Embed Size (px)
Citation preview

Brainy but not too brainy:starting and stopping neuroblastdivisions in DrosophilaCedric Maurange and Alex P. Gould
Medical Research Council, National Institute for Medical Research, The Ridgeway, Mill Hill, London NW7 1AA, UK
Drosophila neuroblasts are similar to mammalian neural
stem cells in that they self-renew and have the potential
to generate many different types of neurons and glia.
They have already proved useful for uncovering asym-
metric division components and now look set to provide
insights into how stem cell divisions are initiated and
terminated during neural development. In particular,
some of the humoral factors and short-range ‘niche’
signals that modulate neuroblast activity during post-
embryonic development have been identified. In
addition, recent studies have begun to reveal how the
total number of cells generated by a single neuroblast is
regulated by spatial and temporal cues from Hox
proteins and a transcription-factor series linked to cell
cycle progression.
During development, the fruit fly Drosophila undergoes aspectacular metamorphosis from burrowing larva intoflying adult. Not surprisingly, these radically differentbody plans are powered by equally distinctive nervoussystems. Whereas the CNS of the newly hatched larvacontains broadly similar numbers of neurons in the head,thoracic and abdominal segments, the vast majority of theadult CNS is derived from just the head and thorax. Thisanterior-heavy CNS reflects the location of the majorsensory and motor organs of the adult such as antennae,eyes, wings and legs. Underlying the two different nervoussystems of Drosophila are two distinct phases of neuro-genesis, embryonic and postembryonic, which are separ-ated by a pause in neuronal proliferation known asquiescence. Only the embryonic period of neurogenesis isresponsible for building the functional CNS of the larva,whereas both phases contribute to forming the muchlarger and more complex adult CNS. Thus, the adult CNSis a composite of 10% remodelled larval cells, born duringembryogenesis, plus 90% imaginal cells, born duringlarval and pupal stages. Interestingly, although most ifnot all adult motoneurons are remodelled from their larvalcounterparts, the vast majority of adult interneurons areformed postembryonically.
How then does the adult CNS grow from its larvalpredecessor, and what are the region-specific develop-mental cues responsible for specifying its final size and
Corresponding author: Alex P. Gould ([email protected]).
www.sciencedirect.com 0166-2236/$ - see front matter Q 2004 Elsevier Ltd. All rights reserved
shape? One key finding is that the larval and adult CNSare both generated from a set of well-characterized neuralprecursors, called neuroblasts, formed early in embryo-genesis [1,2]. These resemble mammalian neural stemcells in that they undergo self-renewing divisions and aremultipotent, capable of generating several different typesof neurons and glia. Neuroblasts divide by a buddingprocess that generates a set of smaller ganglion mothercells (GMCs). Each GMC, however, usually divides onlyonce to produce two postmitotic neurons or glia. Theasymmetric mode of neuroblast division has been inten-sively studied and many cell-polarity proteins andother components regulating it are nowwell characterized[3–5]. Less is known, however, about the nature of thedevelopmental cues that regulate the number of times aneuroblast will divide. This aspect of neural proliferationwill be reviewed here, with focus on how the embryonicand postembryonic phases of neurogenesis contribute tospecifying the final size and highly regionalized shape ofthe adult CNS.
The embryonic phase of neurogenesis
Every thoracic or abdominal hemisegment of the earlyembryo gives rise to an invariant array of 30 identifiableneuroblasts [2]. These are each assigned a unique identityby a grid of anteroposterior and dorsoventral positionalinformation [6]. This influences both the number and thetype of cells that comprise each embryonic neuroblastlineage [7–12]. Within a given hemisegment, the size ofneuroblast clones produced during the embryonic phase ofneurogenesis varies greatly: thus, at one extreme neuro-blast MP2 generates only two cells [7], whereas neuroblastNB7-1 can produce O40 cells [9]. There are also somethoracic versus abdominal differences in the sizes of cer-tain embryonic lineages, notably those of the neuroblastsNB1-1, NB3-1 and NB7-1 [7,9,13]. The anteroposteriorpositional information regulating these segmental differ-ences is highly likely to be provided by the Hox (homeotic)family of homeodomain transcription factors [14,15].Strong evidence for this comes from studies of NB1-1,where the Hox proteins Ultrabithorax (Ubx) andAbdominal-A (AbdA) are required to limit the number ofcells in the abdominal lineage to about half that of itsthoracic counterpart [16]. However, the summation of allthe segmental differences in embryonic lineage sizes hasonly a minor effect on the overall shape of the larval CNS.
Review TRENDS in Neurosciences Vol.28 No.1 January 2005
. doi:10.1016/j.tins.2004.10.009
CasNB
GMC
Progeny
?
