Embed Size (px)
Citation preview

BioSci 145B lecture 9 page 1 ©copyright Bruce Blumberg 2004. All rights reserved
BioSci 145B Lecture #9 6/1/2004
• Bruce Blumberg– 2113E McGaugh Hall - office hours Wed 12-1 PM (or by appointment)– phone 824-8573– [email protected]
• TA – Curtis Daly [email protected]– 2113 McGaugh Hall, 924-6873, 3116– Office hours Tuesday 11-12
• lectures will be posted on web pages after lecture – http://eee.uci.edu/04s/05705/ - link only here– http://blumberg-serv.bio.uci.edu/bio145b-sp2004– http://blumberg.bio.uci.edu/bio145b-sp2004
• DON’T FORGET –TERM PAPERS ARE DUE BY 5 PM on FRIDAY JUNE 4

BioSci 145B lecture 9 page 2 ©copyright Bruce Blumberg 2004. All rights reserved
Requirements for the term paper
• Goals– Analytical thinking– Improved writing
• Select a topic related to genomic or proteomic analysis of an interesting problem– Talk with me about your topic
• Write a short paper (~5 pages) in the style of a research grant describing how you will attack this problem (example is posted).– Specific aims – questions, hypotheses– Background and significance
• What is known, what remains to be learned• why should someone give you money to study this problem?
– Research plan – specific experiments to answer the questions posed in specific aims
• Expected vs unexpected results
BioSci 145B lecture 9 page 3 ©copyright Bruce Blumberg 2004. All rights reserved
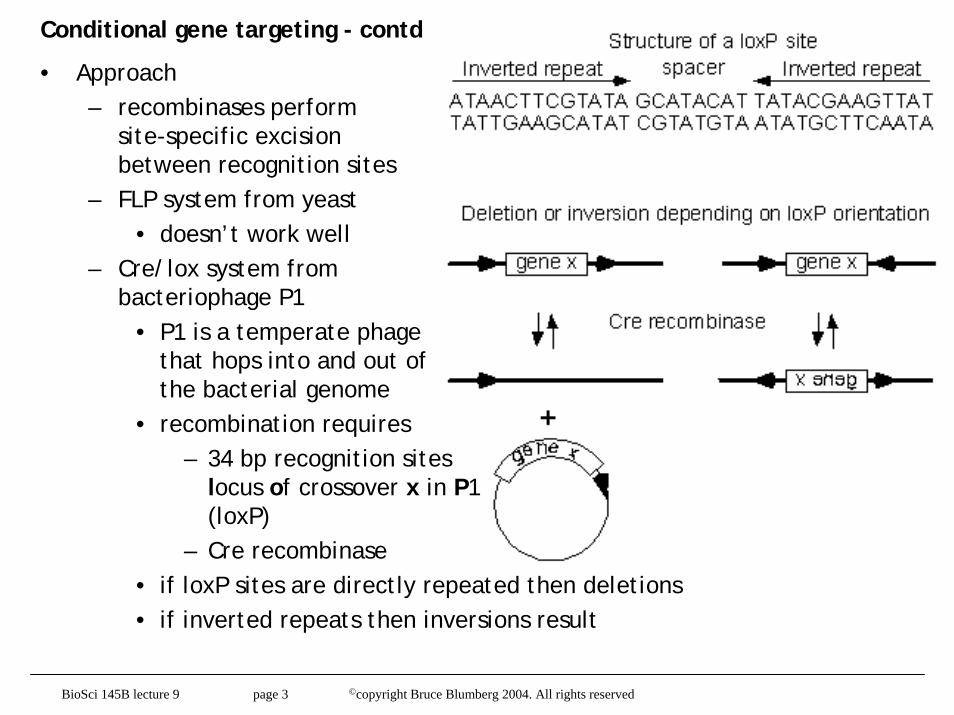
Conditional gene targeting - contd
• Approach– recombinases perform
site-specific excision between recognition sites
– FLP system from yeast• doesn’t work well
– Cre/lox system from bacteriophage P1
• P1 is a temperate phage that hops into and out of the bacterial genome
• recombination requires – 34 bp recognition sites
locus of crossover x in P1(loxP)
– Cre recombinase• if loxP sites are directly repeated then deletions• if inverted repeats then inversions result

