Embed Size (px)
Citation preview

Louisiana State UniversityLSU Digital Commons
LSU Historical Dissertations and Theses Graduate School
1985
Biology, Economic Injury, and Plant ResistanceStudies With the Fall Armyworm, SpodopteraFrugiperda ( J. E. Smith), on Rice, Oryza Sativa L.(Screening, Puerto Rico, Louisiana).Alberto Pantoja-lopezLouisiana State University and Agricultural & Mechanical College
Follow this and additional works at: https://digitalcommons.lsu.edu/gradschool_disstheses
This Dissertation is brought to you for free and open access by the Graduate School at LSU Digital Commons. It has been accepted for inclusion inLSU Historical Dissertations and Theses by an authorized administrator of LSU Digital Commons. For more information, please [email protected].
Recommended CitationPantoja-lopez, Alberto, "Biology, Economic Injury, and Plant Resistance Studies With the Fall Armyworm, Spodoptera Frugiperda ( J.E. Smith), on Rice, Oryza Sativa L. (Screening, Puerto Rico, Louisiana)." (1985). LSU Historical Dissertations and Theses. 4147.https://digitalcommons.lsu.edu/gradschool_disstheses/4147

INFORMATION TO USERS
This reproduction was made from a copy of a m anuscript sent to us for publication and microfilming. While the m ost advanced technology has been used to photograph and reproduce this m anuscript, the quality of the reproduction is heavily dependent upon the quality of the material submitted. Pages in any m anuscript may have indistinct print. In all cases the best available copy has been filmed.
The following explanation of techniques is provided to help clarify notations which may appear on this reproduction.
1. Manuscripts may not always be complete. When it is not possible to obtain missing pages, a note appears to indicate this.
2. When copyrighted materials are removed from the m anuscript, a note appears to indicate this.
3. Oversize materials (maps, drawings, and charts) are photographed by sectioning the original, beginning at the upper left hand com er and continuing from left to right in equal sections with small overlaps. Each oversize page is also filmed as one exposure and is available, for an additional charge, as a standard 35mm slide or in black and white paper format.*
4. Most photographs reproduce acceptably on positive microfilm or microfiche b u t lack clarity on xerographic copies made from the microfilm. For an additional charge, all photographs are available in black and white standard 35mm slide format.*
*For more information about black and white slides or enlarged paper reproductions, please contact the Dissertations Customer Services Department.
IMveraityMkronliris
Internationa]


8610657
Pantoja-L opez, A lberto
BIOLOGY, ECONOMIC INJURY, AND PLANT RESISTANCE STUDIES WITH THE FALL ARMYWORM, SPODOPTERA FRUGIPERDA (J. E. SMITH), ON RICE, ORYZA SATIVA L.
The Louisiana State University and Agricultural and Mechanical Col. Ph.D. 1985
UniversityMicrofilms
International 300 N. Zeeb Road, Ann Arbor, Ml 48106


PLEASE NOTE:
In all cases this material has been filmed in the best possible way from the available copy. Problems encountered with this docum ent have been identified here with a check mark V .
1. Glossy photographs or p ag es_____
2. Colored illustrations, paper or print______
3 . Photographs with dark background_____
4 . Illustrations are poor copy______
5. Pages with black marks, not original copy______
6. Print shows through as there is text on both sides of p a g e _______
7. Indistinct, broken or small print on several pages. /
8. Print exceeds margin requirem ents______
9. Tightly bound copy with print lost in sp ine_______
10. Computer printout pages with indistinct print______
11. Page(s)____________ lacking when material received, and not available from school orauthor.
12. Page(s)____________ seem to be missing in numbering only as text follows.
13. Two pages num bered . Text follows.
14. Curling and wrinkled p ag es_______
15. Dissertation contains pages with print at a slant, filmed a s received_________
16. Other_________________________________________________________________________
UniversityMicrofilms
International


BIOLOGY, ECONOMIC INJURY, AND PLANT RESISTANCE STUDIES WITH
THE FALL ARMYWORM, SPODOPTERA FRUG1PERDA (J. E. SMITH),
ON RICE, ORYZA SATIVA L.
A Dissertation
Submitted to the Graduate Faculty of the Louisiana State University
Agricultural and Mechanical College in partial fulfillment of the requirements for the degree of
DOCTOR OF PHILOSOPHY
in
The Department of Entomology
by
Alberto Pantoja-Lopez B.S., University of Puerto Rico-Mayaguez, 1978
M.S., Louisiana State University, 1982 December, 1985

ACKNOWLEDGEMENTS
The author wishes to express his appreciation to his major professor, Dr. C. M. Smith, Associate Professor of Entomology, for his
patience, guidance, and invaluable suggestions during the completion of
this work and in the preparation of this manuscript.
Gratitude is extended to Dr. J. F. Robinson, USDA-ARS Rice
Entomologist, Louisiana State University Rice Research Station, Crowley,
LA, for his counsel in planning a course of study and assistance in
conducting the field work.
The writer is also indebted to the following members of his advisory committee, for their constructive criticism and suggestions: Dr. J. B. Graves, Dr. E. A. Heinrichs, Dr. S. S. Quisenberry, Entomology
Department; Dr. K. S. McKenzie, Louisiana State University Rice Research
Station; Dr. C. M. Rush, Department of Plant Pathology and Crop
Physiology; and Dr. W. G. Warren, Department of Experimental Statistics.
Thanks are extended to G. B. Trahan, R. W. Webster, Louisiana State
University Rice Research Station; C. Cook, J. Negron, M. Riley, K.Samoil, Entomology Department; and P. Caballero, Chemistry Department,
for technical assistance, friendship, and encouragement during the course of this study.
The writer wishes to acknowledge the help of Mrs. B. Gustavson,
Department of Experimental Statistics, for assistance in the
statistical analyses and interpretation of the data.
The author also wishes to acknowledge the administration of the
Louisiana State University Rice Research Station for providing space,
personnel, and equipment for the field experiments.
ii

Appreciation is also extended to all the student workers, and
secretarial personel in the Entomology Department, who have given of their time and experience to assist in the completion of this research
study and in the preparation of this manuscript.
Finally the author wishes to express his special appreciation to his wife, Alicia and to his parents, Jose Pantoja and Altagracia Lopez
for their enthusiasm, assistance, encouragement, and understanding
throughout this study.
iii

TABLE OF CONTENTS
Page
ACKNOWLEDGEMENT......................................................ii
TABLE OF CONTENTS....................................................iv
LIST OF TABLES.......................................... v
LIST OF FIGURES........ ix
ABSTRACT.............................................................. x
INTRODUCTION.......................................................... 1
LITERATURE REVIEW. ................................... 3CHAPTER I EVALUATION OF RICE GERMPLASM FOR RESISTANCE
TO THE FALL ARMYWORM, SPODOPTERA FRUGIPERDA (J. E. SMITH), (LEPIDOPTERA:NOCTUIDAE)................... 9
CHAPTER II RICE YIELDS AS AFFECTED BY THE FALL ARMYWORM,SPODOPTERA FRUGIPERDA, (J. E. SMITH),(LEPIDOPTERA: NOCTUIDAE)................................ 28
CHAPTER III NATURAL CONTROL AGENTS AFFECTING SPODOPTERAFRUGIPERDA (LEPIDOPTERA:NOCTUIDAE), INFESTINGRICE IN PUERTO RICO................................... 49
CHAPTER IV DEVELOPMENT OF FALL ARMYWORM, SPODOPTERA FRUGIPERDA (J. E. SMITH), (LEPIDOPTERA:NOCTUIDAE) FROM LOUISIANA AND PUERTO RICO:A BIOLOGICAL COMPARISON................................ 56
SUMMARY AND CONCLUSIONS..............................................77
LITERATURE CITED.....................................................80
APPENDIX A ........................................................... 85
APPENDIX B........................................................... 90
APPENDIX C.......................................................... 105
VITA................................................................ 116
iv

LIST OF TABLES
Table Page
CHAPTER I
1. Defoliation and percent of larval population on selected rice cultivars by Spodoptera frugiperda larvae in choiceand no-choice tests. Louisiana, 1983-1985......................... 17
2. Defoliation of selected United States rice cultivars by Spodoptera frugiperda neonate larvae.Baton Rouge, LA 1985............................................18
3. Defoliation of selected Oryza species by Spodopterafrugiperda neonate larvae. Baton Rouge, LA 1985................ 20
CHAPTER II
1. Defoliation, plant stand, panicle density, kernel weight, and yield of rice as affected by Spodoptera frugiperdalarval feeding. Crowley, LA 1983.............................. 35
2. Biomass index, yield, and yield components of rice as affected by Spodoptera frugiperda larval feeding.Crowley, LA 1984................................................37
3. Regression analysis of Spodoptera frugiperda larval density (larvae/m2) on percent yield reduction.Crowley, LA 1983-1984...........................................39
CHAPTER IV
1. Weight of Spodoptera frugiperda from Louisiana (LA) and Puerto Rico (PR) reared on artificial diet. Baton Rouge,LA 1984......................................................... 63
2. Developmental time (days) of Spodoptera frugiperda fromLouisiana (LA) and Puerto Rico (PR) when reared on artificialdiet. Baton Rouge, LA 1984.................................... 64
3. Development of Spodoptera frugiperda from Louisiana (LA)and Puerto Rico (PR) when reared on 'Mars' rice foliage.Baton Rouge, LA 1984............................................65
4. Developmental time (days) and number of instars of Spodoptera frugiperda larvae from Louisiana (LA) and Puerto Rico (PR) when reared on 'Mars' rice foliage.Baton Rouge, LA 1984............................................67
v

TableLIST OF TABLES (CONTINUED)
PageAPPENDIX A
1. Analysis of variance for defoliation rate on selected rice cultivars by Spodoptera frugiperda larvae in a nochoice test. Crowley, LA 1985.................................. 86
2. Analysis of variance for percent larval population on selected rice cultivars by Spodoptera frugiperda larvaein a choice test. Baton Rouge, LA 1985........................ 87
3. Analysis of variance for defoliation rate of selected United States rice cultivars by Spodoptera frugiperdaneonate larvae. Baton Rouge, LA 1985........................... 88
4. Analysis of variance for defoliation rate of selected Oryza species by Spodoptera frugiperda neonate larvae.Baton Rouge, LA 1985.............................................89
APPENDIX B
1. Analysis of variance for defoliation percentage by Spodoptera frugiperda third instar larvae on ’Mars’rice at varying density levels. Baton Rouge, LA 1983...........91
2. Analysis of variance for 'Mars' rice plant density asaffected by Spodoptera frugiperda third instar larvaeat varying density levels. Baton Rouge, LA 1983................92
3. Analysis of variance for 'Mars’ rice panicle densityas affected by Spodoptera frugiperda third instarlarvae at varying density levels. Baton Rouge, LA1983............................................................. 93
4. Analysis of variance for 'Mars' rice gross yield as affected by Spodoptera frugiperda third instar larvaeat varying density levels. Baton Rouge, LA 1983................94
5. Analysis of variance for weight of 500 kernels of 'Mars' rice affected by Spodoptera frugiperda thirdinstar larvae at varing density levels. Baton Rouge,LA 1983.......................................................... 95
6. Analysis of variance for panicle weight of 'Mars' riceaffected by Spodoptera frugiperda third instar larvaeat varying density levels. Baton Rouge, LA 1983................96
vi

LIST OF TABLES (CONTINUED)Table Page
7. Analysis of variance for percent fertile kernels of 'Mars' rice affected by Spodoptera frugiperda third instar larvae at varying density levels. Baton Rouge,LA 1983.......................................................... 97
8. Analysis of variance for biomass index of 'Mars' rice affected by Spodoptera frugiperda fourth instar larvaeat varying density levels. Baton Rouge, LA 1984.... 98
9. Analysis of variance for panicle density of 'Mars' riceaffected by Spodoptera frugiperda fourth instar larvaeat varying density levels. Baton Rouge, LA 1984............... 99
10. Analysis of variance for panicle weight of'Mars' rice affected by Spodoptera frugiperda fourth instar larvae at varying density levels. Baton Rouge,LA 1984......................................................... 100
11. Analysis of variance for mature plant height of 'Mars' rice affected by Spodoptera frugiperda fourth instar larvae at varying density levels. Baton Rouge,LA 1984........................................................ 101
12. Analysis of variance for weight of 1000 kernels of 'Mars' rice affected by Spodoptera frugiperda fourthinstar larvae at varying density levels. Baton Rouge,LA 1984......................................................... 102
13. Analysis of variance for adjusted yield of 'Mars' riceaffected by Spodoptera frugiperda fourth instar larvaeat varying density levels. Baton Rouge, LA 1984................103
14. Analysis of variance for percent filled kernels of 'Mars' rice affected by Spodoptera frugiperda fourthinstar larvae at varying density levels. Baton Rouge,LA 1984.......................................................... 104
APPENDIX C
1. Analysis of variance for consumption index by Spodoptera frugiperda third instar larvae from Louisiana and PuertoRico fed 'Mars' rice foliage. Baton Rouge, LA 1984.............106
2. Analysis of variance for growth ratio of Spodopterafrugiperda third instar larvae from Louisiana and Puerto Rico fed 'Mars' rice foliage. Baton Rouge, LA1984............................................................. 107
vii

LIST OF TABLES (CONTINUED)Table Page
3. Analysis of variance for approximate digestibility bySpodoptera frugiperda third instar larvae from Louisiana and Puerto Rico fed 'Mars' rice foliage. Baton Rouge,LA 1984.......................................................... 108
4. Analysis of variance for efficiency of conversion ofingested food by Spodoptera frugiperda third instar larvae from Louisiana and Puerto Rico fed 'Mars' ricefoliage. Baton Rouge, LA 1984................................... 109
5. Analysis of variance for efficiency of conversion ofdigested food by Spodoptera frugiperda third instarlarvae from Louisiana and Puerto Rico fed 'Mars' ricefoliage. Baton Rouge, LA 1984................................... 110
6. Analysis of variance for preovipositlonal days ofSpodoptera frugiperda adults from Louisiana and PuertoRico. Baton Rouge, LA 1984.....................................Ill
7. Analysis of variance for oviposition period ofSpodoptera frugiperda adults from Louisiana and PuertoRico. Baton Rouge, LA 1984.................................... 112
8. Analysis of variance for adult longevity of Spodopterafrugiperda from Louisiana and Puerto Rico. Baton Rouge,LA 1984.......................................................... 113
9. Analysis of variance for number of egg masses depositedby Spodoptera frugiperda adults from Louisiana andPuerto Rico. Baton Rouge, LA 1984..........................114
10. Analysis of variance for head capsule width ofSpodoptera frugiperda neonate larvae from Louisiana andPuerto Rico. Baton Rouge, LA 1984..........................115
viii

LIST OF FIGURES
CHAPTER II Page
Figure 1. Regression of Spodoptera frugiperda larval density on percent rice yield reduction. Crowley, LA 1983-1984..................................................40
ix

ABSTRACT
A method for screening rice, Oryza sativa L., germplasm for
resistance to the fall armyworm, Spodoptera frugiperda (J. E. Smith),
was developed. Infesting rice seedlings grown in the greenhouse proved
to be an efficient method to identify sources of resistance. In the
evaluation of over 5,000 rice accessions, moderate resistance to S.
frugiperda larval defoliation was detected in plant introductions
160842, 346830, 346833, 346840, and 346853. Moderate levels of
resistance were also detected in the United States cultivars 'Honduras',
'Newrex', and 'Chinese'; and in 0. glaberrima Steud accessions 101800,
102554, and 369453.Effects of j3. frugiperda feeding on rice were studied in the field
during 1983 and 1984. Increased infestation levels resulted in
increased defoliation, reduced plant and panicle density, and reduced
rice yields. Yield component studies indicated that lower yields on infested plots were the result of reduced plant and panicle densities
and reduced panicle size. Kernel weight and filling were not affected.
Yield reductions were found to be linearly related to S. frugiperda
larval density. The linear model E(y) = b x^ accounted for a large
proportion of the variance of yield during both years.Twenty-eight parasitoid adults, larvae, and puparia were recovered
from j>. frugiperda larvae collected from rice fields in Puerto Rico.
Eight were identified as Lespesia sp. (Diptera:Tachinidae), and one as
Archytas mormoratus (Townsend), (Diptera:Tachinidae). The fungus Nomuraea rileyi (Farlow) Sampson killed eight larvae and three were
infected by an unidentified microsporidium.
x

Development, longevity, and reproduction of S_. frugiperda from
Louisiana (LA) and Puerto Rico (PR) were evaluated in larvae reared on
artificial diet and rice foliage. At egg hatch, larvae of PR origin
were smaller than LA larvae. Differences in neonate larval size were
also evident at other stages of development, but did not extend to adult
longevity or reproductive parameters. Larvae, pupae and adults of the
LA strain were heavier and completed development in a shorter period of
time than the PR strain. Differences are probably under genetic control
and may indicate some degree of reproductive isolation.
xi

INTRODUCTIONRice, Oryza sativa L., is attacked by more than 100 insect species.
Among these, armyworms represent a sporadic but potentially important
pest to rice production in the United States. In other countries,
armyworms are among the most serious pests limiting rice production. Of the armyworm species occurring in the New World, the fall armyworm,
Spodoptera frugiperda (J. E. Smith), is the most important.
Since colonial times S. frugiperda has been recognized as an
injurious pest to a wide range of crops, especially the Graminae. The
larvae are particularly injurious to nursery beds and tender young plants, although severe damage can be done to mature plants as well.
Spodoptera frugiperda may suddenly appear in large populations of
foliage feeding larvae and reinvades most of the continental United States and southern Canada after overwintering in the lower tips of
Texas and Florida (Luginbill 1928). During mild winters the insect is
capable of overwintering in Louisiana (Oliver and Chapin 1981). The
development of damaging insect populations is favored by cool, wet
spring conditions followed by warm, humid weather (Sparks 1979). An
outbreak of S_. frugiperda on rice required multiple insecticide
applications to control dense populations covering more than 60% of the
total Texas rice area (Bowling 1978). Louisiana’s geographic location
and climatic conditions are favorable for the development of injurious
_S. frugiperda populations in rice fields.
Little research has been conducted to document j>. frugiperda damage
to rice plants. Chandler et al. (1977), Navas (1974), and Rice et al.
(1982a) documented yield reductions due to armyworm damage on rice;
1

however, most of the data available are based on artificial defoliation
rather than on actual insect feeding.
Soejitno and van Vreden (1976) reported varietal resistance
against the rice swarming caterpillar, Spodoptera mauritia (Boisduval)
in Indonesia. Recently, a greenhouse technique was developed in the
Philippines (Wilde and Apostol 1983) to screen rice lines for resistance
against the rice earcutting caterpillar, Mythimna separata (Walker), and
S_. mauritia. Host-plant resistance is currently used for control of _S.
frugiperda on Bermuda grass (Combs and Valerio 1980; Leuck et al. 1968b;
Leuck and Skinner 1970; Pencoe and Martin 1982; Quisenberry and Wilson
1985; Wiseman et al. 1982), pearl millet (Leuck et al. 1968a; Leuck 1970, 1972), sorghum (McMillan and Starks 1967; Wiseman and Gourley
1982), corn (Brett and Bastida 1963; Manuwoto and Scriber 1982; Ng et
al. 1985; Williams et al. 1983, 1985; Wiseman et al. 1966), and peanuts
(Leuck and Hammons 1974; Leuck and Skinner 1971). Control of _S.
frugiperda larvae in rice is generally accomplished by insecticidal
applications while little attention has been given to the use of host-plant resistance against as a control measure.
The objectives of this study were to: 1. develop a reliable and
efficient greenhouse screening technique for determining J3. frugiperda
resistance in rice and to identify sources of resistance, 2. study the
effects of leaf feeding and larval density of J5. frugiperda on rice
yield, 3. identify natural control agents affecting S_. frugiperda larvae
infesting rice, and 4. compare the development of j5. frugiperda
collected in Puerto Rico and Louisiana.

