Embed Size (px)
Citation preview

Biology Chapter 16
NUCLEIC ACIDS and
INHERITANCE
����������
[email protected] 3366-4165
INTRODUCTIONn In this chapter we will shift our attention to molecular
geneticsn The study of DNA structure and function at the molecular
leveln Recent dramatic advances in techniques and approaches
have greatly expanded our understanding of molecular geneticsn And also of transmission and population genetics
n To a large extent, our knowledge of genetics comes from our knowledge of DNA and RNA
2

n To fulfill its role, the genetic material must meet several criterian 1. Information: It must contain the information necessary
to make an entire organismn 2. Transmission: It must be passed from parent to
offspringn 3. Replication: It must be copied
n In order to be passed from parent to offspring
n 4. Variation: It must be capable of changesn To account for the known phenotypic variation in each species
The Search for the Genetic Material
3
n The data of many geneticists, including Mendel, were consistent with these four propertiesn However, the chemical nature of the genetic material cannot
be identified solely by genetic crosses
!
n Indeed, the identification of DNA as the genetic material involved a series of outstanding experimental approachesn These will be examined next
The Search for the Genetic Material
4

n Griffith studied a bacterium (Diplococcus pneumoniae����) now known as Streptococcus pneumoniae ( ��)
n S. pneumoniae comes in two strains (���
n S --> Smoothn Secrete a polysaccharide capsule (��)
n Protects bacterium from the attack of immune system of animals
n Produce smooth colonies (�) on solid media (agar plate)
!
n R --> Roughn Unable to secrete a capsulen Produce colonies with a rough appearance
Frederick Griffith Experiments with Streptococcus pneumoniae
5
6

Can the genetic trait of pathogenicity be transferred between bacteria?
7
Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Figure 16.2 Can the genetic trait of pathogenicity be transferred between bacteria?
Bacteria of the “S” (smooth) strain of Streptococcus pneumoniae are pathogenic because they have a capsule that protects them from an animal’s defense system. Bacteria of the “R” (rough) strain lack a capsule and are nonpathogenic. Frederick Griffith injected mice with the two strains as shown below:
Griffith concluded that the living R bacteria had been transformed into pathogenic S bacteria by an unknown, heritable substance from the dead S cells.
EXPERIMENT
RESULT
CONCLUSION
Living S (control) cells
Living R (control) cells
Heat-killed (control) S cells
Mixture of heat-killed S cells and living R cells
Mouse dies Mouse healthy Mouse healthy Mouse dies
Living S cells are found in blood sample.
8
CaCl2