(a)
Hb Kr Pdm
Hb Kr Kr Kr Kr
Hb Hb Hb Hb Hb
+Hb
+Kr
NB7-3
NB7-3
NB7-3
(b) (i)
(ii)
MI
Hb Kr Pdm
II
TRENDS in Neurosciences
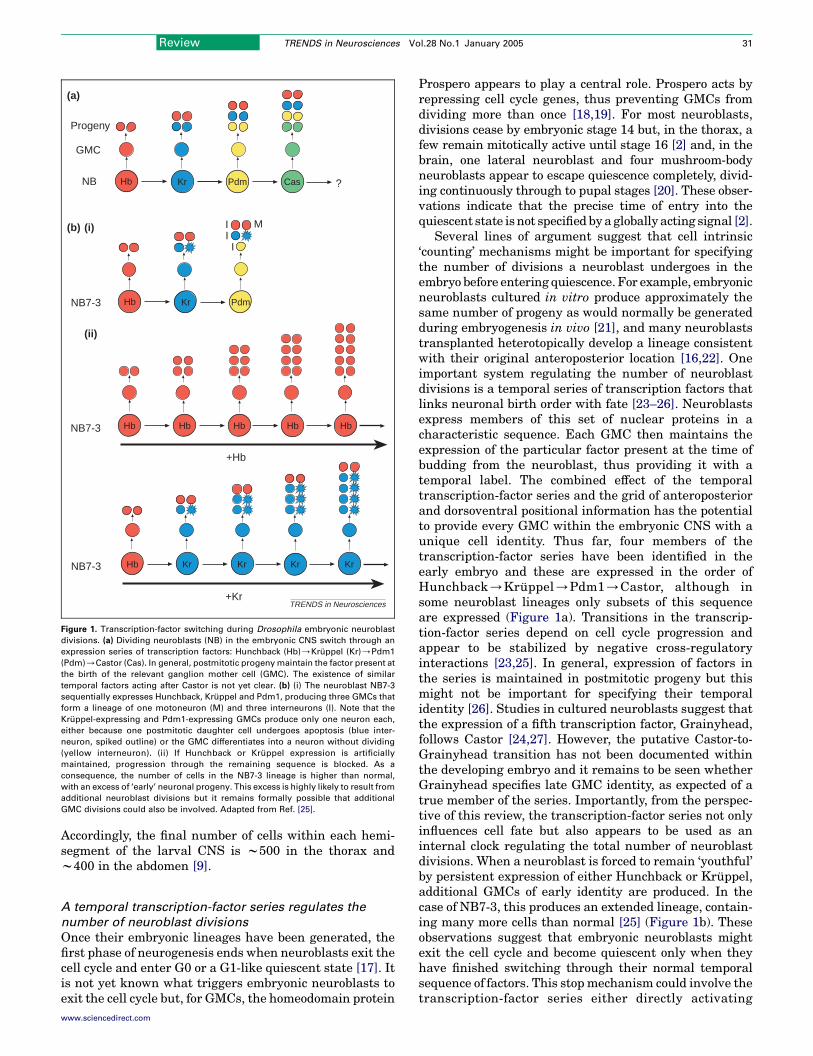
Figure 1. Transcription-factor switching during Drosophila embryonic neuroblast
divisions. (a) Dividing neuroblasts (NB) in the embryonic CNS switch through an
expression series of transcription factors: Hunchback (Hb)/Kruppel (Kr)/Pdm1
(Pdm)/Castor (Cas). In general, postmitotic progeny maintain the factor present at
the birth of the relevant ganglion mother cell (GMC). The existence of similar
temporal factors acting after Castor is not yet clear. (b) (i) The neuroblast NB7-3
sequentially expresses Hunchback, Kruppel and Pdm1, producing three GMCs that
form a lineage of one motoneuron (M) and three interneurons (I). Note that the
Kruppel-expressing and Pdm1-expressing GMCs produce only one neuron each,
either because one postmitotic daughter cell undergoes apoptosis (blue inter-
neuron, spiked outline) or the GMC differentiates into a neuron without dividing
(yellow interneuron). (ii) If Hunchback or Kruppel expression is artificially
maintained, progression through the remaining sequence is blocked. As a
consequence, the number of cells in the NB7-3 lineage is higher than normal,
with an excess of ‘early’ neuronal progeny. This excess is highly likely to result from
additional neuroblast divisions but it remains formally possible that additional
GMC divisions could also be involved. Adapted from Ref. [25].
Review TRENDS in Neurosciences Vol.28 No.1 January 2005 31
Accordingly, the final number of cells within each hemi-segment of the larval CNS is w500 in the thorax andw400 in the abdomen [9].
A temporal transcription-factor series regulates the
number of neuroblast divisions
Once their embryonic lineages have been generated, thefirst phase of neurogenesis ends when neuroblasts exit thecell cycle and enter G0 or a G1-like quiescent state [17]. Itis not yet known what triggers embryonic neuroblasts toexit the cell cycle but, for GMCs, the homeodomain protein
www.sciencedirect.com
Prospero appears to play a central role. Prospero acts byrepressing cell cycle genes, thus preventing GMCs fromdividing more than once [18,19]. For most neuroblasts,divisions cease by embryonic stage 14 but, in the thorax, afew remain mitotically active until stage 16 [2] and, in thebrain, one lateral neuroblast and four mushroom-bodyneuroblasts appear to escape quiescence completely, divid-ing continuously through to pupal stages [20]. These obser-vations indicate that the precise time of entry into thequiescent state isnot specified bya globally acting signal [2].
