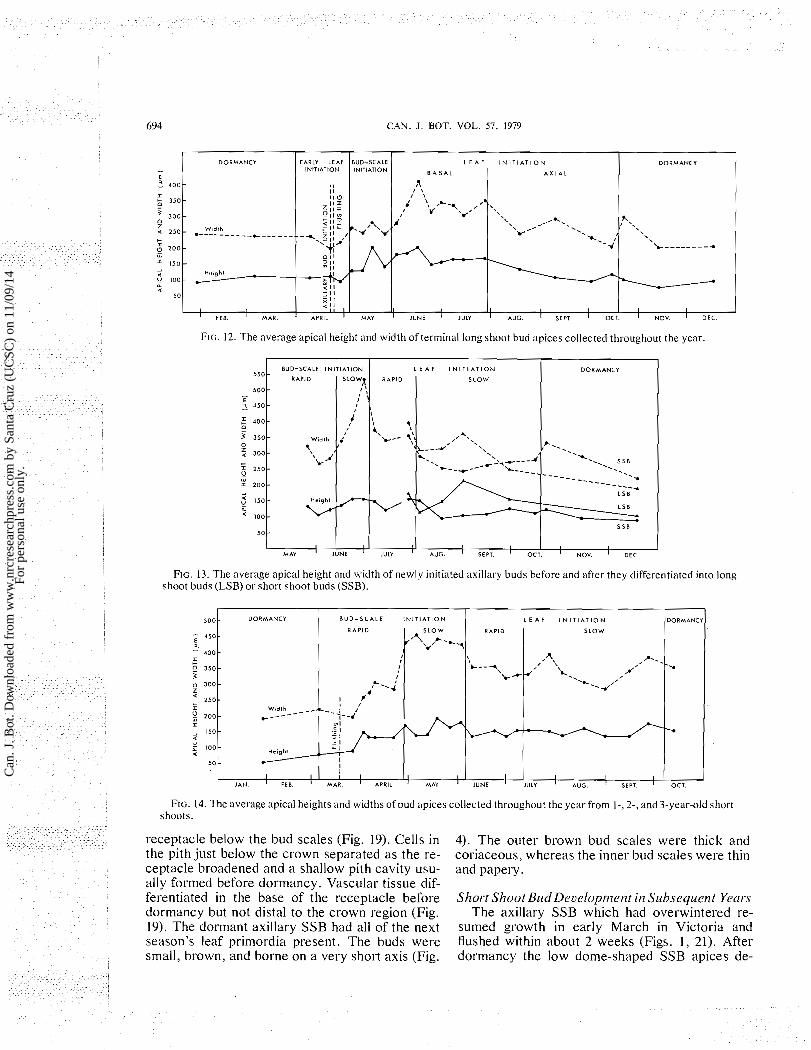
FIG. 13. The average apical height and width of newly initiated axillary buds before and after they differentiated into long shoot buds (LSB) or short shoot buds (SSB).
5 0 0 - DORMANCY B U D - S C A L E I N I T I A T I O N L E A F I N I T I A T I O N
- 4 5 0 - R A P I D S L O W RAPID S L O W
X ,*? *- - 4 0 0 -
\,' C-,
I I
! 3 5 0 - I L - - 1 ,4\
I \
P-.
3 , \
I
D 3 0 0 - b*4 4. /'
z P--.,' -. ._ ' I -. ./'
< 2 5 0 - X
1 7*
2 0 0 - X
L 150- < : 1 0 0 -
5 0 - l
I I f I I
J A N FEB. I MAR. APRIL I I I
MAY I JUNE I I I
JULY I AUG. SEPT I
I
FIG. 14. The average apical heights and widths of bud apices collected throughout the year from I-, 2-, and 3-year-old short shoots.