LITERATURE REVIEW
Economic importance.
Very little information is available on armyworm damage to rice.
The 1979 and 1980 Spodoptera frugiperda Symposia (see Fla. Entomol. vols. 62 and 63) provided a comprehensive review in selected topics on
the biology, rearing, population dynamics, host-plant resistance, and
biological, chemical, and cultural control. Over 175 papers were cited
regarding armyworm damage to various crops but very little reference was made to rice.
Armyworm infestations are considered a limitating factor in rice
production in various countries. Jung-Tsung (1982) reported eight armyworm species from two genera, Mythimna and Spodoptera, causing
significant or serious damage to rice fields in various areas of China.
Pathak (1977) listed 13 armyworm species from three genera, Spodoptera,
Prodenia, and Pseudaletia, as sporadic but serious pests of rice in
various countries. Rice production is also limited by armyworms in
Bangladesh, Pakistan, Sri Lanka, and Thailand (Litsinger 1979; Pathak
and Dhaliwal 1981). In Panama, _S. frugiperda is among the three most
important insect pests on rice (Navas 1966). S_. frugiperda is also animportant economic rice pest in Colombia (Gallego 1967) and Brazil
(Machado 1978). Chandler et al. (1977) listed S_. frugiperda as the main insect affecting rice production in Puerto Rico. In the United States,
S_. frugiperda and Pseudaletia unipuncta are considered as sporadic but important pests on rice (Bowling 1978; Rice et al. 1982a, 1982b).
Spodoptera frugiperda damage to rice in the United States was reported as early as 1845 (Luginbill 1928). During 1899, jS. frugiperda produced

severe damage to rice fields in North Carolina (Chittenden 1900).
In India, two previously unimportant armyworm species became
serious pests after the introduction of semidwarf rice cultivars
(Banerjee and Chaterjee 1982; Papel et al. 1981; Kulshreshtha et al. 1970). Cong Thuat (1982) also reported changes in the incidence of
armyworms on rice in Vietnam after the introduction of new rice
cultivars. Semidwarf rice cultivars tend to have denser crop stands
with upright leaves (Pathak and Dhaliwal 1981). Fall armyworm
ovipositional preferences for plants with dense foliage has been
documented on eight crop plant species, Bermuda grass, Cynodon dactylon
(L.); corn, Zea mays L.; cotton, Gossypium hirsutum L.; sorghum, Sorghum bicolor (L.); soybean Glycine max (L.); rye, Secale cereale L.;
ryegrass, Lolium perenne L.; and wheat, Triticum aestivum (L.) (Pitre et al. 1983). These reports may be an indication that host selection by
armyworms is influenced by plant morphology and growth characteristics.
Rice et al. (1982b) found higher unipuncta larval concentrations on
areas of high rice plant density and low light intensity, possibly due to the semidwarf rice varieties providing a more adequate microclimate
for larval development. Semidwarfism is an agronomic character being
incorporated into adapted California rice cultivars (McKenzie et al.
1980) and selected for in the Louisiana rice breeding program (Mcllrath et al. 1978).
Damage to rice by JP. unipuncta was described by Rice et al.
(1982a). Young larvae skeletonize leaves, mature larvae remove the
entire leaf surface, but the mid vein is not affected by either age
larvae. The amount of feeding damage is dependent on plant density,
temperature, and larval density. In Asia, armyworms have been reported

feeding on the palea and lemma of the grain and anthers of flowers
(Papel et al. 1981) as well as leaves and cutting the spikelets
(Jung-Tsung 1982). Bowling (1978) indicated that JS. frugiperda damage
to flooded rice was restricted to the portion of the plant above water,
but when the seedling was too small to flood, entire plants were consumed.
Little is known about the relationship of jS. frugiperda population
density to yield reductions on rice. Rice et al. (1982a) reported grain
yield reductions ranging from -2.8 to 50.2% due to 25 to 30% defoliation
by P_. unipuncta larvae. A significant reduction in the number of seed
per plant was also observed on defoliated plants, compared to
non-damaged plants. Pophali et al. (1980) reported yield losses ranging
from 3.6 to 22% by populations of 12 to 14 M. separata larvae per m2.
An average population of 1.2 to 5.4 JP. unipuncta larvae per hill
produced 5 to 95% leaf damage (Dar 1979). Young (1979a) indicated that
JL* frugiperda populations can reach 202,500 larvae per ha, but gave no indication of the host-plant or crop on which this infestation was
observed. Approximately 60 to 75% of the total rice area of Texas was
reported as infested in 1973; however, insect population density was not
stated nor yield measured (Bowling 1978). Navas (1974) reported good
yields from upland rice fields in Panama infested with _S. frugiperda
populations of 420 and 251 larvae per m2, but no data were presented to substantiate this conclusion. Thus, the point at which £1 frugiperda
population density reaches the economic damage level on rice has not yet
been determined.
The few references available on armyworm damage to rice are based mainly on artificial defoliation techniques. Bowling (1978) and Navas

6(1966, 1967) reported reductions on rice yields by simulated insect
damage, but concluded that rice plants are capable of recovering rapidly
from extensive damage, particularly if the damage occurs in the early
vegetative stage of growth. Plant response to defoliation by chewing
insects may be different to that produced by artificial defoliation.
Capinera and Roltsch (1980) reported a lower rate of regrowth on wheat,
1L aestivum, affected by grasshopper, Melanoplus sanguinipes (F.), feeding when compared to equal amounts of manual clipping. Data by Rice
et al. (1982a) indicate higher rice yield reductions by _P. unipuncta
defoliation than by yield reductions from an equal percentage of
artificial defoliation, indicating that data based on artificial
defoliation may not be a reliable indication of actual insect damage.
Biological control.Various insects have been reported parasitizing jS. frugiperda
larvae and eggs. Ashley (1979) and Lewis and Norlund (1980) listed 53
species of parasites reared from SL frugiperda eggs and larvae. Only 18
of these are common to the continental United States, while 21 are
present in South America and Central America, including Mexico. Ashley
(1982, 1983), and Ashley et al. (1982, 1983) studied the impact on
frugiperda population of eight native and one imported parasite in south
Florida. These included: Apanteles marginiventris, Campoletis grioti,
Chelonus insularis, Meteorus autographae, Ophion spp., Rogus laphygmae,
Ternelucha spp. and Eiphosoma vitticole Cresson (imported). Although
63% of the first four larval instars were destroyed by parasites, they
concluded that j>. frugiperda has the reproductive potential to increase
its population beyond regulation by native parasites. No EL vitticole

were recovered from the experimental plots» Twenty-one species of
armyworm parasites and predators from five genera are known to occur in
China (Jung-Tsung 1982), yet the efficiency of these parasites in
control of natural armyworm populations has not been determined. Lewis
and Nordlund (1980) stressed the need for research to evaluate the
potential of using natural enemies to control this pest.
Reports by Gardner and Fuxa (1980) indicate that J3. frugiperda is
susceptible to at least 16 species of entomogenous pathogens (viruses,
bacteria, protozoa, fungi, and nematodes). A detailed list indicating
pathogen, geographic location where reported, and host plant association
included no pathogens found in association with rice plants. Hamm and Hare (1982) reported a renewed interest in the use of entomopathogens
for armyworm control; however, no commercial formulation of any
microbial agent is currently available for S. frugiperda control in the
United States.
Chemical control.
Spodoptera frugiperda has demonstrated resistance or tolerance
against to pesticides commonly used on its control (Andrews 1980; Combs
and Chambers 1979; Young 1979b) and multiple insecticide applications
are often necessary to obtain adequate control on rice (Bowling 1978; Chandler et al. 1977; Young 1979b). Harrell et al. (1977) also reported
S. frugiperda tolerance to synthetic pyrethroids.
Cultural control.
Various cultural and mechanical management techniques have been
reported for armyworm control. Small farmers in Central America hand

pick and destroy larvae or apply mud to whorls to inoculate larvae with
soil inhabiting pathogens (Andrews 1980). Damage by armyworms may be
avoided by early planting (Andrews 1980; Mitchell 1978), and
fertilization to stimulate plant regrowth or affect insect response to
plants (Leuck 1972; Lynch et al. 1980). Altieri (1980) suggests
manipulating plant diversity within crops systems to reduce S_. frugiperda incidence.
Host-plant resistance.
No references on genetic resistance to frugiperda in rice
exists, but a few references are available regarding resistance of rice
to other armyworm species. Dar (1979) detected differences in percent
damaged panicles and number of JP. unipuncta larvae among eight rice
varieties in Pakistan. Resistance to S. mauritia was reported by
Soejitno and Vreden (1976) among 462 Asian lines screened under
greenhouse conditions in Indonesia. Pophali et al. (1980) observed
differences in yield loss among 11 rice cultivars due to M. separata.
Wilde and Apostol (1983) developed a greenhouse technique to screen rice
varieties for M. separata and j>. mauritia. The technique consisted of
infesting plants with laboratory-reared first instar larvae dispensed
with a Davis inoculator (Davis and Oswalt 1969).

CHAPTER I
EVALUATION OF RICE GERMPLASM FOR RESISTANCE TO THE FALL ARMYWORM,
SPODOTERA FRUGIPERDA (J. E. SMITH), (LEPIDOPTERA:NOCTUIDAE)
This chapter is written in the style of
Crop Science
9

ABSTRACT
A greenhouse method for screening rice, Oryza sativa L., germplasm
for resistance to the fall armyworm, Spodoptera frugiperda (J. E.
Smith), was developed. In the evaluation of over 5,000 rice cultivars,
moderate resistance to J3. frugiperda larval defoliation was detected in
plant introductions 160842, 346830, 346833, 346840, and 346853.
Moderate levels of resistance were also detected in the United States
cultivars 'Honduras1, 'Newrex', and 'Chinese', and in JD. glaberrima
Steud accessions 101800, 102554, and 369453. Differences in the
performance of plant introduction 160842 in choice and no-choice experiments indicate that non-preference resxstance exists in this
cultivar.
10

INTRODUCTION
Rice, Oryza sativa L., is attacked by more than 100 insect species.
Among these, the fall armyworm, Spodoptera frugiperda (J. E. Smith),
represents a sporadic, but potentially important threat to rice
production in the United States (Bowling, 1978). In Latin American
countries; however, _S. frugiperda is among the most serious pests
limiting rice production. Chandler et al. (1977) listed S. frugiperda
as the main insect affecting rice production in Puerto Rico. In Panama,
£>. frugiperda is among the three most important insect pests of rice
(Navas, 1966). Spodoptera frugiperda is also an economically important
rice pest in Colombia (Gallego, 1967) and Brazil (Machado, 1978).Control of _S. frugiperda in rice is generally accomplished by
insecticidal applications, and little attention has been given to the
use of host-plant resistance. Wiseman and Davis (1979) and Davis (1980)
reviewed the history and progress of JL, frugiperda host-plant resistance
programs on various crops, but no papers were cited regarding resistance
in rice.
Limited research has been conducted regarding rice resistance
to other armyworm species. Dar (1979) detected differences in percent
damaged panicles and number of Pseudaletia unipuncta (Haworth) larvae
among eight rice cultivars in Pakistan. Resistance to Spodoptera mauritia (Boisduval) was reported by Soejitno and van Vreden (1976)
among 462 Asian cultivars screened under greenhouse conditions in
Indonesia. Pophali et al. (1980) observed differences in yield loss due
to Mythimna separata (Walker) among 11 rice cultivars. Wilde and
Apostol (1983) detected various degrees of resistance against M.
11

12
separata and S. mauritia in rice cultivars evaluated in the Philippines.
We report here the development of a greenhouse screening technique for
determining S. frugiperda resistance in rice and the identification of several sources of resistance.
MATERIALS AND METHODSInsects. The insects used in the experiments were obtained from a
laboratory (26 ± 0.5°C, 14:10 photoperiod, 50 ± 10% RH) colony that has
been under culture for 50 generations at the Louisiana State University
Entomology Department, Baton Rouge, LA. Wild adults and larvae were
collected and introduced biannually into the laboratory colony. Adults
were collected from a light trap at the Louisiana State University Rice
Research Station (LSURRS), Crowley, LA.; larvae were field collected
from Bermuda grass, Cynodon dactylon (L.); corn, Zea mays L.; and
sorghum, Sorghum bicolor (L.). Larvae were reared on a pinto bean diet
(Bowling, 1967) following the procedure described by Perkins (1979).
Egg masses were collected daily from rearing containers and allowed to hatch in plastic bags.
Seed and Preliminary Screening. Four media for growing rice plants
in pots under greenhouse conditions were tested. 'Mars' and 'Saturn'
rice were grown in Crowley silt loam soil, peat moss (Premier Brands
Inc., New Rochelle, New Jersey), Jiffy mix (JPA Co. Harvester Road,
Chicago, Illinois) or organic potting soil (Wonder Chemical Co.,
Livingston, Texas) for 30 days in a test replicated four times. The
potting soil produced 30 day-old plants of equal vigor to those grown in
the soil mixture and was selected as the substrate for further experiments. Plants grown in peat moss and Jiffy mix exibited chlorosis

13symptoms after 15 days and many died by day 30.
Rice seed was germinated in a plastic pot ( 5 x 5 x 6 cm) containing
approximately 10 g of potting soil. Due to the large numbers of world
collection accessions to be tested, unreplicated screening tests (two
plants/pot) were initially conducted. Pots (300 - 500/bench) were
arranged side by side on greenhouse benches lined with four mil clear
plastic visqueen to hold water. Conditions were 14:10 photoperiod,
Sylvania Metal Arch 1000 w lamps, 26 ± 10°C, and 70 ± 10% RH. Water was
supplied daily until the 1.0 stage of growth (Yoshida, 1981), and plants
were then flooded (1 cm deep) until termination of the experiment. To
allow comparisons between groups one pot each of the susceptible entries Mars and Saturn were planted for every 35 world collection accessions.
Under the controlled greenhouse conditions described, rice was grown and
evaluated throughout the year ( 2 - 4 tests/year, 300 - 500
entries/test).
Plants were infested at the 3.0 stage of growth (Yoshida, 1981)
with 4 - 6 neonate frugiperda larvae per plant following the
procedure described by Davis and Oswalt (1969). Larvae were allowed to
feed for seven days or until Mars was heavily defoliated (over 70%).
Cultivars with a defoliation rating lower than 70% were selected for
further evaluation in replicated tests.
Rice germplasm (5,000 accessions) was obtained from the USDA
National Small Grain Collection at Beltsville, Maryland. United States
cultivars were obtained from the LSURRS, and twenty-eight accessions of
Oryza species were obtained from the LSURRS and the International Rice
Research Institute (IRRI) at Los Banos, Laguna, Philippines.
No Choice Test. Eleven cultivars selected from the preliminary

14greenhouse screening and two check cultivars were planted by hand in the
field (LSURRS) at the rate of 3.3 g of seed per meter of row and
fertilized with 67-67-67 kg/ha, N-P2 O1.-K.2 O respectively prior to
planting. Additional N (90 kg/ha) was applied just prior to permanent
flood.
Experimental plots (30 cm long) were confined in cylindrical PVC
plastic barriers (38.1 cm diam. x 45.8 cm tall), driven into the soil to
secure them in place. Weeds inside the cylindrical barriers were
removed by hand prior to insect infestation. When plants were in the
3.0 stage of growth, fourth instar larvae (one larva/plant) were placed
on the plants and allowed to feed for four days. A shallow (5 cm) flood was established to prevent larvae from migrating from the test plots.
The barriers were removed at the end of the four day feeding period, a
permanent flood was established and plants were rated for insect
defoliation using a 1-10 scale (based on % leaf area removed) as follows: 1 = 0-10%, 2 = 11-20%, 3 = 21-30%, 4 = 31-40%, 5 = 41-50%, 6 =
51-60%, 7 = 61-70%, 8 = 71-80%, 9 = 81-90%, 10 = 91-100% defoliation.Choice Test. Cultivars selected from the preliminary greenhouse
screening and further tested in the field were germinated in plastic
petri dishes lined with a moistened Whatman #3 filter paper and
transplanted to pots (20 cm tall x 15 cm diam.) containing soil. Plots
were arranged at random on greenhouse benches and flooded (2.5 cm deep).
When plants were in the 4.0 stage of growth (Yoshida, 1981) foliage was
collected, brought to the laboratory and sectioned in 2 cm long pieces.
Two pieces were selected at random and placed on one side of a 100 x 10
mm plastic petri dish lined with a moistened Whatman #3 filter paper.
Two pieces of Mars foliage sectioned in the same way were placed on the

15
opposite side of the dish to serve as a control. Neonate jS. frugiperda
larvae (8 - 15/dish) were released in the center of the dish with a
Davis inoculator (Davis and Oswalt, 1969). Thirteen cultivars were
tested simultaneously using five replications. Dishes were maintained
in total darkness at 26 ± 0.5°C, 50 ± 10% RH. After 24 h, the number of
larvae on test and control cultivar leaves was recorded and the
percentage of the larval population on leaves of each cultivar was
calculated as follows: percentage of larval population = number of
larvae per test cultivar/(number of larvae per test cultivar + number of
larvae per check cultivar) x 100.
United States Cultivars and Oryza Species Tests. Thirty-four
United States cultivars and 28 accessions of Oryza species were planted under greenhouse conditions in two separate tests. Seed was germinated
in the laboratory and transplanted to 5 x 5 x 6 cm plastic pots
containing potting soil. Plants in the 3.0 stage of growth were
infested with a Davis inoculator with 4-6 neonate S_. frugiperda larvae per plant and rated for defoliation when Mars or the most susceptible
cultivar recieved a defoliaion rating greater than 7.0. A defoliation
scale (percentage of the leaf area skeletonized or consumed) was used to
rate entries as described for the no choice test.
All experiments were arranged as a completely randomized design
with four (no choice) five (choice) or six (Oryza species) replications.
Data were subjected to the General Linear Models (GLM) procedure (Freund
and Littell, 1981), and means separated by Duncan's multiple range test
at the 0.05 or 0.01 (United States cultivars test) level of probability.