n In 1928, Griffith conducted experiments using two strains of S. pneumoniae: type S and type R
!
n 1. Inject mouse with live type S bacterian Mouse diedn Type IIIS bacteria recovered from the mouse’s blood
n 2. Inject mouse with live type R bacteria n Mouse survivedn No living bacteria isolated from the mouse’s blood
n 3. Inject mouse with heat-killed type S bacterian Mouse survivedn No living bacteria isolated from the mouse’s blood
n 4. Inject mouse with live type R + heat-killed type S cellsn Mouse diedn Type IIIS bacteria recovered from the mouse’s blood
9
n Griffith concluded that something from the dead type S was transforming type R into type Sn He called this process transformation!
n The substance that allowed this to happen was termed the transformation principle (����)
n Griffith did not know what it was
!
n The nature of the transforming principle was determined using experimental approaches that incorporated various biochemical techniques
10
qqqqqq
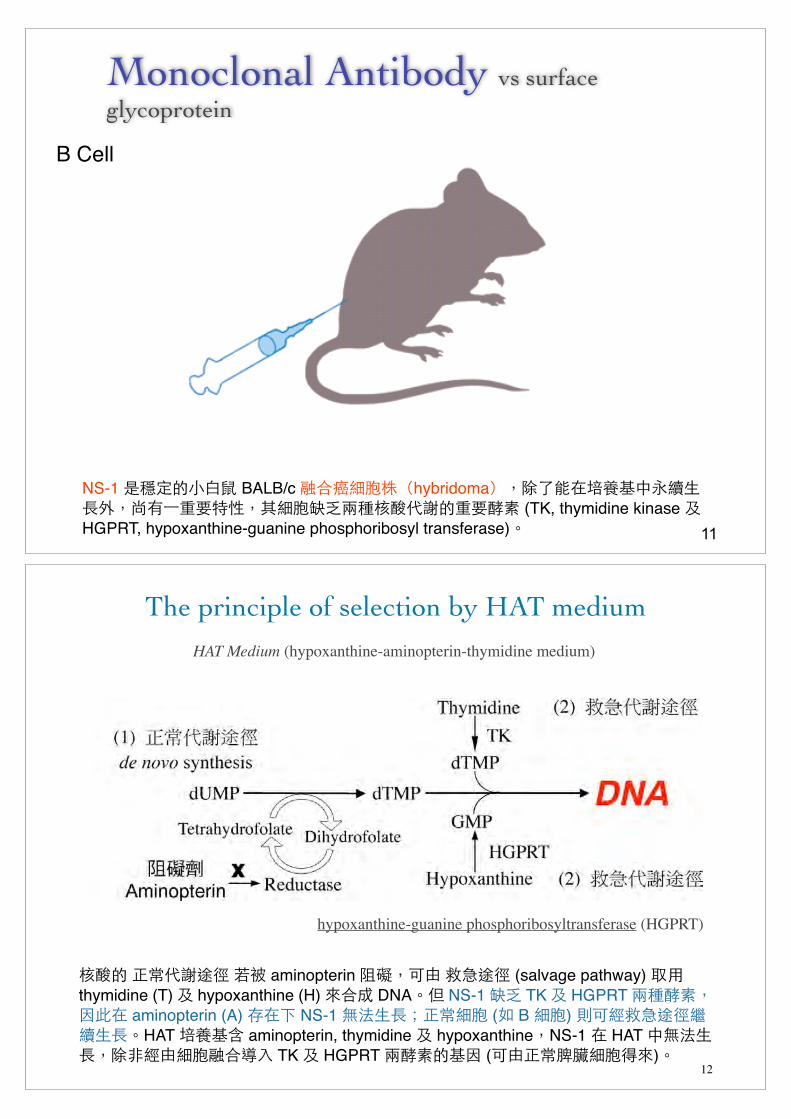
Monoclonal Antibody vs surface glycoprotein
11
B Cell
NS-1 �B���!J BALB/c G&L5FPThybridomaUS8��D0:/-��$S6����)�S�5F=I��A3+*���Q; (TK, thymidine kinase 2 HGPRT, hypoxanthine-guanine phosphoribosyl transferase)R
The principle of selection by HAT medium
12
HAT Medium (hypoxanthine-aminopterin-thymidine medium)
hypoxanthine-guanine phosphoribosyltransferase (HGPRT)
A3� �(+*CN <� aminopterin KMS . >@CN (salvage pathway) � thymidine (T) 2 hypoxanthine (H) �&� DNAR� NS-1 =I TK 2 HGPRT ��Q;S�'� aminopterin (A) 4�� NS-1 �%��V�(5F (� B 5F) 9 �>@CN1-��RHAT D0:? aminopterin, thymidine 2 hypoxanthineSNS-1 � HAT �%��S8,�.5FG&7# TK 2 HGPRT �Q;�:� ( .�(EH5F��)R