receptacle below the bud scales (Fig. 19). Cells in 4). The outer brown bud scales were thick and the pith just below the crown separated as the re- coriaceous, whereas the inner bud scales were thin ceptacle broadened and a shallow pith cavity usu- and papery. ally formed before dormancy. Vascular tissue dif- ferentiated in the base of the receptacle before Short Shoot BsdDeuelop~nent in Subsequent Yerirl dormancy but not distal to the crown region (Fig. The axillary SSB which had overwintered re- 19). The dormant axillary SSB had all of the next sumed growth in early March in Victoria and season's leaf primordia present. The buds were flushed within about 2 weeks (Figs. 1, 21). After small, brown, and borne on a very short axis (Fig. dormancy the low dome-shaped SSB apices de-
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Sa
nta
Cru
z (U
CSC
) on
11/
09/1
4Fo
r pe
rson
al u
se o
nly.
OWENS AND MOLDER 695
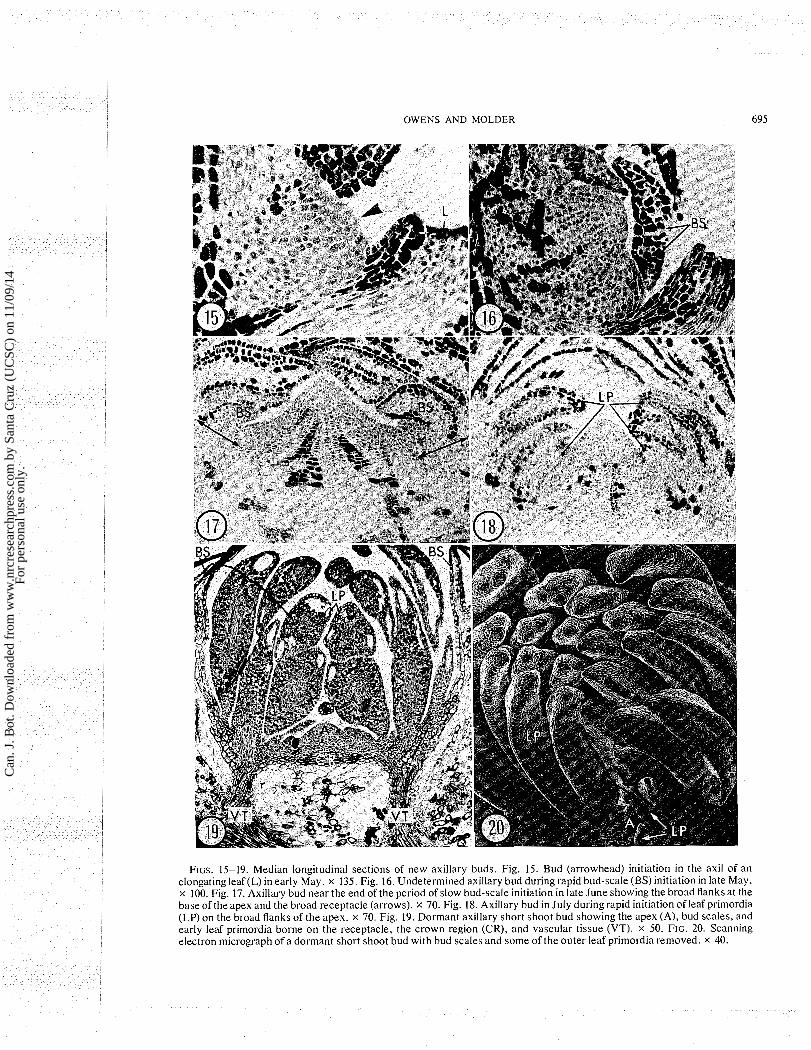
FIGS. 15-19. Median longitudinal sections of new axillary buds. Fig. 15. Bud (arrowhead) initiation in the axil of an elongating leaf(L) in early May. X 135. Fig. 16. Undetermined axillary bud during rapid bud-scale (BS) initiation in late May. X 100. Fig. 17. Axillary bud near the end of the period of slow bud-scale initiation in late June showing the broad flanks at the base of the apex and the broad receptacle (arrows). X 70. Fig. 18. Axillary bud in July during rapid initiation of leaf primordia (LP) on the broad flanks of the apex. X 70. Fig. 19. Dormant axillary short shoot bud showing the apex (A), bud scales, and early leaf primordia borne on the receptacle, the crown region (CR), and vascular tissue (VT). X 50. FIG. 20. Scanning electron micrograph of a dormant short shoot bud with bud scales and some of the outer leaf primordia removed. x 40.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Sa
nta
Cru
z (U
CSC
) on
11/
09/1
4Fo
r pe
rson
al u
se o
nly.

