Embed Size (px)
Citation preview

MARINE MAMMAL SCIENCE, 26(1): 202–212 (January 2010)C© 2009 by the Society for Marine MammalogyDOI: 10.1111/j.1748-7692.2009.00333.x
Bryde’s whale (Balaenoptera brydei) stable associations and dive profiles:New insights into foraging behavior
FILIPE ALVES
ANA DINIS
Museu da Baleia da Madeira,9200-031 Canical, Madeira, PortugalE-mail: [email protected]
IRMA CASCAO
INETI/Departamento de Biotecnologia,Edifıcio E–1◦ andar,
Estrada do Paco do Lumiar 22, 1649-038 Lisboa, Portugal
LUIS FREITAS
Museu da Baleia da Madeira,9200-031 Canical, Madeira, Portugal
Bryde’s whales (Balaenoptera brydei) are the least known of the large baleen whalesand there are still some questions about their taxonomy and the number of species(Kato 2002). Indeed, previous studies of Bryde’s whale feeding ecology were basedonly on observational events (e.g., Siciliano et al. 2004) and on stomach contents(e.g., Kawamura 1980, Murase et al. 2007). Rorqual whales are known to displaylunge feeding behavior (Werth 2000); however, there has been no evidence of Bryde’swhales lunge feeding at depth. Apart from a few studies that have used time depthrecorders (TDRs) to analyze the dive profiles and feeding behaviors of fin whales inthe Mediterranean Sea (Panigada et al. 1999) and of fin and blue whales in the Gulfof California (Croll et al. 2001), knowledge of the dive profiles of other Balaenopteraspecies is unknown. Watkins et al. (1979) successfully tracked separately two adultBryde’s whales with radio tags, providing some surfacing data; however, no dive datawere obtained.
This study is the first to analyze dive data from TDRs attached to Bryde’s whalesto examine foraging behavior from dive profiles. Data on individual associations overconsecutive years, which is scarce for any baleen whale species, are presented and newinsights into foraging behavior given.
In the waters around Madeira Island (southeast North Atlantic, 32◦N, 17◦W)Bryde’s whales can frequently be found alone or in groups of two to five individualsbetween the months of June and November. During tagging surveys, carried outusing the Madeira Whale Museum’s (MWM) 6.5 m inflatable boat Roaz in 2006,two individual adult Bryde’s whales were tagged on 27 June at 6 km off the southeastcoast of the island. The tags used were Mk9 mold 161 (up to 1,000 m) TDR (WildlifeComputers, Redmond, WA) set to sample once per second and very high-frequency(VHF) radio transmitters with a 34 cm semirigid wire antenna (Telonics, Mesa, AZ)
202

NOTES 203
housed in a custom-made syntactic foam body, attached to an 8 cm diameter suctioncup. Each tag weighed approximately 400 g and measured approximately 20 cm ×6 cm × 3 cm (tag body). Before each deployment, the interior of the suction-cupwas covered with a thin layer of silicone grease (Elbesil BM, Boewing Company,Germany) and the tags were deployed using an extensible 5 m pole spear. The firstindividual (Bbr002) was tagged 5 min after the whales were initially sighted, whilethe second individual (Bbr003) was tagged 11 min later. Once tagged, the whaleswere followed using visual observations for 2.5 h. Whenever the boat neared thetagged whales, the animals’ behavior, global positioning system (GPS) position,and travel direction were recorded. No other whales or boats were seen in the areabefore or after tagging. Data from recovered tags were downloaded to a computerand processed with the Wildlife Computer software’s Mk9Host (v1.09.0001) andInstrument Helper (v0.701), respectively.