n Avery, MacLeod and McCarty realized that Griffith’s observations could be used to identify the genetic material
n They carried out their experiments in the 1940sn At that time, it was known that DNA, RNA, proteins and
carbohydrates are major constituents of living cells
n They prepared cell extracts from type S cells containing each of these macromoleculesn Only the extract that contained purified DNA was
able to convert type R into type S
The Experiments of Avery, MacLeod and McCarty
13
Figuren Avery et al also conducted the following experiments
n To further verify that DNA, and not a contaminant (RNA or protein), is the genetic material
14
���� immunoprecipitation
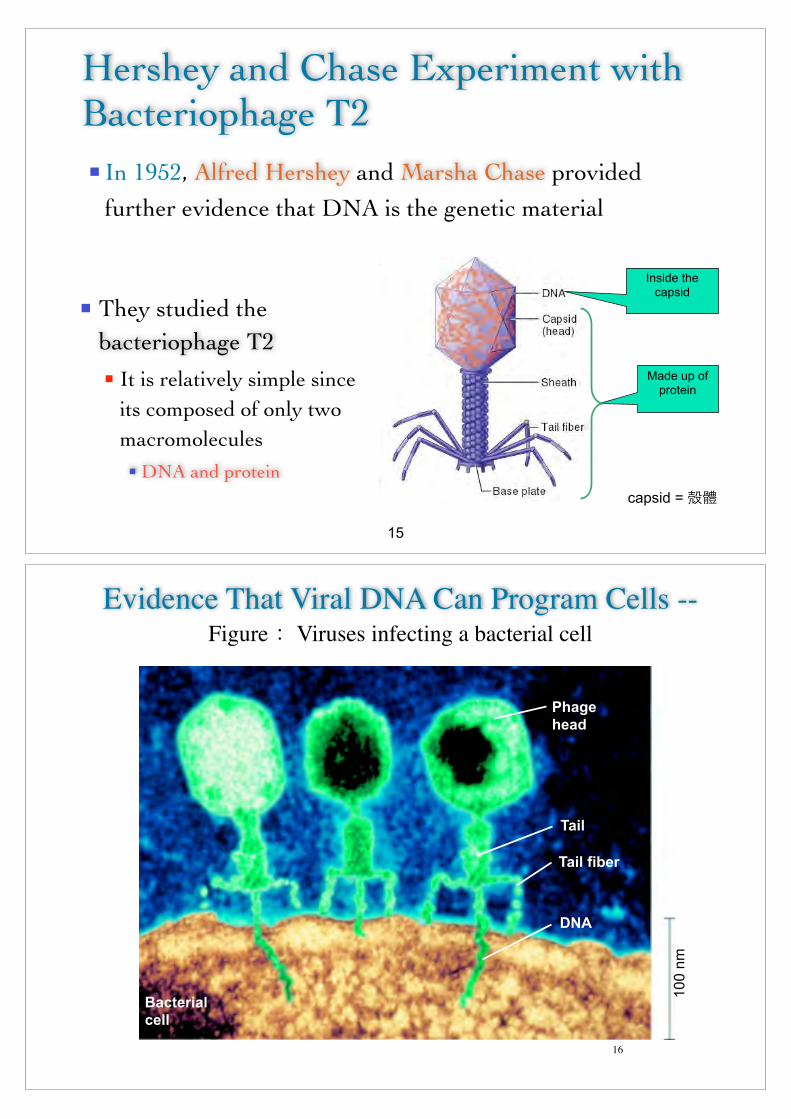
n In 1952, Alfred Hershey and Marsha Chase provided further evidence that DNA is the genetic material
Hershey and Chase Experiment with Bacteriophage T2
n They studied the bacteriophage T2n It is relatively simple since
its composed of only two macromoleculesn DNA and protein
Made up of protein
Inside the capsid
15
capsid = O"
Phage head
Tail
Tail fiber
DNA
Bacterial cell
100
nm
Evidence That Viral DNA Can Program Cells -- Figure Viruses infecting a bacterial cell
16

EM slide
17
Cryo-electron microscopy � ����
18
Cryo-electron microscopy (cryo-EM), or electron cryomicroscopy, is a form of transmission electron microscopy (EM) where the sample is studied at cryogenic temperatures (generally liquid nitrogen temperatures). Cryo-EM is developing popularity in structural biology.The popularity of cryoelectron microscopy stems from the fact that it allows the observation of specimens that have not been stained or fixed in any way, showing them in their native environment, in contrast to X-ray crystallography, which generally requires placing the samples in non-physiological environments, which can occasionally lead to functionally irrelevant conformational changes. In practice, the resolution of cryo-electron microscopy maps is not high enough to allow for unambiguous model construction on the basis of EM maps only, and models obtained by protein crystallography are used to interpret the cryo-EM maps. However, the resolution of cryo-EM maps is improving steadily, and some virus structures obtained by cryo-EM are already at a resolution that can be interpreted in terms of an atomic model.A version of electron cryomicroscopy is cryo-electron tomography (CET) where a 3D reconstruction of a sample is created from tilted 2D images, again at cryogenic temperatures (either liquid nitrogen or helium).
�

Cryo-EM
19
Hershey
20

Figure
Life cycle of the T2 bacteriophage
21
n The Hershey and Chase experiment can be summarized as such:
n Used radioisotopes to distinguish DNA from proteins n 32P labels DNA specificallyn 35S labels protein specifically (methionine ����)
n Radioactively-labeled phages were used to infect nonradioactive Escherichia coli cells
n After allowing sufficient time for infection to proceed, the residual phage particles were sheared off the cells n => Phage hosts and E. coli cells were separated
n Radioactivity was monitored using a scintillation counter
22
qqqqqq
������

The Hypothesis
n Only the genetic material of the phage is injected into the bacteriumn Isotope labeling will reveal if it is DNA or protein
Testing the Hypothesisn Refer to Figure 9.6
23
Figure24

������ 25
fluorophore 螢光團
Emission
Spectrum
scintillation counter ������
Cocktail