696 CAN. J . B 0 7
creased slightly in width as the first bud scales were initiated, then they increased both in height and width as later bud scales were initiated (Fig. 14). In early March, short shoot apices began a phase of rapid bud-scale initiation (Figs. 1, 14) and at flush- ing most bud scales had been initiated (Fig. 21). In late April the rate of bud-scale initiation slowed, apical zonation became more distinct, and the apex broadened considerably at the base (Fig. 14) be- cause of cell divisions and cell enlargement in the future pith and especially in the peripheral zone. As a result, the conical apex developed a broad, flat- tened, basal peripheral zone (Fig. 22). Leaf initia- tion began in early June and several leaf primordia were initiated rapidly, centripetally on the flattened peripheral zone (Figs. 23, 24). The broad base below the new leafprimordia then grew upward as a continuation of the receptacle on which the bud scales were borne (Fig. 24). This early period of rapid leaf initiation reduced the apical width (Figs. 14,22-24). A period of slow leaf initiation followed from mid-July until early October. From midJuly until the end of August apical enlargement kept pace with the rate of leaf initiation and minimal apical change was observed (Fig. 14). From Sep- tember through mid-October the apex broadened and, shortly before dormancy, leaf primordia were no longer being initiated but the apex continued to enlarge and became a broad dome (Fig. 25). Dor- mant SSB had all of the next season's leaf primordia present.
Throughout the period of leaf initiation the re- ceptacle grew upward and all but the last leaf primordia to be initiated were borne on the deep cup-shaped receptacle, the rim of which was equal to, or higher than, the summit of the apex (Fig. 25). In mid-August the radially elongated cells just below the apex began to develop thick walls. By September these cells had differentiated into a dis- tinct concave crown region four to five cells thick at the periphery adjacent to the procambial strands (Fig. 25). Many of the enlarged pith cells below the crown differentiated into sclereids and others sepa- rated or broke down forming a small pith cavity (Fig. 25). During the 2nd year of development, cells of the 1st year's crown region became stretched radially and formed a thin layer over the lower
VOL. 57, 1979
surface of the pith which protruded into the subja- cent pith cavity (Fig. 25).
Anatomy and phenology of successive SSB pro- duced on any one short shoot system were essen- tially similar every year (Figs. 19-23). The SSB on older short shoots generally broke dormancy and flushed a few days before younger SSB. Dormancy ended in early March and each SSB underwent a period of rapid and then slow bud-scale initiation followed by a period of leaf initiation and then dormancy. The phenology of these events was the same as that described for SSB on l-year-old short shoots. Each year a crown region developed which, by the next spring, protruded into the subjacent pith cavity (Fig. 25). Consequently, the age of a short shoot could be determined by counting the number of crown regions in longitudinal sections (Fig. 25) or more simply by counting the rings of leaf bases left each year on short shoots after leaf abscission (Fig. 26).
Apices of successive SSB produced about equal numbers of bud scales and leaves each year for 4 or 5 years then they gradually became less active and produced fewer bud scales and leaves. About 10% of the short shoots died when they were 4or 5 years old, whereas most of the remaining short shoots lived for 8 to 10 years (Fig. 26) depending upon their position on the branch and on the tree. The dead short shoots did not abscise and periderm con- tinued to form around the base. Most of the short shoots produced SSB throughout their life but some 1- and 2-year-old short shoots differentiated into pollen-cone or seed-cone buds. This will be described in a subsequent study.