Several lines of argument suggest that cell intrinsic‘counting’ mechanisms might be important for specifyingthe number of divisions a neuroblast undergoes in theembryo before enteringquiescence. For example, embryonicneuroblasts cultured in vitro produce approximately thesame number of progeny as would normally be generatedduring embryogenesis in vivo [21], and many neuroblaststransplanted heterotopically develop a lineage consistentwith their original anteroposterior location [16,22]. Oneimportant system regulating the number of neuroblastdivisions is a temporal series of transcription factors thatlinks neuronal birth order with fate [23–26]. Neuroblastsexpress members of this set of nuclear proteins in acharacteristic sequence. Each GMC then maintains theexpression of the particular factor present at the time ofbudding from the neuroblast, thus providing it with atemporal label. The combined effect of the temporaltranscription-factor series and the grid of anteroposteriorand dorsoventral positional information has the potentialto provide every GMC within the embryonic CNS with aunique cell identity. Thus far, four members of thetranscription-factor series have been identified in theearly embryo and these are expressed in the order ofHunchback/Kruppel/Pdm1/Castor, although insome neuroblast lineages only subsets of this sequenceare expressed (Figure 1a). Transitions in the transcrip-tion-factor series depend on cell cycle progression andappear to be stabilized by negative cross-regulatoryinteractions [23,25]. In general, expression of factors inthe series is maintained in postmitotic progeny but thismight not be important for specifying their temporalidentity [26]. Studies in cultured neuroblasts suggest thatthe expression of a fifth transcription factor, Grainyhead,follows Castor [24,27]. However, the putative Castor-to-Grainyhead transition has not been documented withinthe developing embryo and it remains to be seen whetherGrainyhead specifies late GMC identity, as expected of atrue member of the series. Importantly, from the perspec-tive of this review, the transcription-factor series not onlyinfluences cell fate but also appears to be used as aninternal clock regulating the total number of neuroblastdivisions. When a neuroblast is forced to remain ‘youthful’by persistent expression of either Hunchback or Kruppel,additional GMCs of early identity are produced. In thecase of NB7-3, this produces an extended lineage, contain-ing many more cells than normal [25] (Figure 1b). Theseobservations suggest that embryonic neuroblasts mightexit the cell cycle and become quiescent only when theyhave finished switching through their normal temporalsequence of factors. This stopmechanism could involve thetranscription-factor series either directly activating
Antp
Ubx
AbdA
AbdB
Br
OL
T1
T2
T3
A
MB
Brain
T1–T3
A3-A7
Term
Pupa
96/0 ~ 96 h
+
Embryo
L1 L2 L3
0 22/0 24 48
Larva
OL
Br
TRENDS in Neurosciences
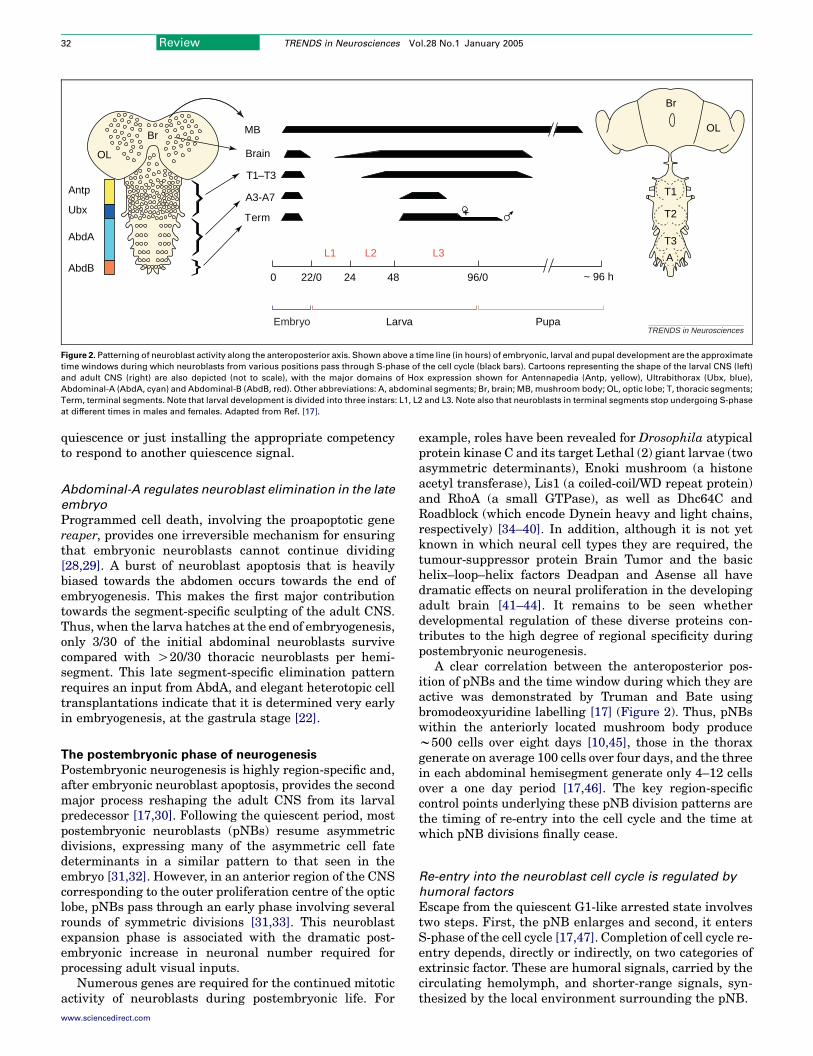
Figure 2. Patterning of neuroblast activity along the anteroposterior axis. Shown above a time line (in hours) of embryonic, larval and pupal development are the approximate
time windows during which neuroblasts from various positions pass through S-phase of the cell cycle (black bars). Cartoons representing the shape of the larval CNS (left)
and adult CNS (right) are also depicted (not to scale), with the major domains of Hox expression shown for Antennapedia (Antp, yellow), Ultrabithorax (Ubx, blue),
Abdominal-A (AbdA, cyan) and Abdominal-B (AbdB, red). Other abbreviations: A, abdominal segments; Br, brain; MB, mushroom body; OL, optic lobe; T, thoracic segments;
Term, terminal segments. Note that larval development is divided into three instars: L1, L2 and L3. Note also that neuroblasts in terminal segments stop undergoing S-phase
at different times in males and females. Adapted from Ref. [17].
Review TRENDS in Neurosciences Vol.28 No.1 January 200532
quiescence or just installing the appropriate competencyto respond to another quiescence signal.
Abdominal-A regulates neuroblast elimination in the late
embryo
Programmed cell death, involving the proapoptotic genereaper, provides one irreversible mechanism for ensuringthat embryonic neuroblasts cannot continue dividing[28,29]. A burst of neuroblast apoptosis that is heavilybiased towards the abdomen occurs towards the end ofembryogenesis. This makes the first major contributiontowards the segment-specific sculpting of the adult CNS.Thus, when the larva hatches at the end of embryogenesis,only 3/30 of the initial abdominal neuroblasts survivecompared with O20/30 thoracic neuroblasts per hemi-segment. This late segment-specific elimination patternrequires an input from AbdA, and elegant heterotopic celltransplantations indicate that it is determined very earlyin embryogenesis, at the gastrula stage [22].