BioSci 145B lecture 9 page 4 ©copyright Bruce Blumberg 2004. All rights reserved
Conditional gene targeting (contd)• Strategy
– Make targeting construct (minimum needed for grant)
– homologous recombination,– transfect CRE, select
for loss of tk– Southern to select
correct event• Result called
“floxed allele”– inject into blastocysts,
select chimeras– establish lines – cross with Cre expressing
line and analyze function

BioSci 145B lecture 9 page 5 ©copyright Bruce Blumberg 2004. All rights reserved
Conditional gene targeting (contd)
– Tissue- or stage-specific knockouts from crossingfloxed mouse with specific Cre-expressing line
– requirement for Cre lines• must be well
characterized– promoters can’t
be leaky• Andras Nagy’s
database of Cre lines and other knockout resources http://www.mshri.on.ca/nagy/cre.htm

BioSci 145B lecture 9 page 6 ©copyright Bruce Blumberg 2004. All rights reserved
Conditional gene targeting (contd)
• advantages– can target recombination to specific tissues and times– can study genes that are embryonic lethal when disrupted– can use for marker eviction– can study the role of a single gene in many different tissues with a single
mouse line– can use for engineering translocations and inversions on chromosomes
• disadvantages– not trivial to set up, more difficult than std ko but more information
possible– requirement for Cre lines
• must be well characterized regarding site and time of expression• promoters can’t be leaky (expressed when not intended)

BioSci 145B lecture 9 page 7 ©copyright Bruce Blumberg 2004. All rights reserved
Genome wide analysis of gene function
• How to mutate all genes in a given genome?– Easy with microbial genomes – can mutate all yeast genes by homologous
recombination– Recombine in selectable marker– Propagate strain and analyze phenotypes
Homology region
Target gene
Selectable marker(antibiotic resistance)
Unique oligonucleotide“barcodes” for PCR

BioSci 145B lecture 9 page 8 ©copyright Bruce Blumberg 2004. All rights reserved
Genome wide analysis of gene function (contd)
• How about gene targeting in other organisms– With more complex genomes and more genes?– Not really feasible to specifically target 20-30K genes
• Difficulty• Expense• Inability to target all possibile loci
– Some efforts to make mouse collection • Lexicon Genetics has a collection
– Drosophila collection as well– Driving force behind these efforts is
• Genome annotation• Drug target discovery (Lexicon)• Functional analysis

BioSci 145B lecture 9 page 9 ©copyright Bruce Blumberg 2004. All rights reserved
Genome wide analysis of gene function (contd)
• Main method for gene targeting in more complex organisms is randominsertional mutagenesis– Transposon mutagenesis
• Bacteria – Tn transposons• Yeast - Ty transposons• Drosophila - P- elements• Vertebrates - Sleeping Beauty transposons
– Viral infection• Typically retroviruses – host range selectivity is obstacle
– Gene or enhancer trapping– modified virusesor transposons

BioSci 145B lecture 9 page 10 ©copyright Bruce Blumberg 2004. All rights reserved
Insertional mutagenesis - Gene trapping
• viruses and transposable elements can deliver DNA to random locations– can disrupt gene function – put inserted gene under the
control of adjacent regulatory sequences
– BOTH
• enhancer trap is designed to bring inserted reporter gene under the control of local regulatory sequences– put a reporter gene adjacent to a weak promoter (enhancer-less), e.g. a
retrovirus with enhancers removed from the LTRs– may or may not disrupt expression

BioSci 145B lecture 9 page 11 ©copyright Bruce Blumberg 2004. All rights reserved
Insertional mutagenesis - Gene trapping (contd)
• enhancer trap (contd)– expression only when integrate into an active transcription unit
• reporter expression duplicates the temporal and spatial pattern of the endogenous gene
– reporters used• β-gal was the most widely used reporter• GFP is now popular• β-lactamase is seeing increasing use
– advantages• relatively simple to perform• active promoters frequently targeted, perhaps due to open chromatin
– Disadvantages• Inactive promoters probably not targeted • insertional mutagenesis not the goal, and not frequent
– overall frequency is not that high• relies on transposon or retroviruses to get insertion
– may not be available for all systems, requires transgenesis or good viral vectors