RESULTS
Preliminary Screening. Eleven (0.22%) promising plant
introductions from the 5,000 accessions evaluated were selected for
further evaluation in the field during the 1984 growing season (Table
1). Seven of the cultivars were from Argentina, one from the
Philippines, and three were from the People's Republic of China.
Choice and no Choice Tests. There were significant differences
among plant introductions (PI) in choice and no-choice tests (Table 1,
Appendix A-Table 1 & 2). Mean field defoliation ratings were
significantly lower for PI 160842 and 346833 than for Mars, the
susceptible check. Pi's 160842, 346833, 346853, 346840, and 346830 had significantly lower field defoliation ratings than the two most
susceptible entries (PI 348782 and 160827) which displayed the highest
field defoliation rate (9.0 rating). Defoliation for Mars and Saturn
was not different from that of the most susceptible entries. Pi's
346840 and 346824 were the least preferred (10.0% and 12.4%) entries in
the choice test, but PI 346824 was heavily defoliated (7.3 rating) under
field conditions.
United States Cultivars Test. An obsolete United States cultivar,
'Honduras', was the least damaged entry (defoliation rating of 3.8), but
this damage was not significantly lower than 59% of the cultivars tested(Table 2, Appendix A-Table 3). The highest defoliation (7.8 rating) was
to 'LA 110'. Honduras, 'Newrex', 'Chinese', and 'Colusa' had
significantly lower defoliation than Mars, the susceptible check.
'Pecos', 'Calrose 76', 'Brazos', 'Bellemont', 'Toro', and LA 110 were
more defoliated than Mars, but the difference was not significant.
16

17
Table 1. Defoliation and percent of larval population on selected rice cultivars by Spodoptera frugiperda larvae in choice and no-choice tests. Louisiana, 1983-1985.
PlantInt.No.
Cultivarname
Countryof
OriginField % Larval Population
defoliation t on cultivar (No choice test) (Choice test)
160842 2869 P. Rep. China 4.3a * 22.9ab *
346833 H62-15-1 Argentina 4.3a 63.6c
346853 H74-14-1 Argentina 5.0ab 65.0c
346840 H65-22-2 Argentina 5.8ab 10.0a
346830 H61-10-1 Argentina 5.8ab 37,4abc
346839 H64-11-1 Argentina 6.7abc 36.Oabc
160823 Chien P. Rep. China 6.8abc 24.lab
009540 Saturn United States 6,8abc 26.Oab
346855 H75-5-1 Argentina 7.0abc 14.7a
346824 LaPlata Argentina 7.3bc 12.3a
009945 Mars United States 7.8bc §348782 11 Philippines 9.0c 51. lbc
160827 Yu-Hsuan P. Rep. China 9.0c 54.6bc
* Means within a column followed by a letter in common do not differ at the 0.05 level of probability according to Duncan's multiple range test, (no choice test, MSE = 3.08, df = 33; choice test, MSE = 0.61, df = 44)
t 1 = 0-10%, 5 = 41-50%, 10 = 91-100% defoliation.
§ Deleted from statistical analyses, larval population of 74% as compared with Saturn.
11 IV410. 1R568-20-11-3. Yukara x TN-1

'iable 2. D efo lia tion of selecteJ United States r ic e c u ltiv a rs by
Spodoptera frugiperda neonate la rvae . Baton Rouge, LA 1985.
C u ltiva r name Mean d e fo lia tio n f
Honduras 3. 0 3 *
Newrex 4 . Dab
Chinese 4.4abColusa 4 .3abcGulfrose 4.4abcdCalrose 4 .4 abedBelle Patna 4 .4abcdDawn 4 .4 abedCaloro 4 .6abcdLebonnet 4.8abcdefStarbonnet 5.0abcdefMagnolia 5.0abcdefB luebelle 5 .OabcdefLabelie 5 .4abcde:gSaturn 5.4abcdefgNewrose 5.4abcdefgArkrose 5.4abcde£gLacrosse 5.8abcdefgBlue Rose 6.0abcdefgNova 66 6 .OabcdefgRexark 6 .OabcdefgLemont 6.3bcdefgNato 6.4bcdefgD ella 6.5cdefgRexoro 6.6cdefgFortuna 6.6cdefgLeah 6.6cdefgMars 6 .8defgPecos 7.0e£gCalrose 76 7. 3tgBrazos 7. 3£gBellemont 7.5gToro 7.6gLA 110 7.8g
* Means w ith in a column followed by a le t t e r in comnon do not d if fe r at the 0.01 le ve l o f p ro b a b ility according to IXmcan’ s m u ltip le range te s t. (MSE - 1 .43 , df - 122)
t 1 - 0-10Z, 5 - 41-50Z, 10 - 91-1002 d e fo lia tio n or skeleton ization .

19Oryza species test. Seed of only six of the 28 Oryza sp.
accessions sown germinated to provide plants for evaluation (Table 3,
Appendix A-Table 4). The defoliation on the Ch glaberrima accessions
101800 and 102554 and PI 369453 was significantly lower than that on
Mars, the most susceptible of the entries in the test. Defoliation on
(). glaberrima accessions 102445, 102435, and (). perennis accession
360790 was not significantly different than that on Mars.
DISCUSSIONRice seedlings grown in potting soil showed no visible signs of
nutrient deficiencies, indicating that the organic potting soil can be
used to grow rice seedlings for screening against S. frugiperda larvae.
Contrary to soil, the commercial mixture does not need sterilization or
sifting, therefore reducing handling time. The use of potting soil is
also advantageous since results are comparable over time. Soil however,
can be used with equal results in programs where the price of commercial
soil mixtures is a limiting factor.
Skeletonization of rice seedlings by _S„ frugiperda larvae was
noticeable by the third or fourth day after infestation and heavy (7.0
rating) defoliation was visible after larvae congregated on preferred
cultivars. Heavy defoliation on Mars occurred about seven days after an
initial infestation of four to six neonate larvae per plant.
Comparison of the field and greenhouse tests indicated that the
non-replicated greenhouse screening technique can be successfully used
to detect resistance to frugiperda in rice seedlings. The relative
defoliation of the susceptible cultivars Mars (6.8 rating) and Saturn
(5.4 rating) in the greenhouse was similar to that observed in the field

20
Table 3. Defoliation of selected Oryza species by Spodoptera frugiperda neonate larvae. Baton Rouge, LA 1985.
Plant introduction or accession number +
Speciesname
Mean defoliation t
101800 0. glaberrima 3.0a *
102554 0. glaberrima 3.3a
369453 0. glaberrima 3.4a
360790 0. perennis 3.9ab
102435 0. glaberrima 4. lab
102445 0. glaberrima 4 .8ab
009945 0. sativa (Mars) 6.2b
* Means within a column followed by a letter in common do not differ at the 0.05 level of probability according to Duncan's multiple range test. (MSE = 2.32, df = 24)
t 369453, 360790,and 009945 represent USDA plant introduction numbers, others represent IRRI accession numbers.
1 = 0-10%, 5 = 41-50%, 10 = 91-100% defoliation or skeletonization.

21
(Mars 7.8, Saturn 6.8 rating), suggesting that screening can be
conducted either in the field or greenhouse. Greenhouse screening
techniques have been used to detect resistance in rice seedlings against
M. separata (Wilde and Apostol, 1983) and _S. mauritia (Soejitno and van Vreden, 1976; Wilde and Apostol, 1983).
Plant introductions 160842, 346833, 346853, 346840, and 346830 were
found to have a moderate degree of _S. frugiperda resistance (Table 1).
Defoliation on Pi's 160842 and 346833 was markedly less than on Mars, a
popular United States medium-grain rice cultivar which was heavily defoliated in both the field and greenhouse test. The low defoliation
rate on PI 346833 does not appear to be associated with non-preference
as indicated by the high larval population (63.6%) on that PI in a
choice test. The lower larval population (22.9%) on PI 160842 and the low defoliation rating (4.3) suggests that non-preference is responsible
for resistance in this PI.
Results of the choice test suggested that non-preference was the
principal factor affecting S>. frugiperda feeding on PI 346840. This PI displayed the lowest larval population (10.0%) feeding on it in a
choice situation. Non-preference was also detected on Pi's 346855 and
346824, but both were heavily defoliated in the no choice test.
Non-preference to S_. frugiperda has been reported in pearl millet
(Leuck, 1970; Leuck et al., 1968a) and Bermuda grass (Leuck and Skinner,
1970). Additional research will be required to test if antibiosis is
present in Pi's 160842, 346833, 346840, and 346853.
Results of the United States cultivar test suggest that resistance
against S. frugiperda may be available in the obsolete United States
cultivars, Honduras, Chinese, or Colusa and in the current cultivar

22Newrex (Table 2). However, 62% of the entries sustained defoliation
ratings greater than 5.0. Although not significantly different from Mars, several semidwarf cultivars were severely defoliated (defoliation
rate > 7.0) by JS. frugiperda. In India, two previously unimportant armyworm species became serious pests after the introduction of
semidwarf rice cultivars (Banerjee and Chaterjee, 1982; Papel et al.,
1981; Kulshreshtha et al., 1970). These reports may be an indication
that host selection by armyworms is influenced by plant morphological and growing characteristics.
The lowest defoliation rate recorded in any test occurred on plants
of the (). glaberrima accessions (Table 3). Oryza glaberrima is a
cultivated species of rice in Africa. The highest degree of resistance
against j3. mauritia and M. separata reported by Wilde and Apostol (1983)
was found to exist in wild rice of the species (). eichingeri Peter, 0.
latifolia Desv., j). officinalis Wall, (). punctata Kotschy, and CL ridleyi Hook.
To our knowledge, this is the first report of resistance to
frugiperda larval feeding in rice plants. Host-plant resistance to JS.
frugiperda is currently available in Bermuda grass (Combs and Valerio,
1980; Leuck et al., 1968b; Leuck and Skinner, 1970; Pencoe and Martin,
1982; Quisenberry and Wilson, 1985; Wiseman et al., 1982), pearl millet
(Leuck et al., 1968a; Leuck, 1970, 1972), sorghum (McMillan and Starks,
1967; Wiseman and Gourley, 1982), corn (Brett and Bastida, 1963;
Manuwoto and Scriber, 1982; Ng et al., 1985; Williams et al., 1983,
1985; Wiseman et al., 1966), and peanuts (Leuck and Hammons, 1974; Leuck
and Skinner, 1971).
The results of this test indicate that the evaluation method

developed is effective in identifying sources of resistance to JS.
frugiperda in rice seedlings. Over 5,000 £. sativa plant introductions,
24 United States cultivars, and six accessions of JO. glaberr-*™** and 0.
perennis were evaluated. Various levels of resistance were identified in the United States cultivars Honduras, Newrex, and Chinese; in Pi's
160842, 346830, 346833, 346840, 346853; and (). glaberrima accessions
101800, 102554, and 369453. Additional research should consider the
nature of resistance to frugiperda and the incorporation of resistance into improved rice cultivars.

ACKNOWLEDGEMENT
The authors acknowledge E. Nowick, Louisiana State University Rice Research Station (LSURRS), Crowley, LA; the Entomology Department of the
International Rice Research Rice Institute at Los Banos Laguna,
Philippines; and Dr. D. H. Smith of the USDA National Small Grain
Collection Laboratory at Beltsville, MD for providing seed for the
various tests, and A. M. Hammond, T. Hardy, E. A. Heinrichs, Louisiana
State University Entomology Department, and K. S. McKenzie, LSURRS for
critical review of the manuscript.
24

LITERATURE CITED
Banerjee, P. K. and P. B. Chaterjee. 1982. Pests of hill rice in West Bengal, India. Int. Rice Res. News. 7(2): 11.
Bowling, C. C. 1967. Rearing two lepidopterous pests of rice on a common artificial diet. Ann. Entomol. Soc. Am. 60: 1215-1216.
Bowling, C. C. 1978. Simulated insect damage to rice: Effects of leaf removal. J. Econ. Entomol. 71: 377-378.
Brett, C. H. and R. Bastida. 1963. Resistance of sweet corn varieties to the fall armyworm, Lyphygma frugiperda. J. Econ. Entomol.56: 162-167.
Chandler, J. V., F. Abruna, J. Lozano, S. Silva, A. Rodriguez, and C. T. Ramirez. 1977. Cultivo intensivo y perspectivas del arroz en Puerto Rico. Univ. de Puerto Rico, Estacion Experimental Agricola, Boletin 25.
Combs, R. L. and J. R. Valerio. 1980. Oviposition by the fall armyworm on four varieties of bermudagrass. J. Ga. Entomol.Soc. 15: 164-167.
Dar, I. A. 1979. Occurrence of the rice ear-cutting caterpillar in the Punjab of Pakistan. Int. Rice Res. News. 4(2): 16.
Davis, F. M.. 1980. Fall armyworm plant resistance programs. Fla.Entomol. 63: 420-424.
Davis, F. M. and T. G. Oswalt. 1969. Hand inoculator for dispensing lepidopterous larvae. USDA/SEA/SR 9.
Freund, R. J. and R. C. Littell. 1981. SAS for linear models. SAS Institute, North Carolina.
Gallego, M., F. L. 1967. Lista preliminar de insectos deimportancia economica y secundarios, que afectan los principales cultivos, animales domesticos y al hombre, en Colombia. Revista Facultad Agronomia, Colombia. 65: 32-66.
Kulshreshtha, J. P., M. B. Kalode, and A. Verna. 1970. Paddy cutworm (Pseudaletia separata Walker) and armyworm (Cirphis compacta Moore) as a serious pest of high yielding varieties of rice. Oryza 7: 143-145.
Leuck, D. B. 1970. The role of resistance in pearl millet in control of the fall armyworm. J. Econ. Entomol. 63: 1579-1681.
Leuck, D. B. 1972. Induced fall armyworm resistance in pearl millet.J. Econ. Entomol. 65: 1608-1611.
25

Leuck, D. B. and R. 0. Hammons. 1974. Nutrients and growth media:Influence on expresion of resistance to the fall armyworm in the peanut. J. Econ. Entomol. 67: 564.
Leuck, D. B., C. M. Taliaferro, G. W. Burton, and M. C. Bowman. 1968a.Fall armyworm resistance in pearl millet. J. Econ. Entomol.61: 693-695.
Leuck, D. B., C. M. Taliaferro, G. W. Burton, and M. C. Bowman. 1968b.Resistance in bermudagrass to the fall armyworm. J. Econ. Entomol61: 1321-1322.
Leuck, D. B. and J. L. Skinner. 1970. Resistance in bermuda grass affecting control of the fall armyworm. J. Econ. Entomol.63: 1981-1982.
Leuck, D. B. and J. L. Skinner. 1971. Resistance in peanuts foliage influencing fall armyworm control. J. Econ. Entomol. 64: 148-150
Machado, S. S. 1978. Alguns fatores limitantes da produtividade das lavoras de arroz. Lavoura Arrozeira. 307: 52-54.
McMillan, W. W. and K. J. Starks. 1967. Greenhouse and laboratory screening of sorghum lines for resistance to the fall armyworm larvae. J. Econ. Entomol. 60: 145-146.
Manuwoto, S. and J. M. Scriber. 1982. Consumption and utilization of three maize genotypes by southern armyworm. J. Econ. Entomol.75: 163-167.
Navas, D. 1966. Evaluacion del dano que ocasionan algunas plagas del arroz en Panama. J!n Programa cooperativo centroamericano para el mejoramiento de cultivos alimenticios, XII: 77-78.
Ng, Sen-Seong, F. M. Davis, and W. P. Williams. 1985. Survival,growth, and reproduction of fall armyworm (Lepidoptera:Noctuidae) as affected by resistant corn genotypes. J. Econ. Entomol.78: 967-971.
Papel, R. K., A. K. Khatri, B. S. Chaudharg. 1981. Rice ear-cutting caterpillar, an injurious pest at panicle stage. Int. Rice Res. News. 6(3): 17
Pencoe, N. L. and P. B. Martin. 1982. Fall armyworm (Lepidoptera:Noctuidae) larval development and adult fecundity on five grass hosts. Environ. Entomol. 11: 720-723.
Perkins, W. D. 1979. Laboratory rearing of the fall armyworm. Fla. Entomol. 62: 87-91.

Pophali, D. J., P. D. Deshmukh, U. K. Kaushik, S. K. Shrivastara, S.L. Patidar, and G. A. Sangrade. 1980. Assessment of yieldlosses caused by earhead-cutting caterpillar Mythimna separata(Walker). Int. Rice Res. News. 5(4): 8-9.
Quisenberry, S. S. and H. K. Wilson. 1985. Consumption andutilization of bermuda grass by fall armyworm (Lepidoptera:Noctuidae) larvae. J. Econ. Entomol. 78: 820-824.
Soejitno, J. and G. van Vreden. 1976. Varietal screening forresistance to the armyworm, Spodoptera mauritia, in rice. InAgric. Coop. Indonesia Netherlands Res. Rept. 1968-1976: 166-167.
Wilde, G. and R. Apostol. 1983. Armyworm (Lepidoptera:Noctuidae) resistance in rice. Environ. Entomol. 12: 376-379.
Williams, W. P., F. M. Davis, and B. R. Wiseman. 1983. Fall armyworm resistance in corn and its supression of larval survival and growth. Agron. J. 75: 831-832.
Williams, W. P., P. M. Buckley, and F. M. Davis. 1985. Larval growth and behavior of the fall armyworm (Lepidoptera:Noctuidae) on callus initiated from susceptible and resistant corn hybryds.J. Econ. Entomol. 78: 951-954.
Wiseman, B. R. and F. M. Davis. 1979. Plant resistance to the fall armyworm. Fla. Entomol. 62: 123-130.
Wiseman, B. R. and L. Gourley. 1982. Fall armyworm(Lepidoptera:Noctuidae): Infestation procedures and sorghumresistance evaluations. J. Econ. Entomol. 75: 1048-1051.
Wiseman, B. R., R. C. Gueldner, and R. E. Lynch. 1982. Resistancein common centipedegrass to the fall armyworm. J. Econ. Entomol. 75: 245-247.
Wiseman, B. R., R. H. Painter and C. E. Wasson. 1966. Detecting corn seedling differences in the greenhouse by visual classification of damage by the fall armyworm. J. Econ. Entomol. 59: 1211-1214.
Yoshida, S. 1981. Fundamentals of rice crop science. Int. Rice Res. Inst., Los Banos, Laguna, Philippines.