The postembryonic phase of neurogenesis
Postembryonic neurogenesis is highly region-specific and,after embryonic neuroblast apoptosis, provides the secondmajor process reshaping the adult CNS from its larvalpredecessor [17,30]. Following the quiescent period, mostpostembryonic neuroblasts (pNBs) resume asymmetricdivisions, expressing many of the asymmetric cell fatedeterminants in a similar pattern to that seen in theembryo [31,32]. However, in an anterior region of the CNScorresponding to the outer proliferation centre of the opticlobe, pNBs pass through an early phase involving severalrounds of symmetric divisions [31,33]. This neuroblastexpansion phase is associated with the dramatic post-embryonic increase in neuronal number required forprocessing adult visual inputs.
Numerous genes are required for the continued mitoticactivity of neuroblasts during postembryonic life. For
www.sciencedirect.com
example, roles have been revealed for Drosophila atypicalprotein kinase C and its target Lethal (2) giant larvae (twoasymmetric determinants), Enoki mushroom (a histoneacetyl transferase), Lis1 (a coiled-coil/WD repeat protein)and RhoA (a small GTPase), as well as Dhc64C andRoadblock (which encode Dynein heavy and light chains,respectively) [34–40]. In addition, although it is not yetknown in which neural cell types they are required, thetumour-suppressor protein Brain Tumor and the basichelix–loop–helix factors Deadpan and Asense all havedramatic effects on neural proliferation in the developingadult brain [41–44]. It remains to be seen whetherdevelopmental regulation of these diverse proteins con-tributes to the high degree of regional specificity duringpostembryonic neurogenesis.
A clear correlation between the anteroposterior pos-ition of pNBs and the time window during which they areactive was demonstrated by Truman and Bate usingbromodeoxyuridine labelling [17] (Figure 2). Thus, pNBswithin the anteriorly located mushroom body producew500 cells over eight days [10,45], those in the thoraxgenerate on average 100 cells over four days, and the threein each abdominal hemisegment generate only 4–12 cellsover a one day period [17,46]. The key region-specificcontrol points underlying these pNB division patterns arethe timing of re-entry into the cell cycle and the time atwhich pNB divisions finally cease.
Re-entry into the neuroblast cell cycle is regulated by
humoral factors
Escape from the quiescent G1-like arrested state involvestwo steps. First, the pNB enlarges and second, it entersS-phase of the cell cycle [17,47]. Completion of cell cycle re-entry depends, directly or indirectly, on two categories ofextrinsic factor. These are humoral signals, carried by thecirculating hemolymph, and shorter-range signals, syn-thesized by the local environment surrounding the pNB.

Review TRENDS in Neurosciences Vol.28 No.1 January 2005 33
One humoral signal that is required to resume pNBactivity is a mitogen that is generated only when adequatenutrition is available. This point of regulation is pre-sumably linked to the ability of many insects to enter areversible low-metabolic state known as diapause, thusarresting larval development during periods of starvation.Co-culture experiments indicate that a source of thepostembryonic CNS mitogen is provided by a specializedlipid-rich organ known as the fat body [48]. Insect fat bodyappears to perform similar functions to vertebrate liverand adipose tissue and is specialized for the synthesis andstorage of protein, lipid and glycogen [49], also acting as aphysiological sensor for amino acid levels [50]. Althoughthe molecular identity of the fat-body mitogen is not yetclear, its action appears to be temporally confined to anamino-acid-sensitive checkpoint controlling cell cycle re-entry [48]. Thus, the five pNBs in the brain that neverundergo quiescence continue to divide normally understarvation conditions. Moreover, if nutrition is withdrawnafter pNBs have left the quiescent period, then the normalnumber of postembryonic cell divisions can still becompleted [48]. Interestingly, this nutritional study alsoshowed that pNBs from different anteroposterior locationsrequire the fat-body mitogen at different times, correlat-ing with their previously described anterior-to-posteriorsequence of cell cycle re-entry.
A second humoral signal affecting pNB activity is20-hydroxyecdysone (20E), a steroid hormone coordinat-ing the timing of many different developmental transi-tions [51]. Although it is not yet clear whether ecdysteroidsact directly on neuroblasts rather than another inter-mediary cell type, pNBs are known to express a nuclearreceptor for ecdysteroids, EcR-B1 [52]. A combination ofin vivo mutant studies and organ culture experimentsindicates that 20E levels affect the speed of the pNB cellcycle but not the final number of divisions [53]. However,the possible functions of 20E during pNB reactivation andits relationship to the fat-body mitogen are less clear. InCNS explant studies, 20E cannot substitute for fat body inactivating pNBs from starved larvae [48] but it doesactivate central brain pNBs from non-starved larvae thatare more than ten hours old [54]. The additionalobservation that 20E is not sufficient for pNB activationin younger larvae [54] suggests that normal pNBactivation could require both signals, perhaps with fat-body mitogen providing the competence to respond to 20E.