BioSci 145B lecture 9 page 12 ©copyright Bruce Blumberg 2004. All rights reserved
Insertional mutagenesis – Gene trapping (contd)
• expressed gene trap (many variations possible)– goal -> ablate expression of endogenous gene, replace with transgene– Make insertion construct – reporter, selection, polyA sites
• No promoter but has a splie-acceptor sequence 5’ of reporter• Can only be expressed if spliced into an endogenous mRNA
– Transfer into embryonic cells, generate a library of insertional mutagens• Mouse, Drospophila, zebrafish, frog
– reporter expression duplicates the temporal and spatial pattern of the endogenous gene
• As in Golling paper we heard about on Thursday

BioSci 145B lecture 9 page 13 ©copyright Bruce Blumberg 2004. All rights reserved
Insertional mutagenesis - Gene trapping (contd)
• Expressed gene trapping (contd)– advantages
• insertional mutagen– gives information about expression patterns– can be homozygosed to generate phenotypes
• higher efficiency than original trapping methods• selectable markers allow identification of mutants
– many fewer to screen– dual selection strategies possible
– disadvantages• overall frequency is still not that high• frequency of integration into transcription unit is not high either• relies on transposon or retroviruses to get insertion
– may not be available in your favorite system.– Uses
• Insertional mutagenesis• Marking genes to id interesting ones• Gene cloning

BioSci 145B lecture 9 page 14 ©copyright Bruce Blumberg 2004. All rights reserved
Generating phenocopies of mutant alleles
• How to inactivate endogenous genes in a targeted but general way?– Important new development is RNAi – RNA
interference– Observation is that introduction of double-
stranded RNAs into cells lead to destruction of corresponding mRNA (if there is one)
– Principle is siRNA – small interfering RNAs– These generate small single stranded RNAs that
target mRNAs for destruction by– RISC – RNA interference silencing complex– First applied in C. elegans where it works
extremely well• Can introduce siRNA into cells even by
feeding to the worms!• Works very well in Drosophila• poorly in mammalian cells• Poorly in Xenopus
BioSci 145B lecture 9 page 15 ©copyright Bruce Blumberg 2004. All rights reserved
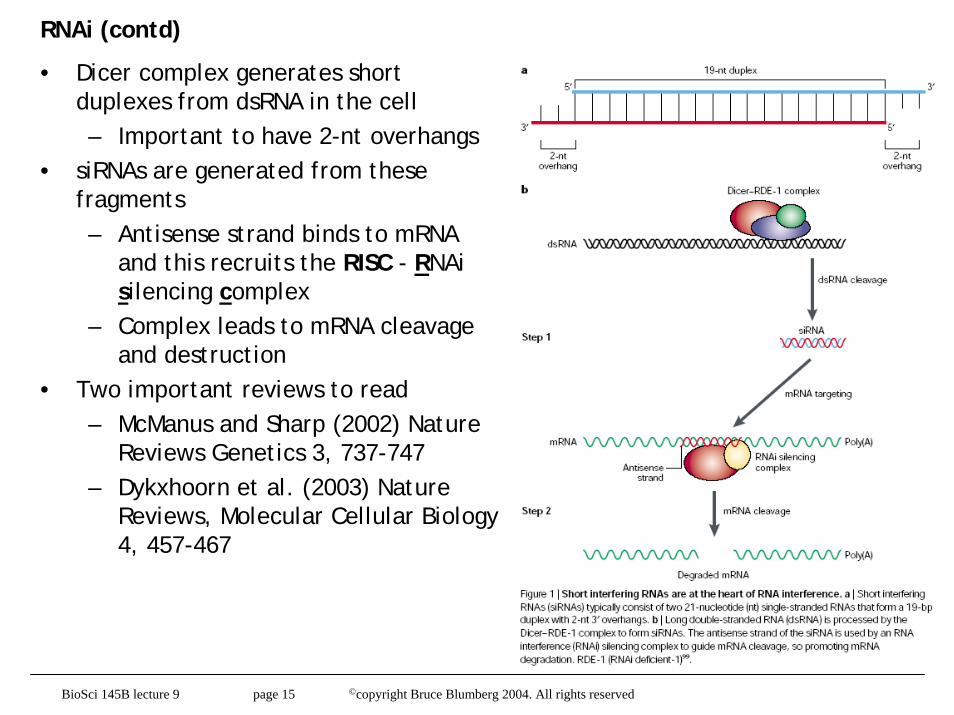
RNAi (contd)
• Dicer complex generates short duplexes from dsRNA in the cell– Important to have 2-nt overhangs
• siRNAs are generated from these fragments– Antisense strand binds to mRNA
and this recruits the RISC - RNAisilencing complex
– Complex leads to mRNA cleavage and destruction
• Two important reviews to read– McManus and Sharp (2002) Nature
Reviews Genetics 3, 737-747– Dykxhoorn et al. (2003) Nature
Reviews, Molecular Cellular Biology 4, 457-467

