Embed Size (px)
Citation preview

Biology of Ixodes rubicundus ticks under laboratoryconditions: observations on oviposition and eggdevelopment
F.J. VAN DER LINGENa, L.J. FOURIEa* D.J. KOKa and J.M. VAN ZYLb
a Department of Zoology and Entomology, University of the Orange Free State, PO Box 339,Bloemfontein, 9300, South Africab Department of Mathematical Statistics, University of the Orange Free State, PO Box 339,Bloemfontein, 9300, South Africa
(Received 17 February 1998; accepted 6 October 1998)
Abstract. Observations on oviposition and egg development of Ixodes rubicundus were made underlaboratory conditions. Engorged females were exposed to temperatures in the range 10–25°C and relativehumidities (RHs) of 33 and 93%. The pre-oviposition period, oviposition period, incubation period,conversion efficiency index (CEI) values and fecundity were determined. The mean pre-ovipositionperiod varied from 13.3 days (temperature 25°C and RH 33%) to 68.3 days (temperature 10°C and RH93%). Oviposition extended from a mean of 39 days (temperature 25°C and RH 93%) to 201.7 days(temperature 10°C and RH 93%). The developmental zero temperature for the pre-oviposition period was9.2°C. The mean total number of eggs produced by engorged I. rubicundus females varied from 2045.7(temperature 10°C and RH 93%) to 3777.7 (temperature 20°C and RH 93%). Both female mass and RHsignificantly (p , 0.01) influenced the number of eggs produced. CEI values varied between 43.1–54.4%(RH 93%) and 34.1–42.5% (RH 33%). At 93% RH females produced between 14.2 and 17.7 eggs per mgbody mass compared to the 13.2–14.6 eggs per mg body mass at 33% RH. The shortest mean incubationperiod recorded was 164.3 days (temperature 25°C and RH 93%). The developmental zero temperaturefor incubation was 6.5°C. Both the pre-oviposition and oviposition periods of I. rubicundus are moreextended compared to other species of the genus. Ixodes rubicundus produces a large number of smalleggs compared to other prostriate ticks.Exp Appl Acarol 23: 513–522 © 1999 Kluwer Academic Publishers
Key words: Ixodes rubicundus, pre-oviposition period, oviposition period, egg yield, conversionefficiency index, fecundity, laboratory study.
Introduction
The Karoo paralysis tick Ixodes rubicundus is of considerable economic importancein South Africa because it causes paralysis in a variety of domestic stock and wildanimals (Spickett and Heyne, 1988; Fourie and Vrahimis, 1989). It inhabits mainlyrocky outcrops and occurs in close association with certain plant species (Stampa,1959; Fourie et al., 1991).
* To whom correspondence should be addressed.
Experimental and Applied Acarology 23: 513–522, 1999.© 1999 Kluwer Academic Publishers. Printed in the Netherlands.

The tick’s geographic distribution range is characterized in some regions by winterand in others by equinoctial rainfall, with mean annual rainfall varying from 100 to600 mm. Mild to severe droughts occur periodically. Air temperatures show majordiurnal and seasonal variations with temperatures as low as 214°C or as high as41°C during winter and summer, respectively. A difference of 25°C betweenmaximum day and minimum night temperatures may occur (Venter et al., 1986). Itis evident that I. rubicundus, in its natural habitat, is subjected to adverse conditionsduring large parts of the year. A field study on the life cycle of the tick has shownthat it extends over 2 years. The two regulating phases in the life cycle of the tick,which undergoes a developmental diapause during the hot and often dry summermonths, are the egg and engorged nymph (Fourie and Horak, 1994).
Comparatively little research on the life cycle of I. rubicundus under controlledlaboratory conditions has been conducted. A better understanding of certain aspectsof its life cycle could contribute towards elucidating its current and potentialbiogeographic distribution, seasonality and mortality factors. The purpose of thepresent controlled laboratory study was to investigate the effect of selected tem-perature and relative humidity regimes on the duration of the pre-oviposition andoviposition periods, egg yield and eclosion in I. rubicundus.