Because the data came from two associated whales, there is unequivocally somedegree of dependence between the two dive tracks. Therefore, in order to minimizeits effects, we assumed independence of samples, that is, each whale was treatedseparately. Mean and standard deviation values were calculated for each whale asparameters were normally distributed (K-S test). From the investigation of thedive profile, the multimodal nature of a three-dimensional (3D) plot of dive depthand duration (not shown), and the log-survivorship plot of dive depth (not shown,following Hooker and Baird 2001) it was possible to discriminate two distinct divetypes: one included shallow dives of less than 40 m, U-shaped, short in time, andwithout lunges (defined below); and the other included deeper, longer dives withor without lunges, and containing both U- and V-shaped dives. In order to excludesurface respiratory behavior, dives shallower than 10 m depth were considered assurface activity. Bottom time was defined as the time between the point when descentstops and depth stabilizes and the point when ascent begins, which is characterizedby a stable and continuous decrease of the depth until the surface is reached. Someof the dives showed lunges. A lunge was defined following Croll et al. (2001) witha minimum amplitude in ascent of 5 m, lasting between 20 s and 2 min (defineda posteriori). Foraging typically includes both searching for prey and prey capture(Heithaus and Dill 2002); thus all deep dives were likely foraging dives, althoughonly a subset involved actual feeding. Therefore, those with lunges were classified asfeeding dives, as has been shown for fin and blue whales (Acevedo-Gutierrez et al.2002), while the other deep dives were considered as foraging dives without preycapture. Because neither Bbr002 nor Bbr003 made lunges during shallow dives ofless than 40 m, we assumed that for the total track period, foraging activity took placeonly during deeper dives. Thus, we decided to compare only deep dives with lunges(feeding dives) vs. without lunges (foraging dives without prey capture). For thiscomparison, and for each whale, one-way analysis of variance (ANOVA) significancetests were performed in order to examine differences for nine dive parameters. Test ofsignificance was assessed at P = 0.05 and the assumptions were tested using the K-Sand Levene’s tests, using STATISTICA 8 software (StatSoft, Tulsa, OK). Additionally,for each whale, two correlations were made: (1) between the dive duration and thepreceding surface interval of the progressively shallower and deeper series of foraging

204 MARINE MAMMAL SCIENCE, VOL. 26, NO. 1, 2010
dives, and (2) between the deep dives with bottom time and the number of bottomlunges. The preceding surface interval was defined as the period between the endof the previous deep dive and the start of the analyzed one. The remaining deepdives were not included in the first correlation because the preceding surface intervalcovered a large period of up to 2 h, which most likely included other purposes ratherthan preparing for the deep dive.
For individual identification purposes, the Bryde’s whales were photographedusing a digital Nikon D2H (Nikon Corporation, Tokyo, Japan) equipped with 70–300 mm F4 and 80–200 mm F2.8 zoom lenses and all photographs were processedusing Corel Photo-Paint 9 software (Corel Corporation, Ottawa, Canada). The goodquality images were sorted following Arnbom (1987), based on the shape and marksof the dorsal fin. These were then compared and included in the MWM Bryde’s whalephoto-identification catalog.
A skin sample from Bbr003 was also obtained from the recovered suction-cup.Genetic analyses, that included DNA extractions and molecular sexing (followingstandard protocols: Gemmel and Akiyama 1996, Abe et al. 2001), revealed that thisindividual was a male. Because Bryde’s whales taxonomic status is still under revision(Kato 2002, Wada et al. 2003) the authors opted for a Blast search in the GenBankdatabase, which retrieved sequences of Bryde’s whales Olsen, 1913 (sample “BEM02”was deposited in GenBank, reference number EU030282).
The visual observations showed that the tagged whales slowly moved togetherin a northeasterly direction following the coastline. The two whales, one slightlybigger than the other (Bbr002 and Bbr003 were approximately 11 m and 13 m,respectively), showed indifference to the boat presence. The tags were recovered onthe next day at about 11 km from the deployment site. A total of 28 h of track timewere obtained from both tags. The whales spent 84.5% of their total track time atthe surface or performing shallow dives of less than 40 m and 15.5% performingdeeper dives (Table 1). The tag from individual Bbr002 was deployed at 1415 andreleased at 0447, resulting in 14.53 h of dive data. Bbr002 performed a total of 155dives, of which 126 were shallow dives of less than 40 m and 29 were deeper dives
Table 1. Summary statistics of total track time. M = male.
