Embed Size (px)
Citation preview

REFERENCECOPY Do Not Remove from the Library
U. S. Fish and Wildlife Service
Biological Report 82(11.100) TR EL-82-4 March 1989 700 Cajun Dome Boulevard
Lafayette, Louisiana 70506
Species Profiles: Life Histories and Environmental Requirements of Coastal Fishes and Invertebrates ( Mid-Atlantic)
BLUE CRAB
Coastal Ecology Group Fish and Wildlife Service Waterways ~xperiment Station
U.S. Department of the Interior U.S. Army Corps of Engineers

Bi 01 ogi cal Report 82( 11.100) TR EL-82-4 March 1989
Species Pro f i les : L i f e H i s t o r i e s and Environmental Requirements o f Coastal Fishes and Inver tebra tes (Mid-At lan t ic )
BLUE CRAB
Jenn i fe r H i l l , Dean L. Fowler,
and Michael J. Van Den Avyle
Georgia Cooperative F i sh and W i l d l i f e Research U n i t School o f Forest Resources
U n i v e r s i t y o f Georgia Athens, GA 30602
P r o j e c t O f f i c e r David Moran
U.S. F ish and W i l d l i f e Serv ice Nat ional Wetlands Research Center
1010 Gause Boulevard S l i d e l 1 , LA 70458
Performed f o r U.S. Army Corps o f Engineers
Coastal Ecology Group Waterways Experiment S ta t i on
Vicksburg, MS 39180
and
U.S. Department o f the I n t e r i o r F ish and Wi ld l i f e Serv ice Research and Development
Nat iona l Wetlands Research Center Washington, DC 20240

DISCLAIMER
The mention o f t rade nmes i n t h i s r e p o r t does n o t c o n s t i t u t e endorsement nor recommendation f o r use by t h e U.S. Fish and W i l d l i f e Service o r Federal Government.
Th is se r ies may be referenced as fo l lows:
U.S. F i sh and W i l d l i f e Service. 1983-19 . Species p r o f i l e s : l i f e h i s t o r i e s and environmental requirements o f coasta l f i s h e s and inver tebra tes . U .S . F ish W i l d l . Serv. B i o l . Rep. 82(11). U.S. Army Corps o f Engineers, TR EL-82-4.
Th is p r o f i l e may be c i t e d as fo l lows:
H i l l , J., D.L. Fowler, and M.J. Van Den Avyle. 1989. Species p r o f i l e s : l i f e h i s t o r i e s and environmental requirements o f coastal f i shes and inver tebra tes (Mid-At lant ic)- -Blue crab. U.S. F i sh W i l d l . Serv. B i o l . Rep. 82(11.100). U.S. Army Corps o f Engineers, TR EL-82-4. 18 pp.

PREFACE
This species p r o f i l e i s one o f a se r ies on coasta l aquatic organisms, p r i n c i p a l l y f i s h , o f sport, commercial, o r ecological importance. The p r o f i l e s are designed t o provide coasta l managers, engineers, and b i o l o g i s t s w i t h a b r i e f comprehensive sketch o f t h e b i o l ogi cal charac ter i s t i c s and envi ronmental requirements o f the species and t o descr ibe how populat ions of t he species may be expected t o r e a c t t o environmental changes caused by coastal devel opnent. Each p r o f i l e has sections on taxonow, l i f e h i s tory , eco log ica l r o l e , environmental requ i rements, and econmi c importance, i f appl i cab1 e. A t h r e e - r i ng b inder i s used f o r t h i s ser ies so t h a t new p r o f i l e s can be added as they are prepared. Th is p r o j e c t i s j o i n t l y planned and f inanced by t h e U.S. Army Corps o f Engineers and the U.S. Fish and W i l d l i f e Service.
M i l l i k i n and Wil l iams (1984) previously publ ished a review o f t he nomenclature, taxononly, morphology, d i s t r i b u t i o n , l i f e h i s t o r y , popu la t ion s t r u c t u r e and dynamics, and t h e f i s h e r y o f t he b lue crab.
Suggestions o r questions regard ing t h i s r e p o r t should be d i rec ted t o one o f t he f o l 1 owing addresses.
In format ion Transfer Special i s t National Wetlands Research Center U.S. F i sh and W i l d l i f e Serv ice NASA-Sl i d e l 1 Computer Complex 1010 Gause Boulevard S l i d e l l , LA 70458
U .S. Army Engi neer Waterways Experiment S t a t i o n At tent ion : WESER-C Post O f f i c e Box 631 Vicksburg, MS 39180.

CONVERSION TABLE
M e t r i c t o U . S . Customary
M u l t i p l y
m i l l i m e t e r s (mm) cen t imete rs (cm) meters (m) meters (m) k i 1 ometers (km) k i l omete rs (km)
square meters (m2) 10.76 square k i l o m e t e r s (km2) 0.3861 hectares (ha) 2.471
l i t e r s (1) cub ic meters (m3) cub ic meters (m3)
m i l l i g r a m s (mg) grams (g) k i lograms (kg) m e t r i c tons (t) m e t r i c tons (t)
k i l o c a l o r i e s ( k c a l ) Ce ls ius degrees (OC)
inches inches f e e t ( f t ) fathoms s t a t u t e m i l e s (mi) n a u t i c a l m i l e s (nmi )
square f e e t ( f t2) square mi 1 es ( m i 2,
acres
g a l l o n s ( g a l ) cub ic f e e t ( f t3) acre - fee t
ounces (oz) ounces (oz) pounds ( l b ) pounds ( l b ) s h o r t tons ( ton )
B r i t i s h thermal u n i t s (Btu) Fahrenhei t degrees (OF)
U . S . Customary t o M e t r i c
25.40 2.54 0.3048 1.829 1.609 1.852
To Obta in
i nches inches f e e t fathoms s t a t u t e m i l e s n a u t i c a l m i l e s
square f e e t square m i l e s acres
ga l 1 ons cub ic f e e t acre- f e e t
ounces ounces pounds pounds s h o r t tons
B r i t i s h thermal u n i t s Fahrenhe i t degrees
m i l l i m e t e r s cen t imete rs meters meters k i 1 ometers k i 1 ometers
square meters square k i lometers hectares
1 i t e r s cub ic meters cub ic meters
mi 11 igrams grams k i 1 ograms m e t r i c tons m e t r i c tons
k i l o c a l o r i e s Cel s i us degrees

CONTENTS
Page
PREFACE .................................................................... iii CONVERSION TABLE ........................................................... i v ACKNOWLEDGMENTS ............................................................ v i
NOMENCLATURE. TAXONOMY. AND RANGE .......................................... 1 MORPHOLOGY AND IDENTIFICATION AIDS ......................................... 1 REASON FOR INCLUSION I N SERIES ............................................. 3 LIFE HISTORY ............................................................... 3
Ma t i ng and Spawning ...................................................... 3 Development .............................................................. 4 Eggs ..................................................................... 4 Larvae ................................................................... 4 Juven i l es ................................................................ 5 Adu l t s ................................................................... 6 M i g r a t i o n s ............................................................... 6
GROWTH AND MOLTING CHARACTERISTICS ......................................... 6 THE FISHERY ................................................................ 7 ECOLOGICAL ROLE ............................................................ 9 ................................................. ENVIRONMENTAL REQUIREMENTS 11 .............................................................. Temperature 11 ................................................................. S a l i n i t y 11
Tempera tu re -Sa l in i t y I n t e r a c t i o n s ........................................ 11 H a b i t a t .................................................................. 12 .............................................. Other Envi ronmenta l Fac to rs 12
LITERATURE CITED ........................................................ 13

ACKNOWLEDGMENTS
We are g r a t e f u l t o Maurice K. Crawford, Douglas E. Facey, and Timothy J. Welch f o r reviews and t o Sue Anthony f o r e d i t i n g and t yp ing t h i s manuscript. Technical reviews were provided by Kenneth Tenore o f the U n i v e r s i t y o f Maryland and W. A. Van Engel o f the V i r g i n i a I n s t i t u t e o f Marine Science a t Gloucester Point . We wish t o thank the authors J. D. Costlow, Jr., and C. G. Bookhout f o r use o f t h e i r drawing o f the Zoea f o r our Figure 4.

Figure 1. Rlue Crab.
BLUE CRAB
NOMENCLATURE ,TAXONOMY, AND RANGE
Scientific name.. . .Callinectes sapidus (Rathbun)
Preferred common name. ...... .Bl ue crab Other common names.. ... .Edible crab,
crab; young females are called sally crabs; adult females are called sooks; and males are called jimies, jirnmydicks , or channelers (Van Engel 1958)
Class ........................ Crustacea Order ......................... Decapoda Family.. ................... .Portunidae Subfamily ................... Portuninae Geographic range: The blue crab
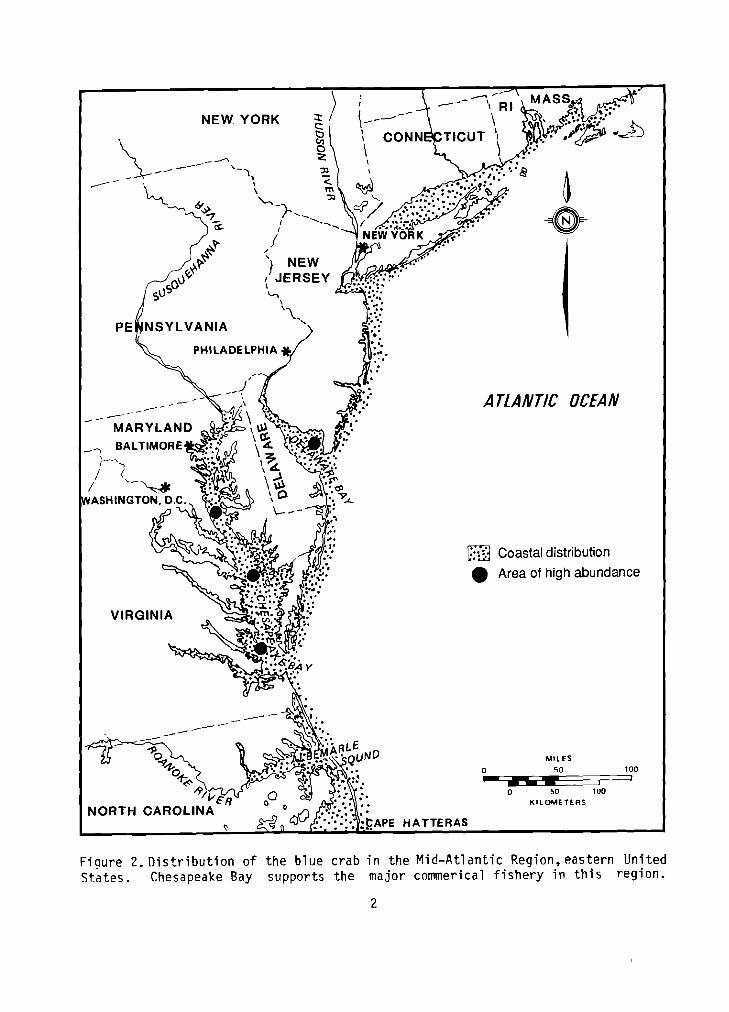
(Figure 1) occurs in coastal waters--primarily bays and brackish estuaries--from Massachusetts Bay southward to the eastern coast of
South America, including the Gulf of Mexico. It has been collected occasionally in Maine and northward to Nova Scotia (Piers 1923; Scatter- good 1960). It occurs throughout the Mid-Atlantic Region (Figure 2) (Sullivan 1909; Churchill 1919; Van Engel 1958).
MORPHOLOGY AND IDENTIFICATION AIDS
The blue crab is grayish or bluish green, with red on carapace spines (Will iams 1965). Pincers on the chelipeds are blue in males and red in mature females. Underparts are off-white with tints of yellow and pink (Williams 1965). Young crabs often are brownish with conspicuous white markings (Newcombe 1945).
ATLANTIC OCEAN
NEW YORK
PHILADELPHIA
Coastal distribution Area of high abundance
K I L O M E T E R S
F iqu re 2. D i s t r i b u t i o n o f t h e b l u e c rab i n t h e M i d - A t l a n t i c Region,eastern Un i t ed S ta tes . Chesapeake Bay suppor ts t h e ma jo r commerical f i s h e r y i n t h i s reg ion .
2