Material and methods
Engorged, female I. rubicundus ticks were collected from Angora goats during Apriland May 1993 and 1994 on the farm Preezfontein, situated 10 km from the townFauresmith (29° 46'S, 25° 19'E) in the south western Free State province of SouthAfrica. Only fully engorged, recently detached females that were still retained in thedense and curly hair of the goats were used. Limited numbers of such ticks wereavailable for the study. The ticks were weighed and placed individually intonumbered cylindrical (15 3 30 mm) Perspex containers (UOFS, instrumentationworkshop). The open ends of the containers were sealed with nylon gauze (Nyboltbolting cloth with pores of 200 mm). Each container consisted of two equally-sizedparts which were screwed together. The small, Perspex containers with ticks wereplaced in larger containers in which the relative humidity (RH) could be controlledthrough the use of saturated salt solutions (Winston and Bates, 1960). Temperature-controlled incubation cabinets were used to expose females in groups of three or fourto temperatures in the range 10–25°C (5°C intervals) and RHs of 33 and 93 6 2% foreach temperature interval. The containers were kept in constant darkness.
Each engorged female was examined every second day to determine the onset ofoviposition, following which each female was examined daily. The eggs depositedby each female were counted with the aid of a stereomicroscope and then transferredto a correspondingly numbered, 7 ml glass vial, covered with 200 mm pore size nylongauze (Nybolt bolting cloth) to ensure sufficient circulation. These vials were placed
514

in the same container as the females. At the completion of egg laying, egg batcheswere weighed to the nearest 0.01 mg. Because of the range of conditions to whicheggs were exposed during the extended oviposition period some egg batches driedout or became infected with fungi. These batches, although counted, could not beweighed. This affected the number of egg batches available for the calculation of theconversion efficiency index (CEI) (see below).
Egg batches used to determine the time to eclosion were not counted or weighedin order to leave them undisturbed. Three to seven egg batches were available for thedetermination of incubation period. Egg batches were monitored twice a week andthe incubation period was taken as the time (in days) from laying of the first egg toeclosion of the first larva. The experimental conditions were extended to include30°C also.
The CEI (Drummond and Whetstone, 1970) was expressed as a percentage torepresent the percentage body mass (engorged female) converted to egg mass.Because some egg batches dried out or became infected with fungi (see above) datawere limited for some of the temperature and humidity regimes (see column n inTable 3).
CEI (%) 5 (m1/m2) 3 100
where m1 is the egg batch mass (g) and m2 is the engorged female mass (g).Fecundity, expressed as the mean number of eggs per mg female engorged mass,
was determined for females at each temperature and RH.The data obtained are presented as mean values. An ANOVA was used to
determine the influence of engorgement mass, temperature and humidity on the pre-oviposition and oviposition periods. A multiple regression analysis and ANOVAwere used to determine the effect of female engorgment mass, temperature andhumidity on the number of eggs laid. A critical low temperature (developmentalzero) for each developmental stage was calculated from the regression equationbetween temperature and the developmental velocity (reciprocal of days of pre-oviposition or incubation period).
Results
Because limited numbers of fully engorged female ticks were available for exposureto a wide range of experimental conditions, only three to four ticks were used at eachtemperature/RH regime. Furthermore, some egg batches were affected by lowhumidity, high temperature, fungal growth or combinations of these, particularly inthose cases where the eggs had to be kept under experimental conditions forextended periods. Therefore, although at least three to four ticks were used for eachcondition in a few cases the results were based on more limited observations.
515

The mean pre-oviposition period varied from 13.3 (temperature 25°C and RH33%) to 68.3 (temperature 10°C and RH 93%) days with minimum and maximumpre-oviposition periods of 12 (temperature 25°C and RH 93%) and 75 days(temperature 10°C and RH 93%) respectively (Table 1). Oviposition extended froma mean of 39 (temperature 25°C and RH 93%) to 201.7 (temperature 10°C and RH93%) days (Table 1). The minimum and maximum oviposition periods were 34(temperature 25°C and RH 33%) and 288 (temperature 10°C and RH 93%) days,respectively (Table 1).