CHAPTER II
RICE YIELDS AS AFFECTED BY THE FALL ARMYWORM, SPODOPTERA FRUGIPERDA
(J. E. SMITH), (LEPIDOPTERA:NOCTUIDAE)
This chapter is written in the style of
Journal of Economic Entomology
28

ABSTRACT
The effects of fall armyworm, Spodoptera frugiperda (J. E. Smith),
feeding on rice, Oryza sativa L., were studied in the field during 1983
and 1984. Increased £. frugiperda infestation levels resulted in
increased rice defoliation, reduced plant and panicle density, and
reduced rice yields. Yield component studies indicated that the lower
yields on infested plots were the result of reduced plant stand and
panicle density. Kernel weight and percent filled kernels were not
affected. Yield reductions were found to be linearly related to S.
frugiperda larval density. The linear model [E(y) = b x^] accounted for
a large proportion of the variance of yield reduction during both years.
29

INTRODUCTION
The fall armyworm, Spodoptera frugiperda (J. E. Smith), is a sporadic but potentially important pest of rice, Oryza sativa L..
Spodoptera frugiperda damage to rice in the United States was reported
as early as 1845 (Luginbill 1928) and severe damage to rice fields in
North Carolina occurred in 1899 (Chittenden 1900). During 1973, an
outbreak of JS. frugiperda on rice required multiple insecticide
applications to control dense larval populations covering more than 60%
of the total Texas rice production area (Bowling 1978).
In Panama, _S. frugiperda is among the most important insect pests
of rice (Navas 1966), and it is also an economically important rice pest
in Colombia (Gallego 1967) and Brazil (Machado 1978). Chandler et al.
(1977) listed j3. frugiperda as the main insect affecting rice production
in Puerto Rico. Multiple insecticide applications are often required to
control this pest on rice; however, the economic injury level for jS.
frugiperda on rice has not yet been determined.
Little is known about the relationship of j>. frugiperda population
density to yield reduction in rice. The few studies available are based
mainly on artificial defoliation techniques rather than on actual insect
feeding damage. Bowling (1978) and Navas (1966, 1967, 1974) reported
reductions on rice yields by simulated insect damage, but concluded that
rice plants are capable of recovering rapidly from extensive damage,
particularly > if the damage occurs in the early vegetative stage of growth.
Plant response to defoliation by chewing insects may be quite
different from the responce produced by artificial defoliation
30

31techniques. Capinera and Roltsch (1980) reported a lower rate of
regrowth on wheat, Triticum aestivum (L.) affected by grasshopper,
Melanoplus sanguinipes (F.), feeding as compared compared to equal
levels of manual clipping. Data collected by Rice et al. (1982a)
indicate higher rice yield reductions when plants were defoliated by the
common armyworm, Pseudaletia unipuncta (Haworth), as compared to similar
levels of artificial defoliation. These artificial defoliation studies
indicate that yield reduction estimates are not as reliable as those
based on actual insect feeding. The research reported here was
initiated to determine the effect of leaf feeding by varying S_.
frugiperda larval densities on rice grain yield.
MATERIALS AND METHODS
Insects. Insects were obtained from a laboratory (26 ± 0.5°C,
14:10 photoperiod, 50 ± 10% RH) colony that had been under culture for
50 generations at the Louisiana State University Entomology Department,
Baton Rouge, LA. Wild insects (larvae and adults) were collected
biannually and introduced into the colony. Larvae were collected from
Bermuda grass, Cynodon dactylon (L.), corn, Zea mays L., and sorghum,
Sorghum bicolor (L.), and adults from a light trap at the Louisiana State University Rice Research Station, Crowley, LA. Larvae were reared
on a pinto bean diet (Bowling 1967) following the method described by Perkins (1979). Eggs masses were collected daily from rearing
containers and allowed to hatch in plastic bags. Neonate larvae were
transferred (20 to 30 per cup) with a Davis inoculator (Davis and Oswalt
1969) to 250 ml cardboard containers supplied with approximately 120 ml
of pinto bean diet. Larvae were reared to the appropiate instar,

32transported to the field, counted, and released over experimental plots at 1700 hours.
Planting and fertilization. 'Mars', a widely grown medium-grain
rice variety, was planted during the 1983 and 1984 growing seasons.
Prior to planting, the experimental area received 67-67-67 kg per ha,
of N-P20^-K20 respectively. The 1983 test was seeded with a single row
cone planter, while the 1984 test was drill seeded with a Kincaid 7 row
planter (Kincaid Equip. Corp., Havens, KS). Both tests were seeded at
the rate of 112 kg seed per ha. An additional 90 kg per ha of N was
applied just prior to flooding. Propanil (3,4-dichlorophenylpropi-
analide) herbicide was applied prior to permanent flood at the rate of
4.48 kg Al per ha for weed control.
Seven days after the permanent flood was established carbofuran
(2,3-dihydro-2,2-dimethyl-7-benzofuranyl methylcarbamate) was applied
with a shaker jar at the rate of 0.56 kg Al per ha for control of the
rice water weevil, Lissorhoptrus oryzophilus Kuschel. Rice stink bug,
Oebalus pugnax (F.), populations were low and assumed to be uniform over
the experimental plots. Therefore, no chemical control tactics were
implemented.
1983 Experiment. Experimental plots were thinned to 20 homogeneous
plants at the 3.0 stage of growth (Yoshida 1981). Cylindrical PVC plastic pipe barriers (0.381 m diam. by 0.458 m tall) were placed around
the plants after thinning. The PVC pipes were driven into the soil and the inner lip of the open end was covered with tack-trap to prevent
insect movement from the plots. Laboratory reared third instar J5. frugiperda larvae were introduced into the plots at density levels of 0,
17.5, 35.1, 52.6, 70.2, and 87.7 larvae per m2. Larvae were allowed

33
to feed for 6 days. After the feeding period, the amount of
defoliation and the number of surviving plants were recorded. The
cylinders were removed and a permanent flood was established.Non-infested plants were also caged to serve as controls.
At grain maturity panicles from plants in each plot were counted
and hand-harvested. Samples were then taken to the laboratory,
individually threshed, and total kernel and 500 kernels weights were
recorded. The percent filled kernels was determined using the
gravimetric method described by Matsushima (1970).
1984 Experiment. Seven-row plots (3.05 m by 1.22 m with 0.178 m
row spacing) were confined by flexible-metal barriers (0.20 m tall) placed 0.15 m from the outside rows and anchored with wooden stakes.
Plant density was determined prior to infestation by counting the number
of plants on 1 m of row (three samples per plot). Plants in the 3.0
stage of growth were infested with laboratory reared fourth instar _S.
frugiperda larvae at densities of 0, 26.9, 53.8, 80.7, 107.6, and 215.1 larvae per m2. The infestation was terminated after 3 days by removing
the metal barriers and permanently flooding the field.
The amount of biomass removed by larval feeding was estimated by
removing three 0.333 m samples per plot prior to permanent flood.
Foliage samples consisted of all above ground plant material. Samples
were weighed and a biomass index calculated using the leaf area index
method described by Yoshida (1981). The leaf area index was modified as follows: biomass index = biomass weight (mg) / plot area (m).
At grain maturity, three random samples (0.333 m per plot) were
hand-harvested from each plot to study the effects of _S. frugiperda
feeding on panicle density. Panicles were taken to the laboratory,

34
counted, hand threshed, and dried at 30°C until constant weights were
obtained. The total number of kernels and filled kernels on each
panicle were recorded. Filled kernels were determined using the
gravimetric technique described by Matsushima (1970). Plant height
(three heights per plot) was recorded at maturity and entire plots were
harvested with a Comper 730 small plot combine (Susue Co., Japan).
Rough rice harvested from each plot was weighed and adjusted to 12%
moisture prior to calculating yields. Percent yield reductions were
calculated for each plot based on the adjusted weight of rice in
uninfested plots.
Experiments were arranged in a randomized complete block design
with four replications. Data were subjected to the SAS GLM procedure
(Freund and Littell 1981) and means compared by Duncan's new multiple
range test (Duncan 1955). Data were also subjected to regression
analyses to relate insect density (larvae per m2) and percent yield
reductions. Various regression models were tested to obtain the
simplest model that could be used to predict yield reductions.
RESULTS
1983 Experiment. Increased insect population densities resulted in
consistent increases in defoliation and reduced plant and panicle
densities in 1983 (Table 1, Appendix B-Table 1 to 5). A significant
(P = 0.05) increase in defoliation was observed at infestation levels of
35.1 and 52.6 larvae per m2. Significant (P = 0.05) reductions in plant
and panicle density occurred at infestations of 17.5 and 52.6 larvae per
m2, respectively. Plant density in the uninfested plots was
significantly (P = 0.05) higher (39.5%) than that for the 17.5 larvae

Table 1. Defoliation, plant stand, panicle density, kernel weight, and yield of rice as affected by Spodoptera frugiperda larval feeding. Crowley, LA 1983.
Number of Larvae/m2
Defoliation * (%)
DensityPlants
(m2)Panicles
Gross Yield (g)
Weight of 500 Kernels (g)
0.0 0.0a ** 27.4a 78.8a 165.0a 10.6a
17.5 4.8a 16.8b 66.2a 135.0a 12.0a
35.1 33.5b 11.4b 54.8a 120.0ab 11.9a
52.6 82.5c 4.1c 15.8b 60.7bc 10.8a
70.2 90.8c 0.0c 0.0b 0.0c --
87.7 93.3c 0.0c *** 0.0b *** 0.0c *** --
MSE 237.70 8.17 173.18 0.0020 1.05DF 15 14 14 14 6
0 = no defoliation, 50 = 50%, and 100 = complete defoliation.
Means within a column followed by the same letter do not differ (P = 0.05) according to Duncan's new multiple range test (Duncan 1955).
*** Mean of three replicates.

36
per m2. However, panicle density was only 16% lower than that in the
non-infested plots and differences were not significant (P = 0.05).
All infestation levels resulted in yield reductions; however, only
the yields of plots infested with population levels greater than 35.1
larvae per m2 were significantly (P = 0.05) less when compared to the
control plots. An infestation level of 70.2 larvae per m2 caused 100%
plant mortality in all replicates. Only six plants from a single
replicate survived (due to unexpectedly high larval mortality) an
infestation of 87.7 larvae per m2. Therefore, rice collected from the
latter treatment was excluded from the statistical analysis. The 17.5
larvae per m2 infestation level reduced yield by 18%, as compared with
the non-infested plots but the difference was not significant (P =
0.05). The weight of 500 kernels ranged from 10.6 g in the control
plots to 12.0 g in plots infested with 17.5 larvae per m2 but the
difference was not significant (P = 0.05). Similarly there was no
significant difference in panicle weight or percent fertile kernels
(Appendix B-Tables 6 & 7).
1984 Experiment. Damage by jS. frugiperda larvae was observed after
the first night of feeding in the 1984 experiment. Generally, upper
leaves were consumed first, but whole plants were consumed as the
infestation period progressed. Although the larvae were uniformly
distributed over the plots at infestation, they congregated in the
shaded areas produced by the metal barriers. As a result, defoliation
was heavy on the outside plot rows and decreased toward the center rows.
Increased larval population densities also resulted in reduced
panicle density and reduced yield in 1984 (Table 2, Appendix B-Table 8
to 13). The calculated biomass index decreased with increased S.

Table 2. Biomass index* yield, and yield components of rice as affected by Spodoptera frugiperda larval feeding. Crowley, LA 1984.
Number of Larvae/m2
Biomass index * (mg/m)
PanicleDensity/m2 Wgt(g)
Mature plant Hght (cm)
Wgt (g)/1000 Kernels
Yield ** (kg/ha)
0.0 996.6a *** 97.0a 2.68ab 107.3d 26.79a 10,267a
26.9 417.6b 84.3a 2.80ab 110.8a 25.89a 9,922a
53.8 556.6b 80.5a 2.98a 110.0a 26.37a 10,003a
80.7 454.3b 82.0a 2.67ab 110.0a 25.42a 9,55lab
107.6 621.1b 103.5a 2.65ab 109.3a 26.60a 9,885a
215.1 329.7b 88.5a 2.52b 101.5b 26.38a 8,511b
MSE 39264.72 401.18 0.049 13.79 0.977 584435.53DF 15 15 15 15 15 14
Biomass index = weight of above ground foliage (mg) / area (m).
Adjusted to 12% moisture.
*** Means within a column followed by the same letter do not differ (P = 0.05) according to Duncan's new multiple range test (Duncan 1955).

38frugiperda larval densities. The smallest infestation of 26.9 larvae
per m2 caused a significant (P = 0.05) decrease in biomass index as compared to the control.
The number of panicles per m2 generally decreased as the number of
insects per m2 increased. With the exception of the 107.6 larvae per m2
infestation density, panicle density was lower in all infested plots
than in control plots, but no significant (P = 0.05) differences were
detected. Significantly (P = 0.05) lower panicle weights were collected
from plots infested with 215.1 larvae per m2 than from plots infested
with 53.8 larvae per m2, but weights of panicles in the control plots
were not significantly (P = 0.05) different than weights of panicles of
plants infested at all other levels (Table 2).
Only the highest population density of 215.1 larvae per m2
significantly (P = 0.05) reduced mature plant height (Table 2). Plants
from all other infested plots were slighty taller than the control
plants, but the differences were not significant (P = 0.05).
Feeding by £. frugiperda larvae had no effect on kernel weight or
the percent filled kernels. The weight of 1000 kernels ranged from
26.79 to 25.42 g, but the difference was not significant (P = 0.05).
Similarly there were no significant (P = 0.05) differences (Appendix
B-Table 14) in the percentage of filled kernels, which ranged from 94.5
to 95.0%.All infestation levels caused yield reductions, but only the 215.1
larvae per m2 infestation level produced a significant (P = 0.05)
reduction. Mean yield of plots infested with 215.1 larvae per m2 was
17% lower than the yield of uninfested plots.
The linear model E(y) (expected % yield loss) = b x^, where b =

Table 3. Regression analysis of Spodoptera frugiperda larval density (larvae/m2) on percent rice yield reduction. Crowley, LA 1983-1984.
Fitted model SE of b r2 F PTest E(y)=b x^ Estimate Value Value Value
1983 E(y)=0.072864 xx 0.00679
1984 E(y)=0.000719 x 0.00013
0.84
0.60
115.26 0.0001
32.57 0.0001

Figure 1. Regression of Spodoptera frugiperda larval density and
percent rice yield reduction. Crowley, LA 1983-1984.
40

Yield
redu
ctio
n (%)
(— * oo
00o 0.841983 E(y) = 0.072864 x1984 E(y) = 0.000719 x
iso
0.60
140 180 200 22020 80 100 120 .16040 602
Number of Spodoptera frugiperda per m “”

regression coeficient and x^ = insect density per m2, fit the yic’ld
reduction data for both years (Table 3, Figure 1). Plots of residuals
indicate no abnormality on the fitted model and r2 values (r2= 0.84,
22 df for 1983 and r2 = 0,60, 22 df for 1984) were highly significant (P
= 0.01). The addition of quadratic and/or cubic components were not
significant, indicating that these terms are not needed in the model.
DISCUSSIONThe lower feeding activity observed during 1984 was probably the
result of the shorter infestation period (3 days) compared to the 6 day
infestation period in 1983. This effect however, is confounded with
year related environmental differences and insect physiological age.The third stage larvae used in 1983 reached the next stage (fourth)
before termination of the experiment, the defoliation recorded was thus a result of comulative feeding by third and fourth stage larvae.
Luginbill (1928) indicated that fourth instar S_. frugiperda larvae
consume more foliage (3.9%) as compared to third instars. The damage
recorded in 1984 was the result of 3 days of feeding by fourth stage
larvae. Additional research will be required to test the effects of
feeding by jS. frugiperda larvae of different physiological ages.The unexpectedly high yields for plots infested with the 53.8 and
107.6 larvae per m2 during the 1984 parallels the low defoliation on
those treatments (Table 2). The relatively low defoliation for those
two treatments was probably the result of high larval mortality. The
larger plots (3.05 by 1.24 m) used in 1984 provided a more appropriate
environment for predators than the smaller (0.381 by 0.458 m)
cylindrical barriers used in 1983. The smaller, and taller PVC pipe

43
barriers also provided protection against the sun during most of the
day. Larvae were uniformly distributed over the plots during both
years, but in 1984 insects congregated along the shaded areas provided
by the metal barriers. This behavior contributed to higher mortality
because S. frugiperda becomes cannibalistic when crowded (Luginbill
1928). Rice et al. (1982a) indicated that larvae of £. unipuncta
concentrate in areas of low light intensity in California rice fields.
Spodoptera frugiperda larvae respond similarly to light (Luginbill 1928).
The effects of larval defoliation on mature plant height were only detected at an infestation of 215.1 larvae per m2. These data suggest
that reduced plant height is associated with reduced regrowth after
heavy defoliation. Capinera and Roltsch (1980) reported a lower rate of
regrowth on wheat defoliated by grasshoppers as compared to comparative
amounts of manual clipping. Rice et al. (1982a) reported similar
effects on rice defoliated by £. unipuncta larvae as compared to similar
amounts of manual defoliation, indicating that insect feeding canv-affeet1’ plant regrowth and yield.
Spodoptera frugiperda larval feeding on rice seedlings had no
significant effect on kernel weight or filling (Tables 1 & 2). Results
of the yield component studies suggest that rice yield reductions ' » <•
associated with jS. frugiperda larval feeding are a combination of a
reduction in panicle density and weight. Panicle densities during 1983
decreased with increased 8. frugiperda densities and were significantly
(P = 0.05) lower for population densities in excess of 52.6 larvae per
m2. With the exception of the 107.6 larvae per m2 infestation, all
infestation levels in the 1984 test reduced panicle density, as compared

44to the uninfested control. The lack of statistical differences was
probably the result of the small sample size or sampling technique.
Three random plot samples were collected in 1984, while whole plots
where examined in 1983. As previously stated, defoliation was heavier
on the outside rows as compared to middle rows. The effect of jS.
frugiperda larval feeding was probably 'diluted' by combining the three
samples. The high panicle density in the 107.6 larvae per m2
infestation was directly related to the low feeding activity (high
biomass index) observed on that treatment (Table 2).
In spite of small irregularities in _S. frugiperda larval feeding
observed during the 1984 test, yield reductions paralleled reductions in panicle density and panicle weight, two of the morphological components
of rice yield. Rice et al. (1982a) reported similar panicle weight
reductions on rice plants defoliated by _P. unipuncta larvae.
Rice yield reduction was linearly related to _S. frugiperda larval
density, but no significant (P = 0.05) effects were detected on rice
kernel weight or percent kernel filling, Rice et al. (1982a) reported a linear relationship between JP. unipuncta larval density and percent
defoliation on rice plants, but did not correlate larval density with defoliation or yield reductions. The difference in the slope of the
linear regression (b) found between years (b = 0.072864 in 1983 and b =
0.000719 in 1984) was probably due to year related variations, duration
of infestation period, cumulative defoliation effects on plant growth,
or a combination of these factors.
The highest infestation level tested in this experiment (215.1
larvae per m2) is lower than the average natural S_. frugiperda larval
density (251 larvae per m2) recovered from upland rice fields in Panama

45
(Navas 1974) and approximately one-half that of the highest infestation
level (420 larvae per m2) reported for rice fields (Navas 1974). Young
(1979) indicated that 33.3 medium to mature j>. frugiperda larvae per m2
would cause economic damage to pastures or broadcast small grains.
Pophali et al. (1980) reported rice yield losses ranging from 3.6 to 22%
by populations of 12 to 14 M. separata larvae per m2. An average
population of 1.2 to 5.4 P. unupuncta larvae per hill produced 5 to 95%
leaf damage to rice plants in Pakistan (Dar 1979), but effects on yield
were not reported. Rice et al. (1982a) recorded yield losses up to 50%
due to 25 to 30% defoliation by £. unipuncta larvae in California rice
fields. The greatest number of JP. unipuncta larvae recovered was 0.37 larvae per plant (Rice et al. 1982b).
The results of this study indicate that _S. frugiperda larval
population levels as low as 26.9 larvae per m2 can reduce rice yields;
however, 215.1 larvae per m2 are required to produce a significant
reduction on rice yield. These population levels are equivalent to one
and nine larvae per ten plant respectively. Reduced yields were
directly related to increased defoliation and reduced panicle density
and weight. Rice yield reduction was linearly related to S_. frugiperda
larval density, but no significant effects were detected on rice kernel
weight or percent kernel filling. Additional research is necessary to consider the effects of S. frugiperda leaf feeding damage on other rice
cultivars.