Although t h e s e a r e t y p i c a l c o l o r pat- t e r n s , r a r e forms have been cap tured t h a t were e n t i r e l y blue (Maryland Tidewater News 1950 ; Haefner 1961).
The carapace , inc lud ing l a t e r a l sp ines , i s u s u a l l y 2.5 times a s wide a s long , moderately convex, and nea r ly smooth, except f o r small t u b e r c l e s on i nne r branchia l and c a r d i a c reg ions . The a n t e r i o r margin of t h e carapace has a median o r f r o n t a l region t h a t extends between t h e compound eyes , and two 1 a t e r a l reg ions .
Members of t h e subfamily Portuninae have n ine a n t e r o l a t e r a l t e e t h on t h e carapace , with t h e l a t e r a l t oo th s i m i l a r t o o t h e r s i n s i z e . The genus Cal l i n e c t e s has an antenna t h a t i s no t excluded from t h e o r b i t ; male C a l l i n e c t e s a l s o have a T-shaped abdomen. The abdomen i s t r i a n g u l a r i n immature females and i s broadly rounded and fo lded 1 oose ly a g a i n s t t h e ven t r a l s i d e of t h e t h o r a c i c s t e r n a i n mature females (F igure 3 ) . The medial reg ion of C . sap idus has two f r o n t a l t e e t h between t he i nne r o r b i t a l t e e t h (Williams 1984).
REASON FOR INCLUSION IN SERIES
The b lue c r ab suppor t s a va luable commercial f i s h e r y throughout t h e
Figure 3 . Ventral view of t h e blue c rab male (A), immature female (B), and mature female (C) ( T r u i t t 1939).
!!id-Atlantic S t a t e s and along most of t h e e a s t e r n and gu l f c o a s t s of t h e United S t a t e s . E s t u a r i e s a r e essen- t i a l i n i t s l i f e h i s t o r y ; t h e s p e c i e s ' high abundance i n e s t u a r i e s and i t s omnivorous feed ing h a b i t s sugges t t h a t i t p lays an important r o l e i n t h e s t r u c t u r e and func t i on of e s t u a r i n e communities. I t i s a p reda tor of commercially va luable clams and o y s t e r s (Newcombe 1945), and young a r e preyed upon by a l a r g e number of e s t u a r i n e and marine animals , inc lud- ing o t h e r c o m e r c i a l l y important spec i e s such a s t h e s t r i p e d bas s , Morone s a x a t i l i s (Manooch 1973).
LIFE HISTORY
Mating and Spawning
Blue c r abs mate i n Chesapeake Bay from May through October (Van Engel 1958; Williams 1984). The spawning season was s i g n i f i c a n t l y s h o r t e r dur ing yea r s i n which temperatures were low f o r extended per iods (Daugherty 1952). Mating occurs p r imar i l y i n r e l a t i v e l y l ow-sa l i n i t y waters i n t h e upper a r e a s of e s t u a r i e s and lower por t ions of r i v e r s (Py l e and Cronin 1950; Darnel 1 1959; Williams 1965; Tagatz 1968).
Will iams (1965) descr ibed t h e b lue c r a b ' s mating behavior . The male may mate dur ing i t s t h i r d o r f o u r t h i n t e rmo l t phase a f t e r i t matures . Females mate on ly once i n t h e i r l i v e s , bu t t h e sperm from t h i s mating i s s t o r e d i n seminal recep- t a c l e s and may be used a s o f t en a s t h e female spawns, g e n e r a l l y two o r more t imes dur ing a 1- o r 2-year per iod (Van Engel 1958; Will iams 1965). When the female i s ready t o molt i n t o t h e mature s t a g e , i t i s c a r r i e d under t h e ma le ' s body; t h e s e p a i r s arc? c a l l e d doublers . The female i s r e l ea sed dur ing mol t ing and i s then rec lasped wi th t h e abdomens f a c i n g each o the r . Spermatophores produced by t h e ma1 e a r e then passed v i a ccpu l a to ry s t y l e t s i n t o t h e spermathecae. Pleopods of

t h e male and swimmerets o f t h e female a re i n t r o m i t t e n t organs which a i d i n copu la t i on . A f t e r copu la t ion , t h e female i s tu rned around and c a r r i e d u n t i l her s h e l l hardens (Wi l l i ams 1984).
A f t e r mating, females m ig ra te t o h igh-sa l i n i t y waters i n lower es tua r i es , sounds, and nearshore spawning areas ( C h u r c h i l l 1919; Darnel 1 1959; F i s c h l e r and Wal burg 1962). These ove rw in te r be fo re spawning by burrowing i n t h e mud a t t h e mouths o f bays (Cook 1981; Schmidt 1985). Most females spawn f o r t h e f i r s t t ime 2 t o 9 months a f t e r ma t i ng ( C h u r c h i l l 1919; W i l l i ams 1965). 1965).
From May through August, t h e female ext rudes f e r t i l i z e d eggs i n t o a cohesive mass o r "sponge" t h a t remains at tached t o se tae on t h e appendages o f t h e abdomen u n t i l t h e l a r v a e emerge ( C h u r c h i l l 1919; Newcombe 1945; P y l e and Cronin 1950). The sponge, which may c o n t a i n 700,000 t o 2 m i l l i o n eggs, i s formed i n about 2 hours ( C h u r c h i l l 1919; T r u i t t 1939; W i l l iams 1965). I ncuba t i on genera l l y requ i res 1-2 weeks. I n Chesapeake Bay, l a r v a l r e l ease appears t o be concentrated between t h e V i r g i n i a capes (McConaugha e t a l . 1986). The presence o f empty egg cases on swimmerets o r t h e occurrence o f 1 arqe. b r i u h t - r e d a d u l t - . nemertean worms (cai-cinonemertes ca rc i no h i l a ) on t h e g i l l s o f a mature --?a- fema e i n i c a t e s t h a t t h e c rab has spawned a t l e a s t once ( C h u r c h i l l 1919; Hopkins 1947). A f t e r reach ing sexual m a t u r i t y , these worms feed on t h e egg masses c a r r i e d by female crabs and l i v e i n t h e g i l l s o f t h e c rab a f t e r t h e eggs ha tch (Hopkins 1947). I n l owe r Chesapeake Bav. mature r e d Carcinonemertes ' o c c u r r e i j n t he g i l l s o f more than 95% o f t h e female crabs t h a t had spawned; immature crabs supported o n l y immature, 1 i g h t - c o l ored worms (Hopkins 1947).
Development
Growth and development o f t h e b l u e crab, as i n o t h e r crustaceans, c o n s i s t o f a s e r i e s o f l a r v a l , j u v e n i l e , and a d u l t stages d u r i n g which a v a r i e t y o f morphologica l , behav io ra l , and phys io l og i ca l changes occur. These changes are most dramat ic when t h e animal m o l t s (sheds i t s r i g i d exoskeleton) p e r m i t t i n g growth and changes i n body shape. Before mo l t i ng , a new s h e l l i s formed beneath t h e o l d exoskeleton, which then loosens and i s c a s t o f f . The new s h e l l i s i n i t i a l l y s o f t , b u t i t expands and hardens i n a few hours. The stage between m o l t s i s termed i n t e r m o l t . Much o f t h e i n f o r m a t i o n summarized here was ob ta ined f rom comprehensive s tud ies o f t h e b l u e c rab i n t h e Chesapeake Bay area by C h u r c h i l l (1919), Newcombe (1945), and Van Engel (1958).
The eggs a re b r i g h t orange when f i r s t depos i ted , b u t become ye1 1 ow, brown, and then dark brown be fore ha t ch ing (Van Engel 1958). The c o l o r change i s caused by abso rp t i on o f t h e ye1 low yo1 k and development o f dark pigment i n t h e eyes and on t h e body. Eggs a re about 0.25 mm i n d iameter (Churchi 11 1919). Sandoz and Rogers (1944) repo r ted t h a t ha tch ing o f b l u e c rab eggs o n l y occurs a t s a l i n i t i e s o f 23-33 p p t and temperatures o f 19-29 O C . M o r t a l i t y o f eggs has been a t t r i b u t e d t o fungal i n f e c t i o n , p redat ion , s u f f o c a t i o n i n s tagnant water, and exposure t o extreme temperatures (Couch 1942 ; Humes 1942 ; Rogers-Tal b e r t 1948) . On t h e average, o n l y one o u t o f every m i l l i o n eggs su rv i ves t o become a mature a d u l t (Van Engel 1958).
Larvae
F i r s t s tage la rvae , c a l l e d zoeae, measure approx imate ly 0.25 mm wide a t ha t ch ing (F igu re 4 ) . C h u r c h i l l