The Data
29
�������� ������
Interpreting the DataMost of the 35S was
found in the supernatantBut only a small
percentage of 32P
n These results suggest that DNA is injected into the bacterial cytoplasm during infectionn This is the expected result if DNA is the genetic material 30
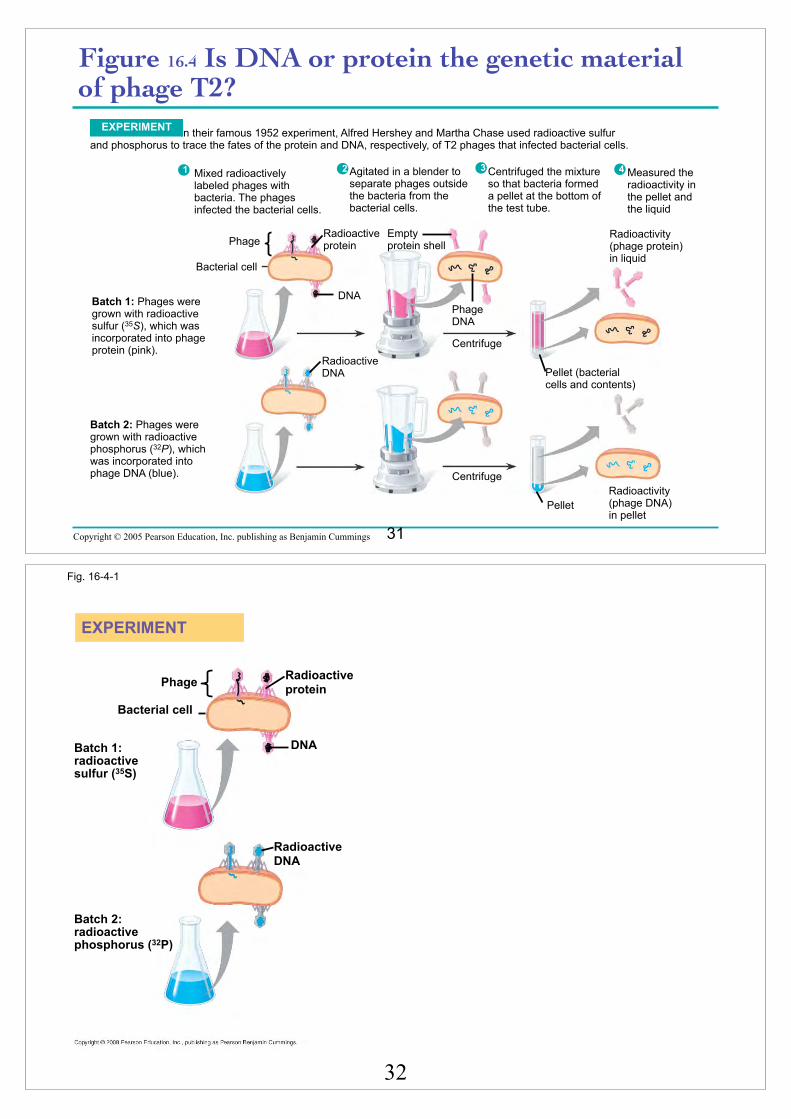
Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Figure 16.4 Is DNA or protein the genetic material of phage T2?
In their famous 1952 experiment, Alfred Hershey and Martha Chase used radioactive sulfur and phosphorus to trace the fates of the protein and DNA, respectively, of T2 phages that infected bacterial cells.
EXPERIMENT
Radioactivity (phage protein) in liquid
Phage
Bacterial cell
Radioactive protein
Empty protein shell
Phage DNA
DNA
Centrifuge
Pellet (bacterial cells and contents)
Radioactive DNA
Centrifuge
PelletRadioactivity (phage DNA) in pellet
Batch 1: Phages were grown with radioactive sulfur (35S), which was incorporated into phage protein (pink).
Batch 2: Phages were grown with radioactive phosphorus (32P), which was incorporated into phage DNA (blue).
1 2 3 4Agitated in a blender to separate phages outside the bacteria from the bacterial cells.
Mixed radioactively labeled phages with bacteria. The phages infected the bacterial cells.
Centrifuged the mixture so that bacteria formed a pellet at the bottom of the test tube.
Measured the radioactivity in the pellet and the liquid
31
Fig. 16-4-1
EXPERIMENT
Phage
DNA
Bacterial cell
Radioactive protein
Radioactive DNA
Batch 1: radioactive sulfur (35S)
Batch 2: radioactive phosphorus (32P)
32