Discussion The patterns of vegetative bud development and
shoot growth in Lnrix are more complex than those of most other western conifers. Lnrix produces vegetative short shoots and terminal and axillary long shoots. SSB are totally preformed, whereas LSB are not. The phenology of axillary SSB de- velopment differs from that of SSB produced on short shoots and neither is similar to that of the LSB. This is further complicated by the occasional formation of LSB on short shoots and terminal SSB on older branches. This presents many intriguing
FIGS. 21-25. Median longitudinal sections of short shoot buds on older short shoots. Fig. 21. Bud at flushing in March showing elongating leaves (L), new bud scales (BS), pith cavity (PC), and crown region (CR). x 70. Fig. 22. Bud during slow bud-scale initiation in May showing the bud scales and the flattened basal peripheral zone (arrows). X 70. Fig. 23. Bud showing early rapid initiation of leaf primordia (LP) from the basal peripheral zone in June. X 70. Fig. 24. Bud in July after early rapid leaf initiation. x 70. Fig. 25. Dormant bud on 2-year-old short shoot showing bud scales, leaf primordia, apex, crown regions, and pith cavities (PC). X 35. Fig. 26. Eight-year-old short shoot showing leaf bases (LB) and, at the distal end, elongated leaves of the current year. X 4.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Sa
nta
Cru
z (U
CSC
) on
11/
09/1
4Fo
r pe
rson
al u
se o
nly.

OWENS AND MOLDER
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Sa
nta
Cru
z (U
CSC
) on
11/
09/1
4Fo
r pe
rson
al u
se o
nly.

698 CAN. J . BOT. VOL. 57, 1979
problems in correlation effects on shoot and bud morphogenesis for which only a description can be given at present but this may provide a basis for future experimental studies.
Two different patterns of bud development occur in temperate-zone trees. In the first pattern, veg- etative shoots are fully preformed in the dormant bud. This pattern is found in most species of Pinaceae past the seedling stage of development and in many woody dicot species. In the second pattern, some shoots are not fully preformed in the winter bud. This pattern is found in some woody dicot species such as Betula and Popul~w. In this pattern early leaves are produced before dor- mancy, and late leaves are produced immediately after dormancy, and some species show heterophylly or leaf dimorphism in that predor- mancy leaves are morphologically different from postdormancy leaves (Kozlowski and Clausen 19666). Larix lrrricina (Clausen and Kozlowski 1970), L. leptolepis, L. decidua, and L. gmelini (Fujimoto 1978), and L. occidentalis have LSB which are not completely preformed but hetero- phylly does not occur.
Based on the developmental anatomy of L. oc- cidentcrlis TLSB throughout the year, the first third of the leaves (initiated after bud-scale initiation) were initiated centripetally at the base of the apex and borne on the broad receptacle inside the bud scales. When shoot elongation began after dor- mancy, little internode elongation occurred be- tween these leaves which resulted in a tuft of basal leaves. All axial leaves were initiated up the flanks of the apex. About half of the axial leaves (distal to the basal leaves) were initiated before dormancy and the remainder after dormancy. There were no external morphological differences between these two axial groups. Consequently, no heterophylly occurred between predormancy and postdormancy leaves as was shown also in L. laricina (Clausen and Kozlowski 1970), nor between basal or axial leaves. The latter distinction resulted from the position of basal leaf primordia on the receptacle and the greatly reduced internodal elongation in this region of the stem. The absence of buds in the axils of basal leaves may be explained partially by the absence of sufficient internodal space (Cannell and Bowler 1978) between basal leaves at the time of axillary bud initiation after dormancy.