Figure 4. Zoea (from Costlow, J r . and Bookhout 1959).
width; development t o t h i s s t a g e r e q u i r e s 31-49 days (Costlow and Bookhout 1959). The megalopa l a r v a , i l l u s t r a t e d by Church i l l (1919) and Newcombe (1945) , is more crab1 i k e i n appearance than t h e zoea; i t s carapace is broader i n r e l a t i o n t o i t s l e n g t h , and i t has p inch ing claws and po in ted j o i n t s a t t h e ends o f t h e l eg s . I t swims f r e e l y , b u t g e n e r a l l y s t a y s near bottom i n nearshore o r lower- e s t u a r i n e , high-sal i n i t y a r e a s (Tagatz 1968) . The megalopal s t a g e l a s t s 6 t o 20 days , a f t e r which t h e l a r v a mol t s i n t o t h e "first c rab" s t a g e , charac- t e r i z e d by a d u l t p ropo r t i ons and appearance.
Juveni 1 es
(1942) , Hopkins (1943) , and Costlow and Bookhout (1959) provided i l l u s - t r a t e d d e s c r i p t i o n s o f t h e morphology of zoeae. The l a r v a e bear l i t t l e morphological resemblance t o a d u l t s (Hopkins 1943) , a r e f i1 ter f e e d e r s , and a r e p lankton ic (Darnel 1 1959).
Evidence sugges t s t h a t b lue c r a b zoeae hatch i n Chesapeake Bay, Chinco- t eague Bay, Delaware Bay, and o t h e r e s t u a r i e s and d r i f t o u t t o s e a , where t hey f e e d and grow (Cook 1981; Finch- ham 1981; Su lk in e t a1. 1982). These l a r v a e may mig ra t e v e r t i c a l l y i n t h e wate r column t o reach f l o o d and ebb t i d e s , which t r a n s p o r t them back i n t o t h e bay a r e a .
The zoeae and a l l subsequent 1 i fe s t a g e s can i n c r e a s e body size on ly by mol t ing (Hay 1905) . Zoeal development depends on s a l i n i t y and tempera ture , b u t development time has been shown t o be v a r i a b l e even i n a s i n g l e s a l i n i t y - t empera ture regime (Costlow and Book- hout 1959). Larvae mol t seven t o e i g h t times be fo re e n t e r i n g t h e nex t s t a g e of development (Costlow and Bookhout 1959). The f i n a l mol t o f t h e zoea is c h a r a c t e r i z e d by a conspicuous change t o t h e second l a r v a l s t a g e (megalops) a t about 2.5 mm carapace
The j u v e n i l e "first c rab" is t y p i c a l l y 2.5 mm wide (from t i p t o t i p o f t h e l a t e r a l s p i n e s o f t h e ca r a - pace) . J u v e n i l e s gradual 1y mig ra t e i n t o sha l l ower , l e s s - s a l i n e wate rs i n upper e s t u a r i e s and r i v e r s , where t h e y grow and mature ( F i s c h l e r and Walburg 1962). Van Engel (1958) r epo r t ed t h a t many j u v e n i l e s had completed t h i s m ig ra t i on by fa1 1. New evidencel, however, sugges t s t h a t t h e bulk may n o t reach t h e upper p a r t s o f t r i b u - t a r i e s and Chesapeake Bay u n t i l t h e fo l lowing summer (W.A. Van Engel', V i rg in i a I n s t i t u t e o f b r i n e Sc ience , Glouces te r Point ; pers . cmm.) . Males, which p r e f e r 1 ow-sa l i n i t y w a t e r s , general l y m i g r a t e f a r t h e r upstream than do f ema le s , which tend t o s t a y i n t h e lower r i v e r s and e s t u a r i e s (Dudley and Judy 1971; Music 1979) .
Growth and matura t ion proceed dur ing a series o f mol t s and intermol t phases; each o f t h e s e c r a b s t a g e s is i d e n t i f i e d by t h e number o f mol t s t h a t have occur red s i n c e t h e megalopal s t a g e . Church i l l (1919) e s t ima t ed t h a t j u v e n i l e s reached t h e 9 t h o r 10 th c r a b s t a g e by October i n Chesapeake Bay. Molt ing and growth s t o p du r ing winter (Church i l l 1919; Darnel 1 1959) , b u t resume a s wate rs warm. The c r abs

generally reach maturity during the spring or summer of the year following the year of hatching.
Adults
In the Chesapeake Bay, sexual maturity i s reached a f t e r 18-20 postlarval molts, a t the age of 1-1.5 years (Van Engel 1958; W i 11 iams 1965). Males continue t o molt and grow a f t e r they reach sexual maturity, b u t females cease to molt and grow when they mature and mate.
After the females mate and migrate t o spawning areas , they e i t he r remain there f o r the r e s t of t h e i r l ives o r move only shor t distances out t o sea (Williams 1965). In warmer months, males generally s tay in 1 ow-sal i n i ty waters such as creeks, r i ve r s , and upper es tuar ies (Churchill 1919; Van Engel 1958; Dudley and Judy 1971; Music 1979). Early work by Fiedler (1930), Trui tt (1937), and Cronin (1949) on blue crabs i n the Chesapeake Bay indicated t ha t females overwintered a t the mouth of the bay and spawned there in spring, whereas the migration of males was nondirec- t iona l . Crabs bury themselves i n mud i n winter and emerge when temperatures r i s e i n spring (Cook 1981; Schmidt 1985). The maximum age f o r most blue crabs in the Mid-Atlantic Region i s 3 years (Churchill 1919; Mil 1 iams 1965) ; adul ts thus l i v e an average of l e s s than 1 year a f t e r reaching maturity (Hay 1905; Truitt 1939).
Migrations
Adult blue crabs a r e excel lent swimmers and a l so can move quickly on land. They ra re ly move from one es tuar ine system t o another (Por ter 1956; Cargo 1958; Fischler and Walburg 1962; Judy and Dudley 1970). When they do leave an es tuary, they usually remain i n adjacent coastal areas , though a few tagged female crabs have been recovered 100-540 bn from t h e i r release s i t e s .
Migrations of blue crabs w i t h i n es tuar ine systems a r e re la ted t o phases of t h e i r l i f e cycle , t o the season, and ( t o a l e s s e r extent) t o searches f o r favorable environmental conditions (Churchill 1919; Fiedler 1930; Truitt 1939; Fischler and Wal burg 1962). Most blue crabs move t o r e l a t i ve ly deeper, warmer waters i n winter and return t o r i ve r s , t i da l creeks, s a l t marshes, and sounds when conditions become more favorable in the spring (Livingston 1976; Subrahmanyam and Coul t as 1980).
GROWTH AND MOLTING CHARACTERISTICS
From the f i r s t crab stage t o the adul t , successive i n term01 t stages of the crab a re morphological l y s imi la r except f o r s i z e . The number of molts during ce r ta in l i f e stages (e.g., larval and juvenile) i s r e l a t i ve ly uniform among crabs, b u t the r a t e of molting (and hence growth) varies considerably and is affected by many environmental fac to rs , including temperature and sal i n i t y . Consequent- l y , i t i s generally not possible t o determine the developmental s tage of a par t i cu la r crab from i t s s i z e or external charac te r i s t i cs (Churchill 1919). Migrations and movements within es tuar ies fu r ther complicate estimation of growth r a t e s , and repeated sampling a t one location can lead t o erroneous conclusions (Darnel 1 1959; Adkins 1972; Palmer 1974). Thus, the growth and molting patterns described here were derived in par t from laboratory s tudies .
Crabs mol t often when small , b u t l e s s frequently as they grow la rger (Van Engel 1958). Each molt typ ica l ly r e su l t s in a 25%-40% increase i n carapace width (Churchi 11 1919; Gray and Newcombe 1939; Van Engel 1958). Resul t s of Churchi 11 ' s (1919) 1 abora- tory s tudies on growth of Chesapeake Bay blue crabs serve as a general guide1 ine f o r growth patterns of blue crabs i n Virginia and adjacent Sta tes
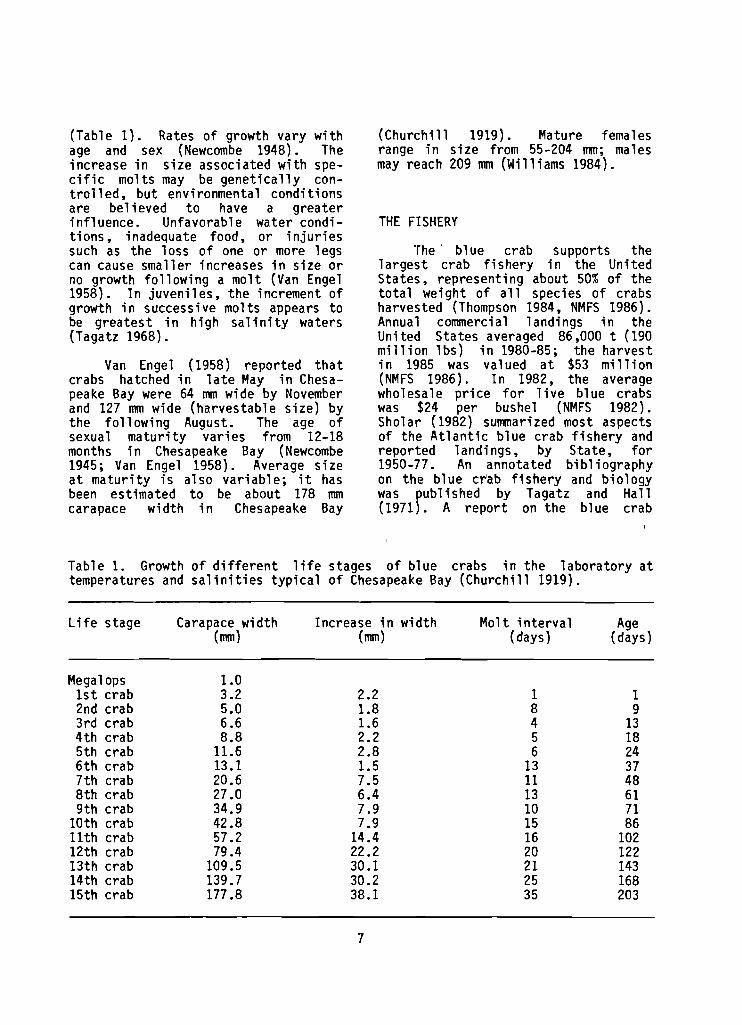
(Table 1) . Rates o f growth vary w i t h age and sex (Newcombe 1948). The increase i n s i z e associated w i t h spe- c i f i c mol ts may be g e n e t i c a l l y con- t r o l l e d , bu t environmental cond i t ions are be l ieved t o have a g rea te r i n f l uence . Unfavorable water condi - t i ons , inadequate food, o r i n j u r i e s such as the l oss o f one o r more legs can cause smal le r increases i n s i z e o r no growth f o l l o w i n g a m o l t (Van Engel 1958). I n juven i les , t h e increment o f growth i n successive mol ts appears t o be g rea tes t i n h igh s a l i n i t y waters (Tagatz 1968).
Van Engel (1958) repor ted t h a t crabs hatched i n l a t e May i n Chesa- peake Bay were 64 mm wide by November and 127 mm wide (harvestable s i z e ) by the f o l l o w i n g August. The age o f sexual m a t u r i t y va r i es from 12-18 months j n Chesapeake Bay (Newcombe 1945; Van Engel 1958). Average s i z e a t m a t u r i t y i s a l so var iab le ; i t has been est imated t o be about 178 mm carapace w id th i n Chesapeake Bay
(Church i l l 1919). Mature females range i n s i z e from 55-204 rnm; males may reach 209 mm ( W i 11 iams 1984).
THE FISHERY
T h e ' b l u e crab supports the l a r g e s t crab f i s h e r y i n t h e Un i ted States, represent ing about 50% o f t he t o t a l weight o f a l l species o f crabs harvested (Thompson 1984, NMFS 1986). Annual commercial landings i n the Uni ted States averaged 86,000 t (190 m i l 1 i o n 1 bs) i n 1980-85; t he harves t i n 1985 was valued a t $53 m i l l i o n (NMFS 1986). I n 1982, t he average wholesale p r i c e f o r l i v e b lue crabs was $24 per bushel (NMFS 1982). Sholar (1982) summarized most aspects o f t h e A t l a n t i c b l u e crab f i s h e r y and repor ted landings, by State, f o r 1950-77. An annotated b ib l i og raphy on t h e b l u e cr'ab f i s h e r y and b io logy was publ ished by Tagatz and H a l l (1971). A r e p o r t on t h e b lue crab
Table 1. Growth o f d i f f e r e n t l i f e stages o f b lue crabs i n t h e l abo ra to ry a t temperatures and s a l i n i t i e s t y p i c a l o f Chesapeake Bay (Church i l l 1919).
L i f e stage Carapace w id th Increase i n w id th Mol t i n t e r v a l Age (mm) (mm) (days) (days)
Mega1 ops 1 s t crab 2nd crab 3 rd crab 4 t h crab 5 t h crab 6 t h c rab 7 t h crab 8 t h crab 9 t h crab
10th crab 11th crab 12th crab 13th crab 14th crab 15th crab