ACKNOWLEDGEMENTS The authors acknowledge the Louisiana State University Rice
Research Station (LSURRS) Crowley, LA for providing land, personnel, and
equipment for the experiments. The authors are indebted to G. B. Trahan
and R. W. Webster, LSURRS; C. Cook, J. Negron, M. Riley, and K. Samoil, Entomology Department; and P. Caballero, Chemistry Department for
technical assistance; and S. J. Johnson, S. S. Quisenberry, and T. J.
Riley, Entomology Department for critical review of this manuscript.
46

LITERATURE CITED
Bowling, C. C. 1967. Rearing two lepidopterous pests of rice in acommon artificial diet. Ann. Entomol. Soc. Am. 60: 1215-1216.
Bowling, C. C. 1978. Simulated insect damage to rice: Effects of leafremoval. J. Econ. Entomol. 71: 377-378.
Capinera, J. E. and W. J. Roltsch. 1980. Response of wheat seedling to actual and simulated migratory grasshopper defoliation. J. Econ. Entomol. 73: 258-261.
Chandler, J. V., F. Abruna, J. Lozano, S. Silva, A. Rodriguez, and C. T. Ramirez. 1977. Cultivo intensivo y perspectivas del arroz en Puerto Rico. Univ. de Puerto Rico, Estacion Experimental Agricola, Boletin 25.
Chittenden, F. H. 1900. The fall armyworm in 1899. USDA Dept. Agric. Entomol. Bull. (S.) 23: 78-85.
Dar, I. A. 1979. Occurrence of the rice ear-cutting caterpillar in the Punjab of Pakistan. Int. Rice Res. News. 4(2): 16.
Davis, F. M. and T. G. Oswalt. 1969. Hand inoculator for dispensing lepidopterous larvae. USDA/SEA/SR 9.
Duncan, D. B. 1955. Multiple range and multiple F tests. Biometrics 11: 1-41.
Freund, R. J. and R. C. Litteil. 1981. SAS for linear models. SAS Institute, North Carolina.
Gallego, M., F. L. 1967. Lista preliminar de insectos de importancia economica y secundarios, que afectan los principales cultivos, animales domesticos y al hombre, en Colombia. Revista Facultad Agronomia, Colombia. 65: 32-66.
Luginbill, P. 1928. The fall armyworm. USDA Tech. Bull. 34.
Machado, S. S. 1978. Alguns fatores limitantes da produtividade das lavoras de arroz. Lavoura Arrozeira. 307: 52-54.
Matsushima, S. 1970. Crop science in rice. Fiji Pub. Co. LTD. Tokyo, Japan.
Navas, D. 1966. Evaluacion del dano que ocasionan algunas plagas del arroz en Panama, jln Programa Cooperative centroamericano para el mejoramiento de cultivos alimenticios, XII: 77-78.
47

48
Navas, D. 1967. Efectos de la destruccion del follaje en losrendimientos del arroz. In Programs cooperative centroamericano para el mejoramiento de cultivos alimenticios. XIII: 85-87.
Navas, D. 1974. Fall armyworm in rice. pp. 99-106 _In Proceedings Tall Timber Conference on Ecological Animal Control by Habitat Management.
Perkins, W. D. 1979. Laboratory rearing the fall armyworm. Fla. Entomol. 62: 87-91.
Pophali, D. J., P. D. Deshmukh, U. K. Kaushik, S. K. Shrivastara, S. L. Patidar, and G. A. Sangrade. 1980. Assessment of yield losses caused by earhead-cutting caterpillar Mythimna separata Walker.Int. Rice Res. News. 5(4): 8-9.
Rice, S. E., A. A. Grigarick, and M. 0. Way. 1982a. Effect of leaf and panicle feeding by armyworm (Lepidoptera: Noctuidae) larvae on rice grain yield. J. Econ. Entomol. 75: 593-595.
Rice, S. E., A. A. Grigarick, and M. 0. Way. 1982b. Relationship of larval density and instars of Pseudaletia unipuncta to rice leaf feeding. Environ. Entomol. 11: 648-651.
Yoshida, S. 1981. Fundamentals of rice crop science. Int. Rice Res. Inst., Los Banos, Philippines.
Young, J. R. 1979. Assessing the movement of the fall armyworm, Spodoptera frugiperda, using insecticide resistance and wind patterns, pp. 344-351. Movements of Highly Mobile Insects.R. C. Rabb, and G. G. Kennedy (eds.) North Carolina State Univ.

CHAPTER III
NATURAL CONTROL AGENTS AFFECTING
SPODOPTERA FRUGIPERDA (LEPIDOPTERA:NOCTUIDAE) INFESTING RICE
IN PUERTO RICO
This chapter is published in
Florida Entomologist 68:488-490.
49

FLORIDA ENTOMOLOGIST49a
4628 NW 40th Street Gainesville FL 32606
November 3; 1985
Alberto Pantoja Department of EntomoIo9y 482 Life Sciences Bu.ildin9 Louisiana State University Baton Rou9e; LA 70803-1718
Dear Mr. PantoJa:Vou are hereby granted Permission to use the material contained in the article ■'Natural control agents affecting JSfijsdoPJfceca ££u9JPerda infesting rice in Puerto Rico' by A. PantoJa; C. M. Smith; and J, F. Robinson that appeared in Florida Entomologist 68:488-498. Articles in Florida Entomologist are not copyrighted.I surmise from your request that you have completed your graduate studies. Congratulations! I hoPe you will consider Florida Entomologist when publishing your future research.
Sincerely;
John R. McLaughlin Editor

ABSTRACT
Twenty-eight parasitoid adults, larvae, and puparia were recovered
from Spodoptera frugiperda (J. E. Smith) collected from rice fields in
Puerto Rico. Eight were identified as Lespesia sp., one as Archytas
mormoratus (Townsend), and 19 were unidentifiable (possibly Chelonus sp.) due to premature S_. frugiperda larval death which prevented
parasitoid development into puparia. The fungus Nomuraea rileyi
(Farlow) Sampson killed eight larvae and three were infected by an
unidentified microsporidium. About 70% of the J3. frugiperda larvae died
after refusing to eat the diet or died as pupae of unknown causes.
50

INTRODUCTION
The fall armyworm, Spodoptera frugiperda (J. E. Smith), is a
sporadic but important pest of rice, Oryza sativa L., in the United
States (Bowling 1978). Chandler et al. (1977) listed _S. frugiperda as
the main insect pest affecting rice production in Puerto Rico,. Ashley
(1979), Gardner and Fuxa (1980), and Lewis and Nordlund (1980) listed
parasites and pathogens recovered from JS. frugiperda larvae infesting
various host plants but over one-half of the pathogens and the
parasitoids listed do not indicate the host plant from which _S.
frugiperda larvae were recovered. To date, no parasitoids or pathogens
have been reported as being recovered from j>. frugiperda larvae infesting rice fields.
MATERIALS AND METHODS
S. frugiperda larvae (second and third instars) were collected from
infested rice fields at Arecibo, Puerto Rico on January 4 and February
2, 1984. A total of 720 larvae were collected and placed in plastic
cups containing 30 ml of pinto bean diet capped, and transported to the
Louisiana State University (LSU) Entomology Department under Animal and
Plant Health Inspection Service permit 20 PPQ 576 and 599. Upon return
to the laboratory, larvae were removed from the collection containers,
placed on fresh diet, and held at a 13:11 light: dark cycle and constant
temperature of 26°C. Larvae were monitored daily for mortality and/or
parasitoid emergence. Parasitoid puparia were removed from the diet container and placed in individual containers for emergence of adult
parasitoids. Pupae of _S. frugiperda were also monitored for emergence
51

52
of parasitoids. Adult parasitoids were identified by N. E. Woodley of
the USDA Insect Identification Institute at Beltsville, Maryland (IIBII). Voucher specimens were deposited in the museum of the LSU
Entomology Department, Baton Rouge, Louisiana. Pathogens were
identified by personnel in the insect pathology project at LSU.
RESULTS AND DISCUSSION
Twenty-eight parasitoid adults, larvae, and puparia were recovered.
Eight were identified as Lespesia sp. (Diptera:Tachinidae), one as
Archytas mormoratus (Townsend) (Diptera:Tachinidae) and 19 were
unidentifiable (possibly Chelonus sp., (Hymenoptera:Braconidae)) due to
premature _S. frugiperda larval death which prevented parasitoid
development into puparia. The fungus Nomuraea rileyi (Farlow) Sampson
killed eight larvae and three were infected by an unidentified
microsporidium. About 70% of the S_. frugiperda larvae died after
refusing to eat the diet or died as pupae of unknown causes.
Lespesia spp. have been recorded as parasitoids of S_. frugiperda on
corn and peanuts in Argentina, Colombia, Guatemala, Puerto Rico, the
United States, Uruguay, and Venezuela (Ashley 1979). The group needs
revision, as species are not identifiable (F. C. Tompson; IIBII,
personal communication). To our knowledge this is the first time
Lespesia has been recovered from _S. frugiperda infesting rice.Archytas sp. has been recovered from S_. frugiperda collected on
alfalfa, corn, and peanuts in Argentina, Brazil, Chile, Colombia, Cuba,
Mexico, Paraguay, Peru, Puerto Rico, the United States, Uruguay, and
Venezuela (Ashley 1979). A. mormoratus; however, has not been reported
previously as parasitizing frugiperda in Puerto Rico, and has not

53been reported in relation to rice.
The fungus N. rileyi has been reported previously from S.
frugiperda in Puerto Rico (Luginbill 1928) and Colombia (Gardner and
Fuxa 1980), but the host plant from which J3. frugiperda larvae were
recovered was not stated. It is most likely that the bacterium was a
saprophyte or wound invader and probably not the primary cause of death.
Microsporidia and bacteria have been reported as pathogens of S.
frugiperda in the United States and Venezuela (Gardner and Fuxa, 1980),
but not from larvae collected on rice.
Although j3. frugiperda destruction by parasitoids in Puerto Rican
rice fields is lower than that reported in south Florida corn fields
(Ashley et al. 1982), this study provides important information on
parasite and pathogen host plant interactions. To our knowledge, this
is the first report of parasitoids and/or entomopliagous pathogens being
recovered from S. frugiperda larvae infesting rice. Furthermore, this study is the first report of A. mormoratus parasitizing _S. frugiperda
larvae in Puerto Rico.

ACKNOWLEDGEMENTS We thank Rafal Olmeda and Jose Pantoja of the Puerto Rico
Agricultural Extension Service for their help in the location and
collection of the larvae. We are also grateful to J. R. Fuxa, LSU for
identification of the pathogens and to N. E. Woodley, USDA IIBII for
identification of the parasitoids. We appreciate the comments of J. R.
Fuxa, R. A. Goyer, and D. J. Boethel on an early draft of this note.
54

REFERENCES CITED
Ashley, T. R. 1979. Classification and distribution of fall armyworm parasitoids. Fla. Entomol. 62: 114-123.
Ashley, T. R., V. H. Waddill, E. R. Mitchell, and J. Rye. 1982. Impact of native parasitoids on the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), in South Florida and release of the exotic parasite, Eiphosoma vitticole (Hymenoptera: Ichneumonidae). Environ. Entomol. 11: 833-837.
Bowling, C. C. 1978. Simulated insect damage to rice: effects of leaf removal. J. Econ. Entomol. 71: 377-378.
Chandler, J. V., F. Abruna, J. Losano, S. Silva, A. Rodriguez, and C. T. Ramirez. 1977. Cultivo intensivo y perspectivas del arroz en Puerto Rico. Univ. of Puerto Rico, Agricultural Exp. Stat. Bull. 250.
Gardner, W. A. and J. R. Fuxa. 1980. Pathogens for the suppression of the fall armyworm. Fla. Entomol. 63: 439-447.
Lewis, W. J. and D. A. Nordlund. 1980. Employment of parasitoids and predators for fall armyworm control. Fla. Entomol. 63: 433-438.
Luginbill, P. 1928. The fall armyworm. USDA Tech. Bull. No. 34.
55

CHAPTER IV
DEVELOPMENT OF FALL ARMYWORM, SPODOPTERA FRUGIPERDA (J. E. SMITH),
(LEPIDOPTERA:NOCTUIDAE) FROM LOUISIANA AND PUERTO RICO:
A BIOLOGICAL COMPARISON
This chapter is written in the style of
Environmental Entomology

ABSTRACT
Development, longevity, and reproduction of fall armyworm,
Spodoptera frugiperda (J. E. Smith) from Louisiana (LA) and Puerto Rico (PR) were studied on artificial diet and rice foliage. At egg hatch,
larvae of PR origin were smaller than LA larvae. Differences found in
neonate larval size were also evident at all stages of development but
did not extend to adult longevity or reproductive parameters. Larvae, pupae, and adults of the LA strain were heavier and completed
development in a shorter period of time than the PR strain. Differences
in developmental parameters were not determined by diet, but were
influenced by sex. Differences are probably under genetic control and
may indicate some degree of reproductive isolation.
57

INTRODUCTION
Since its first recorded appearance in the United States (Luginbill
1928), the fall armyworm (FAW), Spodoptera frugiperda (J. E. Smith), has
been responsible for annual crop losses estimated in excess of $300
million (Mitchell 1979). Due to its migratory habits and the lack of a
diapause mechanism to overwinter (Sparks 1979), considerable effort has
been devoted to defining FAW dispersal patterns and the factors
governing them. Young (1979) stressed the importance of using
insecticide resistance and wind patterns to assess FAW movement. Others
have used insecticide susceptibility as affected by host-plant
interactions (Combs and Chambers 1979), and morphological characters
(Wiltshire 1977, as cited by Sparks 1979) to differentiate FAW from various geographical areas.
Although it has been suggested that morphological races of FAW may
exist (Wiltshire 1977, as cited by Sparks 1979) and that mainland
populations may originate from the Caribbean area, little effort has
been devoted to document the development of FAW originating from various
geographical areas. Snow et al. (1968) and Sparks (1979) found no
difference in pheromone responses between FAW populations from Georgia
and the United States Virgin Islands, or between populations from
Florida and Bolivia. Recently, Pashley et al. (1985) examined genetic
relationships among FAW populations from South and Central America,
Mexico, the United States, and Puerto Rico. Differences were largely
due to a divergent Puerto Rican population. There is a current need for
additional information on FAW host-plant interaction and migration.
Here, we report biological comparisons of FAW collected in Puerto Rico
and Louisiana.
58

MATERIALS AND METHODSDevelopment on an artificial diet. The Louisiana strain was
obtained from a laboratory colony at Louisiana State University
Entomology Department. Wild insects (larvae and adults) were field
collected and introduced biannually into the laboratory colony. Larvae
were collected from Bermuda grass (Cynodon dactylon (L.)), corn, (Zea
mays L.), and sorghum (Sorghum bicolor ( L.)) Adults were collected
when available (May to September) from a light trap at the Louisiana
State University Rice Research Station, Crowley, LA. As a final
measure to avoid genetic inbreeding, wild larvae were collected from (L
dactylon at Clinton, LA, and adults from a light trap at Crowley, LA and introduced into the colony two generations prior to the first
experiment. The Puerto Rican strain was obtained as described by
Pantoja et al. (1985) and allowed to adapt to captivity for five
generations prior to experimentation. A small sample of individuals of
the Louisiana and Puerto Rican strains were examined electrophoretically
after four generations in the laboratory and genetic variability was
comparable to field collected populations.
Both strains were reared on a pinto bean diet (Bowling 1967)
following the procedure described by Perkins (1979). The diet was
dispensed in 30 ml plastic diet cups (10 to 15 ml of diet per cup) and arranged on trays (22 or 44 cups per tray). One neonate fall armyworm
larvae was placed on each diet cup with a camel's hair brush, and cups
were capped and numbered. The cups on each tray were divided evenly
between the two fall armyworms strains. Trays were kept in a rearing
room maintained at 26 ± 0.5°C, 14 h photophase, 50 ± 10% RH. A total of
59

60132 insects per strain were studied on four separate replicated
experiments conducted over time.
Larval weight was recorded 8 days after experiments began and daily
from day 10 to pupation. Weight (24 h post-pupation), days to pupation, pupal development time, days to eclosion, and adult weight were also
recorded. Adults were sacrificed prior to weighing by freezing at -5°C for 24 h.
Development on rice foliage. Larval development on 'Mars' rice
foliage was studied in the laboratory using rice grown in the greenhouse
(Sylvania Metal Halide 1000 W lighting, 14 h photophase, 26 ± 10°C, 70 ±
10% RH). Twenty-four neonate larvae per strain were confined
individually on 100 by 10 mm plastic petri dishes containing 40 day old
rice foliage. Petri dishes were lined with filter paper (Whatman it3)
and moistened with distilled water. Each dish was inspected daily for
discarded head capsules (evidence of moulting) and to replace uneaten
foliage. Developmental parameters were recorded as described above.
Rice consumption and utilization. Rice consumption and utilization
was measured using newly moulted third instar larvae reared on pinto
bean diet. Larvae (n = 34 per strain) were removed from pinto bean diet
and fed 'Mars' rice foliage for 24 h following the procedure described
above and 24 larvae per strain were then transferred to a clean petri dish and allowed to feed for 48 h on a known amount of rice. The
remaining larvae were sacrificed and dried (48 h at 60°C) to calculate
initial larval dry weight. After feeding, larvae were sacrificed, dried
and weighed. Remaining food and feces were also dried and weighed.
Approximate digestibility (AD), efficiency of conversion of ingested
food (ECI), and efficiency of conversion of digested food (ECD) were

61
calculated as described by Beck and Reese (1976). Larval growth rate
(GR) was calculated as "relative growth rate" (Kogan and Cope 1974) with
the exception that gut content was not subtracted from the final larval
dry weight. Consumption index (Cl) was calculated as described by
Waldbauer (1968). Nutritional indices were defined as follows:
AD = (dry weight food ingested) - (dry weight feces / dry weight food ingested) x 100.
ECI = (dry weight gained / dry weight food ingested) x 100.
ECD = (dry weight gained) / (dry weight food ingested - weight of feces)
x 100.
GR = (1 / duration test (days)) x (In (final dry larval weight / initial
larval weight)).
Cl = dry weight rice ingested / dry weight gain x duration test (days).
Adult fecundity and longevity. Adult fecundity and longevity were
studied on FAW reared on pinto bean diet, as described above. Pupae
were segregated by sex and adults allowed to emerge in the diet cups.One newly emerged female was confined with two males of equal age and
strain and held in 500 ml cardboard containers supplied with a honey-beer solution. Containers were randomized and kept in rearing
room (14 h photophase, 50 ± 10% RH). Fecundity was measured on 30
females (15 per strain) by counting the number of egg masses deposited
daily until moths died. Egg masses were held in plastic bags and
allowed to hatch (evidence of mating). Head capsule width of neonate
larvae (n = 75 per strain) were measured under magnification (50 x) to
compare first instar larval size.
All experiments were arranged as a randomized complete block design
(block = tray) and data were subjected to the SAS (Freund and Littell

62
1981) General Linear Models (rice consumption and utilization and adult
fecundity and longevity) or paired t test (development on diet and rice
foliage).
RESULTSDevelopment on artificial diet. The Puerto Rican (PR) FAW strain
attained larval weights ranging from 33 to 52% of that of the Louisiana
(LA) FAW, but attained approximately equal pupal and adult fresh weights
(Table 1). With two exceptions days 12 and 14 female LA FAW attained
significantly higher (P = 0.04) larval weight than female PR FAW.
Significantly (P = 0.05) larger male LA FAW larvae were recorded for
days 10, 13, and 14. Pupal weights of female PR FAW were significantly
(P = 0.05) lower than those of LA FAW. Female adult weight was
significantly (P = 0.02) greater for LA FAW than for PR FAW, but weights
of male adults were not significantly different (P = 0.59).
Total development, egg to adult, was significantly (P = 0.03)
shorter for LA FAW of both sexes than for PR FAW (Table 2). Length of
larval life was significantly (P = 0.02) shorter for LA FAW of both
sexes than for PR FAW. No significant differences (P = 0.10) existed in
length of pupal development.The highest average larval weight was for LA FAW males (503.9 mg on
day 14), while the highest pupal weight recorded was for female LA FAW
(240.3 mg). With one exception (day 12) PR male FAW larvae were
slightly larger than PR females, while the opposite was true for the LA
strain, with the exception of day 14.Development on rice foliage. Development of FAW larvae on rice foliage
(Table 3) was similar to that of larvae reared on pinto bean diet.