RNAi (contd)
• Micro RNAs are small cellular RNAs that previously lacked any known function– Always form a hairpin structure with
mismatches in stem• Turn out that micro RNAS direct gene
silencing via translational repression– (miRNAs) are mismatched duplexes
that dicer processes into stRNAs– Use the same cellular complex as
siRNAs– Perfect matches -> target cleavage– Imperfect matches -> translational
repression of target
BioSci 145B lecture 9 page 16 ©copyright Bruce Blumberg 2004. All rights reserved
BioSci 145B lecture 9 page 17 ©copyright Bruce Blumberg 2004. All rights reserved
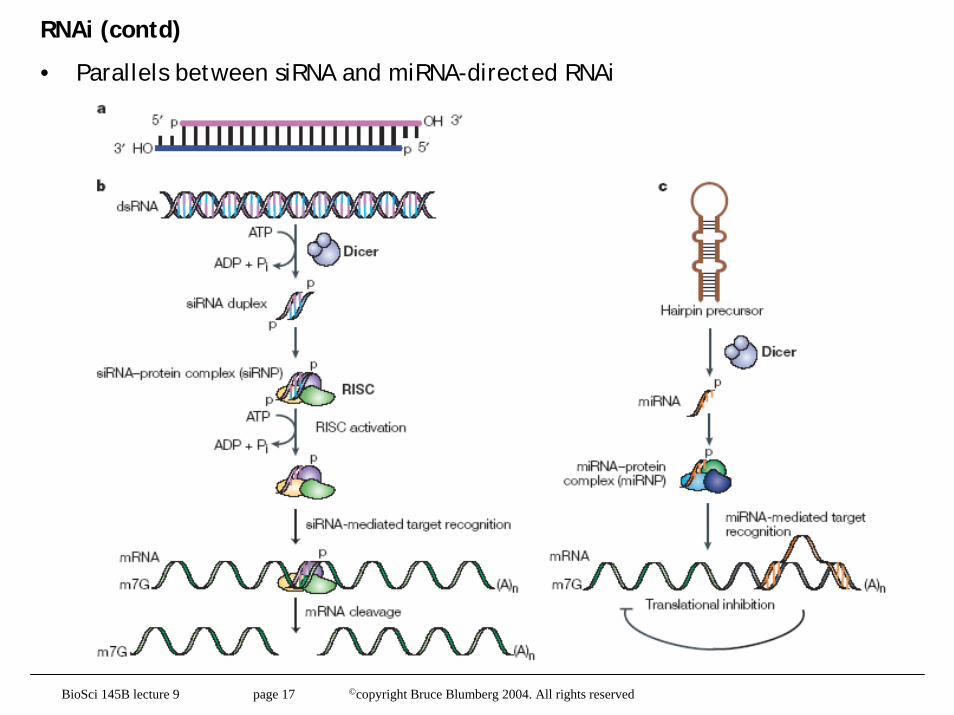
RNAi (contd)
• Parallels between siRNA and miRNA-directed RNAi