Surface (<10 m) orshallow dives of less
than 40 m
Number Deep divesof shallow TotalTrack dives of Number Number
Start track time Time less than Time of ofWhale Sex (local time) (h) (h) 40 m (h) dives dives
Bbr002 ? 14 h 15 min 14.53 11.90 126 2.62 29 155Bbr003 M 14 h 26 min 13.42 11.70 136 1.70 23 159
Total 27.95 23.60 262 4.32 52 314(84.5%) (15.5%)

NOTES 205
(Table 1). The maximum dive duration recorded was 7.9 min; however, the whale wasstill at 211 m depth when the tag came off, and thus we estimated a maximum diveduration of 9.4 min, based on the highest mean ascent rate from this individual (seeTable 2). The maximum dive depth was 267 m. The tag from Bbr003 was deployedat 1426 and released at 0351, providing 13.4 h of dive data. Bbr003 performed atotal of 159 dives, of which 136 were shallow dives of less than 40 m and 23 weredeeper dives (Table 1). The maximum dive duration recorded was 6.5 min and themaximum dive depth was 292 m.
Analysis of the dive profiles for each whale during the period when the tags weresimultaneously attached showed a temporal synchrony in the dive behavior betweenthe two whales (Fig. 1). During this period the whales simultaneously performed 23deep dives, with a single exception occurring at 2116, when Bbr002 dived alone.The dive synchrony between Bbr002 and Bbr003 is highlighted when analyzing thedeep dive data presented in Table 2.
The analysis of the dive profiles during the total track time showed that both whalespresented specific periods of day for the two distinct dive types. Both performed alarge series of shallow dives of less than 40 m until approximately 1.5 h before sunset,when they made the two deepest dives. During night, series of sequentially deepdives took place, spaced for short time intervals of 2–5 min. Generally, the deep divestended to be sequentially shallower from evening to night, and deeper from night toearly morning, as highlighted in Figures 1 and 2.
A total of 52 foraging dives were recorded (29 from Bbr002 and 23 from Bbr003)and occurred with darkening. From these, 27 were considered to be feeding dives(18 from Bbr002 and 9 from Bbr003) and 25 were considered to be foraging diveswithout prey capture (11 and 14 for each whale, respectively). A total of 43 lungeswere recorded in the feeding dives, 6 during the descent phase (all from Bbr002), 33during the bottom period (20 from Bbr002 and 13 from Bbr003), and 4 during theascent phase (all from Bbr002). For each whale, the comparison between these twodive types showed that feeding dives were longer, deeper, and with a longer bottomtime than foraging dives without lunges (Table 3). The correlations showed high r2
values (Fig. 3A, B).During the period that the whales were tagged, from early afternoon to early
morning, the majority of their time was dedicated to surface activity, which could bemostly related to resting and respiration. During this surface period, it was unlikelythat traveling and feeding occurred, mainly because the tags were recovered close tothe deployment site, and because the whales were not seen feeding during the visualfollow-up and no lunges were detected from the dive analyses. Nevertheless, Bryde’swhales feeding at the surface on small schooling fish have been previously observedby the authors in the same area where these two whales were tagged (southeastMadeira), and have been described in other areas of the world (Tershy et al. 1993).
During the short period of time that the whales spent performing foraging behavior(only 15.5% of total track time), lunges occurred in about half of the deep dives,indicating feeding activity, as suggested by other studies made on other Balaenopterasp. (Panigada et al. 1999, Croll et al. 2001). Despite lunges having occurred in allphases of the deep feeding dives, most of them were recorded in the bottom period,

206 MARINE MAMMAL SCIENCE, VOL. 26, NO. 1, 2010
Tabl
e2.
Div
eda
taof
the
sim
ulta
neou
s23
deep
dive
sof
Bbr
002
and
Bbr
003
(one
inco
mpl
ete)
.
Des
cent
Bot
tom
Asc
ent
Div
eM
ean
Max
.N
umbe
rM
ax.
Num
ber
Mea
nM
ax.