Fig. 16-4-2
EXPERIMENT
Phage
DNA
Bacterial cell
Radioactive protein
Radioactive DNA
Batch 1: radioactive sulfur (35S)
Batch 2: radioactive phosphorus (32P)
Empty protein shell
Phage DNA
33
Fig. 16-4-3
EXPERIMENT
Phage
DNA
Bacterial cell
Radioactive protein
Radioactive DNA
Batch 1: radioactive sulfur (35S)
Batch 2: radioactive phosphorus (32P)
Empty protein shell
Phage DNA
Centrifuge
Centrifuge
Pellet
Pellet (bacterial cells and contents)
Radioactivity (phage protein) in liquid
Radioactivity (phage DNA) in pellet
34

n In 1956, A. Gierer and G. Schramm isolated RNA from
the tobacco mosaic virus (TMV), a plant virus
n Purified RNA caused the same lesions as intact TMV viruses n Therefore, the viral genome is composed of RNA
!
n Since that time, many RNA viruses have been found
RNA Functions as the Genetic Material in Some Viruses
35
DNA RNA Protein
Identify RNA also as a genetic material
36
Life cycle of Retrovirus
DNA RNA Protein
DNA and RNA are genetic material
In 1928, Griffith conducted experiments using two strains of S. pneumoniae: type IIIS and type IIR
In the 1940s, Avery, MacLeod and McCarty, further verify that DNA, and not a contaminant (RNA or protein), is the genetic material by transfroming S. pneumoniae
In 1952, Alfred Hershey and Marsha Chase provided further evidence that DNA is the genetic material by using Bacteriophage T2

n The nucleotide is the repeating structural unit of DNA (�uj�jS) and RNA (j�jS)
n It has three componentsn A phosphate groupn A pentose sugar �Dt��n A nitrogenous base �g��^��^�!
n Refer to Figure
Nucleotides 核 苷 酸
39
�0c����|L.��t)G=>�"�c�x��zK�~U((�!�c��LX�WH��\�"rsc�q�c((�
Figure
Nucleotides are the building blocks of nucleic acids
40
��
� �
1
23
4
5

Figure The structure of nucleotides found in (a) DNA and (b) RNA
A, G, C or T
n These atoms are found within individual nucleotidesn However, they are removed when nucleotides join together to make
strands of DNA or RNA
A, G, C or U
41
核 苷 酸去 氧 核 苷 酸
n Base + sugar → nucleoside n Example
n Adenine + ribose = Adenosine 腺核苷 n Adenine + deoxyribose = Deoxyadenosine 去氧腺核苷
!
n Base + sugar + phosphate(s) → nucleotiden Example
n Adenosine monophosphate (AMP) 腺核苷單磷酸 n Adenosine diphosphate (ADP) 腺核苷雙磷酸 n Adenosine triphosphate (ATP) 腺核苷參磷酸 !
n Refer to Figure
42
核 苷 或 核 醣
核 苷 酸
qqqqqq

Figure
Base always attached here
Phosphates are attached there
Nucleoside and nucleotide
Nucleoside triphosphate
43
qqqqqq
n Nucleotides are covalently linked together by phosphodiester bonds (磷脂鍵)n A phosphate connects the 5’ carbon of one nucleotide to the 3’
carbon of another!
n Therefore the strand has directionalityn 5’ to 3’ orientation (5’ to 3’ directionality) of the
nucleotides!
n The phosphates and sugar molecules form the backbone of the nucleic acid (jS) strandn The bases project from the backbone
44
qqqqqq

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Figure 16.5 The structure of a DNA strandSugar-phosphate
backboneNitrogenous
bases
5ʹ endO–
O P O CH2
5ʹ
4ʹO–
HH
OH
HH
3ʹ
1ʹH O
CH3
N
O
NH
Thymine (T)
O
O P O
O–
CH2
HH
OH
HH
HN
N
N
H
NH
H
Adenine (A)
O
O P O
O–
CH2
HH
OH
HH
HH H
HN
NN
OCytosine (C)
O
O P O CH2
5ʹ
4ʹO–
H
O
H
H3ʹ
1ʹ
OH2ʹ
H
N
NN H
ON
N HH
H H
Sugar (deoxyribose) 3ʹ end
Phosphate
Guanine (G)
DNA nucleotide
2ʹ
N
45
Figure
Nucleotides are linked together to form a strand
46

n In 1953, James Watson and Francis Crick discovered the double helical structure of DNA
!
n The scientific framework for their breakthrough was provided by other scientists includingn Linus Paulingn Rosalind Franklin and Maurice Wilkinsn Erwin Chargaff
Building a Structural Model of DNA: A Few Key Events Led to the Discovery of the Structure of DNA
47
n In the early 1950s, he proposed that regions of protein can fold into a secondary structuren α-helix
Linus Pauling
Figure48
���*e7�Electronegativity��Y{2J�Resonance��Fm2J�Valence bond
theory��i�}n� Molecular Orbital
Theory��T5I1E<o�Secondary
structure���b��QA:v�%NCh8�&��Quantum mechanics�Quantum
physics or Quantum theory��
���� Molecular Evolution