Temperature significantly (p , 0.05) influenced the pre-oviposition and oviposi-tion periods. Mass and RH, however, had no significant effect. The relationship(r 5 0.8059) between temperature and the reciprocal of the pre-oviposition period isgiven by the following equation:
y 5 b 3 log(x 2 a)
where a 5 8.15, b 5 2.68, y is 1/pre-oviposition period (days) and x is the tem-perature (°C). A developmental zero temperature of 9.2°C was calculated.
The oviposition pattern of females maintained at 20°C and 93% RH is presentedin Fig. 1. The number of eggs deposited rose sharply to reach a mean peakproduction of 174 eggs day–1 on day 10, after which it decreased gradually up to day45. Few eggs (, 25) were deposited daily from day 45 up to day 67 (Fig. 1). Themean total number of eggs produced by engorged I. rubicundus females varied from2045.7 (temperature 10°C and RH 93%) to 3777.7 (temperature 20°C and RH 93%).The absolute minimum and maximum numbers of eggs produced were 1717 and5092, respectively (Table 2). Both female mass and RH significantly (p , 0.01)influenced the number of eggs produced. The relationship (r 5 0.83) between femalemass and number of eggs is reflected by the following linear regression equation:
y 5 a 1 bx
Table 1. Pre-oviposition and oviposition periods (days) of engorged I. rubicundus females exposed todifferent temperature and RH regimes
Pre-oviposition period (days) Oviposition period (days)Temperature(°C)
RH(±2%)
Femalex– mass(g) x– Minimum Maximum x– Minimum Maximum n
10 93 0.116 68.3 64.0 75.0 201.7 133 288 310 33 0.180 58.3 57.0 60.0 116.3 105 136 315 93 0.205 16.0 15.0 17.0 49.7 46 52 315 33 0.195 19.7 16.0 23.0 50.7 46 59 320 93 0.112 17.0 15.0 19.0 61.3 51 67 320 33 0.166 18.3 16.0 22.0 48.0 42 57 325 93 0.187 14.0 12.0 20.0 39.0 36 44 425 33 0.170 13.3 13.0 14.0 60.7 34 108 3
516

where a 5 2989.602, b 5 20324.95, y is the number of eggs and x is the femalemass (g).
The number of eggs produced by females exposed to different temperature and RHregimes can be calculated by using the following multiple regression equation:
y 5 ax1 1 bx2 1 cx3 1 d
where a 5 21.8, b 5 400.8, c 5 19 138.07, d 5 21342.419, y is the number of eggs,x1 is the temperature (°C) (r 5 0.006), x2 is the RH (%) (r 5 0.1849) and x3 is thefemale mass (g) (r 5 0.8678).
CEI values for females exposed to the different temperatures and RHs are given inTable 3. RH and CEI values were significantly (p 5 0.0266) correlated (r 5 0.7662)whereas temperature and CEI values were not significantly (p 5 0.8549) correlated(r 5 0.0777). At an RH of 93% between 43.1 and 54.4% of the engorged body mass
Figure 1. Mean (±S.E.) number of eggs laid per day by I. rubicundus females at 20°C and 93% RH.
Table 2. Total numbers of eggs laid and fecundity of I. rubicundus females
Egg number
Temperature (°C) RH (±2%)Female x–
mass (g) x– Minimum Maximum Fecundity n
10 93 0.144 2045.7 1717 2590 14.20 310 33 0.180 2382.7 1869 2985 13.20 315 93 0.198 3501.0 3111 3858 17.70 315 33 0.189 2661.7 1879 3520 14.10 320 93 0.221 3777.7 1791 5092 17.10 320 33 0.166 2277.7 2074 2385 13.72 325 93 0.187 2805.0 2592 3224 15.00 425 33 0.170 2477.0 2267 2719 14.60 3
517

was converted into eggs. At an RH of 33% this conversion varied between 34.1 and42.5%. The fecundity (x number of eggs/x mg body mass) for females exposed to thedifferent temperatures and RH is given in Table 2. Engorged females at an RH of93% produced between 14.2 and 17.7 eggs per mg body mass. At an RH of 33% thefecundity varied between 13.2 and 14.6. Neither temperature (p 5 0.6203 andr 5 0.2085) nor RH (p 5 0.0557 and r 5 0.6951) were significantly correlated withfecundity.