Num
ber
dura
tion
Dur
atio
nra
tera
teof
Dur
atio
nde
pth
ofD
urat
ion
rate
rate
of(m
in)
(min
)(m
/s)
(m/s
)lu
nges
(min
)(m
)lu
nges
(min
)(m
/s)
(m/s
)lu
nges
Bb
r002
Mea
n±
SD5.
13±
1.20
1.73
±0.
711.
2±
0.4
1.7
±0.
50.
3±
0.5
0.83
±0.
7513
2±
590.
5±
0.7
2.59
±1.
020.
8±
0.5
1.8
±0.
60.
2±
0.5
Tota
l39
.82
(33.
7%)
619
.19
(16.
3%)
1158
.9(5
0.0%
)4
Bb
r003
Mea
n±
SD4.
48±
0.92
1.62
±0.
521.
3±
0.4
2.0
±0.
60.
0±
0.0
0.84
±0.
7613
6±
640.
6±
0.8
2.05
±0.
631.
1±
0.6
2.0
±1.
10.
0±
0.0
Tota
l:37
.34
(36.
6%)
019
.43
(19.
1%)
1345
.17
(44.
3%)
0

NOTES 207
Figure 1. Dive profiles of Bbr002 and Bbr003 of total track time showing individualsynchrony and with emphasis on the dives. Sunset at 2119 and sunrise at 0700. The arrowshighlight the sequentially shallower (during early night) and deeper (during early morning)series of deep dives.
Figure 2. Detailed dives profile of the sequentially shallower (during early night) anddeeper (during early morning) series of deep dives of Bbr002 and Bbr003. ↑ = lunges ofBbr002; � = lunges of Bbr003.
indicating a higher peak in feeding activity during this phase, suggesting that, likefin and blue whales, Bryde’s whales also feed at depth and prey consumption takesplace mainly during the bottom phase of the deep dive. Additionally, this peak infeeding activity explains the significantly longer bottom period of feeding dives whencompared with foraging dives without prey capture.

208 MARINE MAMMAL SCIENCE, VOL. 26, NO. 1, 2010
Tabl
e3.
Div
esst
atis
tics
(mea
n±
stan
dard
devi
atio
n,pe
rcen
tage
,and
one-
way
AN
OV
AP
valu
es)f
orth
etw
ode
epdi
vety
pes
ofB
br00
2an
dB
br00
3.∗ P
<0.
05.
Des
cent
Bot
tom
Asc
ent
Type
ofD
ive
dura
tion
Dur
atio
nM
ean
rate
Max
.rat
eD
urat
ion
Max
.dep
thD
urat
ion
Mea
nra
teM
ax.r
ate
dive
(min
)(m
in)
(m/s
)(m
/s)
(min
)(m
)(m
in)
(m/s
)(m
/s)
Bb
r002
Wit
hlu
nges
5.92
±1.
451.
81±
0.79
1.3
±0.
52.
0±
0.6
1.79
±1.
6415
9±
692.
37±
1.02
1.1
±0.
62.
3±
1.4
n=
1810
0%31
%31
%38
%W
itho
utlu
nges
4.74
±0.
981.
55±
0.60
1.2
±0.
41.
7±
0.5
0.59
±0.
7111
9±
552.
58±
0.85
0.8
±0.
41.
9±
1.0
n=
1110
0%33
%13
%54
%P
valu
es0.
03∗
0.36
0.67
0.19
0.03
∗0.
120.
560.
140.
44
Bb
r003
Wit
hlu
nges
4.98
±0.
921.
76±
0.6
1.5
±0.
32.
1±
0.6
1.54
±0.
7417
3±
561.
73±
0.56
1.6
±0.
52.
9±
1.4
n=
910
0%35
%31
%31
%W
itho
utlu
nges
4.18
±0.
771.
54±
0.45
1.2
±0.
41.
9±
0.7
0.4
±0.
2911
2±
582.
24±
0.60
0.8
±0.
31.
5±
0.4
n=
1410
0%37
%9%
54%
Pva
lues
0.04
∗0.