Table 1. Weight of Spodoptera frugiperda from Louisiana (LA) and Puerto Rico (PR) reared onartificial diet. Baton Rouge, LA 1984.
Stage __________________________________ Mean wgt. (mg) i SE________________
duration Females Males
(days) _______ LA_______ PR_______ LA________ PR
Larva n n P * n n P *
8 79.7 ± 11.3 55 33.1 + 7.6 29 0.01 73.8 + 11.4 57 48.0 + 10.5 41 0.15
10 255.3 ± 29.3 55 110.5 + 18.3 29 0.01 241.2 + 30.7 57 124.8 + 20.0 41 0.01
11 308.2 + 33.2 55 179.6 + 33.2 29 0.01 220.2 + 25.1 55 205.1 + 31.0 41 0.71
12 363.4 + 36.2 49 269.7 + 37.3 27 0.08 300.0 + 27.0 52 254.9 + 27.8 38 0.16
13 426.3 + 46.3 37 293.7 + 37.4 21 0.04 425.4 + 28.6 43 306.5 + 28.9 28 0.02
14 437.2 ± 64.6 26 366.7 + 41.2 11 0.26 503.9 + 57.7 28 386.9 + 63.7 20 0.05
Pupa 240.3 ± 4.8 41 219.6 + 8.2 28 0.05 234.6 + 4.3 43 220.4 + 6.5 41 0.23
Adult 142.0 ± 4.9 25 125.3 + 6.4 10 0.02 129.6 + 3.0 30 133.8 + 9.9 10 0.59
* P = T, Contrast LA versus PR, n = number of observations.

Table 2. Developmental time (days) of Spodoptera frugiperda from Louisiana (LA) andPuerto Rico (PR) on artificial diet. Baton Rouge, LA 1984.
Mean ± SE
Days to pupation Pupal development Days to eclosionStrain Females Males Females Males Females Males
LA 15.1 ± .26 14.9 ± .37 9.1 ± .27 10.0 ± .13 24.5 ± .53 24.8 ± .36
PR 16.7 ± .37 16.1 ± .48 9.8 ± .35 10.6 ± .28 27.2 ± .32 27.0 ± .87
P = X * 0.01 0.02 0.12 0.10 0.01 0.03
* Contrast LA versus PR, n females: LA = 41, PR = 28; n males: LA = 43, PR = 23.

Table 3. Development of Spodoptera frugiperda from Louisiana (LA) and Puerto Rico (PR) on 'Mars' ricefoliage. Baton Rouge, LA 1984.
Mean weight (mg) ± SE Mean (± SE) no. days to
Larva @ 11 days Pupa @ 24 h Adult @ emergence Pupation
Females Males Females Males Females Males Females Males
LA
PR
115.8 ±24.6
55.8 ± 5.5
99.0 ±17.8
75.4 ± 9.9
187.0 ± 6.7
153.8 ±11.0
184.7 ± 5.3
197.2 ±17.5
90.0 ± 5.0
82.5 ±13.3
90.8 ±4.5
109.3 ±7.7
17.5 ±.65
19.1 ±.48
17.6 ±.38
18.6 ±.24
P = T * 0.02 0.21 0.04 0.57 0.41 0.31 0.02 0.07
* Contrast LA versus PR, n females: L A = 8 , P R = 1 2 ; n males: LA = 16, PR = 10.

66Female LA FAW attained significantly (P = 0.04) greater larval and pupal
weights and completed larval development in a significantly (P = 0.02)
shorter period of time than PR females, but differences in adult fresh
weights and total development times (LA = 28.5, PR = 29.0) were not
significant. No significant differences were detected with respect to
male developmental parameters except larval development, which was
longer (P = 0.07) for PR males than for LA males. These values were
consistent with those observed among larvae fed pinto bean diet (Table
1). For both FAW strains growth (larval, pupal, and adult weight) on
rice was markedly reduced and time to complete development was prolonged as compared to growth on pinto bean diet.
Females and males of both strains completed development with
approximately equal numbers of instars (Table 4). Female LA larvae
spent a significantly (P = 0.01) shorter time in the second stage than
the PR females. The shortest development time observed in any stage was
1.6 days for LA females at instar group three. Male PR larvae required
an extra 1.5 and 2.6 days to complete development (P = 0.07) in instars
three and four, respectively, than LA males. Only one PR male and one
LA female reached the seventh instar, therefore no within origin
comparisons were possible.
Rice consumption and utilization. No significant (Appendix C-Tables 1
& 2) differences in consumption index (LA = 4.81, PR = 5.58%) or growth
rate (LA = 0.11, PR = 0.13) were detected between the two FAW strains.
Similarly there was no significant (Appendix C-Tables 3 & 4) differences
in approximate digestibility (LA = 59.7, PR = 58.2%) or efficiency of conversion of ingested food (LA = 21.8, PR = 20.7%). Efficiency of
convertion of digested food was identical (38.4%) for both FAW strains

Table 4. Developmental time (days) and number of instars of Spodoptera frugiperda larvae from Louisiana(LA) and Puerto Rico (PR) fed 'Mars' rice foliage. Baton Rouge, LA 1984.
Mean (± SE) days at instar group Mean No.
Strain Sex n I 11 III IV V VI VII Instars ± SE
LA Females 8 4.5 + .3 1.8 ± .2 1.6 ± .2 2.0 ± .3 3.5 ± 1 4.0 ± 1 2.0 5.7 ± .9
PR Females 12 3.7 + .3 3.0 ± .3 2.2 ± .2 2.8 ± .8 3.5 ± .9 4.2 ± 1 5.0 ± .6 5.7 ± .4
LA Males 16 3.0 + .0 2.1 ± .1 2.1 ± .2 1.8 ± .2 1.9 ± .3 2.4 ± .4 5.0 ± .5 5.8 ± .6
PR Males 10 3.2 + .2 2.2 ± .2 3.6 ± .8 2.0 ± .3 3.2 ± 1 5.0 ± 1 5.0 6.0 ± .2
ContrastComparison of Instar Group
P = T
LA vs. PR Females 0.08 0.01 0.05 0.36 0.72 0.53 — 0.98
LA vs. PR Males 0.65 0.82 0.01 0.79 0.27 0.07 — 0.77
<y>

68(Appendix C-Table 5).
Adult fecundity and longevity. There were no significant (Appendix
C-Tables 6 to 8) differences in the length of the preoviposition (LA =
3.8, PR = 3.5 days) and oviposition (LA = 4.1, PR = 4.3 days) periods or
adult longevity (LA = 10.0, PR = 10.4 days) between the two strains.
Similarly, there was no difference (Appendix C-Table 9) in the number of
egg masses per female (LA = 8.6, PR = 12.1 masses). At egg hatch, LA
larvae were 9% larger (significant P = 0.01, Appendix C-Table 10) than
PR larvae, as indicated by head capsule widths of 0.32 and 0.29 mm for
the LA and PR strains, respectively.
DISCUSSION
When larval and adult weights, length of development, and neonate
larval size of PR FAW were compared to LA FAW differences were evident.
The differences were influenced by sex, as trends were similar on both
diets. However, these differences do not extend to reproductive
parameters or adult longevity. Generally, LA larvae, pupae and adults
were heavier (Table 1) and completed development in a shorter period of
time than PR larvae (Table 2). Differences in development between the
two strains are more evident for females than for males.
Geographic isolation of Puerto Rico from the mainland (no
interbreeding between strains) and/or reproductive isolation due to
host-plant preferences may be a possible explanations for the
differences observed between strains. Pashley et al. (1985) reported
genetic differences between PR and mainland FAW populations of a
magnitude supporting reproductive isolation of the PR population. This
population was from rice, the same sample examined in the present study,

69and it is likely that both the allozymic and developmental differences
are due to underlying genetic differences in the two strains.
Development on ’Mars' foliage followed a similar pattern to that
observed on artificial diet, although growth on rice was more retarded
and development was delayed. Assuming that developmental differences
observed on artificial diet are due to the effect of diet alone and not
genetically and/or physiologically determined, then developmental
differences between strains should not be apparent when both are reared
on rice. Our results (Table 3) do not support this hypothesis. Thus,
the differences observed between strains are not due to diet effects
alone. The lack of differences in growth ratio, and in consumption and
utilization of rice foliage present additional evidence that growth and
development differences between strains are genetically controlled and
not determined by diet. The reduction in growth as indicated by lower
larval weight in both strains reared on rice when compared to pinto bean
diet is more likely to be an indication of food quality supporting
growth and not diet effect.
Piedra (1974) studied FAW development on maize Z. mays, napier
grass Pennisetum perpureum Schumach, sorghum S_. vulgare, and velvetbean
Stizolobioum deringianum Bort, and concluded that diet had little effect
on FAW adult longevity or pupal weight. Diet quality of corn was cited
as the principal factor causing a significant effect on larval
development. Recent findings (D. P. Pashley, LSU Entomology Department,
personal communication) of allozymic identity between FAW collected from
Bermuda grass in Louisiana and the PR strain from rice suggest that the
differences in our experiments may be a result of reproductive isolation
due to host-plant preference or availability.

70Differences in adult size did not extend to oviposition (under
laboratory conditions) or adult longevity but were evident at egg hatch.
Head capsule width of PR neonate larvae were 9% smaller than the LA
counterparts. These differences were detectable through larval
development as lower larval weight and increased duration of each instar
(Table 4). In Manduca sexta (L.), Trichoplusia ni (Hubner), and
Diatraea saccharalis (F.), pupation was determined by a critical body
size (threshold size) in relation to head capsule size (Jones et al.
1981; Ninjhout 1975; Roe et al. 1982). Faster growing larvae reach the
threshold size in a shorter period of time and in fewer stadia than
slower growing larvae. Although this explains why the smaller PR larvae
required longer time to complete each stadium, it does not explain why
PR males completed development in a shorter number of stadia than LA
males. Probably differences in male larval growth are the result of
genetic differences (Pashley et al. 1985) and not determined by
threshold size alone. Roe et al. (1982) indicated than in ID.
saccharalis sexual dimorphism is under genetic control and not
determined by a critical body size alone.
The results of this study indicate that differences in development
between PR and LA FAW strains are under genetic control probably due to
reproductive isolation. Because a FAW genotype similar to that
collected in Puerto Rican rice fields has been found on the mainland and
the corn genotype of the mainland has been recovered in Puerto Rico (D.
P. Pashley, personal communication) differences in chemical
communication (pheromone) and host plant preference for oviposition
might be present between the two genotypes. If the physiological
differences observed in this experiment are the result of isolation due

71to host-plant preference and geographical isolation, characterization of
various FAW populations will be necessary in host-plant resistance
programs. Additional research is needed to determine if other
morphological measurements are similar to head capsule widths, and to
correlate morphological and physiological differences to genetic
studies. Research is also needed to characterize the nature of the
barriers governing isolation under field conditions in areas of sympatry
if isolation is due to factors other than geographical. The latter study represents a greater challenge since both strains readily
interbreed under laborabory conditions (Pantoja, unpublished data).

ACKNOWLEDGEMENTS We appreciate the comments of P. Dowd, D. P. Pashley, S. S.
Quisenberry, R. Rose, and T. C. Sparks on an early draft of this paper.
72

REFERENCES CITED
Beck, S. D. and J. C. Reese. 1976. Insect-plant interactions:Nutrition and metabolism. Recent Adv. Phytochem. 10: 41-92.
Bowling, C. C. 1967. Rearing two lepidopterous pests of rice on a common artificial diet. Ann. Entomol. Soc. Am. 60: 1215-1216.
Combs, R. L. and H. W. Chambers. 1979. Comparison of toxicity ofinsecticides to larvae of a laboratory strain of fall armyworm to those collected from cotton and corn in Central Mississippi. J.Ga. Entomol. Soc. 14: 8-11.
Freund, R. J. and R. C. Littell. 1981. SAS For Linear Models. SASInstitute, North Carolina.
Jones, D., G. Jones, and B. D. Hammock. 1981. Growth parametersassociated with endocrine events in larval Trichopulsia ni (Hubner) and timing of these events with developmental markers. J. Insect Physiol. 27: 779-788.
Kogan, M. and D. Cope. 1974. Feeding and nutrition of insectsassociated with soybeans. 3. Food intake, utilization, and growth in the soybean looper, Pseudoplusia includens. Ann. Entomol. Soc. Am. 67: 66-72.
Luginbill, P. 1928. The fall armyworm USDA Tech. Bull. 34.
Mitchell, E. R. 1979. Preface to the Fall Armyworm Symposium, Fla. Entomol. 62: 81-133.
Ninjhout, H. F. 1975. A threshold size for metamorphosis in thetobacco hornworm, M. sexta (L.). Biol. Bull. Mar. Biol. Lab. Woods Hole 149: 214-225.
Pantoja, A., C. M. Smith, and J. F. Robinson. 1985. Natural control agents affecting Spodoptera frugiperda (Lepidoptera:Noctuidae) infesting rice in Puerto Rico. Fla. Entomol. 68: 488-490.
Pashley, D. P., S. J. Johnson, and A. N. Sparks. 1985. Genetic population structure of migratory noctuids: The fall armyworm (Lepidoptera:Noctuidae). Ann. Entomol. Soc. Am. 78: 756-762.
Perkins, W. D. 1979. Laboratory rearing the fall armyworm. Fla. Entomol. 62: 87-91.
Piedra, F. 1974. Effect of different forage diets on the biology ofSpodoptera frugiperda Smith (Lep. Noctuidae). Cuban J. Agric. Sci. 8: 99-103.
73

74
Roe, R. M., A. M. Hammond, Jr., and T. C. Sparks. 1982. Growth of larval Diatraea saccharalis (Lepldoptera; Pyralidae) on an artificial diet and synchronization of the last larval stadium.Ann. Entomol. Soc. Am. 75: 421-429.
Snow, J. W., W. C. Cantelo, R. L. Burton, and S. D. Hensley. 1968.Populations of fall armyworm, corn earworm, and sugarcane borer on St. Croix, U. S. Virgin Islands. J. Econ. Entomol. 61: 1757-1760.
Sparks, A. N. 1979. A review of the biology of the fall armyworm.Fla. Entomol. 62: 82-87.
Waldbauer, G. P. 1968. The consumption and utilization of food by insects. Ad. Insect Physiol. 5: 229-573.
Young, J. R. 1979. Assessing the movement of the fall armyworm (Spodoptera frugiperda) using insecticide resistance and wind patterns, pp. 344-351. JLn Movement of Highly Mobile Insects.R. C. Rabb, and G. G. Kennedy (eds.) North Carolina State Univ.

SUMMARY AND CONCLUSIONS Results of the studies reported here indicate that larval feeding
by the fall armyworm, Spodoptera frugiperda (J. E. Smith), can cause
considerable damage to rice, Oryza sativa L., seedlings. Increased
frugiperda infestation levels during the seedling stage resulted in
increased defoliation, reduced plant and panicle density, and reduced
rice yields. Yield component studies indicated that the reduced yields
on the infested plots were the result of reduced plant and panicle
density and reduced panicle size. Kernel weight and percent filled
kernels were not affected. Rice yield reduction was linearly related to
£. frugiperda larval density. Defoliation was heavier during the 1983
experiment than that in 1984, probably due to a longer feeding period
or difference in insect physiological age. Third instar larvae were
allowed to feed for 6 days during the 1983 test, while the fourth
instars used in the 1984 test only fed for 3 days. In spite of lower
larval feeding activity during the 1984 experiments, the linear model
E(y) = b accounted for a large proportion of the variance of yield
reductions during both years. Additional research is needed to
determine if the effects of larval fedding are the same for other rice
cultivars.
A technique for infesting rice seedlings grown in the greenhouse was
developed which proved to be an efficient method to screen rice
germplasm for resistance to JS. frugiperda larval feeding. Over 5,000
rice accessions were evaluated and moderate resistance to S. frugiperda
larval defoliation was detected in five plant introductions, four United
States cultivars, and three accessions of Oryza glaberrima Steud. Low
75

levels of resistance were also detected in some of the obsolete United
States rice cultivars. The United States cultivars 'Honduras',
'Newrex', and 'Chinese'; plant introductions 160842, 346833, 346853,
346840, and 346830; and (). glaberrima accessions 101800, 102554, and
369453 were found to be resistant to _S. frugiperda larval feeding.
Defoliation to the susceptible check cultivars 'Mars' and 'Saturn' in
the greenhouse was similar to that recorded in the field, indicating
that screening can be accurately conducted in the field or greenhouse.
The use of potting soil reduced handling time and allowed comparisons of
results of tests conducted over an extended period of time. The results
of this study will ultimately benefit host-plant resistance and breeding
programs in areas were S. frugiperda is considered an economically
important pest to rice production. However, further research is needed to study the nature of resistance and to incorporate resistance into
improved rice cultivars.
Important data on the general biology and development of
_S. frugiperda on rice was collected. When larval and adult weights,
length of development, and neonate larval size of j>. frugiperda larvae
collected in Puerto Rico (PR) were compared to larvae from Louisiana
(LA), differences were evident. At egg hatch, larvae of PR origin were
smaller than LA larvae. Differences in neonate larval size were also
evident at all stages of development but did not extend to adult
longevity or reproductive parameters. Larvae, pupae and adults of the
LA strain were heavier and completed development in a shorter period of
time than the PR strain. Differences in developmental parameters were not determined by diet, but were affected by sex, and may indicate some
degree of reproductive isolation. If the physiological and

77
morphological differences observed in this experiment were the result of
isolation due to host-plant preference and/or geographical isolation,
characterization of j>. frugiperda populations will be necessary in
host-plant resistance studies. These results will benefit the
development of jS. frugiperda host-plant resistance programs and
understanding j>. frugiperda migration.
The identification of parasites and pathogens recovered from
S. frugiperda larvae collected from rice fields in Puerto Rico have
contributed important information on the abundance and distribution of
natural control agents affecting S_. frugiperda larvae. Twenty-eight
parasitoid adults, larvae, and puparia were collected. Eight were identified as Lespesia sp., one as Archytas mormoratus (Townsend), and
19 were unidentifiable. The fungus Nomuraea rileyi (Farlow) Sampson
killed eight larvae and three were infected by an unidentified
microsporidium. To our knowledge this is the first report of
parasitoids and entomophagous pathogens being recovered from
S. frugiperda larvae infesting rice. However, research is needed to
evaluate the potential of using these natural enemies in conjunction
with current pest management programs and agronomic practices.