dynamics i n Chesapeake Bay was pub- l i s h e d by the Chesapeake B io log ica l Laboratory (Jones e t a1 . 1983).
Harvests from the Mid-At lant ic Region dur ing 1977-85 composed about h a l f o f the t o t a l U.S. commercial b lue crab harvest; commercial landings were about 40,000 t o r 90 m i l l i o n I b s (NMFS 1982). Almost 90% o f commercial b lue crab landings i n the Mid-At lant ic are from the Chesapeake Bay reg ion i n Maryland and V i rg in ia ; landings are much smal ler i n Delaware and New Jersey (NMFS 1981, 1986). Much o f the harvest i s processed f o r commercial packaging s i n g u l a r l y o r mixed w i t h o ther food products and represents a $60 m i l 1 i o n indus t ry (NMFS 1985).
Blue crabs are caught throughout the year, bu t most are taken du r ing the summer and e a r l y f a l l (Music 1979). Hard crabs (having hardened exoskeletons) are taken p r i m a r i l y i n shallow water i n the warmer months w i t h crab pots o r t r o t l i n e s . The use of t r o t l i n e s f o r b lue crabs has genera l ly diminished i n the New England s ta tes (Sholar 1982) , whereas the use o f crab pots has increased (Tang 1983). Dredges (and less f requent ly, scrapes) are used i n deeper o f fshore waters i n w in te r t o take crabs burrowed i n t o the mud (Adkins 1972; Tang 1983; Schmidt 1985). F ish ing pressure has been s t e a d i l y increasing i n the Chesapeake Bay area; numbers o f fishermen, boats, and pots increased th ree fo ld between 1950 and 1975 (Table 2). Cronin (1983) reported a 7.6-fol d increase between 1948 and 1981 i n the number o t l icenses f o r crabbing issued i n Mary1 and.
Recently molted "soft-she1 1" crabs represent a smal ler po r t i on o f the t o t a l indust ry , a1 though they have a higher value per crab (Haefner and Garten 1974). Methods o f processing used i n the so f t - she l l i ndus t ry were reviewed by Haefner and Garten (1974). Few fishermen engage i n the s o f t s h e l l
business because these crabs must be tended cont inuously (Adkins 1972).
Management o f the commercial b lue crab f i s h e r y i s u s u a l l y l o c a l , and has included measures such as carapace wid th l i m i t s , n e t mesh-size t i m i t s , const ra in ts on gear type, closed sea- sons and areas, p r o h i b i t i o n o f the harvest o f sponge crabs o r o ther fe- males, quotas, and 1 icens ing (Bearden 1978). Even so, an assessment o f the blue crab f i s h e r y i n Chesapeake Bay suggested t h a t b lue crabs are cur- r e n t l y being overf ished (Tang 1983).
The b lue crab a lso supports a rec rea t iona l f i s h e r y and a v a r i e t y o f small-scale commercial harvests and sales by "weekend operators ." Gears used inc lude handlines, pots, and c o l l a p s i b l e t raps. Recreational fishermen genera l ly are l i m i t e d t o a maximum o f f i v e pots (Sholar 1982). Landings from these a c t i v i t i e s have r a r e l y been quant i f ied .
Commercial harvests o f b lue crabs f l u c t u a t e widely. For example, the annual harvest i n Chesapeake Bay f l uc tua ted between 45 and 94 m i l l i o n I b s from 1966 t o 1980, t y p i c a l o f harvests over the l a s t 50 years (Finchham 1981). Rees (1963) and More (1969) found no d i r e c t r e l a t i o n s h i p between commercial catches and recru i tment o f harvestabl e crabs i n subsequent years. Pearson (1948) found t h a t the s i z e o f the spawning stock a lso f a i l e d t o determine the s i z e o f the populat ion t h a t surv ived t o l e g a l s i z e f o r commercial f i sh ing . He noted t h a t f l uc tua t ions i n abundance were r e l a t e d p r i m a r i l y t o ra tes o f s u r v i v a l dur ing the f i r s t year o f l i f e .
Apparently, there are no r e l i a b l e methods f o r p r e d i c t i n g harvests. Pop- u l a t i o n f l u c t u a t i o n s have apparent ly been caused by extreme c o l d weather, reduced s a l i n i t i e s from heavy r a i n s (Pearson 1948), paras i t ism by the leech Myzobdella l u u b r i s (Hutton and S o g a n d a r e s - B e r n a l k t o x i c wastes
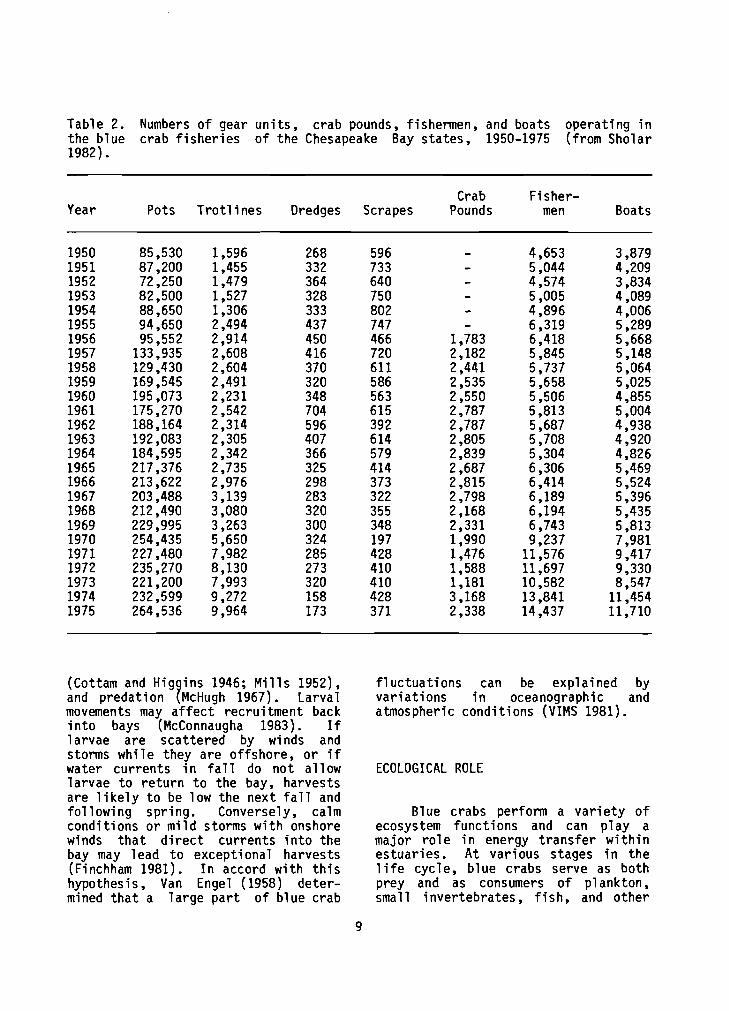
Table 2. Numbers of gear un i t s , crab pounds, fishermen, and boats operating i n the blue crab f i s h e r i e s of the Chesapeake Bay s t a t e s , 1950-1975 (from Sholar 1982).
Crab Fisher- Year Pots Trot1 i nes Dredges Scrapes Pounds men Boats
(Cottam and Hig ins 1946; Mills 1952), and predation qAcHugh 1967). Larval movements may a f f e c t recruitment back in to bays (McConnaugha 1983). I f larvae a r e scat tered by winds and storms while they a r e offshore, o r i f water currents in f a l l do not allow larvae t o re turn t o the bay, harvests a re l i k e l y t o be low the next f a l l and following spring. Conversely, calm conditions o r mild storms w i t h onshore winds t h a t d i r e c t currents i n t o the bay may 1 ead t o exceptional harvests (Finchham 1981). In accord w i t h t h i s hypothesis, Van Engel (1958) deter- mined t h a t a large pa r t of blue crab
f l uctuations can be explained by var ia t ions i n oceanographic and atmospheric conditions (VIMS 1981).
ECOLOGICAL ROLE
Blue crabs perform a va r ie ty of ecosystem functions and can play a major r o l e i n energy t rans fe r w i t h i n e s tua r ies . A t various s tages in the l i f e cycle, blue crabs serve as both prey and as consumers of plankton, small inver tebrates , f i s h , and other