(1950)
49
�pa·_`·�b���G���8�G��<o�>��My ��1954�/�G�m%+�9-4 l]`G�B�1963
�/P�jk��+V6�'�3 1962�7�l]`�@B���3 �,l]`BZ�#� ����f���;$��OR�3 l]`B�3B��
n She worked in the same laboratory as Maurice Wilkins!
n She used X-ray diffraction (X光繞射) to study wet fibers of DNA
Rosalind Franklin
The diffraction pattern is interpreted (using mathematical theory)
This can ultimately provide information concerning the structure of the molecule
50

X-ray diffraction
n She made marked advances in X-ray diffraction techniques with DNA!
n The diffraction pattern she obtained suggested several structural features of DNA!
n Helicaln More than one strandn 10 base pairs per complete turn
Rosalind Franklin
52

n Chargaff pioneered many of the biochemical techniques
for the isolation, purification and measurement of
nucleic acids from living cells
!
n It was already known then that DNA contained the four
bases: A, G, C and T
Erwin Chargaff’s Experiment
53
54

The Hypothesis
n An analysis of the base composition of DNA in
different species may reveal important features
about the structure of DNA
n ……..Evolutionary Conservation
Testing the Hypothesis
n Refer to Figure
55
Figure
An analysis of base composition among different DNA samples
56
�w chloroformT5� protease
6M HCl
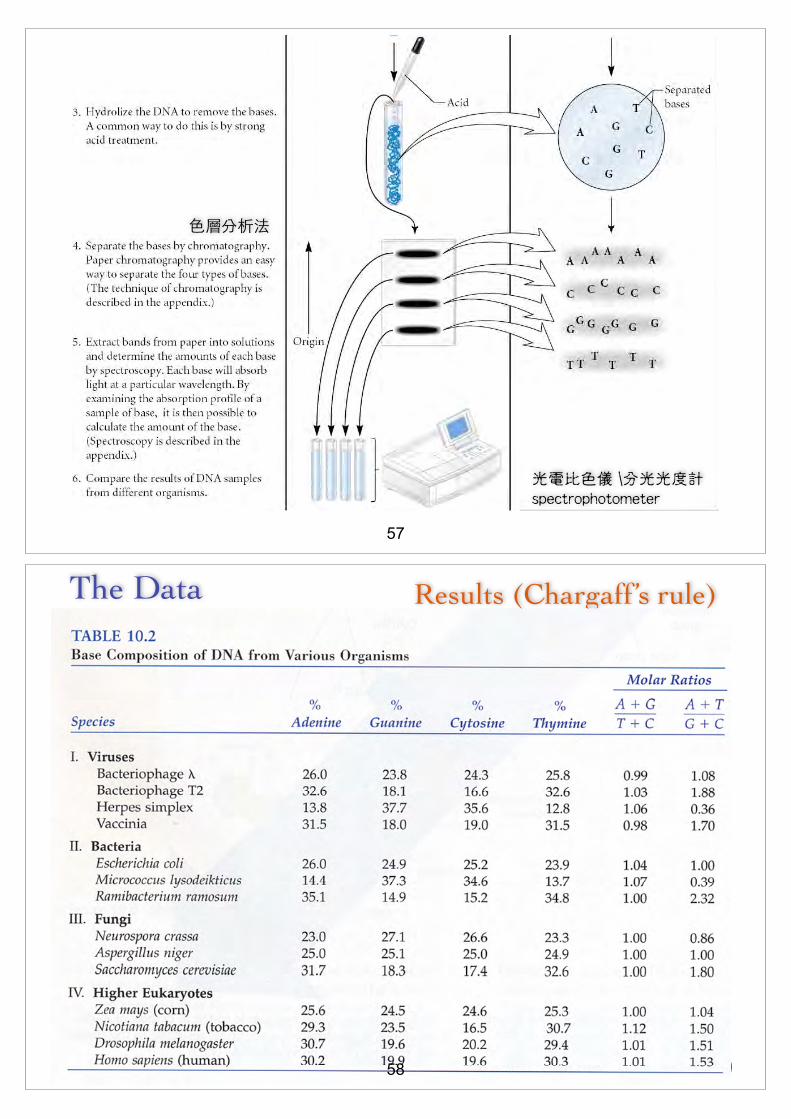
57
�����
光電比色儀 \分光光度計 spectrophotometer
The Data
9-39
Results (Chargaff’s rule)
58