Re-entry into the neuroblast cell cycle is regulated by a
stem cell niche
In addition to humoral signals, there is mounting evidencethat pNB activity in larvae is modulated by more localcues. By analogy with other stem cell systems, these arelikely to constitute a specialized cellular and molecularmicroenvironment known as a niche [55–57]. Although itis unclear whether such a niche exists for neuroblasts inthe early embryo, culture assays do suggest that embry-onic divisions can proceed without close proximity to othercell types [21,24]. For larval pNBs, however, one import-ant component of the stem cell niche is provided by anetwork of surface glia. Hence, although the DrosophilaE-cadherin homologue DE-cadherin is widely expressed
www.sciencedirect.com
by glia, pNBs and their progeny, blocking its functionspecifically in glia using a dominant-negative approachreduces the mitotic activity of central brain neuroblasts[58]. A possible role of cadherin-based cell adhesion withinthe pNB niche is to facilitate local glial-to-neuroblastsignalling. For example, it is known that glia secrete theglycoprotein Anachronism (Ana) and that this is requiredto keep central brain pNBs arrested in the quiescentstate for the appropriate period of time [59]. Release fromAna-mediated repression requires terribly reduced opticlobes (trol), a gene encoding a heparin sulfate proteoglycanrelated to vertebrate Perlecan [60,61]. The distribution ofTrol has yet to be characterized in detail but it is known tobe expressed by midline glia in the embryo [60,62]. Acombination of genetic interaction data, CNS culturestudies and coimmunoprecipitation experiments suggeststhat Trol could promote pNB progression from G1 toS-phase by facilitating Fibroblast growth factor (FGF) andHedgehog (Hh) signalling [61]. It has also been suggestedthat Trol could promote exit from quiescence by somehowsequestering Ana [60].
What then, is the relationship between humoral factors,Trol and Ana during pNB reactivation? There is evidence tosupport the notion that the Trol–Ana pathway acts on theG1-to-S transition downstream of pNB enlargement,whereas the nutritional checkpoint mediated by the fat-body mitogen precedes enlargement. For example, pNBs introlmutantbutnot starved larvae canbe forced intoS-phaseby ectopic expression of cyclin E [47,48,61].
pNB divisions are stopped in the abdomen by a pulse of
Hox protein
Once pNBs re-enter the cell cycle, what determines whenthey stop dividing? Several of the extrinsic factors thatregulate the duration and exit from quiescence such asAna, 20E and nutritional status do not appear to influencefinal pNB clone size [48,53,59]. This argues that a separ-ate mechanism is responsible for cell counting: stoppingeach pNB from dividing after the appropriate number ofprogeny has been generated. As previously suggested forthe embryo, this might involve counting cell divisions viaswitching through a transcription-factor series. Never-theless, it remains to be determined whether there areadditional transcription-factor transitions during post-embryonic neuroblast divisions.
One component of the postembryonic counting mech-anism could be the coiled-coil protein Mushroom bodydefect. Larvae lacking activity of this protein displaydefects in several CNS regions, including the abdomenwhere two of the three different types of pNB generatemany more progeny than normal by escaping quiescenceand also continuing to divide past the normal endpoint[63,64]. Although the genetic pathway governing cessa-tion of pNB divisions has yet to be worked out in detail, itis clear that region-specific inputs must be important.For the central abdomen, it is known that the Hox proteinAbdA is required to terminate pNB divisions at theappropriate time [46]. During the last larval instar, abrief pulse of AbdA expression within the three abdominalpNBs activates proapoptotic genes and thus triggersprecursor death. Neuroblast apoptosis, rather than
Larva
)
Abdominal
NBAbdA
Thoracic
NBUbx
Embryo
Quiescent NB
Cycling NB
Mitogenic niche signals (e.g. Trol, Hh, FGF)
Humoral mitogenic signals (e.g. FB-derived, 20E)
Apoptotic NBQuiescence niche signals (e.g. Ana)
Niche (e.g. glial)
Ubx expression
AbdA expression TRENDS in Neurosciences
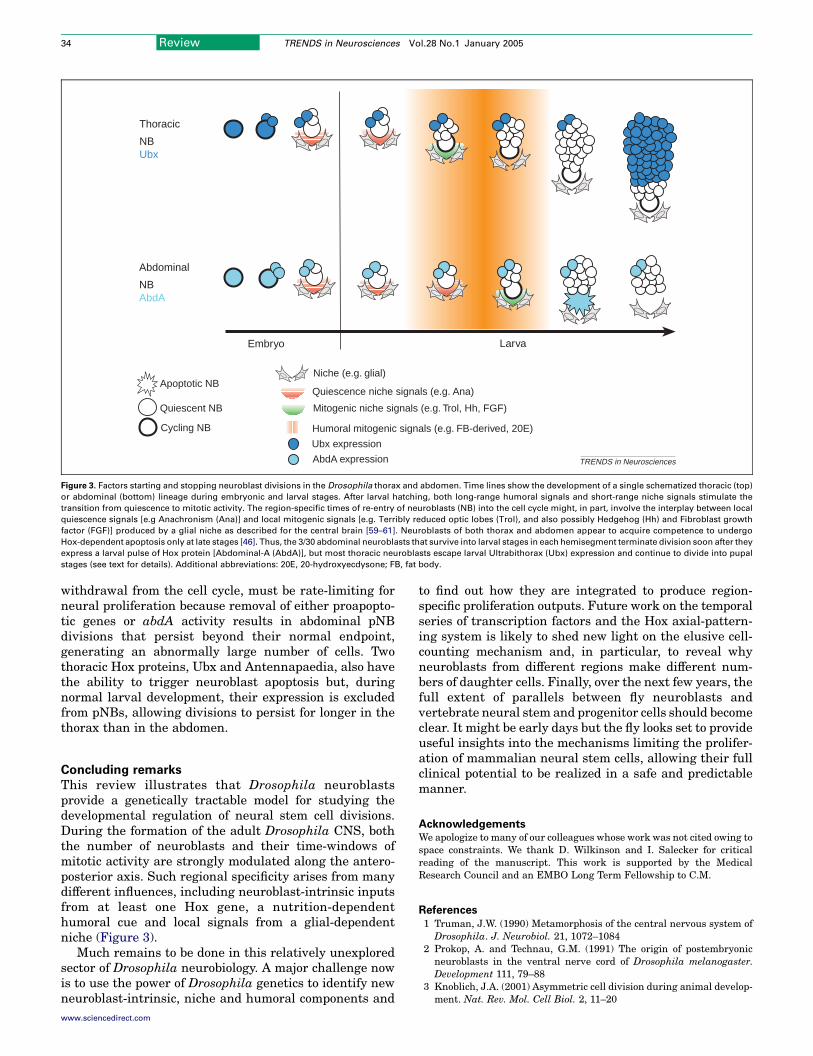
Figure 3. Factors starting and stopping neuroblast divisions in the Drosophila thorax and abdomen. Time lines show the development of a single schematized thoracic (top)
or abdominal (bottom) lineage during embryonic and larval stages. After larval hatching, both long-range humoral signals and short-range niche signals stimulate the
transition from quiescence to mitotic activity. The region-specific times of re-entry of neuroblasts (NB) into the cell cycle might, in part, involve the interplay between local