crabs. They are important d e t r i ti- vores and scavengers throughout t h e i r range.
Zoeae are phytopl ankt ivorous (Darnel1 1959), and r e a d i l y consume d i no f l age1 1 ates and copepod naupl ii (Tagatz 1968). The omnivorous megalopa eats f i s h larvae, small she l l f i sh , and aquat ic p lan ts (Van Engel 1958; Darnel1 1959; Tagatz 1968). Cannibalism i s common among a l l l i f e stages o f b lue crabs (Hay 1905; Church i l l 1919; Darnel1 1959; Tagatz 1968) .
Post - la rva l crabs are considered general scavengers, bottom carnivores, d e t r i t i v o r e s , and omnivores (Hay 1905; Darnel1 1959; Adkins 1972). Food h a b i t s tud ies have shown t h a t the predominant foods consumed vary g r e a t l y among 1 ocal i t i e s . Some comon
, items are dead and l i v e f i s h , crabs, organic debr is , shrimp, mollusks ( i n c l u d i n g mussels, clams, oysters, and s n a i l s ) , and aquat ic p lan ts (Newcombe 1945; Darnel 1 1959; W i l l iams 1965; Tagatz 1968; Seed 1980; Arnold 1984; Warren 1985). T r u i t t (1939) found t h a t roo ts , shoots, and leaves o f eel grass (Zos t e r a ) , d i t c h grass (Ruppia), sea l e t t u c e (K), and s a l t marsh grass (S a r t i n a ) were comon ly eaten by crabs + i n sa t marshes, t i d a l creeks, and o ther shal low es tuar ine areas. Darnel 1 (1958) concluded t h a t mollusks were the dominant food of crabs l a r g e r than 120 mm wide. A1 though predator-prey i n t e r a c t i o n s are complex (West and Wi l l iams 1986), these r e l a t i o n s h i p s may be s t a b i l i z e d by predator avoidance mechanisms. For example, the pe r iw ink le ( L i t t o r i n a i r r o r a t a ) reduces i n j u r y and mor ta l i t y ra tes caused by b lue crab by c l imb ing t a l l grass dur ing h igh t i d e s (Warren 1985). Some b i va l ves a l so escape predat ion as they develop t h i c k e r she l ls . Blue crab feeding on in fauna l b iva lves was found t o be a func t i on o f prey avai 1 ab i 1 i t y and she1 1 s t rength of the re1 a t i v e t o predator s t rength r z n d o n and Kennedy 1982).
Blue crabs are the prey o f a v a r i e t y o f animals. Egg masses, c a r r i e d by females, are o f t e n s p e c i f i c a l l y at tacked by some f i s h e s (Adkins 1972). Larva l stages are eaten by f i s h , s h e l l f i s h , j e l l y f i s h , combjel l ies, and var ious o ther plank- t i v o r e s (Van Engel 1958) . Juveni 1es are i d p o r t a n t prey o f many f i s h such as spot ted sea t r o u t (C noscion nebulosus), red drum (* ocel l a t u s ) , black drum cromis), and sheepshead (Archosar us r o b a t o c e halus), as we d *wading b i r d s (Fontenot and R o g i l l i o 1970; Adkins 1972; Barrass and K i t t i n g 1986). Barrass and K i t t i n g (1986) showed t h a t crabs l ess than 10 m long respond t o recorded vocal i z a t i o n s o f 1 aughing g u l l s (Larus a t r i c i l l a ) by f l e e i n g o r h id ing; thus, the absence o f v i sua l cues i n t u r b i d waters apparent ly does no t h inder the detec t ion o f av ian pre- dators by b lue crabs. Juven i le and a d u l t b lue crabs are consumed by mammals, a v a r i e t y o f b i rds , and several f i shes, i n c l u d i n g s t r i p e d bass (Manooch 1973), American eel , An ui11a r o s t r a t a (Wenner and Musick 1 9 h sandbar shark, Carcharhinus p l umbeus (Medved and Marshal 1 1981).
The b lue crab i s hos t t o sevd era1 paras i tes and diseases, b u t many i n f e c t i o n s are temporari 1y e l i m i nated dur ing mol t ing . A f t e r t h e i r l a s t mol t , a d u l t b lue crabs may serve as a 1 odgi ng place f o r barnacles , bryo- zoans, and o the r s e s s i l e organisms ,
(Darnel 1 1959; W i l l iams 1965). The barnacles ~ a l a n u s amphi tri t e and Chelonib ia a t u f a t t a c h t o t h e cara- pace bu t genera h y have 1 i t t l e physio- i o g i c a l e f f e c t on the crab (Darnel1 1959; W i 11 iams 1965)'. a1 thoush the s t a l ked barnacle octa lasmi s 1 o i e i may c l o a a c rab 's s i l t s and s i l l c h a m b e r s ( c a k e y 1961) and saccul i n i d barnacles may prevent m o l t i n g (Steele 1982). I n f e c t i o n s by the protozoan Paramoeba pe rn i c iosa have been responsib le f o r numerous crab morta l i t i e s along the eastern seaboard (Mahood e t a1. 1970).

Blue c r abs have been impl ica ted a s c a r r i e r s of s t r a i n s of t h e bac- ter ium Vibr io cho le rae which a r e respons ib-or outbreaks of human cho le ra (Moody 1982; Welsh and Sizemore 1985; Huq e t a1 . 1986). This Vibr io is apparent ly a na tu ra l p a r t o f t h e e s t u a r i n e ecosystem and i n h a b i t s Chesapeake Bay and o t h e r major f i s h i n g a r eas (Greer 1981) ; however, t h e s e and o t h e r p a r a s i t e s pose no t h r e a t t o humans i f t h e b lue c r abs a r e proper ly s t o r e d , c leaned , and cooked.
ENVIRONMENTAL REQUIREMENTS
Temperature
Water temperature i n f 1 uences growth and su rv iva l o f b lue c rabs . Williams (1965) found t h a t l a r v a l c r abs , r ea red a t temperatures 1 e s s than 21 "C, d i d no t develop beyond t h e f i r s t zoeal s t a g e and d i d not progress p a s t t h e t h i r d zoeal s t a g e when reared a t 30 O C o r higher . Blue c r abs a r e more t o l e r a n t of low temperatures than a r e many spec i e s of f i s h e s and shrimp (Music 1979). Thei r a b i l i t y t o burrow i n t o t h e s u b s t r a t e apparent ly enables them t o be i n s u l a t e d from cold water (Music 1979; Weinstein 1979). The upper i n c i p i e n t l e t h a l temperature f o r j uven i l e b lue c r abs is 33 O C (Holland e t a l . 1971).
L e f f l e r (1972) measured growth of c rabs a t f o u r temperatures , s t a r t i n g wi th 22-nnn c rabs , and found t h e fol lowing mean carapace widths a f t e r 70 days: 56 mn a t 34 O C , 48 mn a t 27 "C, 40 m a t 20 O C , and 38 mm a t 15 "C. The r e l a t i o n s h i p between growth r a t e and water temperature was repor ted by Churchi 11 (1919), Winget e t a l . (1976), and L e f f l e r (1972). Growth r a t e was propor t iona l t o water temperature; growth and molt ing ceased below 15.5 "C (Churchi l l 1919) and below 1 3 O C ( L e f f l e r 1972).
Mortal i t y was determined t o be d i r e c t l y propor t iona l t o temperature
wi th in a range of 15-34 O C ( L e f f l e r 1972). Experiments by Holland e t a1 . (1971) i nd i ca t ed t h a t mortal i t y increased a t temperatures above 30 O C .
L e f f l e r (1972) noted t h a t c r abs a l s o accl imated t o 34 O C were hyperac t ive . A c t i v i t y and aggression of c r abs a l s o decreased with temperature u n t i l a t 1 3 O C almost no movement occurred.
Sal i n i t y
Blue c r abs occupy water ranging from a near-ocean s a l i n i t y of 34 ppt t o f reshwater i n r i v e r s a s f a r a s 195 km upstream from t h e c o a s t (Tagatz 1968; Palmer 1974). Newcombe (1945) wrote t h a t s a l i n i t i e s of 22-28 ppt a r e needed f o r normal ha tch ing of eggs and f o r normal development of zoeae, bu t su rv iva l and growth of megalopae and small j uven i l e c r abs may be normal a t s a l i n i t i e s a s low a s 5 ppt . When s a l i n i t y is very low, l a r v a e may hatch prematurely and d i e i n t h e prezoeal s t a g e (Van Engel 1958). Gunter (1938) noted t h a t pos t - la rva l b lue c r abs move i n t o f reshwater and may do s o through- ou t t h e spec i e s ' range. S p e c i f i c s a l i n i t y l e v e l s a r e not c r i t i c a l f o r pos t l a rva l c r abs (Odum 1953; Costlow 1967; Adkins 1972; Palmer 1974), a l though t h e occurrence of mature ma1 e s general l y decreases with i nc reas ing s a l i n i t y above 10 ppt (Music 1979). Holland e t a1 . (1971) found t h a t s a l i n i t i e s w i th in t h e range of 2-21 ppt had l i t t l e e f f e c t on growth and su rv iva l of j uven i l e s .
Temperature - S a l i n i t y I n t e r a c t i o n s
Optimal temperatures may vary with o t h e r environmental v a r i a b l e s inc luding s a l i n i t y (Winget e t a l . 1976). Costlow (1967) found t h a t su rv iva l of megalopae exceeded 70% a t 20-30 "C when s a l i n i t y was g r e a t e r than 10 ppt , but never exceeded 50% a t 15 O C . Larval development progressed normally a t 25 O C when s a l i n i t y was between 20.1 and 31.1 pp t , bu t d i d no t progress normally beyond t h i s s a l in - i t y range (Nil l iams 1965).

H a b i t a t
The b l u e crab i nhab i t s a l l areas o f es tua r ies t o some ex ten t (Church i l l 1919; Newcombe 1945; Palmer 1974; Music 1979). Weinste in (1979) found t h a t shal low s a l t marsh h a b i t a t s were important nurser ies f o r j uven i l es . Mature ma1 es p r e f e r creeks, r i v e r s , and upper es tuar ies , b u t t h i s may be a response t o s a l i n i t y r a t h e r than t o o ther phys ica l o r b i o l o g i c a l fea tures o f t h e h a b i t a t such as re fuge (Church i l l 1919; W i l 1 iams 1965; Music 1979). When n o t mating, mature females tend t o remain i n h igh s a l i n - i t y areas o f lower es tuar ies and surrounding waters (Church i l l 1919; Van Engel 1958; Palmer 1974; Music 1979).
The opt imal h a b i t a t f o r small crabs i s shal low es tuar ine water w i t h bottoms o f s o f t d e t r i t u s , mud, o r mud-she1 1 (Adkins 1972). Larger crabs p re fe r red deeper es tuar ine waters having harder bottom substrates.
Other Environmental Factors
Among the many st resses t h a t a f f e c t b lue crabs w h i l e they occupy
nursery areas a re pes t ic ides , domestic and i n d u s t r i a l wastes, a1 t e r a t i o n o f cur ren ts , and des t ruc t i on o f marsh- lands (Adkins 1972). Blue crabs are adversely a f f e c t e d by a wide range o f tox icants , i n c l u d i n g naphthalene (Pearson 1979; Sabourin 1982), d imethyl naphthalene (Mantel e t a1 . 1985), methoxychlor (Bookhout and Costlow 1976) , DOT (Commercial F i she r ies Review 1946; Mahood e t a l . 1970), benzene ( S a i f f and C r i s t i n i 1982; Mantel e t a l . 1985), Kepone (F isher e t a l . 1983), t he organophos- phate f e n i t r o t h i o n (Johnston and Corbet t 1985), cadmium (Brouwer e t a l . 1984), and a c i d r u n o f f ( L i v i n ston e t a,. 1976; Laugh l in e t a1 . 19787.
E f f e c t s from these tox i can ts range from sub1 e tha l responses, such as decreases i n i o n exchange e f f i c i e n - cy a t t he g i l l s (Sabourin 1982) and decreased growth (Mantel e t a l . 1985), t o d i r e c t mor ta l i t y (Commercial F i she r ies Review 1946). Seve r i t y o f the e f f e c t s depends on the tox i can t , concentrat ion, t ime exposed, s a l i n i t y , t i d a l cyc le , age and m o l t phase o f crab, and o the r va r i ab les . Many o f t he tox i can ts a re bioaccumulated i n b l u e crabs and passed t o humans and o the r na tu ra l predators.