330.
050.
370.
00∗
0.02
∗0.
060.
00∗
0.00
∗
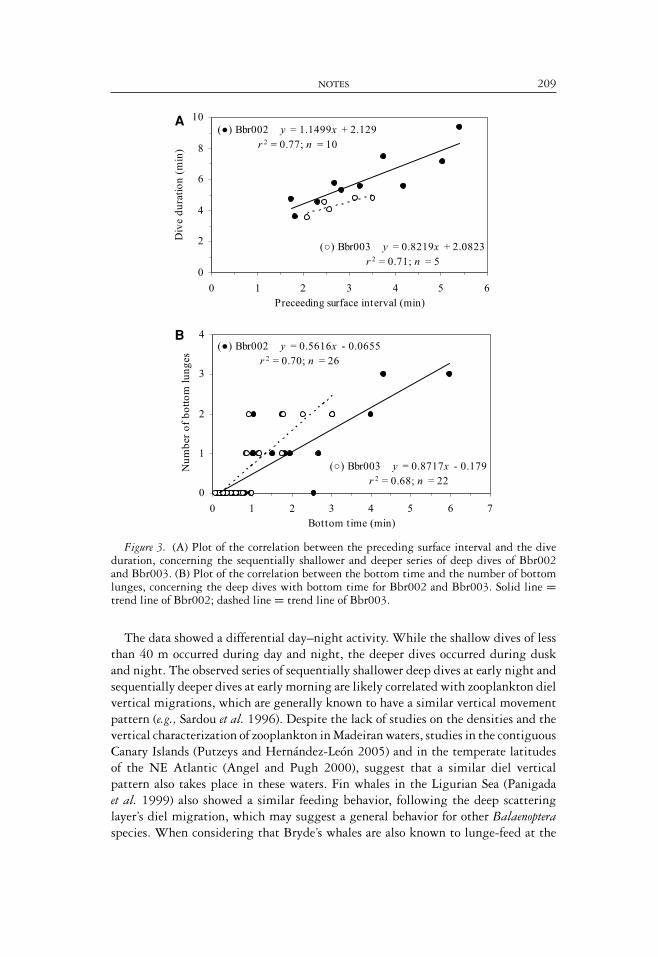
NOTES 209
Figure 3. (A) Plot of the correlation between the preceding surface interval and the diveduration, concerning the sequentially shallower and deeper series of deep dives of Bbr002and Bbr003. (B) Plot of the correlation between the bottom time and the number of bottomlunges, concerning the deep dives with bottom time for Bbr002 and Bbr003. Solid line =trend line of Bbr002; dashed line = trend line of Bbr003.
The data showed a differential day–night activity. While the shallow dives of lessthan 40 m occurred during day and night, the deeper dives occurred during duskand night. The observed series of sequentially shallower deep dives at early night andsequentially deeper dives at early morning are likely correlated with zooplankton dielvertical migrations, which are generally known to have a similar vertical movementpattern (e.g., Sardou et al. 1996). Despite the lack of studies on the densities and thevertical characterization of zooplankton in Madeiran waters, studies in the contiguousCanary Islands (Putzeys and Hernandez-Leon 2005) and in the temperate latitudesof the NE Atlantic (Angel and Pugh 2000), suggest that a similar diel verticalpattern also takes place in these waters. Fin whales in the Ligurian Sea (Panigadaet al. 1999) also showed a similar feeding behavior, following the deep scatteringlayer’s diel migration, which may suggest a general behavior for other Balaenopteraspecies. When considering that Bryde’s whales are also known to lunge-feed at the

210 MARINE MAMMAL SCIENCE, VOL. 26, NO. 1, 2010
surface, it is suggested that this species, and possibly other baleen whales, may usedifferent strategies to feed upon different prey species during the day (e.g., schools ofsmall pelagic fishes) and during the night (e.g., masses of zooplankton). It should benoted that our observations are based only on a relatively short duration track of twoindividuals, both in the same group, so clearly additional dive data from this specieswill be needed to confirm these suggestions.