LITERATURE CITED
Altieri, M. A. 1980. Diversification of corn agroecosystems as a means of regulating fall armyworm population. Fla. Entomol. 63: 450-456.
Andrews, K. L. 1980. The whorlworm, Spodoptera frugiperda, in Central America and neighboring areas. Fla. Entomol. 63: 456-457.
Ashley, T. R. 1979. Classification and distribution of fall armyworm parasites. Fla. Entomol. 62: 114-123.
Ashley, T. R. 1982. Impact of native parasites on the fall armyworm, Spodoptera frugiperda, (Lepidoptera: Noctuidae) in South Florida and release of the exotic parasite, Eiphosoma vitticole (Hymenoptera: Ichneumonidae). Environ. Entomol. 11: 833-837.
Ashley, T. R. 1983. Growth pattern alterations in fall armyworm,Spodoptera frugiperda, larvae after parasitization by Apanteles marginiventris, Campoletis grioti, Chelenus insularis, and Eiphosoma vitticole. Fla. Entomol. 66: 260-266.
Ashley, T. R., C. S. Barfield, V. H. Waddill, and E. R. Mitchell. 1983. Parasitization of fall armyworm larvae on volunteer corn, bermudagrass, and paragrass. Fla. Entomol. 66: 267-271.
Ashley, T. R., V. H. Waddill, E. R. Mitchell, and J. Rye. 1982.Impact of native parasitoids on the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), in South Florida and release of the exotic parasite, Eiphosoma vitticole (Hymenoptera: Ichneumonidae). Environ. Entomol. 11: 833-837.
Banerjee, P. K. and P. B. Chaterjee. 1982. Pests of hill rice in West Bengal, India. Int. Rice Res. News. 7(2): 11.
Beck, S. D. and J. C. Reese. 1976. Insect plant interactions:Nutrition and metabolism. Recent Adv. Phytochem. 10: 41-92.
Bowling, C. C. 1967. Rearing two lepidopterous pest of rice on a common artificial diet. Ann. Entomol. Soc. Am. 60: 1215-1216.
Bowling, C. C. 1978. Simulated insect damage to rice: Effects of leafremoval. J. Econ. Entomol. 71: 377-378.
Brett, C. H. and R. Bastida. 1963. Resistance of sweet corn varieties to the fall armyworm, Lyphygma frugiperda. J. Econ. Entomol.56: 162-167.
Capinera, J. E. and W. J. Roltsch. 1980. Response of wheat seedling to actual and simulated migratory grasshopper defoliation. J. Econ. Entomol. 73: 258-261.
78

79Chandler, J. V., F. Abruna, J. Lozano, S. Silva, A. Rodriguez, and C.
T. Ramirez. 1977. Cultivo intensivo y perspectivas del arroz en Puerto Rico. Univ. de Puerto Rico, Estacion Experimental Agricola, Boletin 25.
Chittenden, F. H. 1900. The fall armyworm in 1899. USDA Dept.Agric. Entomol. Bull. (.S.) 23: 78-85.
Combs, R. L. and H. W. Chambers. 1979. Comparison of toxicity ofinsecticides to larvae of a laboratory strain of fall armyworm to those collected from cotton and corn in Central Mississippi.J. Ga. Entomol. Soc. 14: 8-11.
Combs, R. L. and J. R. Valerio. 1980. Oviposition by the fall armyworm on four varieties of bermudagrass. J. Ga. Entomol.Soc. 15: 164-167.
Cong Thuat, N. 1982. Rice insect pest in Vietnam. Int. Rice Res.News. 7(2): 10-11.
Dar, I. A. 1979. Occurrence of the rice ear-cutting caterpillar in the Punjab of Pakistan. Int. Rice Res. News. 4(2): 16.
Davis, F. M.. 1980. Fall armyworm plant resistance programs. Fla.Entomol. 63: 420-424.
Davis, F. M. and T. G. Oswalt. 1969. Hand inoculator for dispensing lepidopterous larvae. USDA/SEA/SR 9.
Duncan, D. B. 1955. Multiple range and multiple F tests. Biometrics 11: 1-41.
Freund, R. J. and R. C. Littell. 1981. SAS for linear models. SAS Institute, North Carolina.
Gallego, M., F. L. 1967. Lista preliminar de insectos de importancia economica y secundarios, que afectan los principales cultivos, animales domesticos y al hombre, en Colombia. Revista Facuitad Agronomia, Colombia. 65: 32-66.
Gardner, W. A. and J. R. Fuxa. 1980. Pathogens for the suppression of the fall armyworm. Fla. Entomol. 63: 439-447.
Hamm, J. J. and W. W. Hare. 1982. Application of entomopathogens in irrigation water for control of fall armyworms and corn earworms (Lepidoptera: Noctuidae) on corn. J. Econ. Entomol. 75:1074-1079.
Harrel, E. A., J. R. Young, and W. W. Hare. 1977. Insect control on late planted sweet corn. J. Econ. Entomol. 70: 129-131.

80
Jones, D., G. Jones, and B. D. Hammock. 1981. Growth parametersassociated with endocrine events in larval Trichopulsia ni (Hubner) and timing of these events with developmental markers.J. Insect Physiol. 27: 779-788.
Jung-Tsung, W. 1982. Distribution, seasonal occurrence, and natural enemies of armyworm attacking rice in China. Int. Rice. Res. News 7: 9-10.
Kogan, M. and D. Cope. 1974. Feeding and nutrition of insects associated with soybeans. 3. Food intake, utilization, and growth in the soybean looper, Pseudoplusia includens. Ann.Entomol. Soc. Am. 67: 66-72.
Kulshreshtha, J. P., M. B. Kalode, and A. Verna. 1970. Paddy cutworm (Pseudaletia separata Walker) and armyworm (Cirphis compacta Moore) as a serious pest of high yielding varieties of rice. Oryza 7: 143-145.
Leuck, D. B. 1970. The role of resistance in pearl millet in control of the fall armyworm. J. Econ. Entomol. 63: 1579-1681.
Leuck, D. B. 1972. Induced fall armyworm resistance in pearl millet.J. Econ. Entomol. 65: 1608-1611.
Leuck, D. B. and R. 0. Hammons. 1974. Nutrients and growth media:Influence on expression of resistance to the fall armyworm in the peanut. J. Econ. Entomol. 67: 564.
Leuck, D. B., C. M. Taliaferro, G. W. Burton, and M. C. Bowman. 1968a.Fall armyworm resistance in pearl millet. J. Econ. Entomol.61: 693-695.
Leuck, D. B., C. M. Taliaferro, G. W. Burton, and M. C. Bowman. 1968b.Resistance in bermudagrass to the fall armyworm. J. Econ. Entomol.61: 1321-1322.
Leuck, D. B. and J. L. Skinner. 1970. Resistance in bermuda grassaffecting control of the fall armyworm. J. Econ. Entomol.63: 1981-1982.
Leuck, D. B. and J. L. Skinner. 1971. Resistance in peanuts foliageinfluencing fall armyworm control. J. Econ. Entomol. 64: 148-150.
Lewis, W. J. and D. A. Norlund. 1980. Employment of parasitoids andpredators for fall armyworm control. Fla. Entomol. 63: 433-438.
Litsinger, J. A. 1979. Major insect pest of rainfed wetland rice in Tropical Asia. Int. Rice Res. News. 4(2): 14-15.
Luginbill, P. 1928. The fall armyworm. USDA Tech. Bull. 34.

81
Lynch, R. E., P. B. Martin, and J. W. Garner. 1980. Culturalmanipulation of coastal bermudagrass to avoid losses from the fall armyworm. Fla. Entomol. 63: 411-419.
Machado, S. S. 1978. Alguns fatores limitantes da produtividade das lavoras de arroz. Lavoura Arrozeira. 307: 52-54.
Manuwoto, S. and J. M. Scriber. 1982. Consumption and utilization of three maize genotypes by southern armyworm. J. Econ. Entomol.75: 163-167.
Matsushima, S. 1970. Crop science in rice. Fiji Pub. Co. LTD.Tokyo, Japan.
Mcllrath, W. 0., G. J. Trahan, N. E. Jodon, and C. J. Leger. 1978.Rice variety improvement research program, pp. 6-21 _In LSU Rice Exp. Stat. Annual Rept. for 1978, Crowley, LA.
McKenzie, K. S., J. N. Rutger, and M. L. Peterson. 1980. Relation of seedling vigor to semidwarfism, early maturity, and pubescence in closely related rice lines. Crop Sci. 20: 169-172.
McMillan, W. W. and K. J. Starks. 1967. Greenhouse and laboratory screening of sorghum lines for resistance to the fall armyworm larvae. J. Econ. Entomol. 60: 145-146.
Mitchel, E. R. 1978. Relationship of planting date to damage byearworm in commercial sweet corn in north central Florida. Fla. Entomol. 61: 251-255.
Mitchel, E. R. 1979. Preface to the Fall Armyworm Symposium. Fla Entomol. 62: 81-133.
Navas, D. 1966. Evaluacion del dano que ocasionan algunas plagas del arroz en Panama. La Programs cooperativo centroamericano para el mejoramiento de cultivos alimenticios, XII: 77-78.
Navas, D. 1967. Efectos de la destruccion del follaje en losrendimientos del arroz. La Programs cooperativo centroamericano para el mejoramiento de cultivos alimenticios. XIII: 85-87.
Navas, D. 1974. Fall armyworm in rice. pp. 99-106 In Proceedings Tall Timber Conference on Ecological Animal Control by Habitat Management.
Ng, Sen-Seong, F. M. Davis, and W. P. Williams. 1985. Survival,growth, and reproduction of fall armyworm (Lepidoptera:Noctuidae) as affected by resistant corn genotypes. J. Econ. Entomol.78: 967-971.
Ninjhout, H. F. 1975. A threshold size for the metamorphosis in the tobacco hornworm, Manduca sexta (L.). Biol. Bull. Mar. Biol.Lab. Woods Hole. 149: 214-225.

82
Oliver, A. D. and J. B. Chapin. 1981. Biology and illustrated keyfor the identification of twenty species of economically important noctuid pests. Louisiana Agri. Exp. Stat. Bull. 733.
Pantoja, A., C. M. Smith, and J. F. Robinson. 1985. Natural control agents affecting Spodoptera frugiperda (Lepidoptera:Noctuidae) infesting rice in Puerto Rico. Fla. Entomol. 68: 488-490.
Papel, R. K., A. K. Khatri, and B. S. Choudharg. 1981. Riceear-cutting caterpillar, an injurious pest at panicle stage. Int. Rice Res. News. 6(3): 17.
Pashley, D. P., S. J. Johnson, and A. N. Sparks. 1985. Geneticpopulation structure of migratory noctuids: The fall armyworm (Lepidoptera:Noctuidae). Ann. Entomol. Soc. Am. 78:756-762.
Pathak, M. D. 1977. Insect pests of rice. Int. Rice Res. Inst., Los Banos, Philippines.
Pathak, M. D. and G. S. Dhaliwal. 1981. Trends and strategies for rice insect problems in tropical Asia. Int. Rice Res. Inst. Res. Paper Series 64: 15.
Pencoe, N. L. and P. B. Martin. 1982. Fall armyworm (Lepidoptera: Noctuidae) larval development and adult fecundity on five grass hosts. Environ. Entomol. 11: 720-723.
Perkins, W. D. 1979. Laboratory rearing the fall armyworm. Fla. Entomol. 62: 87-91.
Piedra, F. 1974. Effects of different forage diets on the biology of Spodoptera frugiperda Smith (Lep:Noctuidae). Cuban J. Agric.Sci. 8: 99-103.
Pitre, H. N., J. E. Murooney, and D. B. Hogg. 1983. Fall armyworm (Lepidoptera: Noctuidae) oviposition: Crop preferences and eggdistribution on plants. J. Econ. Entomol. 76: 463-466.
Pophali, D. J., P. D. Deshmukh, U. K. Kaushik, S. K. Shrivastara, S. L. Patidar, and G. A. Sangrade. 1980. Assessment of yield losses caused by earhead-cutting caterpillar Mythimna separata Walker. Int. Rice Res. News. 5(4): 8-9.
Quisenberry, S. S. and H. K. Wilson. 1985. Consumption and utilization of bermuda grass by fall armyworm (Lepidoptera:Noctuidae) larvae. J. Econ. Entomol. 78: 820-824.
Rice, S. E., A. A. Grigarick, and M. 0. Way. 1982a. Effect of leaf and panicle feeding by armyworm (Lepidoptera: Noctuidae) larvae on rice grain yield. J. Econ. Entomol. 75: 593-595.

Rice, S. E., A. A. Grigarick, and M. 0. Way. 1982b. Relationship of larval density and instars of Pseudaletia unipuncta to rice leaf feeding. Environ. Entomol. 11: 648-651.
Roe, R. M., A. M. Hammond, and T. C. Sparks. 1982. Growth of larval Diatraea saccharalis (Lepidoptera:Pyralidae) on an artificial diet and synchronization of the last larval stadium. Ann. Entomol. Soc Am. 75: 421-429.
Snow, W. W., W. C. Cantelo, R. L. Burton, and S. D. Hensley. 1968.Populations of fall armyworm, corn earworm and sugarcane borer on St. Croix, U.S. Virgin Islands. J. Econ. Entomol. 61: 1757-1760.
Soejitno, J. and G. van Vreden. 1976. Varietal screening forresistance to the armyworm, Spodoptera mauritia, in rice. pp. 166-167 Ijl Agric. Coop. Indonesia Netherlands Res. Rept. 1968-1976
Sparks, A. N. 1979. A review of the biology of the fall armyworm. Fla Entomol. 62: 82-87.
Waldbauer, G. P. 1968. The consumption and utilization of food by insects. Ad. Insect Physiol. 5: 229-573.
Wilde, G. and R. Apostol. 1983. Armyworm (Lepidoptera: Noctuidae) resistance in rice. Environ. Entomol. 12: 376-379.
Williams, W. P., F. M. Davis, and B. R. Wiseman. 1983. Fall armyworm resistance in corn and its suppression of larval survival and growth. Agron. J. 75: 831-832.
Williams, W. P., P. M. Buckley, and F. M. Davis. 1985. Larval growth and behavior of the fall armyworm (Lepidoptera:Noctuidae) on callus initiated from susceptible and resistant corn hybryds.J. Econ. Entomol. 78: 951-954.
Wiseman, B. R. and F. M. Davis. 1979. Plant resistance to the fall armyworm. Fla. Entomol. 62: 123-130.
Wiseman, B. R. and L. Gourley. 1982. Fall armyworm(Lepidoptera:Noctuidae): Infestation procedures and sorghumresistance evaluations. J. Econ. Entomol. 75: 1048-1051.
Wiseman, B. R., R. C. Gueldner, and R. E. Lynch. 1982. Resistancein common centipedegrass to the fall armyworm. J. Econ. Entomol. 75: 245-247.
Wiseman, B. R., R. H. Painter, and C. E. Wasson. 1966. Detecting corn seedling differences in the greenhouse by visual classification of damage by the fall armyworm. J. Econ. Entomol. 59: 1211-1214.
Yoshida* S. 1981. Fundamentals of rice crop science. Int. Rice Res. Inst., Los Banos, Philippines.

Young, J. R. 1979a. Assessing the movement of the fall armyworm, Spodoptera frugiperda, using insecticide resistance and wind patterns, pp. 344-351. ^n Movements of Highly Mobile Insects. R. C. Rabb, and G. G. Kennedy (eds.) North Carolina State Univ.
Young, J. R. 1979b. Fall armyworm: Control with insecticides. Fla.Entomol. 62: 130-133.