BioSci 145B lecture 9 page 18 ©copyright Bruce Blumberg 2004. All rights reserved
RNAi (contd)
• Ways to generate short RNAs that silence gene expression in vitro– a) chemical synthesis of siRNA, introduce into cell– b) synthesize long dsRNA, use dicer to chop into siRNA– c) introduce perfect duplex hairpin, dicer generates siRNA– d) make miRNA based hairpin, dicer generates silencing RNA
• Introduce into cells or organism by microinjection, transfection, etc.– Expression is transient– can only generate phenotypes for a short time after introduction

BioSci 145B lecture 9 page 19 ©copyright Bruce Blumberg 2004. All rights reserved
RNAi (contd)
• Ways to generate short silencing RNAs in vivo– Continuing expression to generate stable phenotype– a) produce long hairpin
from pol II promoter, letdicer make siRNA
– b) produce two transcriptsfrom pol III promoter, letanneal in cells
– c) produce a short hairpinfrom pol III promoter, letdicer generate siRNAs
– d) produce imperfecthairpin from pol II promoter, let dicergenerate miRNAs that direct gene silencing

BioSci 145B lecture 9 page 20 ©copyright Bruce Blumberg 2004. All rights reserved
RNAi (contd)
• RNAi for whole genome functional analysis– First generate library of constructs that generates siRNA or stRNA– Introduce these into cells, embyos (fly, frog, mouse) or animals (C.
elegans, plants)• For C. elegans, make the library in E. coli and simply feed bacteria to
worms• Must microinject or transfect with other animals
– Evaluate phenotypes

BioSci 145B lecture 9 page 21 ©copyright Bruce Blumberg 2004. All rights reserved
Antisense methods to knock out gene function
• Antisense oligonucleotides can transiently target endogenous RNAs– For destruction
• Many methods and oligo chemistries available• Most are very sensitive to level of antisense oligo, these are degraded
and rapidly muck up cellular nucleotide pools leading to toxicity– For translational inhibition
• Morpholino oligos appear to work the best– Morpholine sugar is substituted for deoxyribose– Is not a substrate for cellular DNAses or RNAse H– Base-pairs with RNA or DNA more avidly than standard DNA– The oligo binds to the area near the ATG in the transcript and
inhibits translation of the protein
– Deoxyribose morpholine
N
OO

BioSci 145B lecture 9 page 22 ©copyright Bruce Blumberg 2004. All rights reserved
Antisense methods to knock out gene function (contd)
Oligodeoxyribonucleotide Morpholino Oligonucleotide
B = A, C, T, G
N
O B
P N
O
O
N
O B
O
P N
O
O
O
O
OB
O
O
P O
O
O
B
P O
O
O

BioSci 145B lecture 9 page 23 ©copyright Bruce Blumberg 2004. All rights reserved
Most Molecules Function in Complexes
• Given a target, how can we identify interacting proteins?
• Complex members may be important new targets– pharmacology– toxicology– Endocrine disrupter action
• High throughput, genome wide screen is preferred– 20 years is too long

BioSci 145B lecture 9 page 24 ©copyright Bruce Blumberg 2004. All rights reserved
How can we approach whole genome analysis of protein complex formation?
• Each protein interacts with average of 3 others
• Many are much more complex
• Two papers this Thursday and one next Thursday describe two different approaches to this problem.

BioSci 145B lecture 9 page 25 ©copyright Bruce Blumberg 2004. All rights reserved
How to identify protein-protein interactions on a genome wide scale?
• You have one protein and want to identify proteins that interact with it– straight biochemistry
• Co-immunoprecipitation• GST-pulldown
– Library based methods• phage display• Yeast two hybrid• in vitro expression cloning
• You want to identify all proteins that interact with all other proteins– Proteomic analysis– Protein microarrays– Large scale two-hybrid

BioSci 145B lecture 9 page 26 ©copyright Bruce Blumberg 2004. All rights reserved
Mapping protein-protein interactions• biochemical approach
– purify cellular proteins that interact with your protein• co-immunoprecipitation• GST-pulldown• affinity chromatography• biochemical fractionation
– pure protein(s) are microsequenced
– advantage• functional approach• stringency can be manipulated• can identify multimeric proteins or complexes• will work if you can purify proteins
– disadvantages• much skill required• low throughput• considerable optimization required