The mean dive duration of deep dives found in this study (5.0 ± 1.2 min, n =50) is in agreement with the values described by Best et al. (1984) that, based onvisual observations off the South African coast, calculated a mean dive duration forthe longest dives of 4.9 ± 0.8 min (n = 14). Also, the longest dive from these data(estimated 9.4 min) is coincident with the longest dive reported in Best et al. (1984)(9.1 min). However, longer dives have been recorded for Bryde’s whales, for instance,the 11.5 min dive observed by di Sciara (1983).
The synchrony of the dive profile between the two tagged whales in this studyindicates a strong temporal association during the entire track time, including thedeep dives related to feeding activity. Despite the tags not being capable of providinginformation on spatial associations, such association is suggested by the 2.5 h periodwhen the whales were followed and observed coming back to surface together. Thus,a strong spatio-temporal association between these individuals is suggested for thetotal track period. Our results differ from the visual observations of surface feedingof Bryde’s whales in the Central Gulf of California made by Tershy et al. (1993),which registered that in feeding aggregations, each whale appears to respond to themovements of its prey independently from each other.
Photo-identification analysis revealed that Bbr002 and Bbr003 have been identi-fied together on seven occasions, during the summers of 2005 (1 and 24 July and11 August), 2006 (27 and 30 June and 13 July), and 2007 (26 August). In 2008,Bbr003 was photographed alone on 22 June, 3, 13, and 21 July. All sightings oc-curred between 4 km and 9 km off the south and east coast of Madeira Island and noother whales were observed in the vicinity.
The dive synchrony and the association between the two individuals during threeconsecutive years in this study provide new insights into the foraging behavior andsocial organization of Bryde’s whales. Although from this study, we cannot infer thepossibility of long-term continued associations for Bryde’s whale individuals, theobserved associations during consecutive summers in Madeiran waters suggests thatthese individuals can form stable associations at least during part of their life. Themolecular sexing helped to exclude the hypothesis that the pair was a mother andher young. Obtaining a biopsy sample from Bbr002 will provide useful informationto analyze further whether the association between these two individuals was one oftwo males or of a male and female.
ACKNOWLEDGMENTS
We would like to thank the crew members of the MWM for logistical support, the CamaraMunicipal de Machico for financial support, Sophie Querouil for genetic assistance, AdalbertoCarvalho for mapping using GIS, Claudia Ribeiro and Catia Nicolau for useful comments,

NOTES 211
Luıs Dias and Rita Ferreira for contributing with photographs, and Helen Mitcheson forreviewing English. The LIFE and FEDER/INTERREG III-B EU programs funded the “Pro-jecto para a Conservacao dos Cetaceos no Arquipelago da Madeira” (LIFE99 NAT/P/006432)and the MACETUS project (MAC/4.2/M10), respectively. We also thank Robin Baird andtwo anonymous reviewers that commented on previous draft and contributed with usefulinformation.
LITERATURE CITED
Abe, H., M. Goto and L. A. Pastene. 2001. Practical use of multiplex fluorescent PCR forcetacean sex identification. Marine Mammal Science 17:657–664.
Acevedo-Gutierrez, A., D. A. Croll and B. R. Tershy. 2002. High feeding costs limit divetime in the largest whales. Journal of Experimental Biology 205:1747–1753.
Angel, M. V., and P. R. Pugh. 2000. Quantification of diel vertical migration by 24 mi-cronektonic taxa in the northeast Atlantic. Hydrobiologia 440:161–179.
Arnbom, T. 1987. Individual identification of sperm whales. Report of the InternationalWhaling Commission 37:201–204.
Best, P. B., D. S. Butterworth and L. H. Rickett. 1984. An assessment cruise for the SouthAfrican inshore stock of Bryde’s Whales (Balaenoptera edeni). Report of the IntenationalWhaling Commission 34:403–423.