The relationship between temperature, RH and mean egg incubation period issummarized in Table 4. No eggs hatched at either an RH of 33% or a temperature of30°C. Only one out of six egg batches exposed at temperature 10°C and RH 93%hatched after 562 days. The shortest mean incubation period recorded was 164.3 daysat 25°C. The relationship (r 5 0.3964) between temperature and the reciprocal of theincubation period of eggs is described by the following equation:
y 5 b 3 log(x 2 a)
where a 5 6.53, b 5 2.19, y is 1/incubation time (days) and x is the temperature (°C).The developmental zero temperature for incubation was calculated as 6.5°C.
Table 3. Engorged I. rubicundus female mass, egg mass and CEI values (%)
Female Mass (g) Egg Mass (g)Temperature(°C)
RH(±2%) x– Minimum Maximum x– Minimum Maximum CEI (%) n
10 93 – 0.116 0.116 – 0.050 0.050 43.1 110 33 0.180 0.169 0.200 0.061 0.044 0.082 34.1 315 93 0.205 0.202 0.208 0.112 0.111 0.112 54.4 215 33 0.195 0.155 0.235 0.083 0.072 0.093 42.5 220 93 – 0.112 0.112 0.054 – 0.054 48.1 120 33 0.166 0.170 0.174 0.057 0.052 0.062 34.2 325 93 0.187 0.166 0.218 0.083 0.075 0.096 44.5 425 33 0.170 0.159 0.192 0.069 0.060 0.076 40.5 3
Table 4. Incubation period of I. rubicundus eggs
Incubation period (days)
Temperature (°C) RH (±2%) x– Minimum Maximum n
10 93 – 562 562 110 33 –a – – 615 93 180.0 131 201 615 33 – – – 620 93 170.5 138 190 620 33 – – – 625 93 164.3 150 179 725 33 – – – 630 93 – – – 330 33 – – – 3
a No hatching of eggs.
518

Discussion
A decrease in pre-oviposition time for I. rubicundus with an increase in temperatureis in accordance with that generally found for ixodid ticks (Fujimoto, 1992;Guglielmone, 1992). In comparison to other Ixodes species the mean pre-ovipositionperiod of 14 days recorded for I. rubicundus at 22–25°C is long. The mean pre-oviposition periods recorded for Ixodes persulcatus (Fujimoto, 1992), Ixodes hex-agonus (Toutoungi et al., 1995) and Ixodes ricinus (MacLeod, 1935) were 5.8, 8.4and 10 days, respectively. A possible explanation for the longer than expected pre-oviposition period is perhaps the fact that I. rubicundus eggs undergo a devel-opmental diapause (Fourie and Horak, 1994). The delay in onset of oviposition maybe a reflection of reduced metabolic activity in engorged females. The developmentalzero or critical low temperature for the onset of oviposition in I. rubicundus is high(9.2°C) compared to the 5.6 and 2.9°C for Ixodes nipponensis and I. persulcatus,respectively (Fujimoto, 1992). A value of 8.2°C for Ixodes ovatus, which occurs inmore moderate environments, has been recorded (Fujimoto, 1989). The availabledata suggest that the developmental zero temperature is a reflection of the environ-ment in which the tick occurs. Lower values are adaptations for cool conditions.