Interpreting the Data
n The data shown in Table 10.2 are only a small sampling of Chargaff’s results
!
n The compelling observation was thatn Percent of adenine = percent of thyminen Percent of cytosine = percent of guanine !
n This observation became known as Chargaff’s rule (d[)
n It was crucial evidence that Watson and Crick used to elucidate the structure of DNA
59
qqqqqq
A few key events led to discovery of the double-helix structure1. α-helix protein secondary structure (Linus Pauling)
2. X-ray diffraction (Rosalind Franklin)
1). Helical structure
2). Helical structure of DNA was too wide to be only single-
stranded helix
3). 10 base pairs per complete turn
3. The amount of A equals T and the amount of G equals C
(Chargaff)60
qqqqqq

n Familiar with all of these key observations, Watson and
Crick set out to solve the structure of DNA
n They tried to build ball-and-stick models that incorporated all
known experimental observations
!
n A critical question was how the two strands (or more
strands) would interact
Watson and Crick
61
Fig. 9.16b
62

n Familiar with all of these key observations, Watson and Crick set out to solve the structure of DNA n They tried to build ball-and-stick models that incorporated all
known experimental observations
!
n A critical question was how the two (or more strands) would interactn An early hypothesis proposed that the strands interact through
phosphate-Mg++ crosslinksn Refer to Figure 9.15
Watson and Crick
63
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.
Fig09.14
Sugar
–O
OO
OP
Base
Sugar
Base
Sugar
O
OOP
Base
Sugar
BaseO–
Mg2+
n This hypothesis was, of course, incorrect!
Why doesn’t this model ?

n They went back to the ball-and-stick units
n They then built models with the n Sugar-phosphate backbone on the outside
n Bases projecting toward each other
!
n They first considered a structure in which bases form H bonds with identical bases in the opposite strandn ie., A to A, T to T, C to C, and G to G
n Model building revealed that this also was incorrect
Watson and Crick
65
n They then realized that the hydrogen bonding of A and T resembled that between C and G n So they built ball-and-stick models with AT and CG
interactions n These were consistent with all known data about DNA structure
!
n Watson, Crick and Maurice Wilkins were awarded the Nobel Prize in 1962n Rosalind Franklin died in 1958, and Nobel prizes are not
awarded posthumously
Watson and Crick
66

1962 Nobel Prize winners
67
Fig. 16-1
Awarded the Nobel Prize in 1962 68

Francis Crick, Maurice Wilkins, James Watson and Rosalind Franklin
69
Watson and Crick
70