Buds occurred in the axils of only some axial leaves and in groups rather than being evenly dis- tributed along the lateral shoots of L. occidentalis. In this study it appeared that this arrangement re- sulted from buds being initiated in the axils of leaf primordia which were in close proximity. Cannell
and Bowler (1978) described the special arrange- ment of lateral buds on leader shoots of L. decihla as being evenly (but not randomly) dispersed to maximise their distances apart and offered an alter- native explanation for this arrangement. In their model, various staggered, whorled, or clustered (as in L. occidentalis) branch arrangements could occur as a result of originally evenly distributed axillary primordia being dispersed unequally when the parent shoot elongated. They were careful to point out that their findings may not be extended to lateral branches, as used in the present study, be- cause gravitational and nutritional factors may favour lateral bud initiation (and development) at particular orientations.
Few detailed studies have been made of differ- ences between SSB and LSB development in gym- nosperms. In Ginkgo, there was no anatomical dif- ference between preformed SSB and LSB (Foster 1938; Gunckel and Wetmore 19460, 1946b) but dif- ferences became apparent after dormancy during shoot elongation. Both short and long shoots showed similar early rises in auxin levels followed by comparable decreases at flushing. Long shoots showed a second steep rise in auxin levels causing more shoot elongation and it was this second rise which was absent in short shoots. This was the first apparent difference between long shoot and short shoot development and was believed to initiate the difference between the two shoot types (Gunckel and Thimann 1949). In Larix the difference be- tween axillary SSB and ALSB appeared during the 1st year of development, that is, before dormancy. They were initiated at essentially the same time and were indistinguishable until midway through the early period of rapid leaf initiation. At that time, distal axillary apices which were more vigorous, as evidenced by greater apical size during rapid leaf initiation, differentiated into LSB at the same time as less vigorous distal and proximal axillary apices differentiated into SSB. Leaves were then initiated up the flanks of the LSB apex but not up the flanks of the SSB apex. Consequently, unlike Ginkgo, LSB and SSB in Lnrix were anatomically distinct and predetermined before dormancy and not de- termined only as a result of differential elongation after dormancy. Predetermination in SSB and LSB occurred each year before dormancy throughout the life of the short or long shoot. Conversion of long shoots to short shoots and vice versa is com- mon in L. occidentalis and superficially appears as an abrupt change; however, factors causing this change act on bud development the summer before flushing and shoot expansion. They do not act on internode elongation since most leaves of SSB are
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Sa
nta
Cru
z (U
CSC
) on
11/
09/1
4Fo
r pe
rson
al u
se o
nly.

OWENS A N D MOLDER
borne on a broad, flat to concave receptacle rather than on an axis which could undergo elongation. In L. occidentrrlis, SSB which converted to TLSB passed through a year or more of more vigorous growth, as shown by the larger size of the bud and greater number of leaves produced before differ- entiating into TLSB. Conversely, TLSB of low vigor, as shown by the smaller size of the bud, smaller leaves, and shorter internodes after elon- gation, were those that often differentiated into SSB at the tips of long shoots.
Descriptions have been given which provide a comparison of TLSB (Doak 1935; Hanawa 1966; Owens and Molder 1975, 19776) and SSB (Doak 1935; Hanawa 1967) development in Pinus. These show that differences between TLSB and SSB in Pinus, as in Larix, result from development before dormancy rather than as a result of differential elongation after dormancy as in Girlkgo (Gunckel and Thimann 1949).
SSB development in Larix differs in several ways from that in P~IIIIS (Doak 1935; Hanawa 1967). In Larix, SSB passed through the same developmen- tal cycle each year (bud-scale initiation, leaf initia- tion, and dormancy). In Pir?us, however, on1 y a few bud scales were initiated by a very reduced apex after the 1st year and no additional leaves were normally initiated. In both genera, apices on short shoots may be induced to develop into LSB in response to natural or artificial pruning. Lariat- SSB, unlike those of Pirlus (Hanawa 1967), produced a crown region each year even though shoot elonga- tion was only 1 mm or less. Consequently, the age of short shoots in L ~ ~ r i x , which in this study was up to 8 to 10 years, could be accurately determined by the number ofcrown regions as well as by counting rings of leaf bases. Also, leaves abscised in Lmix rather than entire short shoots as in Pinus. HOW- ever, short shoots of Lnrix were similar to short shoots ofPir7us (Hanawa 1967) in that they became less vigorous after several years. In Lnrix, produc- tion of bud scales and leaves by the short shoot apex diminished in the years just before the short shoot died. SSB development also is of particular interest in Larix because the short shoot apices may differentiate into pollen-cone and seed-cone buds after the 1st year of development, whereas in Pi t~us, cone buds differentiate from newly initiated axillary buds.