Croll, D. A., A. Acevedo-Gutierrez, B. Tershy and J. Urban-Ramırez. 2001. The divingbehavior of blue and fin whales: Is dive duration shorter than expected based on oxygenstores? Comparative Biochemistry and Physiology-Part A: Molecular and IntegrativePhysiology 129:797–809.
di Sciara, G. N. 1983. Bryde’s whales (Balaenoptera edeni Anderson 1878) off easternVenezuela (Cetacea, Balaenopteridae). Technical Report 83-153, Hubbs-Sea World Re-search Institute, San Diego, CA. 27 pp.
Gemmel, N. J., and S. Akiyama. 1996. An efficient method for the extraction of DNA fromvertebrate tissues. Trends in Genetics 12:338–339.
Heithaus, M. R., and L. M. Dill. 2002. Feeding strategies and tactics. Pages 412–422 in W.F. Perrin, B. Wursig and J. G. M. Thewissen, eds. Encyclopedia of marine mammals.Academic Press, San Diego, CA.
Hooker, S. K., and R. W. Baird. 2001. Diving and ranging behaviour of odontocetes: Amethodological review and critique. Mammal Review 31:81–105.
Kato, H. 2002. Bryde’s whales: Balaenoptera edeni and B. brydei. Pages 171–177 in W. F. Perrin,B. Wursig and J. G. M. Thewissen, eds. Encyclopedia of marine mammals. AcademicPress, San Diego, CA.
Kawamura, A. 1980. Food habits of the Bryde’s whales taken in the South Pacific and IndianOceans. Scientific Reports of the Whales Research Institute, Tokyo 32:1–23.
Murase, H., T. Tamura, H. Kiwada, Y. Fujise, H. Watanabe, H. Ohizumi, S. Yonezaki,H. Okamura and S. Kawahara. 2007. Prey selection of common minke (Balaenopteraacutorostrata) and Bryde’s (Balaenoptera edeni) whales in the western North Pacific in2000 and 2001. Fisheries Oceanography 16:186–201.
Panigada, S., M. Zanardelli, S. Canese and M. Jahoda. 1999. How deep can baleen whalesdive? Marine Ecology Progress Series 187:309–311.
Putzeys, S., and S. Hernandez-Leon. 2005. A model of zooplankton diel vertical migration offthe Canary Islands: Implication for active carbon flux. Journal of Sea Research 53:213–222.
Sardou, J., M. Etienne and V. Andersen. 1996. Seasonal abundance and vertical distributions ofmacroplankton and micronecton in the northwestern Mediterranean Sea. OceanologicaActa 19:645–656.
Siciliano, S., M. C. de O. Santos, A. F. Vicente, F. Alvarenga, E. Zampirolli, J. Lailson-Brito,A. Azevedo and J. L. Pizzorno. 2004. Strandings and feeding records of Bryde’s whales(Balaenoptera edeni) in south-eastern Brazil. Journal of the Marine Biological Associationof the UK 84:857–859.

212 MARINE MAMMAL SCIENCE, VOL. 26, NO. 1, 2010
Tershy, B., A. Acevedo-Gutierrez, D. Breese and C. Strong. 1993. Diet and feeding behavior offin and Bryde’s whales in the Central Gulf of California, Mexico. Revista de InvestigacionCientifica de la Universidad Autonoma de Baja California Sur. Serie Ciencias del Mar(No. Esp. SOMEMMA) 1:31–37.
Wada, S., M. Oishi and T. K. Yamada. 2003. A newly discovered species of living baleenwhale. Nature 426:278–281.
Watkins, W. A., G. N. di Sciara and K. E. Moore. 1979. Observations and radio taggingof Balaenoptera edeni near Puerto de la Cruz, Venezuela. Woods Hole OceanographicInstitution Technical Report WHOI-79-89. 8 pp.
Werth, A. J. 2000. Feeding in marine mammals. Pages 475–514 in K. Schwenk, ed. Feeding:Form, function and evolution in tetratpod vertebrates. Academic Press, New York, NY.
Received: 4 April 2008Accepted: 14 April 2009





