BioSci 145B lecture 9 page 27 ©copyright Bruce Blumberg 2004. All rights reserved
Mapping protein-protein interactions (contd)
• GST (glutathione-S-transferase) pulldown assay– Versatile and general– Fuse protein of interest
to GST– Incubate with cell or
tissue extracts– Mix with glutathione-
sepharose beads• Binds GST-fusion
protein and anything bound to it
– Run SDS-PAGE– Identify bands
• Co-Ip (immunoprecipitation) is identical except that antibody is used to pull down protein X

BioSci 145B lecture 9 page 28 ©copyright Bruce Blumberg 2004. All rights reserved
Mapping protein-protein interactions (contd)
• scintillation proximity assay– Target is bound to solid phase –
bead or plate– radioactive protein or ligand is added
and allowed to reach equilibrium• 35S, 125I, 3H work best
– radioactive decay is quenched in solution, only detected when in “proximity” of the solid phase, e.g. when bound to target
– applications• ligand-receptor binding with 3H small molecules• protein:protein interaction• protein:DNA

BioSci 145B lecture 9 page 29 ©copyright Bruce Blumberg 2004. All rights reserved
Mapping protein-protein interactions (contd)
• FRET - fluorescent resonance energy transfer– based on the transfer of energy
from one fluor to another that is not normally excited at that wavelength
– Many types of fluorescent moieties possible
• rare earth metals – europium cryptate
• fluorescent proteins – GFP and variants– allophycocyanin
• Tryptophan residues in proteins– application
• very commonly used for protein:protein interaction screening in industry
• FRET microscopy can be used to prove interactions between proteins within single cells
– Roger Tsien

BioSci 145B lecture 9 page 30 ©copyright Bruce Blumberg 2004. All rights reserved
Mapping protein-protein interactions (contd)
• FRET (contd)– advantages
• can be very sensitive• may be inexpensive or not depending on materials• non-radioactive• equilibrium assay• single cell protein:protein interactions possible• time resolved assays possible
– disadvantage• poor dynamic range - 2-3 fold difference full scale• must prepare labeled proteins or ligands
– Not suitable for whole genome analysis• tunable (or multiwavelength capable) fluorometer required (we have
one here)

BioSci 145B lecture 9 page 31 ©copyright Bruce Blumberg 2004. All rights reserved
Mapping protein-protein interactions (contd)
• Biacore (surface plasmon resonance)– surface plasmon waves are excited at a metal/liquid interface– Target bound to a thin metal foil and test sample flowed across it– Foil is blasted by a laser from behind
• SPR alters reflected light intensity at a specific angle and wavelength• Binding to target alters refractive index which is detected as change
in SPR• Change is proportional to change in mass and independent of
composition of binding agent

BioSci 145B lecture 9 page 32 ©copyright Bruce Blumberg 2004. All rights reserved
Mapping protein-protein interactions (contd)
• Biacore (contd)– Advantages
• Can use any target• Biological extracts possible• Measure kinetics• Small changes detectable with correct instrument
– 360 d ligand binding to 150 kd antibody• Can use as purification and identification system
– Disadvantages• Machine is expensive (we have two)• “high throughput” very expensive• Not trivial to optimize

BioSci 145B lecture 9 page 33 ©copyright Bruce Blumberg 2004. All rights reserved
Library-based methods to map protein-protein interactions (contd)
• Phage display screening (a.k.a. panning)– requires a library that expresses
inserts as fusion proteins with a phage capsid protein
• most are M13 based• some lambda phages used
– prepare target protein• as affinity matrix• or as radiolabeled probe
– test for interaction with library members• if using affinity matrix you purify phages from a mixture• if labeling protein one plates fusion protein library and probes with
the protein– called receptor panning based on similarity with panning for
gold

BioSci 145B lecture 9 page 34 ©copyright Bruce Blumberg 2004. All rights reserved
Library-based methods to map protein-protein interactions (contd)
• Phage display screening (a.k.a. panning) (contd)– advantages
• stringency can be manipulated• if the affinity matrix approach works the cloning could go rapidly
– disadvantages• Fusion proteins bias the screen against full-length cDNAs• Multiple attempts required to optimize binding• Limited targets possible• may not work for heterodimers• unlikely to work for complexes• panning can take many months for each screen
– Greg Weiss in Chemistry is local expert