As with pre-oviposition, the mean oviposition period for I. rubicundus at 23–25°Cis long (39 days) compared to other Ixodes species. Values of 19.1 and 22 days forI. ovatus and I. ricinus, respectively, have been recorded (MacLeod, 1935; Fujimoto,1989). In the case of I. hexagonus, egg laying of mated females lasted for 8–22 days,with the majority of the eggs being deposited during the first 6 days of theoviposition period (Toutoungi et al., 1995). The mean number of eggs laid by I.rubicundus is greater compared to other Ixodes species (Table 5) and may accountfor the comparatively longer oviposition period. With reference to developmentalzero temperatures for incubation, the value is normally close to that recorded foroviposition (Yano et al., 1987; Fujimoto 1989, 1992). The 6.5°C recorded for I.rubicundus is lower than expected and is probably due to a small sample size foreggs incubated at 10°C.
Available records show that Ixodes species convert between 40.3 and 54.4% oftheir engorged body mass into eggs. Compared to Ixodes species, values forAmblyomma species (between 53.3 and 65%) and Hyalomma truncatum (56%) seemto be higher, with a CEI for Hyalomma dromedarii being much higher (74%). TheCEI values for Aponomma hydrosauri (48.6%) and Boophilus annulatus (53.3 and44%) were in the same range as that recorded for Ixodes (Table 5). Temperature didnot affect CEI values in H. dromedarii (Hagras and Khalil, 1988). There is a trend forthe CEI in I. rubicundus to increase with a decrease in temperature from 25 to 15°C.These measurements may, however, be artefactual, since at higher temperaturesmoisture loss from eggs would have been considerably greater, resulting in lowerCEI values. Furthermore, the parameter estimates are based on limited observations
519
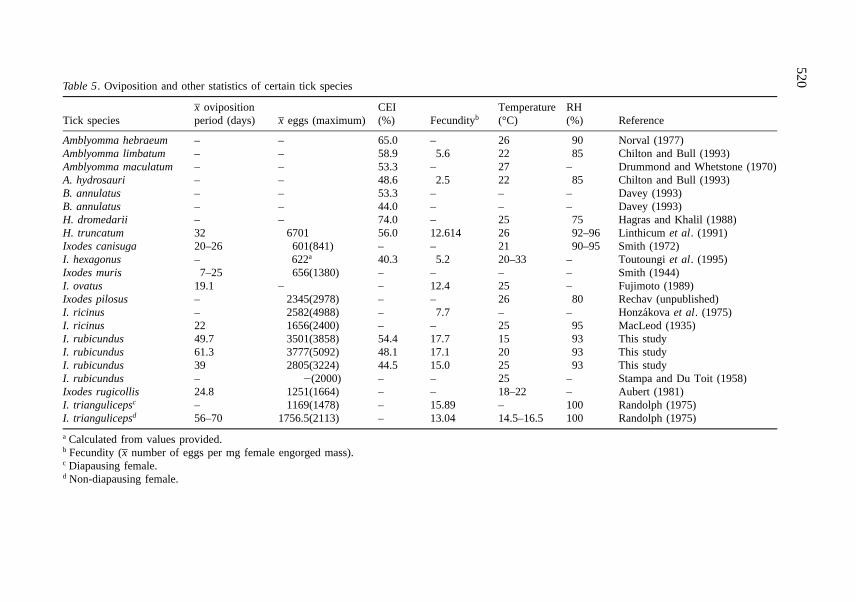
Table 5. Oviposition and other statistics of certain tick species
Tick speciesx– ovipositionperiod (days) x– eggs (maximum)
CEI(%) Fecundityb
Temperature(°C)
RH(%) Reference
Amblyomma hebraeum – – 65.0 – 26 90 Norval (1977)Amblyomma limbatum – – 58.9 5.6 22 85 Chilton and Bull (1993)Amblyomma maculatum – – 53.3 – 27 – Drummond and Whetstone (1970)A. hydrosauri – – 48.6 2.5 22 85 Chilton and Bull (1993)B. annulatus – – 53.3 – – – Davey (1993)B. annulatus – – 44.0 – – – Davey (1993)H. dromedarii – – 74.0 – 25 75 Hagras and Khalil (1988)H. truncatum 32 6701 56.0 12.614 26 92–96 Linthicum et al. (1991)Ixodes canisuga 20–26 601(841) – – 21 90–95 Smith (1972)I. hexagonus – 622a 40.3 5.2 20–33 – Toutoungi et al. (1995)Ixodes muris 7–25 656(1380) – – – – Smith (1944)I. ovatus 19.1 – – 12.4 25 – Fujimoto (1989)Ixodes pilosus – 2345(2978) – – 26 80 Rechav (unpublished)I. ricinus – 2582(4988) – 7.7 – – Honzakova et al. (1975)I. ricinus 22 1656(2400) – – 25 95 MacLeod (1935)I. rubicundus 49.7 3501(3858) 54.4 17.7 15 93 This studyI. rubicundus 61.3 3777(5092) 48.1 17.1 20 93 This studyI. rubicundus 39 2805(3224) 44.5 15.0 25 93 This studyI. rubicundus – 2(2000) – – 25 – Stampa and Du Toit (1958)Ixodes rugicollis 24.8 1251(1664) – – 18–22 – Aubert (1981)I. triangulicepsc – 1169(1478) – 15.89 – 100 Randolph (1975)I. triangulicepsd 56–70 1756.5(2113) – 13.04 14.5–16.5 100 Randolph (1975)