quiescence signals [e.g Anachronism (Ana)] and local mitogenic signals [e.g. Terribly reduced optic lobes (Trol), and also possibly Hedgehog (Hh) and Fibroblast growth
factor (FGF)] produced by a glial niche as described for the central brain [59–61]. Neuroblasts of both thorax and abdomen appear to acquire competence to undergo
Hox-dependent apoptosis only at late stages [46]. Thus, the 3/30 abdominal neuroblasts that survive into larval stages in each hemisegment terminate division soon after they
express a larval pulse of Hox protein [Abdominal-A (AbdA)], but most thoracic neuroblasts escape larval Ultrabithorax (Ubx) expression and continue to divide into pupal
stages (see text for details). Additional abbreviations: 20E, 20-hydroxyecdysone; FB, fat body.
Review TRENDS in Neurosciences Vol.28 No.1 January 200534
withdrawal from the cell cycle, must be rate-limiting forneural proliferation because removal of either proapopto-tic genes or abdA activity results in abdominal pNBdivisions that persist beyond their normal endpoint,generating an abnormally large number of cells. Twothoracic Hox proteins, Ubx and Antennapaedia, also havethe ability to trigger neuroblast apoptosis but, duringnormal larval development, their expression is excludedfrom pNBs, allowing divisions to persist for longer in thethorax than in the abdomen.
Concluding remarks
This review illustrates that Drosophila neuroblastsprovide a genetically tractable model for studying thedevelopmental regulation of neural stem cell divisions.During the formation of the adult Drosophila CNS, boththe number of neuroblasts and their time-windows ofmitotic activity are strongly modulated along the antero-posterior axis. Such regional specificity arises from manydifferent influences, including neuroblast-intrinsic inputsfrom at least one Hox gene, a nutrition-dependenthumoral cue and local signals from a glial-dependentniche (Figure 3).
Much remains to be done in this relatively unexploredsector of Drosophila neurobiology. A major challenge nowis to use the power of Drosophila genetics to identify newneuroblast-intrinsic, niche and humoral components and
www.sciencedirect.com
to find out how they are integrated to produce region-specific proliferation outputs. Future work on the temporalseries of transcription factors and the Hox axial-pattern-ing system is likely to shed new light on the elusive cell-counting mechanism and, in particular, to reveal whyneuroblasts from different regions make different num-bers of daughter cells. Finally, over the next few years, thefull extent of parallels between fly neuroblasts andvertebrate neural stem and progenitor cells should becomeclear. It might be early days but the fly looks set to provideuseful insights into the mechanisms limiting the prolifer-ation of mammalian neural stem cells, allowing their fullclinical potential to be realized in a safe and predictablemanner.
AcknowledgementsWe apologize to many of our colleagues whose work was not cited owing tospace constraints. We thank D. Wilkinson and I. Salecker for criticalreading of the manuscript. This work is supported by the MedicalResearch Council and an EMBO Long Term Fellowship to C.M.
References
1 Truman, J.W. (1990) Metamorphosis of the central nervous system ofDrosophila. J. Neurobiol. 21, 1072–1084
2 Prokop, A. and Technau, G.M. (1991) The origin of postembryonicneuroblasts in the ventral nerve cord of Drosophila melanogaster.Development 111, 79–88
3 Knoblich, J.A. (2001) Asymmetric cell division during animal develop-ment. Nat. Rev. Mol. Cell Biol. 2, 11–20

Review TRENDS in Neurosciences Vol.28 No.1 January 2005 35
4 Jan, Y.N. and Jan, L.Y. (2001) Asymmetric cell division in theDrosophila nervous system. Nat. Rev. Neurosci. 2, 772–779
5 Chia, W. and Yang, X. (2002) Asymmetric division of Drosophilaneural progenitors. Curr. Opin. Genet. Dev. 12, 459–464
6 Skeath, J.B. and Thor, S. (2003) Genetic control of Drosophila nervecord development. Curr. Opin. Neurobiol. 13, 8–15
7 Bossing, T. et al. (1996) The embryonic central nervous systemlineages of Drosophila melanogaster. I. Neuroblast lineages derivedfrom the ventral half of the neuroectoderm. Dev. Biol. 179, 41–64
8 Schmidt, H. et al. (1997) The embryonic central nervous systemlineages of Drosophila melanogaster. II. Neuroblast lineagesderived from the dorsal part of the neuroectoderm. Dev. Biol.189, 186–204
9 Schmid, A. et al. (1999) Clonal analysis of Drosophila embryonicneuroblasts: neural cell types, axon projections and muscle targets.Development 126, 4653–4689
10 Ito, K. et al. (1997) The Drosophila mushroom body is a quadruplestructure of clonal units each of which contains a virtually identicalset of neurones and glial cells. Development 124, 761–771
11 Urbach, R. et al. (2003) The pattern of neuroblast formation, mitoticdomains and proneural gene expression during early brain develop-ment in Drosophila. Development 130, 3589–3606