In most other members of the Pinaceae having predetermined buds (Owens and Molder 1977a), there is a relationship in time between the end of shoot elongation and the end of bud-scale initiation. This is the important morphogenetic period when apices differentiate into specific bud types. This
was also true for TLSB and SSB on l-year-old and older short shoots in L. occider7trrlis (Figs. 1, 2). Bud scale initiation stopped and buds differentiated on elongating lateral shoots at about the end of the rapid phase of shoot elongation. Axillary SSB dif- ferentiation did not occur until lateral shoot elon- gation was completed, perhaps because they began development later than other vegetative bud types.
Periods of slow or no bud-scale initiation and rapid apical enlargement have been observed in vegetative apices of other conifers (Cecich et al. 1972; Owens and Molder 1973~1, 19736, 1976, 19770) and these have been interpreted as an im- portant morphogenetic stage when many cytologi- cal changes occur. Cecich et a / . (1972) reported an increase in DNA in Picell glaucn during this stage. A similar increase in DNA and mitotic frequency was shown in vegetative apices of Pseudotsuga rnenziesii (Owens and Molder 1973~1) during transi- tion from bud-scale to leaf initiation. The longer more noticeable period of slow bud-scale initiation and apical enlargement in L. occider7t~llis SSB re- sulted from a much greater radial enlargement of the apex and receptacle relative to the apical height compared with other species which have been studied. The end of bud-scale initiation has been shown to be the time when cone buds begin to differentiate in members of the Pinaceae having preformed buds (Owens and Molder 1979). These studies also have shown that an increase in mitotic frequency, greater than that shown during transi- tion from bud-scale to leaf initiation, occurs when apices undergo transition from bud-scale initiation to bract or microsporophyll initiation. This will be the subject of a subsequent study.
BOE. K. N. 1958. Silvics ofwestern larch. U.S. Dep. Agric. For. Ser. Misc. Publ. No. 16. p. 17.
CANNELL, M. G. R., and K. C. BOWLER. 1978. Spatial arrange- ment of lateral buds at the time that they form on leaders of Picerr and Lrrrix. Can. J . For. Res. 8: 129-137.
CECICH, R. A., N. R. LERSTEN, and J . P. MIKSCHE. 1972. A cytophotometric study of nucleic acids and proteins in the shoot apex of white spruce. Am. J . Bot. 59: 442-449.
CLAUSEN, J. J . , and T. T. KOZLOWSKI. 1967. Seasonal growth characteristics of long and short shoots of tamarack. Can. J. Bot. 45: 1643-1651.
1970. Observations on growth of long shoots in Lari-u lriricitzri. Can. J . Bot. 48: 1045- 1048.
CONN, H., J . M. A. DARROW, and V. M. EMMEL. 1960. Staining procedures used in Biological Stain Commission. William and Wilkins, Baltimore.
DOAK, C. C. 1935. Evolution of foliar types, d w a ~ f shoots, and cone scales of Pinus. Ill. Biol. Monogr. 13(3): 1-106.
FOSTER, A. S. 1938. Structure and development of the shoot apex of Gir~kgo bilobn. Bull. Torrey Bot. Club, 65: 531-556.
FOWELLS, H . A. (Compiler). 1965. Silvics of forest trees of the United States. U.S. Dep. Agric., WA, Agric, Handb. NO. 271. pp. 5 4 6 5 5 6 .
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Sa
nta
Cru
z (U
CSC
) on
11/
09/1
4Fo
r pe
rson
al u
se o
nly.