LITERATURE CITED
Adkins, G. 1972. A s tudy o f the b lue crab f i s h e r y i n Louisiana. La. W i ld l . Fish. Corn., Oyster, Water Bottoms and Seafood Div. Tech. B u l l . No. 3. Baton Rouge, La. 57 pp.
Arnold, W . . 1984. The e f fec ts o f prey s ize, predator s ize, and sedi- ment composition on the r a t e o f predat ion o f the b lue crab, C a l l i nectes sapidus Rathbun, on the hard c K Mercenari a mercenaria (Linne). J. Exp. B i o l . Ecol. 80 ~207-219.
Barrass, A.N., and C.L. K i t t i n g . 1986. Responses of Ca l l i nec tes sa idus bluecrabs t o recorded group 7%- ee i n g voca l iza t ions o f Sterna terns and Larus a r t r i c i l l a - & i T s . Annual. Meeting o f the American Soc ie ty o f Zool ogi s t s , American Microscopical Society, Animal Behavior Society, Crustacean Society, I n t e r n a t i o n a l Associat ion o f Astacology, and the Soc ie ty o f Systematic Zoology. L o u i s v i l l e , Ky, USA, Dec. 27-30, 1982. Am. Zool . 22(4) :853.
Bearden, C.M. 1978. Blue crab laws and regu la t i ons o f the South A t l a n t i c and Gu l f s tates. Pages 53-70 - i n R.J. Rhodes and W.A. Van Engel, eds. Report o f t he Nat ional Blue Crab Indus t r y Morkshop. Spec. Rep. Appl ied Mar. Sci . Ocean Eng. 185, Va. I n s t . Mar. Sc i . and Spec. Rep. S.C. Mar. Resour. Cent.
Blundon, J .A., and V.S. Kennedy. 1982. Mechanical and behavioral aspects o f b lue crab, C a l l inectes sapidus (Rathbun) , predat ion on
Chesapeake Bay b iva lves . J. Exp. Mar. B i o l . Ecol . 65:47-65.
Bookhout, C.G., and J.D. Costlow, J r . 1976. Ef fects o f mirex, methoxy- ch lo r , and ma1 a th ion on development o f crabs. U.S. Environ. Prot . Agency Environ. Res. Cent. Ecol . Res. Ser. EPA 600/3-76-007. 86 pp.
Brouwer, M., T. Brouwer-Hoexum, and D. M. Engel. 1984. Cadmium accumu- l a t i o n by the b lue crab, Ca l l inec tes sapidus : involvement o f hemocyani n and cha rac te r i za t i on o f cadmium- b ind ing pro te ins . Mar. Environ. Res. 14:71-88.
Cargo, D.G. 1958. The m ig ra t i on o f a d u l t female b lue crabs, Ca l l inec tes sapidus Rathbun, i n Chincoteaque Bay and adjacent waters. J. Mar. Res. 16 : 180-291.
Causey, D. 1961. The barnacle genus Octolasmis i n the Gu l f o f Mexico. Turtox News 39(2) : 51-55.
Church i l l , E.P., Jr. 1919. L i f e h i s - t o r y o f the b lue crab. B u l l . Bur. Fish. 36:95-128, co lored f r o n t i s - piece.
Church i l l , E.P., J r . 1942. The zoeal stages o f the b lue crab, Ca l l i nec tes sapidus Rathbun. Publ. No. 49, Chesapeake B i o l . Lab., Sol omons, MD 26 PP.
Cornercia1 F isher ies Review. 1946. DDT. Commercial F isher ies Review 8(6) :30-32.

Cook, D. 1981. I n search o f t he b lue crab. Mar. Resour. B u l l . 13(1) : 2-3.
Costlow, J.D., J r . 1967. The e f f e c t o f s a l i n i t y and temperature on sur - v i v a l and metamorphosis o f mega1 ops o f t he b l u e crab, C a l l i nec tes sapidus. He1 go1 . Wiss . Meeresunters. 15~84-97 .
Costlow, J.D., Jr., and C. G. Book- hout. 1959. The 1 a r v a l development o f C a l l i n e c t e s sapidus Rathbun reared i n t he labora to ry . B i o l . Bu l l . (Woods Hole) 116(3):373-396. 373-396.
Cottam, C., and E. Higgins. 1946. DDT: i t s e f f e c t s on f i s h and w i l d l i f e . U.S. F i s h W i l d l . Serv. C i r c . No. 11. 14 pp.
Couch, J.N. 1942. A newfungus on c rab eggs. J . E l i sha M i t c h e l l Sc i . SOC. 58(2) : 158-162.
Cronin, L.E. 1949. Comparison o f methods o f tagg ing t h e b l u e crab. E C O ~ ogy 30(3) : 390-394.
Cronin, L.E. 1983. Ana lys is o f l o c a l popu la t ions o f b l ue crab. Pages 112-127 i n P. Jones, D. Heimbuch and C. S taggTeds . Report o f t he work- shop on b l u e c rab stock dynamics i n Chesapeake Bay. Chesapeake B i o l . Lab. UMCEES 83-303 CBL, Solomons. 168 pp.
Da rne l l , R.M. 1958. Food h a b i t s o f f i s h e s and 1 a rger i nve r teb ra tes o f Lake Pontchar t ra in , Louis iana, an es tua r i ne community. Pub1 . I n s t . Mar. Sc i . Univ. Tex. 5:353-416.
Darne l l , R.M. 1959. Studies o f t h e l i f e h i s t o r y o f t h e b lue c rab (Ca l l inec tes- sapidus Rathbun) i n Lou is iana waters. Trans. Am. F ish.
Daugherty, F.M., J r . 1952. The b lue c rab i n v e s t i g a t i o n , 1949-1950. Tex. J. Sc i . 4:77-84.
Dudley, D.L., and M. H. Judy. 1971. Occurrence o f 1 a r va l , juven i 1 e, and mature crabs i n t he v i c i n i t y o f Beaufor t I n l e t , No r th Carol ina. U.S. N a t l . Mar. F ish. Serv. Spec. Sc i . Rep. F ish. 637. 10 pp.
F i e d l e r , R.H. 1930. So l v i ng t he ques t ion o f c rab m ig ra t i ons . F ish. Gazette 47(6) : 18-21.
Finchham, M.W. 1981. The o f f s h o r e odyssey o f t h e A t l a n t i c B lue Crab. MD. Sea Grant Prog. Newsl . 4(4) :2-5.
F i s c h l e r , K.J., and C.H. Walburg. 1962. Blue c rab movement i n coas ta l South Carol ina, 1958-59. Trans. Am. F ish. Soc. 91:275-278.
F isher , D.J., M.E. Bender, and M.H. Roberts, J r . 1983. E f f e c t s o f i n g e s t i o n o f Kepone-contaminated food by j u v e n i l e b l u e crabs ( c a l l i nec tes - sa i dus Rathbun) . Aquat. Toxic01 .&234.
Fontenot, B.J., and H.E. R o g i l l i o . 1970. A s tudy o f e s t u a r i n e spo r t - f i s h e s i n t he B i l o x i marsh complex. Compl . Rep., D i n g e l l -Johnson P r o j e c t F-8, La. W i l d l . Fish. Comm., Baton Rouge. 172 pp.
Gray, E.H., and C.L. Newcombe. 1939. S tud ies o f m o l t i n g i n C a l l i n e c t e s m. Rathbun. Growth 2 (4 ) :
Greer, J . 1981. The cho le ra connec- t i o n . Md. Sea Grant Prog. Newsl . 4(4) :101-112.
Gunter, G. 1938. The common b l u e c rab i n f r e s h waters. Science 87 : 87-88.
Haefner, P.A., J r . 1961. A b lue b lue crab. Es tua r i ne B u l l . 6(3-4) :3-5.
Haefner, P.A., Jr., and D. Garten. 1974. Methods o f hand l ing and shedding b l u e crabs, C a l l i nec tes sapidus. Va. I n s t . Mar. Sc i . Resour. Advis. 8, and Spec. Rep.