BioSci 145B lecture 9 page 35 ©copyright Bruce Blumberg 2004. All rights reserved
Mapping protein-protein interactions (contd)
• Two hybrid screening– originally used in yeast, now
other systems possible– prepare bait - target protein
fused to DBD (GAL4) usual• stable cell line is commonly
used– prepare fusion protein library
with an activation domain - prey– Key factor required for success is
no activation domain in bait!
– approach• transfect library into cells and either
select for survival or activation of reporter gene
• purify and characterize positive clones
BioSci 145B lecture 9 page 36 ©copyright Bruce Blumberg 2004. All rights reserved
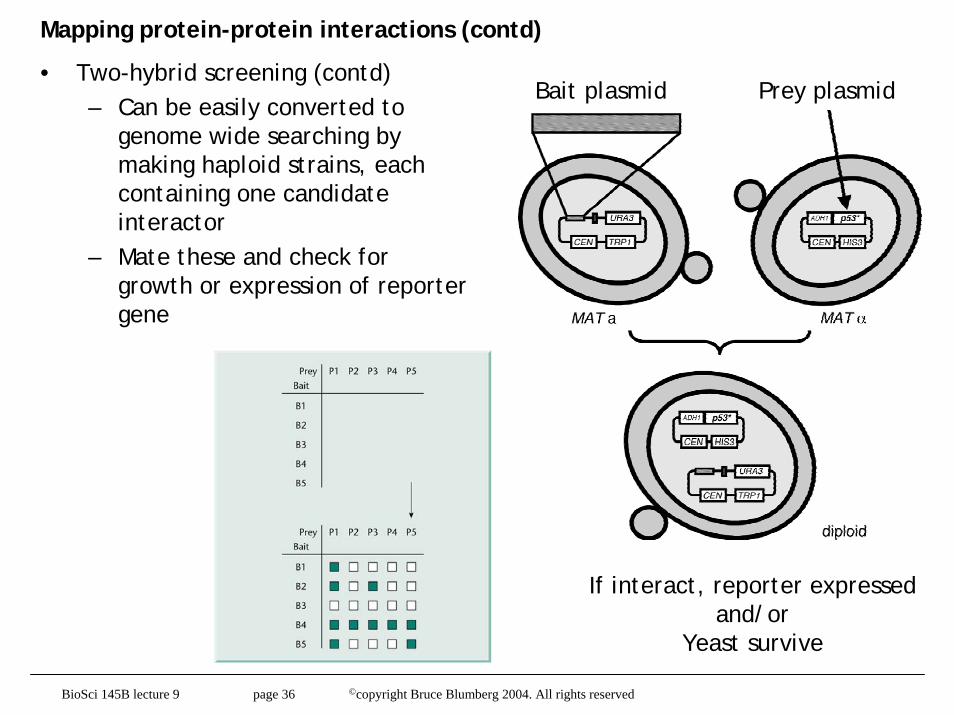
Mapping protein-protein interactions (contd)
• Two-hybrid screening (contd)– Can be easily converted to
genome wide searching by making haploid strains, each containing one candidate interactor
– Mate these and check for growth or expression of reporter gene
Bait plasmid Prey plasmid
If interact, reporter expressed and/or
Yeast survive

BioSci 145B lecture 9 page 37 ©copyright Bruce Blumberg 2004. All rights reserved
Mapping protein-protein interactions (contd)
• In vitro interaction screening - based on in vitro expression cloning (IVEC)– transcribe and translate cDNAs in vitro into small pools of proteins (~100)– test for their ability to interact with your protein of interest
• EMSA• co-ip• FRET• SPA
– advantages• functional approach• smaller pools increase sensitivity• diversity of targets
– proteins, complexes, nucleic acids, protein/nucleic acid complexes, small molecule drugs
– very fast– disadvantages
• can’t detect heterodimers unless 1 partner known• expensive consumables (but cheap salaries)
– Typical screen may cost $10-15K• expense of automation





