12 Urbach, R. and Technau, G.M. (2003) Segment polarity and DVpatterning gene expression reveals segmental organization of theDrosophila brain. Development 130, 3607–3620
13 Udolph, G. et al. (1993) A common precursor for glia and neurons inthe embryonic CNS of Drosophila gives rise to segment-specificlineage variants. Development 118, 765–775
14 Mann, R.S. and Morata, G. (2000) The developmental and molecularbiology of genes that subdivide the body ofDrosophila.Annu. Rev. CellDev. Biol. 16, 243–271
15 McGinnis, W. and Krumlauf, R. (1992) Homeobox genes and axialpatterning. Cell 68, 283–302
16 Prokop, A. and Technau, G.M. (1994) Early tagma-specific commitmentof Drosophila CNS progenitor NB1-1. Development 120, 2567–2578
17 Truman, J.W. and Bate, M. (1988) Spatial and temporal patterns ofneurogenesis in the central nervous system of Drosophila melano-gaster. Dev. Biol. 125, 145–157
18 Li, L. and Vaessin, H. (2000) Pan-neural Prospero terminates cellproliferation duringDrosophila neurogenesis. Genes Dev. 14, 147–151
19 Liu, T.H. et al. (2002) Transcription of theDrosophilaCKI gene dacapois regulated by a modular array of cis-regulatory sequences. Mech.Dev. 112, 25–36
20 Ito, K. and Hotta, Y. (1992) Proliferation pattern of postembryonicneuroblasts in the brain of Drosophila melanogaster. Dev. Biol. 149,134–148
21 Furst, A. and Mahowald, A.P. (1985) Cell division cycle of culturedneural precursor cells from Drosophila. Dev. Biol. 112, 467–476
22 Prokop, A. et al. (1998) Homeotic regulation of segment-specificdifferences in neuroblast numbers and proliferation in the Drosophilacentral nervous system. Mech. Dev. 74, 99–110
23 Kambadur, R. et al. (1998) Regulation of POU genes by castor andhunchback establishes layered compartments in the Drosophila CNS.Genes Dev. 12, 246–260
24 Brody, T. and Odenwald, W.F. (2000) Programmed transformations inneuroblast gene expression during Drosophila CNS lineage develop-ment. Dev. Biol. 226, 34–44
25 Isshiki, T. et al. (2001) Drosophila neuroblasts sequentially expresstranscription factors which specify the temporal identity of theirneuronal progeny. Cell 106, 511–521
26 Pearson, B.J. and Doe, C.Q. (2003) Regulation of neuroblastcompetence in Drosophila. Nature 425, 624–628
27 Uv, A.E. et al. (1997) Tissue-specific splicing and functions of theDrosophila transcription factor Grainyhead. Mol. Cell. Biol. 17,6727–6735
28 White, K. et al. (1994) Genetic control of programmed cell death inDrosophila. Science 264, 677–683
29 Peterson, C. et al. (2002) reaper is required for neuroblast apoptosisduring Drosophila development. Development 129, 1467–1476
30 White, K. and Kankel, D.R. (1978) Patterns of cell division and cellmovement in the formation of the imaginal nervous system ofDrosophila melanogaster. Dev. Biol. 65, 296–321
www.sciencedirect.com
31 Ceron, J. et al. (2001) Patterns of cell division and expression ofasymmetric cell fate determinants in postembryonic neuroblastlineages of Drosophila. Dev. Biol. 230, 125–138
32 Akong, K. et al. (2002) Drosophila APC2 and APC1 have overlappingroles in the larval brain despite their distinct intracellular localiza-tions. Dev. Biol. 250, 71–90
33 Hofbauer, A. and Campos-Ortega, J.A. (1990) Proliferation patternand early differentiation of the optic lobes inDrosophila melanogaster.Rouxs Arch. Dev. Biol. 198, 308–325
34 Rolls, M.M. et al. (2003) Drosophila aPKC regulates cell polarityand cell proliferation in neuroblasts and epithelia. J. Cell Biol. 163,1089–1098
35 Betschinger, J. et al. (2003) The Par complex directs asymmetric celldivision by phosphorylating the cytoskeletal protein Lgl. Nature 422,326–330
36 Gateff, E. (1978) Malignant neoplasms of genetic origin in Drosophilamelanogaster. Science 200, 1448–1459
37 Scott, E.K. et al. (2001) enok encodes a Drosophila putative histoneacetyltransferase required for mushroom body neuroblast prolifer-ation. Curr. Biol. 11, 99–104
38 Lee, T. et al. (2000) Essential roles of Drosophila RhoA in theregulation of neuroblast proliferation and dendritic but not axonalmorphogenesis. Neuron 25, 307–316
39 Liu, Z. et al. (2000) Drosophila Lis1 is required for neuroblastproliferation, dendritic elaboration and axonal transport. Nat. CellBiol. 2, 776–783
40 Reuter, J.E. et al. (2003) A mosaic genetic screen for genes necessaryforDrosophilamushroom body neuronal morphogenesis.Development130, 1203–1213
41 Arama, E. et al. (2000) Mutations in the beta-propeller domain of theDrosophila brain tumor (brat) protein induce neoplasm in the larvalbrain. Oncogene 19, 3706–3716
42 Frank, D.J. et al. (2002) The Drosophila melanogaster gene braintumor negatively regulates cell growth and ribosomal RNA synthesis.Development 129, 399–407
43 Wallace, K. et al. (2000) The pan-neural bHLH proteins DEADPANand ASENSE regulate mitotic activity and cdk inhibitor dacapoexpression in the Drosophila larval optic lobes. Genesis 26, 77–85