App l ied Mar. Sc i . Ocean. Eng. 52. 14 PP.
Hav. W.P. 1905. The l i f e h i s t o r v o f u ,
t h e b l u e c rab (Ca l l i nec tes sa i d i s ) . Rep. U.S. Bur. F ish. 1 9 0 4 : 3 h
Hol land, J.S., D.V. A l d r i c h , and K. Strawn. 1971. E f f e c t s o f tempera- t u r e and sa l i n i t y on growth, food convers ion, s u r v i v a l , and tempera- t u r e r e s i s t a n c e o f j u v e n i l e b l u e crabs, C a l l i nec tes sapidus Rathbun. Tex. A&M Univ. Sea Grant Publ. TAMU-SG-71-222, C o l l ege S ta t i on . 166 pp.
Hopkins, S.H. 1943. The ex te rna l morphology o f t h e f i r s t and second zoeal stages o f t h e b l u e crab, C a l l i nec tes sapidus Rathbun. Trans. Am. Microsc. Soc. 62:85-90.
Hookins. S.H. 1947. The nemertean ~a rc i nonemer tes as an i n d i c a t o r o f the spawning h i s t o r y o f t h e host , C a l l i n e c t e s sapidus. J. P a r a s i t o l . 33(2):146-150.
Humes, A.G. 1942. The morphology, taxonomv. and bionomics o f t he nemertean genus Carcinonemertes. Univ. 111. B i o l . Monoar. 18(41:
Huq, A,, S.A. Huq, D.J. Grimes, M. O'Brien, K.H. Chu, J.M. Capuzzo, and R.R. Co lwe l l . 1986. Co lon i za t i on o f t he q u t o f t he b lue c rab ( ~ a l l i n e c t e s sa i d u s ) by ( V i b r i o
+nviron. M ic ro - cho l erae) . App blol.: 586-588.
Hutton, R. F., and F. Sogandares- Bernal . 1959. Notes on t he d i s t r i - b u t i o n o f t he leech, Myzobdel la l u g u b r i s Leidy, and i t s assoc ia t i on w i t h mor ta l i t y o f t he b lue crab, Ca l l i nec tes sapidus Rathbun. J. Paras i t o 1 . 45:384,404,430.
Johnston, J.J., and M.D. Corbet t . 1985. The e f f e c t s o f temperature, s a l i n i t y and a s imu la ted t i d a l c y c l e on t he t o x i c i t y o f f e n i t r o t h i o n t o
C a l l i n e c t e s sa idus. Comp. Biochem. ~ h y s i o l . ~ o m p +armacol. Toxic01 . 80C 145-150.
Jones, P., D. Heimbuch, and C. Stagg. 1983. Report o f t he workshop on b l u e crab s tock dynamics ' i n Chesapeake Bay. Univ . o f Maryland Chesapeake B i o l . Lab. UMCEES 83-303 CBL , Sol omons . 168pp.
Judy, M.H., and D.L. Dudley. 1970. Movements o f tagged b lue crabs i n Nor th Ca ro l i na waters. Comer. F ish . Rev. 32(11) :29-35.
Laughl in , R.A., C.R. Cr ipe, and R.A. L i v i ngs ton . 1978. F ie1 d and 1 abora to rv avoidance reac t i ons b.y b l ue crabs ( C a l l i n e c t e s sa i dus ) tb storm water r u n o f f . dk- Am. F ish . Soc. 107:78-86.
L e f f l e r , C.W. 1972. Some e f f e c t s o f temperature on t he growth and meta- bo l i c r a t e o f j u v e n i l e b l ue crabs, C a l l i nec tes sa idus, i n t h e labora- t o r y . Mar. *(N.Y .) 14:104-110.
L i v i ngs ton , R.J. 1976. D iu rna l and seasonal f l u c t u a t i o n s o f organisms i n a n o r t h F l o r i d a es tuary . Estua- r i n e Coastal Mar. Sc i . 4:373-400.
L i v i ngs ton , R.J., C.R. Cr ipe, R.A. Laugh l in , and F.G. Lewis 111. 1976. Avoidance responses o f es tua r i ne organisms t o storm water r u n o f f and pul p m i l 1 e f f l u e n t s . Es tua r i ne Processes 1:313-331.
Mahood, R.J., M.D. McKenzie, D.P. Middlaugh, S.J. B o l l a r , J.R. Davis, and D. Spitzbergen. 1970. A r e p o r t on t h e coopera t i ve b l u e crab s tudy - South A t l a n t i c States. F la. Dep. Nat. Resour. Cont r ib . Ser. No. 139. 32 PP.
Manooch, C.S., 111. 1973. Food h a b i t s o f v e a r l i n q and a d u l t s t r i o e d bass, ~ o k n e s a x a t i l i s ( ~ a l baum) , from A1 bemarle Sound. Nor th Caro- l i n a . Chesapeake Sc i . 14:73-86.

Mantel, L.H., E. Flynn, M. Katz, and L. Knapp. 1985. E f f e c t s o f ben- zene and d imethy l naphthalene on homeostat ic processes i n two species o f crabs. Mar. Envi ron. Res. 258-261.
Mary1 and Tidewater News. 1950. T r u l y a b l ue crab. Md. T idewater News' 7(4):5.
McConaugha, J.R. 1983. Blue c rab l a r v a l r ec ru i tmen t . Pages 128-152 i n P. Jones, D. Heimbuch and C. - Stagg, eds. Report o f t h e workshop on b lue c rab s tock dynamics i n Chesapeake Bay. Univ. o f Maryland Chesapeake B i o l . Lab. UMCEES 83-303 CBL, Solomons. 168 pp.
McConaugha, J.R., C.M. Jones, and R. Mar is . 1986. Assessment o f C. sa idus spawning s tock s i z e repro- + u c t i v i t y p e r i o d i c i t y and f e c u n d i t y i n Chesapeake Bay. Am. Zoo1 . 26(4) : 129A (abs t r . ) .
McHugh, J.L. 1967. Es tua r i ne nekton. Pages 581-620 i n G.H. Lau f f , ed. Es tuar ies . Am. Assoc. Adv. Sc i . Pub1 . No. 83.
Medved, R.J., and J.A. Marsha l l . 1981. Feeding behav io r and b i o l o g y o f younq sandbar sharks, ca rcha rh in i s p l umbeus ( ~ i s c e s , Carcharh in i dael i n Chincoteaaue Bav. V i r g i n i a . U.S: N a t l . Mar. ' ~ i s l h : Serv. F ish. B u l l . 79(3) :441-447.
M i l l i k i n , M.R., and A.B. Wi l l iams. 1984. Synopsis o f b i o l o g i c a l da ta on t h e b lue crab, C a l l i nectes sapidus. NOAA Tech. Rep. MFS 1. ?-A0 !-I sher ies Synopsi s 138.
M i l l s , H.R. 1952. Deaths i n F l o r i d a marshes. Audubon Mag. 54:285-291.
Moody, M.W. 1982. Zoonot ic diseases. Pages 65-69 - i n H.M. Pe r r y and W. A. Van Engel, eds. Proceedings o f t h e b lue c rab col loquium, 16-19 October, B i l o x i , M i s s i s s i p p i . G u l f S ta tes
Mar. F ish. Comm. Tech. Rep. 7, Ocean Springs, Miss. 235 pp.
More, W.R. 1969. A c o n t r i b u t i o n t o t he b i o l o g y of t he b l u e c rab ( C a l l i nec tes - sapidus Rathbun) i n Texas, w i t h a d e s c r i ~ t i o n o f t h e f i she ry . Tex. Parks ' W i l d l . Dep. Tech. Ser. No. 1. 31 pp.
Music, J.L. 1979. Assessment o f Georgia 's shrimp and c rab resources. Ga. Dep. Nat. Resour. Coastal Resour. Div., Con t r i b . Ser. No. 30. 75 PP.
Na t i ona l Mar ine F i she r i es Serv ice (NMFS). 1981. F i s h e r i e s o f t h e Un i t ed States, 1980. U.S. Na t l . Mar. Fish. Serv. Curr. F ish. S ta t . No. 8100. 132 pp.
Nat iona l Mar ine F i she r i es Serv ice (NMFS). 1982. She1 l f i s h Market Review. U.S. Dep. Corn., Washing- ton, D.C. 47 pp.
Nat iona l Mar ine F i she r i es Serv ice (NMFS) . 1985. Processed f i s h e r y products, 1985. U.S. N a t l . Mar. F ish. ~ e r v . Curr. F ish. S ta t . No. 8357. 37 pp.
Nat iona l Marine F i s h e r i e s Serv ice (NMFS). 1986. F i s h e r i e s o f t he Un i t ed States, 1985. U.S. N a t l . Mar. F ish. Serv. Curr. F ish. S t a t . No. 8368. 122 pp.
Newcombe, C.L. 1945. The b i o l o g y and conserva t ion o f t h e b lue crab, C a l l i nec tes sapidus Rathbun. Va. F ish. Lab. Ed. Ser. No. 4, Glou- c e s t e r Po in t . 39 pp.
Newcombe, C.L. 1948. An appl i c a t i o n o f t h e a l l ome t r y equat ion t o t h e s tudy o f i n C a l l i nec tes sapidus Rathbun. Am. Nat. 82:315-325.
Odum, H. F. 1953. Fac to rs c o n t r o l 1 i ng mar ine i nvas ion i n t o F l o r i d a f r esh - waters. B u l l . Mar. Sc i . G u l f Ca r i bb. 3(2) : 134-156.

Palmer, b l ue Georg
R.A. 1974. Studies on the crab (Cal l inectes sapidus) i n
i a . Ga. Dep. Nat. Resour., ~ame- F ish Div., Contr ib. Ser. No. 29. 59 pp.
Pearson, J .C. 1948. F luc tuat ions i n the abundance o f the blue crab i n Chesapeake Bay. U.S. F ish Wi ld l . Serv. Res. Rep. No. 14. 26 pp.
Pearson, W.H. 1979. Detect ion o f n a ~ t h a l e n e bv the b lue crab. ~ a i 1 inectes ;apidus. Estuaries 2 : 64-65.
Piers, H. 1923. The b lue crab (Cal l i nectes sapidus, Rathbun) : Extension o f i t s range northward t o near Ha l i fax , Nova Scotia. Proc. Nova Scot ia I n s t . Sci. 15:83-90.
Porter , H.J. 1956. Delaware b lue crab. Estuar ine Bu l l . 2(2) :3-5.
Pyle, R.W., and L.E. Cronin. 1950. The aeneral anatomv o f the b lue crab c a l l inectes sa idus Rathbun. Chesapeake Biol* Pub1 . No. 87, Solomons, Md. 40 pp.
Rees, G.H. 1963. Progress on blue crab research i n the South A t l a n t i c . Proc. Gu l f Caribb. Fish. I n s t . Annu. Sess. 15:llO-115.
Roaers-Tal ber t . R. 1948. The funaus La enidium c a l l inectes Couch (1942) * the blue crab i n Chesa- -.,a- -
peake Bay. B i o l . B u l l . (Woods Hole) 95(2) :214-228.
Sabourin, T.D. 1982. Respiratory and c i r c u l a t o r y responses of the b lue crab t o naphthalene and the e f fec t o f accl imat ion sal i n i t y . Aquat. Toxic01 . 2:301-318.
S a i f f , E., and A.C. C r i s t i n i . 1982. E f fec ts of benzene on the c e l l u l a r s t ruc tu re o f the mid gu t gland o f j u v e n i l e Ca l l inec tes sapidus. Am. Zoo1 . 22(4) :907.
Sandoz, M. , and R. Rogers. 1944. The ef fect of environmental f ac to rs on hatchinq, mol t i nq , and su rv i va l of zoea ia rvae o f - - t h e blue crab Ca l l inectes Rathbun. ECOI ogy 25 : 216-
Scattergood, L .W. 1960. B l ue crabs (Cal l inectes sa idus) i n Maine. Maine F i e l d N a t h : 59-63.
Schmidt, S. 1985. Dredging f o r b lue crabs: V i r g i n i a ' s w in te r f i she ry . Mar. Resour. B u l l . 17(1) :2-3.
Seed, R. 1980. Predator-prey re1 a- t ionsh ips between the mud crab Pano eus h e r b s t i i , the blue crab & t e T and the A t l a n t i c r ibbed musse Geukensia (=Modiol us) demissa. Estuar ine C o a s t a l . Sci.: 445-458.
Sholar, T.M. 1982. Blue crab f i s h - e r ies of the A t l a n t i c coast. Pages 111-127 i n H.M. Perry and W.A. Van Engel, e z . Proceedings o f the b lue crab colloquium, 16-19 October, B i l o x i , M iss i ss ipp i , Gu l f States Mar. Fish. Corn. Tech. Rep. 7, Ocean Springs, Miss. 235 pp.
Steele. P. 1982. A s v n o ~ s i s o f the b io logy of the b l ue Gab ' C a l l i nectes sa idus Rathbun i n F lor ida. Pages d35- i n H.M. Perry and W.A. Van Engel ,eds. proceedings o f the b lue crab co l loauium. 16-19 October. B i l o x i , ~ i s s i s s i ~ p i . Gu l f s ta tes Mar. Fish. Corn. Tech. Rep. 7, Ocean Springs, Miss. 235 pp.
Subrahmanyam, C.B., and C.L. Coultas. 1980. Studies on the animal com- muni t ies i n two no r th F l o r i d a s a l t marshes. P a r t 3. Seasonal f l uctua- t i o n s of f i s h and macroinverte- brates. B u l l . Mar. Sci. 30:790-818.
Sulkin, S.D., C.E. Epi fanio, and A.J. Provenzano, J r . 1982. The b lue crab i n the mid-At lant ic b i g h t es tuar ies : a proposed recru i tment model. MD. Sea Grant Program Tech.