a Calculated from values provided.b Fecundity (x– number of eggs per mg female engorged mass).c Diapausing female.d Non-diapausing female.
520

because of desiccation of egg batches or fungal growth which made it impossible todetermine the weight of such egg batches.
Between 12.4 and 17.7 eggs per mg body mass are produced for some Ixodesspecies, such as I. rubicundus, I. ovatus and Ixodes trianguliceps (fecundity; Tables2 and 5). This is evidently higher than other species such as I. hexagonus (5.2) andI. ricinus (7.7). The values recorded for the last two species were closer to the valuesrecorded for some metastriate ticks (Table 5). It is evident that I. rubicundusproduces a large number of relatively small eggs. The mean egg mass (0.03 mg) forI. rubicundus is considerably less than the 0.07 mg recorded for I. hexagonus(Toutoungi et al., 1995). Although the engorgement mass (maximum of 0.324;Petney et al., 1989) of I. rubicundus is less compared to I. ricinus (maximum of0.539 g; Honzakova et al., 1975) the mean number of eggs laid is considerably more(Table 5), further illustrating this point. The relatively large number of eggs producedmay be an adaptation of I. rubicundus, which is frequently exposed to adverseenvironmental conditions, to maximize the number of progeny reaching the adultstage.
Acknowledgements
This study was funded by the Foundation for Research Development and theUniversity of the Orange Free State.
References
Aubert, M.F.A. 1981. Breeding of the tick Ixodes (Pholeoixodes) rugicollis (Acari: Ixodidae) underlaboratory conditions. J. Med. Entomol. 18: 324–327.
Chilton, N.B. and Bull, C.M. 1993. Oviposition by two Australian species of reptile tick. Acarology 34:115–121.
Davey, R.B. 1993. Stagewise mortality, ovipositional biology and egg viability of Boophilus annulatus(Acari: Ixodidae) on Boselaphus tragocamelus (Artiodactyla: Bovidae). J. Med. Entomol. 30:997–1002.
Drummond, R.O. and Whetstone, T.M. 1970. Oviposition of the Gulf Coast tick. J. Econ. Entomol. 63:1547–1551.
Fourie, L.J. and Horak, I.G. 1994. The life cycle of Ixodes rubicundus (Acari: Ixodidae) and itsadaptation to a hot, dry environment. Exp. Appl. Acarol. 18: 23–35.
Fourie, L.J. and Vrahimis, S. 1989. Tick-induced paralysis and mortality of gemsbok. S. Afr. J. Wildl.Res. 19: 118–121.
Fourie, L.J., Kok, O.B. and Van Zyl, J.M. 1991. Spatial distribution of the Karoo paralysis tick Ixodesrubicundus (Acari: Ixodidae) within a false upper Karoo veld type. Exp. Appl. Acarol. 11: 37–49.