700 CAN. J . BOT. VOL. 57, 1979
FRAMP-I~ON, C. V. 1960. Some aspects of the developmental anatomy of the "long" shoots in Lrrri.~ rdecidria Mill., with p;rrticular reference to seasonal periodicity. New Phytol. 59: 175-191.
FUJIMOTO, S. 1978. Studies on the shoot formation in L0ri.r. lpptol~~pi.s Cordon. Res. Bull. Coll. Exp. For., Coll. Agric. Hokkaido Univ.. Sapporo, Jpn. 35: 1-28.
GUNCKEL, J . E., ilnd K. V. T H I M A N N . 1949. Studies of de- velopnlent in long shoots and short shoots of Ci11kgo hilobtr L. 111. Auxin production in shoot growth. Am. J . Bot. 36: 145-151.
GUNCKEL, J . E. . and R. H. WETMORE. 19460. Studies of de- velopment in long shoots and short shoots of Citrkgo Oilohrr L. I. Origin and patterns of development of the cortex, pith and procambium. Am. J . Bot. 33: 285-295.
1946h. Studies of development in long shoots and short shoots of Gitrligo hilobrr L. 11. Phyllotaxis and the organiza- tion of the primary vascular system; primary phloem and primary xylem. Am. J . Bot. 33: 345-358.
HANAWA, J . 1966. Growth and development in the shoot apexof Pitrrts rlc,tr.s~flora I. Growth periodicity and structure of the terminal vegetative shoot apex. Bot. Map. (Tokyo), 79: 736-746.
----- 1967. Growth and development in the shoot apex of Pitfrts C / L , I I S ( ~ ~ ~ N 11. Ontogeny of the dwarf shoot and lateral branch. Bot. Mag. (Tokyo), 80: 248-256.
Hosle, R. C. 1973. Native treesofcanada. Dep. Environ., Can. For. Serv., Ottawa.
JOHANSEN, D. A. 1940. Plant microtechnique. McGraw-Hill, New York.
KOZLOWSKI, T. T. 1964. Shoot growth in woody plants. Bot. Rev. 30: 335-392.
KOZLOWSKI, T. T . , and J. J . CLAUSEN. 1966~1. Seasonal de- velopment of long and short shoot components of taniarack. Bull. Ecol. Soc. Am.47: 113-114.
19660. Shoot growth characteristics of heterophyllous woody plants. Can. J . Bot. 44: 827-843.
OWENS, J . N., i l ~ d M. MOLDER. 1973rr. A study of DNA and mitotic activity in the vegetative apex of Do~~glas-fir during the annual growth cycle. Can. J . Bot. 51: 1395-1409.
1973h. Bud development in Western hemlock: I. Annual growth cycle of vegetative buds. Can. J. Bot. 51: 2223-2231.
1975. Development of long-shoot terminal buds of Pitiris cotltorta spp. cot~/ortci. 111 Management of lodgepole pine ecosystems. Vol. l . Proc. Pullman, Wash. pp. 86-104.
1976. Bud development in Sitka spruce. I. Ann~lal growth cycle of the vegetative buds and shoots. Can. J . Bot. 54: 313-325.
19770. The times and patterns of cone differentiation in western conifers. Proceedings of the Third World Consulta- tions on Forest Tree Breeding. Canberra.
19770. Development of long-shoot terminal buds of western white pine (Pitriis t~otiticola). Can. J. Bot. 55: 1308-1321.
1979. The times and patterns of cone differentiation in western North American conifers. Proceedings of IUFRO Symposium on flowering and seed development in trees. Starkville, Mississippi. In press.
SASS, J . E. 1958. Botanical microtechnique. 3rd ed. Iowa State University Press, Ames, Iowa.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Sa
nta
Cru
z (U
CSC
) on
11/
09/1
4Fo
r pe
rson
al u
se o
nly.





