Rep. No. UM-SG-TS-82-04. U n i v e r s i t y o f Maryland. 36 pp.
Su l l i van , W.E. 1909. Notes on t he crabs found i n Narraganset t Bay. Comnissioners o f I n l a n d F i she r i es o f Rhode Is land . 39 th Annu. Rep:56-78.
Tagatz, M.E. 1968. B io l oqy o f t h e -- b l u e crab, C a l l i nec tes sapidus Rathbun, i n t h e S t . Johns River . F l o r i da . U.S. F i s h W i l d l . Serv. F ish. B u l l . 67:17-33.
Tagatz, M.E., and A.B. H a l l . 1971. Annotated b i b l i o q r a p h y on t h e f i s h - i n g i n d u s t r y and b i b l o g y of t he b l u e crab, C a l l i nec tes sapidus. U.S. Nat. Mar. F ish . Serv. Spec. Sc i . Rep. F ish . 640. 94 pp.
Tang, Q. 1983. Assessment o f t he b lue c rab commercial f i s h e r y i n Chesapeake Bay, 1983. Pages 86-110 i n P. Jones, D. Heimbuch and C. - Stagg, eds. Report o f t h e workshop on b l u e c rab s tock dynamics i n Chesapeake Bay. Univ. o f Maryland Chesapeake B i o l . Lab. UMCEES 83-303 CBL, Solomons. 168 pp.
Thompson, B.G. 1984. F i she r i es o f t h e Un i t ed States, 1984. (1.5,. N a t l . F ish. Serv. Curr . F i sh . S ta t . No. 7500.
T r u i t t , R . V . 1937. Crab tagg ing opera t ions . Chesapeake R i o l . Lab., Mimeo. 4 pp.
T r u i t t , R . V . 1939. Our water resources and t h e i r conserva t ion . Chesapeake B i o l . Lab. Cont r i b. No. 27, Solomons, Md. 103 pp.
Van Engel, W .A. 1958. The b lue c rab and i t s f i s h e r y i n Chesapeake Bay. P a r t 1. Reproduct ion, e a r l y devel - opment, growth, and m ig ra t i on . Commer. F ish . Rev. 20(6) :6-17.
VA. I n s t . Mar. Sc i . Sea Grant Program. (vIMS). 1981. P r e d i c t i n g V i r g i n i a ' s
b lue crab catch. Commer. F ish ing News1 . l ( 3 ) :1-2.
Warren, J.H. 1985. C l imb ing as an avoidance behavior i n t he s a l t marsh pe r iw ink le , L i t t o r i n a i r r o r a t a (Say). J. Exp. Mar. B i o l x 89: 11-28.
Weinstein, M.P. 1979. Shal low marsh h a b i t a t s as p r imary nu rse r i es f o r f i s h and s h e l l f i s h , Cape Fear R iver , Nor th Caro l ina . U.S. N a t l . Mar. F ish. Serv. F ish. B u l l . 77~339-357.
Welsh, P.C., and R.K. Sizemore. 1985. Inc idence o f bacteremia i n s t r essed and unstressed ~ o ~ u l a t i o n s o f t h e b lue crab, ~ a i l i n e c t e s sa idus. ADD^ . Env i ron . Mic ro -%T l o .
U e n n ~ r , C.A., and J.A. Musick. 1975. Food h a b i t s and seasonal abundance o f t h e American ee l , A n g u i l l a r o s t r a t a , f rom t h e lower Chesapeake Bay. Chesapeake Sci . 16: 62-66.
West, D.L., and A.H. U i l l i a m s . 1986. Predat ion by C a l l i nec tes sa idus (Rathbun) w i t h i n S a r t i n a a& f l o r a ( L o i s e l ) ma* m. K B i o l . Ecol . 100:75-95.
Wi l l i ams, A.B. 1965. Mar ine decapod crustaceans o f t he Caro l inas . U.S. F i s h W i l d l . Serv. F ish. B u l l . 65 : 1-298.
W i l l iams, A.B. 1984. Shrimps, l ob - s te r s , and crabs o f t he A t l a n t i c coas t o f t h e eas te rn Un i t ed States, Maine t o F lo r i da . S m i thsonian -.
I n s t i t u t i o n Press, Washington, D.C. 550 pp.
Winget, R.R., C.E. Ep i f an io , T. Run- ne ls , and P . Aus t in . 1976. E f f e c t s o f d i e t and temperature on growth and m o r t a l i t y o f t he b lue crab, C a l l i n e c t e s sa idus, mainta ined i n a r e c i r c u l a t i n g -f cu t u r e system. Proc. N a t l . She1 1 f i s h . Assoc. 66:29-33.

l O t 7 Y - 1 0 1
R-RT DOCUMENTATION 1. "Emrn NO. 2 PAGE IB~ 01 og i cal Report 82( 11.100) *I
L tnr m d a ~ w n ~ e Species P r o f i l e s : L i f e H i s t o r i es and Environmental Requirements o f Coastal Fishes and Inver tebrates (Mid-At lant ic ) - -B l ue Crab.
I IS. Sumemantaty Not-
*U.S. Army Corps o f Engineers Report No. TR EL-82-4 10. Abstnct (Urnik 200 word.)
Species p r o f i l e s are sumnaries o f the l i t e r a t u r e on taxonomy, l i f e h i s t o r y , and environmental requirements o f coastal f i shes and aquat ic inver tebra tes . They are prepared t o a s s i s t w i t h impact assessment. The b lue crab, Ca l l i nec tes sapidus, occurs i n lower reaches o f f reshwater r i v e r s , estuar ies, and coastal waters along the A t l a n t i c seaboard and G u l f o f Mexico, and the species supports the l a r g e s t crab f i s h e r y i n the Uni ted States Chesapeake Bay provides the g rea tes t product ion o f b lue crabs on the eas t coast. The b lue crab 's h igh abundance i n estuar ies, d iverse feeding habi ts , and importance as prey f o r o t he r marine animals i n d i c a t e i t s impor tant r o l e i n the s t r uc tu re and func t ion of es tuar ine communities. Female b lue crabs spawn i n high-sal i n i t y lower es tuar ies of coasta l areas; the r e s u l t i n g la rvae are p lank ton ic and develop i n t o j u ven i l es a t 5 t o 10 weeks o f age. Juveni les g radua l l y migrate i n t o shal lower, less-sa l i n e upper es tuar ies and r i v e r s where they grow and mature a t 1-2 y r o f age. Mating occurs i n the upper es tuar ies a f t e r which females migrate t o areas having h igher s a l i n i t i e s . Growth and su r v i va l of b lue crabs are s t r ong l y a f f ec ted by water temperature and s a l i n i t y , b u t tolerances vary w i t h l i f e stage. Larvae requ i re temperatures o f 20-30 OC and s a l i n i t i e s o f 10-30 pp t f o r proper development, bu t s a l i n i t y and tempe~atu re tolerances are broad f o r advanced juven i les and adu l t s . Blue crabs use near l y a l l areas w i t h i n es tuar ies as nursery h a b i t a t and crab populat ions are sens i t i ve t o changes i n phys ica l features o f contamination of these areas. 17. Dorumnt Amlysls a. k r l p t o n
Estuar ies Feeding She1 l f i s h Spawning Crustacea Growth
b. Idmtl8anlOpn.lnd.d Tarms
Blue crab Temperature requi remen t s Ca l l inectes sapi dus Hab i ta t requirements
I nuiglenrs kcn.lon NO.
L "ma 0.1.
mrch 1989 6,
7. ~uthods)
& n n i f ~ r H i l l , [lebn I F n w l ~ r , andMiCh i lp1 J - Van Den Avvle i. hrlormlnl O~..nlxatlon Name and M d m s
Georg i a Cooperative F ish and W i l d l i f e Research U n i t School o f Forest Resources Un i ve r s i t y o f Georgia
GA 30602 12 Spon.orlnl O~..nlmlon Name and Addms
U.S. Department o f the I n t e r i o r U.S. Army Corps o f Engineers F ish and W i l d l i f e Service Waterways Experiment S ta t i on Nat ional Wetlands Research Center P.O. Box 631 Washington, DC 20240 Vicksburg, MS 39180
a hfi-lna Omnlxatlm map(. NO.
10. Cloi.c(ffaskmorR UnH No.
11. con(m~t(c) w cren((o) WO.
(c)
(C)
11 TYD. of Awa 4 hrlod wmd
11
1 Unl imi ted b ~ n c l a s s j f i ~ d 18 1
c. COSATl fleldlCmuo
la Awallabllity Statement I 19. k u f i t y Class CThi. R e m ) 21. No. of P u n
20. k u r l t y class ( T ~ I S P.-) I Unc lass i f i ed
I a mu I
LI.. ANSU39.18l O ~ O N A L row m (&m

As the Nation's principal conservation agency, the Department of the Interior has responsibility for most of our nationally owned public lands and natural resources. This includes fostering the wisest use of our land and water resources, protecting our fish and wildlife, preserving the environmental and cultural values of our national parks and historical places, and providing for the enjoy- ment of life through outdoor recreation. The Department assesses our energy and mineral resources and works to assure that their development is in the best interests of all our people. The Depart- ment also has a major responsibility for American Indian reservation communities and for people who live in island territories under U.S. administration.
U.S. DEPARTMENT OF THE INTERIOR FlSH AND WILDLIFE SERVICE
TAKE PRIDE in America
UNITED STATES DEPARTMENT OF THE INTERIOR
FlSH AND WILDLIFE SERVICE National Wetlands Research Center
NASA-Slidell Computer Complex 101 0 Gause Boulevard
Slidell. LA 70458
POSTAOE AND FEE8 PAID U 8 DEPARTMENT OF THE INTERIOR
lNl423
OFFICIAL BUSINESS PENALTY FOR PRIVATE USE, $300





