APPENDIX A

86Table 1. Analysis of variance for defoliation rate on selected rice cultivars by Spodoptera frugiperda larvae in a no choice test. Crowley, LA 1985.
Source of variation df Mean Square F P
Error 33 3.08
Replicate 3 8.16 2.65 0.0649Cultivar 12 8.11 2.63 0.0137

87
Table 2. Analysis of variance for percent larval population on selected rice cultivars by Spodoptera frugiperda larvae in a choice test. Baton Rouge, LA 1985.
Source of variation df Mean Square F P
Error 44 0.0611
Replicate 4 0.0543 0.89 0.4784
Cultivar 11 0.1940 3.18 0.0030
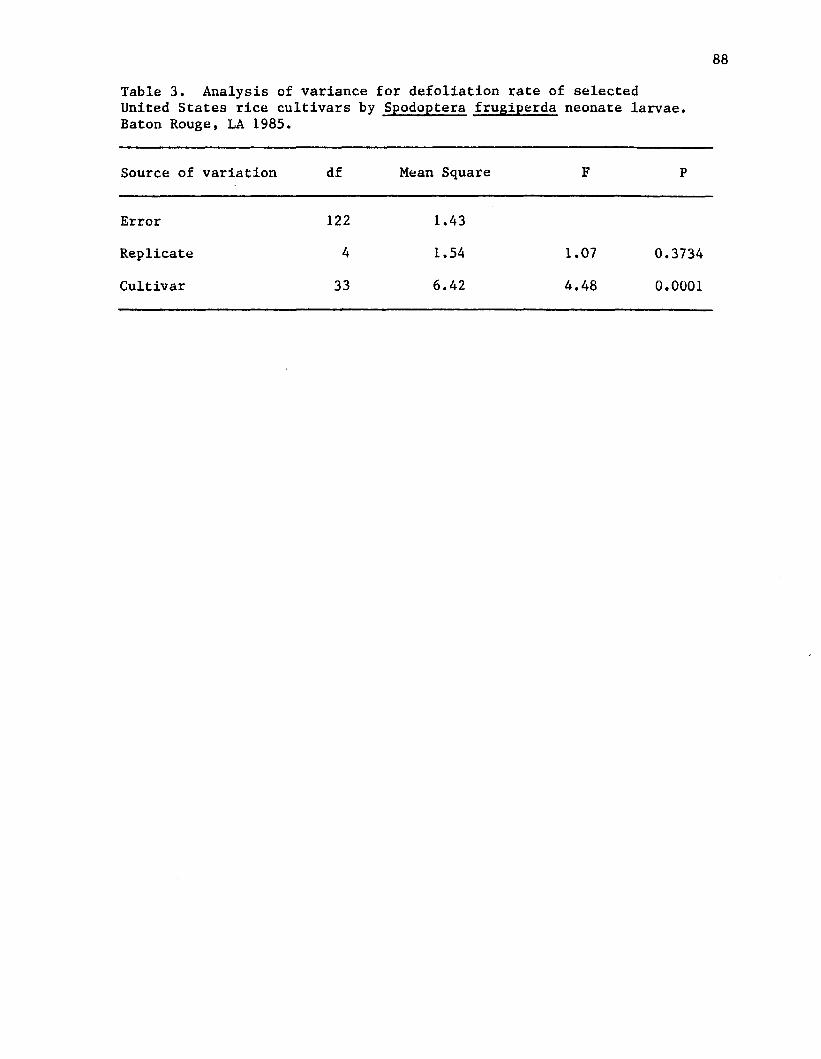
88Table 3. Analysis of variance for defoliation rate of selected United States rice cultivars by Spodoptera frugiperda neonate larvae. Baton Rouge, LA 1985.
Source of variation df Mean Square F P
Error 122 1.43
Replicate 4 1.54 1.07 0.3734
Cultivar 33 6.42 4.48 0.0001

89
Table 4. Analysis of variance for defoliation rate of selected Oryza species by Spodoptera frugiperda neonate larvae. Baton Rouge, LA 1985,
Source of variation df Mean Square F P
Error 24 2.32
Replicate 6 2.84 1.23 0.3282
Cultivar 6 5.67 2.73 0.0364

APPENDIX B

91
Table 1. Analysis of variance for defoliation percentage by Spodopterafrugiperda third instar larvae on 'Mars’ rice at varying density levels.Baton Rouge, LA 1983.
Source of variation df Mean Square F P
Error 15 237.70
Replicate 3 388.26 1.63 0.2238
Treatment 5 7522.74 31.65 0.0001

92
Table 2. Analysis of variance for 'Mars' rice plant density as affectedby Spodoptera frugiperda third instar larvae at varying density levels.Baton Rouge, LA 1983.
Source of variation df Mean Square F P
Error 14 8.17
Replicate 3 18.67 2.29 0.1235
Treatment 5 238.25 13.84 0.0001

93
Table 3. Analysis of variance for 'Mars’ rice panicle density asaffected by Spodoptera frugiperda third instar larvae at varying densitylevels. Baton Rouge, LA 1983.
Source of variation df Mean Square F P
Error 14 173.18
Replicate 3 466.08 2.69 0.0863Treatment 5 2396.90 13.84 0.0001

94
Table 4. Analysis of variance for 'Mars' rice gross yield as affectedby Spodoptera frugiperda third instar larvae at varying density levels.Baton Rouge, LA 1983.
Source of variation df Mean Square F P
Error 14 0.0020
Replicate 3 0.0046 2.23 0.1299Treatment 5 0.0184 9.00 0.0005

95
Table 5. Analysis of variance for weight of 500 kernels of 'Mars' riceaffected by Spodoptera frugiperda third instar larvae at varing densitylevels. Baton Rouge, LA 1983.
Source of variation df Mean Square F P
Error 6 1.0461
Replicate 3 2.2093 2.11 0.2001
Treatment 3 0.8978 0.86 0.5117

96
Table 6. Analysis of variance for panicle weight of 'Mars' riceaffected by Spodoptera frugiperda third instar larvae at varying densitylevels. Baton Rouge, LA 1983.
Source of variation df Mean Square F P
Error 22 0.0244Replicate 3 0.0270 1.11 0.3799
Treatment 5 0.0723 2.96 0.0498

97
Table 7. Analysis of variance for percent fertile kernels of 'Mars'rice affected by Spodoptera frugiperda third instar larvae at varyingdensity levels. Baton Rouge, LA 1983.
Source of variation df Mean Square F P
Error 6 61.00
Replicate 3 30.85 0.51 0.6924
Treatment 3 64.34 1.05 0.4347

98
Table 8. Analysis of variance for biomass index of 'Mars' rice affectedby Spodoptera frugiperda fourth instar larvae at varying density levels.Baton Rouge, LA 1984.
Source of variation df Mean Square F P
Error 15 39264.72
Replicate 3 44415.74 1.13 0.3681
Treatment 5 223016.98 5.68 0.0039

99
Table 9. Analysis of variance for panicle density of 'Mars' riceaffected by Spodoptera frugiperda fourth instar larvae at varyingdensity levels. Baton Rouge, LA 1984.
Source of variation df Mean Square F P
Error 15 401.18
Replicate 3 827.71 2.18 0.1334
Treatment 5 334.24 0.83 0.5461

100Table 10. Analysis of variance for panicle weight of 'Mars' riceaffected by Spodoptera frugiperda fourth instar larvae at varyingdensity levels. Baton Rouge, LA 1984.
Source of variation df Mean Square F P
Error 15 0.0485
Replicate 3 0.3424 7.06 0.0035
Treatment 5 0.0970 2.00 0.1369

101Table 11. Analysis of variance for mature plant height of 'Mars' riceaffected by Spodoptera frugiperda fourth instar larvae at varyingdensity levels. Baton Rouge, LA 1984.
Source of variation df Mean Square F P
Error 15 13.79
Replicate 3 10.15 0.74 0.5464
Treatment 5 47.88 3.47 0.0277

102Table 12. Analysis of variance for weight of 1000 kernels of 'Mars'rice affected by Spodoptera frugiperda fourth instar larvae at varyingdensity levels. Baton Rouge, LA 1984.
Source of variation df Mean Square F P
Error 15 0.9768
Replicate 3 1.6571 1.70 0.2105
Treatment 5 1.0036 1.03 0.4366

103Table 13. Analysis of variance for adjusted yield of 'Mars' riceaffected by Spodoptera frugiperda fourth instar larvae at varyingdensity levels. Baton Rouge, LA 1984.
Source of variation df Mean Square F P
Error 14 584434.53
Replicate 3 202862.88 0.35 0.7918
Treatment 5 1552049.10 2.66 0.0686

104
Table 14. Analysis of variance for percent filled kernels of 'Mars'rice affected by Spodoptera frugiperda fourth instar larvae at varyingdensity levels. Baton Rouge, LA 1984.
Source of variation df Mean Square F P
Error 15 0.0010Replicate 3 0.0015 1.49 0.2565
Treatment 5 0.0023 2.25 0.1026

APPENDIX C
105

106
Table 1. Analysis of variance for consumption index by Spodopterafrugiperda third instar larvae from Louisiana and Puerto Rico fed 'Mars’rice foliage. Baton Rouge, LA 1984.
Source of variation df Mean Square F P
Error
Strain
45
12.37
5.91 2.50 0.1209

107
Table 2. Analysis of variance for growth ratio of Spodoptera frugiperdathird instar larvae from Louisiana and Puerto Rico fed 'Mars1 ricefoliage. Baton Rouge, LA 1984.
Source of variation df Mean Square F P
Error 45 0.0057
Strain 1 0.0183 3.21 0.0798

108
Table 3. Analysis of variance for approximate digestibility bySpodoptera frugiperda third instar larvae from Louisiana and Puerto Ricofed 'Mars' rice foliage. Baton Rouge, LA 1984.
Source of variation df Mean Square F P
ErrorStrain
451
4.7738.11 0.45 0.5060

109
Table 4. Analysis of variance for efficiency of conversion of ingestedfood by Spodoptera frugiperda third instar larvae from Louisiana andPuerto Rico fed 'Mars' rice foliage. Baton Rouge, LA 1984.
Source of variation df Mean Square F P
Error
Strain
451
51.18
6.51 0.13 0.7229
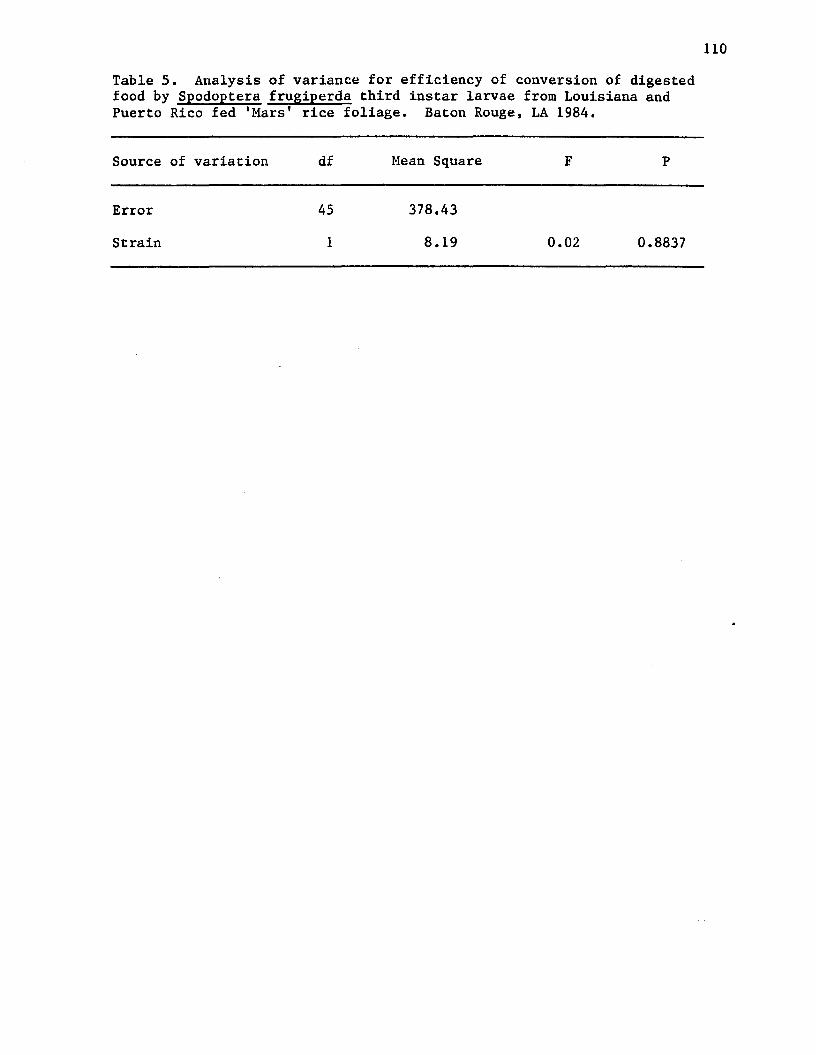
110Table 5. Analysis of variance for efficiency of conversion of digestedfood by Spodoptera frugiperda third instar larvae from Louisiana andPuerto Rico fed 'Mars’ rice foliage. Baton Rouge, LA 1984.
Source of variation df Mean Square F P
Error
Strain
45 378.43
1 8.19 0.02 0.8837

ill
Table 6. Analysis of variance for preovipositional days of Spodopterafrugiperda adults from Louisiana and Puerto Rico. Baton Rouge, LA 1984.
Source of variation df Mean Square F P
Error 25 4.1371
Run 1 0.1429 0.03 0.8541
Strain 1 0.1429 0.03 0.8541
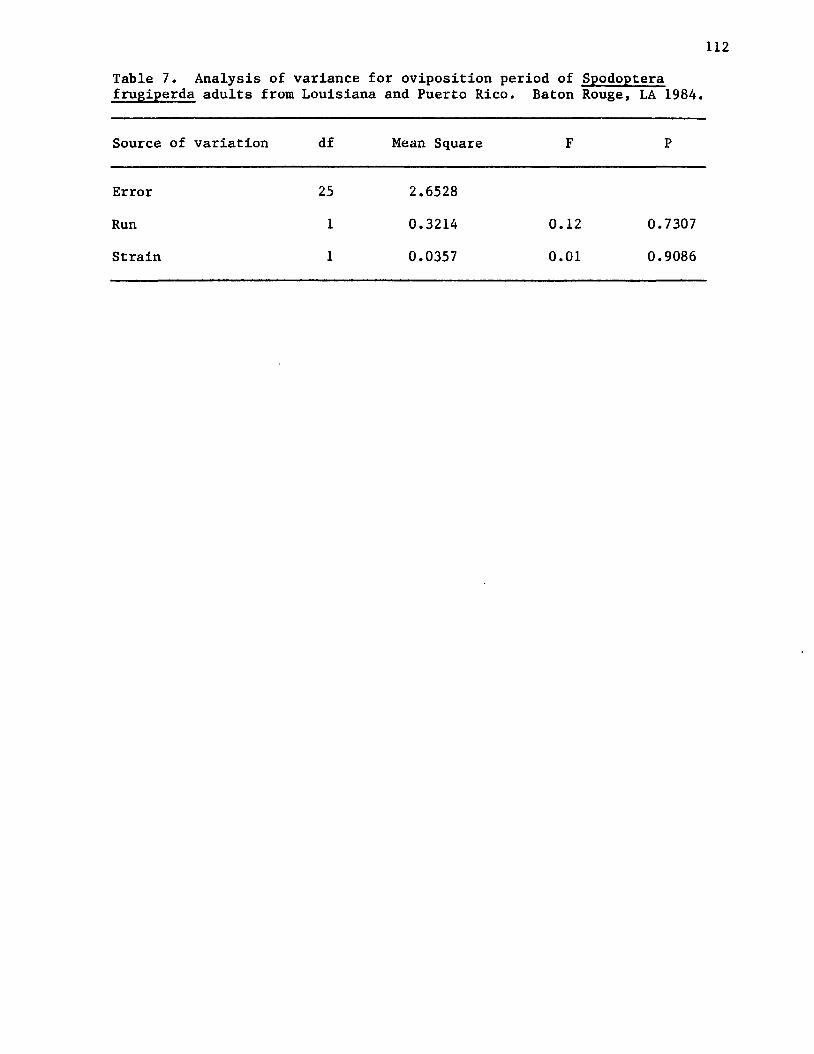
112Table 7. Analysis of variance for oviposition period of Spodopterafrugiperda adults from Louisiana and Puerto Rico. Baton Rouge, LA 1984.
Source of variation df Mean Square F P
Error 25 2.6528
Run 1 0.3214 0.12 0.7307
Strain 1 0.0357 0.01 0.9086
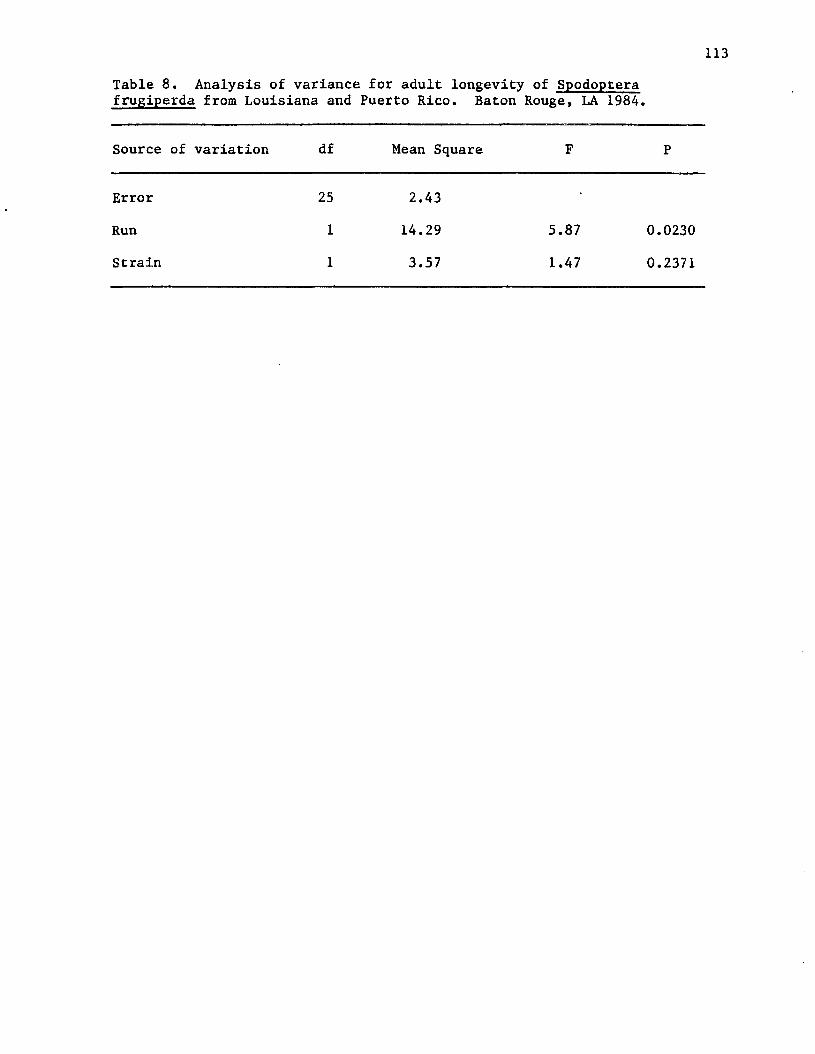
113
Table 8. Analysis of variance for adult longevity of Spodopterafrugiperda from Louisiana and Puerto Rico. Baton Rouge, LA 1984.
Source of variation df Mean Square F P
Error 25 2.43
Run 1 14.29 5.87 0.0230
Strain 1 3.57 1.47 0.2371

114
Table 9. Analysis of variance for number of egg masses deposited by Spodoptera frugiperda adults from Louisiana and Puerto Rico. Baton Rouge, LA 1984.
Source of variation df Mean Square
Error 25 28.16
Run 1 41.29 1.47 0.2373Strain 1 69.14 2.39 0.1296

115Table 10. Analysis of variance for head capsule width of Spodoptera frugiperda neonate larvae from Louisiana and Puerto Rico. Baton Rouge, LA 1984.
Source of variation df Mean Square F P
Error 147 0.00024
Run 1 0.01066 83.30 0.0001
Strain 1 0.02509 106.32 0.0001

VITA
Alberto Pantoja-Lopez was born on March 10, 1956, in Vega Baja,
Puerto Rico. He graduated from Lino Padron Rivera High School in May
1974.
In August 1974, he entered the University of Puerto Rico at
Mayaguez, and graduated in July 1978, with a Bachelor of Science in
Agronomy.
From August 1978 to August 1980, he was employed by the Puerto Rico
Department of Agriculture in a pesticides enforcement program sponsored
by the United States Environmental Protection Agency (EPA).
He entered the Louisiana State University Graduate School in August
1980, and graduated in December 1982, with a Master of Science degree in
Agronomy. He is currently a candidate for the Doctor of Philosophy
degree in Entomology.
116

DOCTORAL EXAMINATION AND DISSERTATION REPORT
Candidate: Alberto Pantoja-Lopez
Major Field: Entomology
Title of Dissertation: Biology, Economic Injury, and Plant Resistance Studies With the Fall Armyworm, Spodoptera frugiperda (J. E. Smith), on Rice, Orzya sativa L.
Approved:n
M ajor Professor and Chairman
DiL-UDean of the Gradue 1
EXAMINING COMMITTEE:
JL/H-C. hlu&lx
Date of Examination:
November 21, 1985





