Plos Wilson: ants vs DNA
71
Figure72
Antiparallel

Double helix DNA ... commonly B form
73
Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Figure 16.7 The double helix
O
–O O
OHP
H2C
O
–O O
OP
H2C
O
–O O
OP
H2C
O
–OO
OP
O
OH
O
O
OT A
C
GC
A T
O
O
O
O
OH
CH2
O O–
OOP
CH2
O O–
OOP
CH2
O O–
OO
P
CH2
OO–
OOP
5ʹ end
Hydrogen bond3ʹ end
5ʹ end
3ʹ end
C
T
A
A
T
CG
GC
A T
C G
AT
AT
A T
TA
C
TA0.34 nm
3.4 nm
(a) Key features of DNA structure (b) Partial chemical structure (c) Space-filling model
G
1 nm
G
H2C
G
74
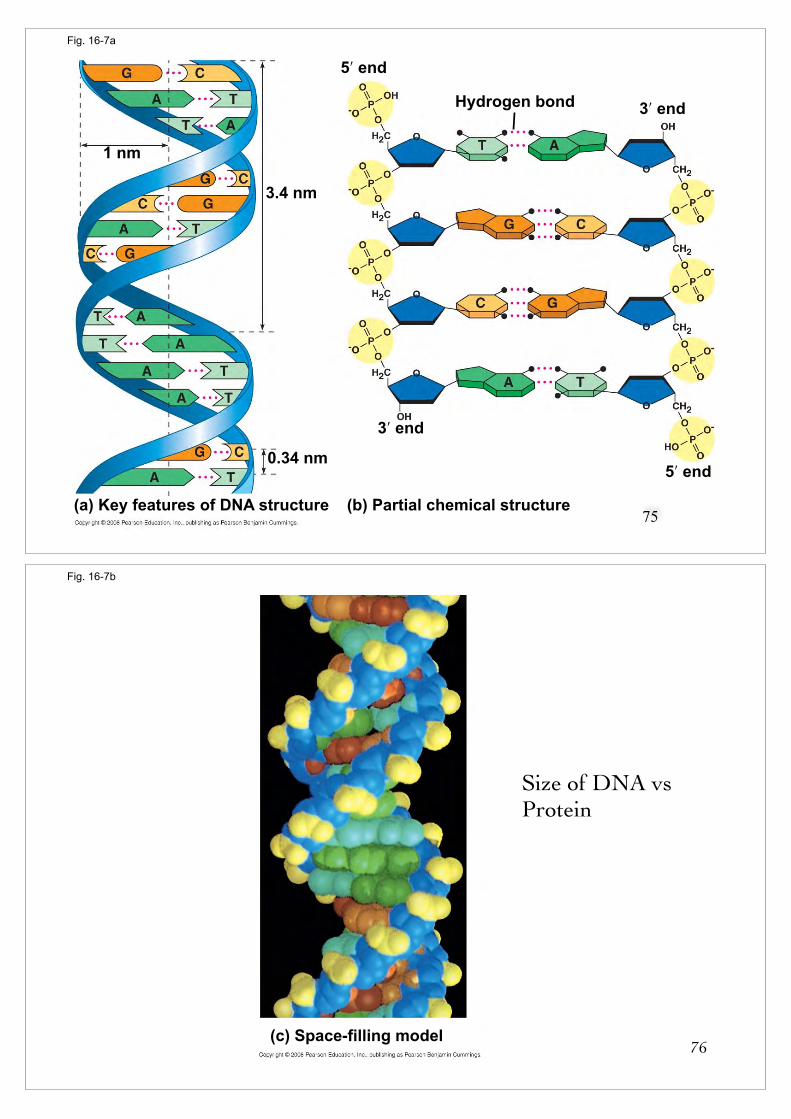
Fig. 16-7a
Hydrogen bond 3ʹ end
5ʹ end
3.4 nm
0.34 nm
3ʹ end
5ʹ end
(b) Partial chemical structure(a) Key features of DNA structure
1 nm
75
Fig. 16-7b
(c) Space-filling model76
Size of DNA vs Protein

n General structural features (Figures)
The DNA Double Helix
n Two strands are twisted together around a common axis
n There are 10 bases per complete twist
n The two strands are antiparallel (P@'�P?@')n One runs in the 5’ to 3’ direction and the other 3’ to 5’
n The helix is right-handedn As it spirals away from you, the helix turns in a clockwise
direction77
qqqqqq
n The helix is right-handed

n General structural features
The DNA Double Helix
n The double-bonded structure is stabilized by!
n 1. Hydrogen bonding between complementary basesn A bonded to T by two hydrogen bonds
n C bonded to G by three hydrogen bonds
!
n 2. Base stackingn Within the DNA, the bases are oriented so that the flattened
regions are facing each other79
n General structural features (Figures )
The DNA Double Helix
n There are two asymmetrical grooves (溝槽) on the outside of the helixn 1. Major groove
!
n 2. Minor groove
!
nCertain proteins can bind within these groovesnThey (proteins) can thus interact with a particular
sequence of bases80

The DNA Double Helix
81
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.
Space-filling model of DNA
Ball-and-stick model of DNA
Minor groove Major
grooveMinor groove
Major groove
Fig. 9.18 (TE Art)
82

Protein and DNA interaction
83
n The primary structure of an RNA strand is much like that of a DNA strand
!
n RNA strands are typically several hundred to several thousand nucleotides in length
!
n In RNA synthesis, only one of the two strands of DNA is used as a template
RNA Structure
84

Figure
RNA molecules
85
n Although usually single-stranded, RNA molecules can form short double-stranded regions and tertiary structuren This secondary structure is due to complementary base-pairing
n A to U and C to G
n This allows short regions to form a double helix
n RNA double helices typicallyn Are right-handedn With 11 to 12 base pairs per turn!
n Different types of RNA secondary structures are possiblen Refer to Figure
86

Figure
Also called hair-pin
Complementary regions
Noncomplementary regions
Held together by hydrogen bonds
Have bases projecting away from double stranded regions
Possible secondary structures of RNA molecules
RNA double helices are antiparallel and typically of the right-handed A form with 11 to 12 base pairs per turn
87
One role of RNA secondary structure
88

DNA repl icat ion
天 然 的 尚 好 !
89





