44 Loop, T. et al. (2004) Transcriptional signature of an adult brain tumorin Drosophila. BMC Genomics 5, 24
45 Lee, T. et al. (1999) Development of the Drosophila mushroom bodies:sequential generation of three distinct types of neurons from aneuroblast. Development 126, 4065–4076
46 Bello, B.C. et al. (2003) A pulse of the Drosophila Hox proteinAbdominal-A schedules the end of neural proliferation via neuroblastapoptosis. Neuron 37, 209–219
47 Caldwell, M.C. and Datta, S. (1998) Expression of cyclin E or DP/E2Frescues the G1 arrest of trol mutant neuroblasts in the Drosophilalarval central nervous system. Mech. Dev. 79, 121–130
48 Britton, J.S. and Edgar, B.A. (1998) Environmental control of the cellcycle in Drosophila: nutrition activates mitotic and endoreplicativecells by distinct mechanisms. Development 125, 2149–2158
49 Dean, R.L. et al. (1985) Structure of the fat body. In ComprehensiveInsect Physiology, Biochemistry and Pharmacology (Kerkut, G.A. andGilbert, L.I. eds), pp. 155–210, Pergamonn Press
50 Colombani, J. et al. (2003) A nutrient sensor mechanism controlsDrosophila growth. Cell 114, 739–749
51 Riddiford, L.M. (1993) Hormones and Drosophila Development. InThe Development of Drosophila melanogaster (Vol. II) (Bate, M. andMartinez Arias, A., eds.), pp. 899–939, Cold Spring Harbor LaboratoryPress
52 Truman, J.W. et al. (1994) Ecdysone receptor expression in the CNScorrelates with stage-specific responses to ecdysteroids duringDrosophila and Manduca development. Development 120, 219–234
53 Truman, J.W. et al. (1993) Formation of the adult nervous system. TheDevelopment of Drosophila melanogaster (Vol. II) (Bate, M. andMartinez Arias, A., eds.), pp. 1245–1276, Cold Spring HarborLaboratory Press
54 Datta, S. (1999) Activation of neuroblast proliferation in explantculture of the Drosophila larval CNS. Brain Res. 818, 77–83
55 Spradling, A. et al. (2001) Stem cells find their niche. Nature 414,98–104

Review TRENDS in Neurosciences Vol.28 No.1 January 200536
56 Fuchs, E. et al. (2004) Socializing with the neighbors: stem cells andtheir niche. Cell 116, 769–778
57 Wurmser, A.E. et al. (2004) Neuroscience. Cellular interactions in thestem cell niche. Science 304, 1253–1255
58 Dumstrei, K. et al. (2003) Role of DE-cadherin in neuroblastproliferation, neural morphogenesis, and axon tract formation inDrosophila larval brain development. J. Neurosci. 23, 3325–3335
59 Ebens, A.J. et al. (1993) The Drosophila anachronism locus: aglycoprotein secreted by glia inhibits neuroblast proliferation. Cell74, 15–27
60 Voigt, A. et al. (2002) Perlecan participates in proliferation activationof quiescent Drosophila neuroblasts. Dev. Dyn. 224, 403–412
61 Park, Y. et al. (2003) Drosophila perlecan modulates FGF and
Endea
the quarterly magaziand philosophy
You can access EndeScienceDirect, whecollection of beaut
articles on the historreviews and edito
Featur
Sverre Petterssen and the Contentious (and Momentous) WFood of Paradise: Tahitian breadfruit and the Autocritique
Two Approaches to Etiology: The Debate Over SmokinSicily, or sea of tranquility? Mapping a
The Prehistory of the PerioTwo portraits of Edmo
and comin
Fighting the ‘microbe of sporting mania’: Australian scienceby P. Ro
Learning from Education to Communicate ScienceThe Traffic and Display of Body Parts in the Ear
The Rise, Fall and Resurrection ofPomet’s great ‘‘Compleat Histo
Sherlock Holmes: scientificThe Future of Electricity in
The First Personal CompBaloonmania: news in
and much, muc
Locate Endeavour on ScienceDirect
www.sciencedirect.com
hedgehog signals to activate neural stem cell division. Dev. Biol.
253, 247–25762 Friedrich, M.V. et al. (2000) Perlecan domain V of Drosophila
melanogaster. Sequence, recombinant analysis and tissue expression.Eur. J. Biochem. 267, 3149–3159
63 Prokop, A. and Technau, G.M. (1994) Normal function of themushroom body defect gene of Drosophila is required for the
regulation of the number and proliferation of neuroblasts. Dev. Biol.
161, 321–33764 Guan, Z. et al. (2000) Mushroom body defect, a gene involved in the
control of neuroblast proliferation in Drosophila, encodes a coiled-coilprotein. Proc. Natl. Acad. Sci. U. S. A. 97, 8122–8127
vour
ne for the historyof science
avour online viare you’ll find aifully illustratedy of science, bookrial comment.
ing
eather Forecasts for D-Day, 6 June 1944 by J.R. Flemingof European Consumption by P. White and E.C. Spary
g and Lung Cancer in the 1950s by M. Parascandoland naming the moon by J. Vertesi
dic Table by D. Rouvraynd Halley by P. Fara
g soon
and Antarctic exploration in the early twentieth centurybertsas a Good Story by A. Negrete and C. Lartiguely-19th Century by S. Alberti and S. ChaplinGroup Selection by M. Borrellory of Drugs’’ by S. Shermandetective by L. Snyder1892 by G.J.N. Goodayuter by J. Novemberthe air by M.G. Kim
h more . . .
(http://www.sciencedirect.com)