Fujimoto, K. 1989. Ecological studies on ixodid ticks. 6. The effects of temperature on the oviposition,development and survival of Ixodes ovatus Neumann (Acarina: Ixodidae). Jpn J. Sanit. Zool. 40:187–193.
Fujimoto, K. 1992. Comparative observations on oviposition and development of two ixodid ticks,Ixodes persulcatus Schultze and Ixodes nipponensis Kitaoka and Saito, under different temperatures.Jpn J. Sanit. Zool. 43: 105–112.
521

Guglielmone, A.A. 1992. The effect of temperature and humidity on development and longevity ofAmblyomma triguttatum triguttatum (Acari: Ixodidae). Bull. Entomol. Res. 82: 203–208.
Hagras, A.E. and Khalil, G.M. 1988. Effect of temperature on Hyalomma (Hyalomma) dromedarii Kock(Acari: Ixodidae). J. Med. Entomol. 25: 354–359.
Honzakova, E., Olejnıcek, J., Cerny, V., Daniel, M. and Dusbabek, F. 1975. Relationship betweennumber of eggs deposited and body weight of engorged Ixodes ricinus female. Folia Parasitol. 22:37–43.
Linthicum, K.J., Logan, T.M., Kondig, J.P., Gordon, S.W. and Bailey, C.L. 1991. Laboratory biology ofHyalomma truncatum (Acari: Ixodidae). J. Med. Entomol. 28: 280–283.
MacLeod, J. 1935. Ixodes ricinus in relation to its physical environment. III Climate and reproduction.Parasitology 27: 489–500.
Norval, R.A.I. 1977. Studies on the ecology of the tick Amblyomma hebraeum Koch in the eastern Capeprovince of South Africa. II. Survival and development. J. Parasitol. 63: 740–747.
Petney, T.N., Fourie, L.J. and De Jager, C. 1989. The effect of the pattern of attachment weights offemale Ixodes rubicundus on Karoo paralysis in a naturally infested flock of sheep. Vet. Parasitol. 33:177–186.
Randolph, S.E. 1975. Seasonal dynamics of a host-parasite system: Ixodes trianguliceps (Acarina:Ixodidae) and its small mammal hosts. J. Animal Ecol. 44: 425–429.
Smith, C.N. 1944. Biology of Ixodes muris Bishopp and Smith (Ixodidae). Ann. Entomol. Soc. Am. 37:221–234.
Smith, M.W. 1972. The life history of Ixodes canisuga (Johnston, 1849) under laboratory conditions.Ann. Trop. Med. Parasitol. 66: 281–286.
Spickett, A.M. and Heyne, H. 1988. A survey of Karoo paralysis in South Africa. Onderstepoort J. Vet.Res. 55: 89–92.
Stampa, S. 1959. Tick paralysis in the Karoo areas of South Africa. Onderstepoort J. Vet. Res. 28:169–227.
Stampa, S. and Du Toit, R. 1958. Paralysis of stock due to the Karoo paralysis tick (Ixodes rubicundusNeumann). S. Afr. J. Sci. 54: 241–246.
Toutoungi, L.N., Gern, L. and Aeschlimann, A. 1995. Biology of Ixodes (Pholeoixodes) hexagonus underlaboratory conditions: part II. Effect of mating on feeding and fecundity of females. Exp. Appl. Acarol.19: 233–245.
Venter, J.M., Mocke, C. and De Jager, J. M. 1986. Climate. In The Karoo Biome: A preliminarysynthesis. Part I – physical environment, R.M. Cowling, P.W. Roux and A.J.H. Pieterse (eds), pp.39–52. South African National Scientific Programmes Report No. 124.
Winston, P.W. and Bates, D.H. 1960. Saturated solutions for the control of humidity in biologicalresearch. Ecology 41: 232–237.
Yano, Y., Shiraishi, S. and Uchida, T.A. 1987. Effects of temperature on development and growth in thetick Haemaphysalis longicornis. Exp. Appl. Acarol. 3: 73–78.
522